异长春花苷内酰胺在制备抗炎药物中的应用的制作方法
本发明属于医药技术领域,具体涉及异长春花苷内酰胺在制备抗炎药物中的应用。
背景技术:
胆木(Nauclea officinalis Pierre ex Pitard),又名乌檀,是茜草科植物胆木的干燥根茎,主要分布于我国海南、广东、广西等省。其性味苦、寒,具有清热解毒、消肿止痛之功效,民间常用于感冒发热、肺炎、肠炎、痢疾、湿疹、皮疹、脓疡等病的治疗。文献报道胆木中主要含有生物碱类成分,而异长春花苷内酰胺(Strictosamide)是含量最高的生物碱成分(Fenxia Zhu*,Jiaquan Chen,Hui Wang,et al Journal of Chromatography B,2015,1007:54-66;胡欣.乌檀化学成分分离分析与相关成分活性、药动学研究.沈阳药科大学,2007;陈家全,徐光富,王慧,等.UPLC法同时测定胆木中6种成分的含量.中国药科大学学报,2014,45(4):434-437)。
发明人前期利用柱色谱技术结合自动纯化系统制备了异长春花苷内酰胺单体化合物(朱粉霞,王静静,宋捷,等.胆木的化学成分研究.药学学报,2013,48(2):276-280)。异长春花苷内酰胺在胆木中含量丰富,易于分离,其的分子式为C26H30N2O8,分子量为498.52,CAS登录号为23141-25-5。
目前的专利报道中,异长春花苷内酰胺主要有以下应用:(1)异长春花苷内酰胺在制备抗皮肤色素沉着药物中的应用(CN103585167A),(2)异长春花苷内酰胺制备治疗咳嗽和哮喘药物中的用途(CN102600197A),(3)含有异长春花苷内酰胺等成分的组合制剂在制备清热解毒、消肿止痛药物方面的用途(CN102600269A)等;但目前尚无异长春花苷内酰胺在抗炎方面应用的报道。
抗炎药物一般是通过调节对炎症具有重要作用的促炎症因子(TNF-α、IL-1β、IL-6等)和炎症介质的水平,从而发挥其抗炎作用。关于抗炎机制方面的研究,文献对很多信号通路在炎症方面的作用进行了描述(Guha M,Mackman N.LPS induction of gene expression in human monocytes.Cell Signal.2001,13:85-94),其中,核因子-κB(NF-κB)与丝裂原激活的蛋白激酶 (MAPK)通路是与炎症相关的两条重要通路。研究表明,NF-κB主要是通过IκB激酶(IKK)介导的IκB-α的磷酸化与降解而激活的;在MAPK信号通路中,ERK、JNK和p38被认为是抗炎药物作用的重要靶点。
技术实现要素:
本发明的目的在于提供异长春花苷内酰胺在制备抗炎药物中的医药用途。
本发明的第一方面,发明人研究了异长春花苷内酰胺对LPS刺激的RAW 264.7细胞中炎症因子IL-1β和TNF-α的水平以及IL-1β和TNF-αmRNA表达的影响,从而研究了异长春花苷内酰胺的抗炎活性。
本发明的第二方面,发明人研究了异长春花苷内酰胺在LPS刺激的RAW 264.7细胞中发挥抗炎作用的分子机制。异长春花苷内酰胺经LPS刺激的RAW 264.7细胞体外实验,发现该化合物可以抑制NF-κB信号通路中IKKα、IκB-α和p65的磷酸化;同时,也能够抑制MAPK信号通路中ERK、JNK和p38的磷酸化。
实验结果表明,本发明中的化合物异长春花苷内酰胺能够显著地抑制IL-1β和TNF-α水平及其mRNA的表达,对炎症有明显的抑制作用,因此可以用于制备抗炎药物。此外,抗炎机制研究表明,异长春花苷内酰胺能够抑制LPS刺激的RAW 264.7细胞中的NF-κB和MAPK信号通路。
附图说明
图1是异长春花苷内酰胺的化学结构式;
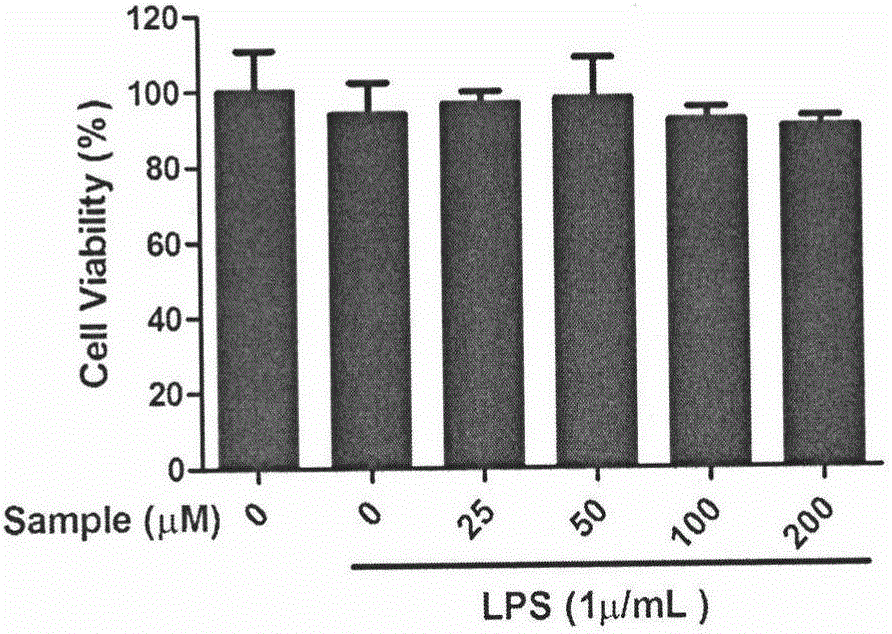
图2是异长春花苷内酰胺对LPS刺激的RAW 264.7细胞活力的影响;
图3是异长春花苷内酰胺对LPS刺激的RAW 264.7细胞中炎症因子水平的影响,其中A是异长春花苷内酰胺对IL-1β水平的影响,B是异长春花苷内酰胺对TNF-α水平的影响;
图4是异长春花苷内酰胺对IL-1β和TNF-αmRNA表达的影响,其中A是异长春花苷内酰胺对IL-1βmRNA水平的影响,B是异长春花苷内酰胺对TNF-αmRNA水平的影响;
图5是异长春花苷内酰胺对LPS刺激的RAW 264.7细胞中NF-κB信号通路的抑制作用。其中A是异长春花苷内酰胺对IκB磷酸化的的影响,B是异长春花苷内酰胺对p65磷酸化的影响,C是异长春花苷内酰胺对IKKα磷酸化的影响;
图6是异长春花苷内酰胺对LPS刺激的RAW 264.7细胞中MAPK信号通路的抑制作用。其中A是异长春花苷内酰对ERK磷酸化的影响,B是异长春花苷内酰胺对JNK磷酸化的影响,C是异长春花苷内酰胺对p38磷酸化的影响。
具体实施方式
现结合实施例和附图,对本发明作详细描述,但本发明的实施不仅限于此。本发明所用试剂和原料均市售可得或可按文献方法制备。
下列实施例中未注明具体条件的实验方法,通常按照常规条件,或按照制造厂商所建议的条件。
实施例:异长春花苷内酰胺对LPS刺激的RAW 264.7细胞中炎症具有良好的抑制作用。
(一)细胞培养
小鼠RAW 264.7巨噬细胞,购自中科院上海细胞库。在37℃,5%CO2条件下,用含10%HyClone胎牛血清、100U/mL青霉素及100mg/mL链霉素的DMEM不完全培养基培养,0.5%胰酶消化传代。所有操作均为无菌操作。
(二)细胞活力实验
RAW 264.7细胞接种于96孔板中(4×106个/mL),每组六个复孔,每孔含200μL不完全高糖DMEM培养基。细胞贴壁后,用PBS洗涤细胞一遍,对照组加入不完全高糖培养基,诱导组加入含1μg/mL LPS的不完全高糖培养基,给药组加入25、50、100和200μM的异长春花苷内酰胺溶液和含1μg/mL LPS的不完全高糖培养基,于37℃,5%CO2孵育箱中培养24h后,加20μL 5mg/mL MTT培养4h,弃去培养液,用PBS洗涤细胞一遍,加150μL DMSO,微波震荡15min,用酶标仪检测570nm和490nm处的OD值。细胞活力=[(OD570-OD490)sanple/(OD570-OD490)control]×100%。
结果如图1所示,25、50、100和200μM浓度的异长春花苷内酰胺对细胞活性的影响与对照组和诱导组相比无明显差异(P>0.05);表明浓度为25~200μM异长春花苷内酰胺对LPS刺激的RAW 264.7细胞没有显著的细胞毒性。
(三)异长春花苷内酰胺对LPS刺激的RAW 264.7细胞中IL-1β产生的影响
本实验通过ELISA试剂盒检测异长春花苷内酰胺对LPS刺激的RAW 264.7细胞产生IL-1β的影响。将RAW 264.7细胞接种于96孔板中(4×106个/mL),每组六个复孔,每孔含200μL不完全高糖DMEM培养基。细胞贴壁后,用PBS洗涤细胞一遍,对照组加入不完全高糖培养基,诱导组加入含1μg/mL LPS的不完全高糖培养基,给药组加入25、50、100和200μM的异长春花苷内酰胺溶液和含1μg/mLLPS的不完全高糖培养基,于37℃,5%CO2孵育箱中培养24h后,吸50μL上清液,根据ELISA试剂盒(杭州联科生物技术有限公司)的操作说明书进行操作,检测IL-1β含量。
结果如图2A所示,LPS诱导组的IL-1β水平显著高于空白对照组(P<0.001);25、50、100和200μM异长春花苷内酰胺组的IL-1β水平与诱导组相比,有显著差异(P<0.01),25μM水平的异长春花苷内酰胺能够将IL-1β的产量降至15.00±2.86pg/mL(P<0.01)。上述结果表明异长春花苷内酰胺显著地且呈剂量依赖性的抑制LPS刺激的RAW 264.7小鼠巨噬细胞产生IL-1β。
(四)异长春花苷内酰胺对LPS刺激的RAW 264.7细胞中TNF-α产生的影响
本实验通过ELISA试剂盒检测异长春花苷内酰胺对LPS刺激的RAW 264.7细胞产生TNF-α的影响。RAW 264.7细胞接种于96孔板中(4×106个/mL),每组六个复孔,每孔含200μL不完全高糖DMEM培养基。细胞贴壁后,用PBS洗涤细胞一遍,对照组加入不完全高糖培养基,诱导组加入含1μg/mL LPS的不完全高糖培养基,给药组加入25、50、100和200μM的异长春花苷内酰胺溶液和含1μg/mL LPS的不完全高糖培养基,于37℃,5%CO2孵育箱中培养24h后,吸50μL上清液,根据ELISA试剂盒(杭州联科生物技术有限公司)的操作说明书进行操作,检测TNF-α含量。
实验结果如图2B所示,LPS诱导组的TNF-α水平显著高于空白对照组(P<0.001);25、50、100和200μM异长春花苷内酰胺给药组的TNF-α水平与诱导组相比,有显著差异(P<0.001),且该化合物对LPS刺激的RAW264.7细胞中TNF-α生成的抑制作用呈剂量依 赖性,25μM水平的异长春花苷内酰胺能够将TNF-α的水平降至2370.34±22.28pg/mL(P<0.001)。上述结果表明异长春花苷内酰胺能显著抑制LPS刺激的RAW264.7细胞产生TNF-α。
(五)异长春花苷内酰胺对IL-1βmRNA表达的影响
本实验使用Trizol试剂提取对照组、诱导组和异长春花苷内酰胺给药组的RAW 264.7细胞产生的IL-1βmRNA,利用RT-PCR确定mRNA的相对含量。RAW 264.7细胞接种于96孔板中(4×106个/mL),每组六个复孔,每孔含200μL不完全高糖DMEM培养基。细胞贴壁后,用PBS洗涤细胞一遍,对照组加入不完全高糖培养基,诱导组加入含1μg/mL LPS的不完全高糖培养基,给药组加入25、50、100和200μM的异长春花苷内酰胺溶液和含1μg/mL LPS的不完全高糖培养基,于37℃,5%CO2孵育箱中培养24h后,根据Trizol试剂供应商(美国Invitrogen公司)提供的方法提取RNA,并测定在260nm和280nm的吸收值,确定RNA的浓度和纯度;随后,利用RT-PCR扩增RNA,以GAPDH为内参,采用2-ΔΔCt法计算IL-1βmRNA的相对表达量。
实验结果如图3A所示,结果表明,25~200μM水平的异长春花苷内酰胺能够显著抑制IL-1βmRNA的表达(P<0.001),并且抑制作用呈剂量依赖性;当异长春花苷内酰胺的浓度达到200μM时,与LPS诱导组相比,IL-1βmRNA的表达被抑制了80.59%。
(六)异长春花苷内酰胺对TNF-αmRNA表达的影响
本实验使用Trizol试剂提取对照组、诱导组和异长春花苷内酰胺给药组的RAW 264.7细胞产生的TNF-αmRNA,利用RT-PCR确定mRNA的相对含量。RAW 264.7细胞接种于96孔板中(4×106个/mL),每组六个复孔,每孔含200μL不完全高糖DMEM培养基。细胞贴壁后,用PBS洗涤细胞一遍,对照组加入不完全高糖培养基,诱导组加入含1μg/mL LPS的不完全高糖培养基,给药组加入25、50、100和200μM的异长春花苷内酰胺溶液和含1μg/mL LPS的不完全高糖培养基,于37℃,5%CO2孵育箱中培养24h后,根据Trizol试剂供应商(美国Invitrogen公司)提供的方法提取RNA,并测定在260nm和280nm的吸收值,确定RNA的浓度和纯度;随后,利用RT-PCR扩增RNA,以GAPDH为内参,采用2-ΔΔCt法计算TNF-αmRNA的相对表达量。
实验结果如图3B所示,结果表明,25~200μM水平的异长春花苷内酰胺能够显著抑制TNF-αmRNA的表达(P<0.001),并且抑制作用呈剂量依赖性;当异长春花苷内酰胺 的浓度达到200μM时,与LPS诱导组相比,TNF-αmRNA的表达也被抑制了80.59%。
(七)异长春花苷内酰胺对NF-κB通路的抑制作用
RAW 264.7细胞接种于96孔板中(4×106个/mL),每组六个复孔,每孔含200μL不完全高糖DMEM培养基。细胞贴壁后,用PBS洗涤细胞一遍,对照组加入不完全高糖培养基,诱导组加入含1μg/mL LPS的不完全高糖培养基,给药组加入25、50、100和200μM的异长春花苷内酰胺溶液和含1μg/mL LPS的不完全高糖培养基,于37℃,5%CO2孵育箱中培养24h后,胰酶消化,于冷PBS中洗涤2遍,各加入200ul冰预冷裂解缓冲液,混匀后冰浴30min,每隔5min涡旋震荡10s,充分裂解,4℃离心10min,上清液分装,存于-70℃保存,待检测。利用Western blot法,以GAPDH为内参,计算IκBα、p-IκBα、IKKα、p-IKKα、p65、p-p65的相对表达量。
实验结果如图4所示,结果表明,异长春花苷内酰胺能够显著地降低LPS-刺激的RAW264.7细胞中p-IκBα、p-IKKα和p-p65的表达,并且抑制作用呈剂量依赖性;当异长春花苷内酰胺的浓度达到200μM时,p-p65/p65比值下降至0.43±0.05,与空白对照组接近。
(八)异长春花苷内酰胺对MAPK通路的抑制作用
RAW 264.7细胞接种于96孔板中(4×106个/mL),每组六个复孔,每孔含200μL不完全高糖DMEM培养基。细胞贴壁后,用PBS洗涤细胞一遍,对照组加入不完全高糖培养基,诱导组加入含1μg/mL LPS的不完全高糖培养基,给药组加入25、50、100和200μM的异长春花苷内酰胺溶液和含1μg/mL LPS的不完全高糖培养基,于37℃,5%CO2孵育箱中培养24h后,胰酶消化,于冷PBS中洗涤2遍,各加入200uL冰预冷裂解缓冲液,混匀后冰浴30min,每隔5min涡旋震荡10s,充分裂解,4℃离心10min,上清液分装,存于-70℃保存,待检测。利用Western blot法,以GAPDH为内参,计算ERK、p-ERK、JNK、p-JNK、p38、p-p38的相对表达量。
实验结果如图5所示,结果表明,异长春花苷内酰胺能够显著地降低LPS-刺激的RAW 264.7细胞中p-ERK、p-JNK和p-p38的表达,并且抑制作用呈剂量依赖性;当异长春花苷内酰胺的浓度达到200μM水平时,对ERK、JNK和p-38磷酸化的抑制率分别为90.00%、71.69%和75.49%。
上述实验结果表明,异长春花苷内酰胺可显著抑制LPS刺激的RAW 264.7细胞中IL-1β和TNF-α的生成,同时也抑制了IL-1β和TNF-αmRNA的表达,这些结果说明异长春花苷内酰胺具有良好的抗炎活性,且无细胞毒性,因此可用于制备抗炎药物。抗炎机制方面的研究结果显示,异长春花苷内酰胺能够抑制RAW 264.7细胞中LPS刺激的NF-κB和MAPK通路的激活。
以上已对本发明创造的较佳实例进行了具体说明,但本发明创造不限于所述实施例,熟悉本领域的技术人员在不违背本发明创造精神的前提下还可做出其他的等同的变型或替换,这些等同的变型或替换均包含在本申请权力要求所限定的范围。
- 还没有人留言评论。精彩留言会获得点赞!