干细胞组合物和生产用于治疗应用的干细胞的方法与流程
背景技术:
干细胞是用于治疗应用、医学研究、药物测试等的有吸引力的细胞来源。不幸的是,细胞作为同种异体移植物的使用是有问题的。干细胞的扩增至大量是细胞治疗的先决条件,但是成体msc具有有限的增殖能力。这是现有技术中的公知常识,即用于来自骨髓(bm)分离的msc的主要方法是当将全骨髓置于塑料培养皿中时其粘附在塑料上的能力,并且在4小时后,将非粘附细胞洗出。该规程还可以包括在具有低粘度和低渗透压的高密度溶液(例如ficoll、percoll)中密度离心bm,以获得含有msc的bm的单核部分。另外,来自固体组织(比如骨)的msc可以通过将小块骨放置在瓶内并通过骨外植体(explant)长出的细胞而分离。可选地,也可以使用胶原酶消化过程。然而,细胞与其它类型的细胞如造血干细胞的污染导致可能发生异质群体,损害(compromising)msc样本的纯度。为了克服这个问题,已经开发了两种类似的技术,磁珠分选和荧光激活细胞分选(facs)。另外,荧光激活细胞分选(facs)是一种分离msc的方法。离体扩增是msc分离后的下一个必要步骤,其允许msc的自由离体自动更新(renovation),目标是生产治疗上显著的细胞数量,这取决于几个因素,比如供体依赖性(年龄、性别、创伤的存在和全身性疾病的存在)或方法依赖性。
即使msc可以在相对短的时间段内由于快速增殖而极大地扩增,然而在现有技术中没有允许在没有广泛的体外培养的情况下,生产治疗上相关数量的msc的方法,这意味着使用酶处理从瓶至瓶的多轮细胞传代。在这种条件下的msc扩增已经显示逐渐降低其msc最大分化潜能。广泛的继代培养(subcultivation)损害细胞的功能,导致与生长停滞和细胞凋亡相关的细胞衰老。另一方面,有数据表明延长的培养可导致自发的转化获得致瘤潜能。此外,显示msc的特定特性在培养期间丧失。第5代和第10代以上的细胞与该代次以下的细胞相比,msc的心脏保护作用降低。这可以通过降低的血管内皮生长因子释放潜能来解释(crisostomo等人,2006;digirolamo等人,1999;muraglia等人,2000;rubio等人,2005;stenderup等人,2003)。因此,现有技术中存在的所有方法应该在扩增细胞的产量和质量之间做出折衷,这限制了它们在细胞治疗中的使用。
本发明中描述的方法和细胞组合物克服了上述限制,比如多次细胞传代、细胞衰老、分化降低和生长因子生产降低、冷冻保存细胞数量有限。相比之下,我们提供了降低细胞传代、降低体外细胞操作和相应增加分化能力和生长因子生产、稳定的核型、实际上无限数量的冷冻保存的细胞。
引用的所有出版物、专利和专利出版物以所有目的通过引用整体并入本文。
技术实现要素:
本发明涉及在体外获得和/或扩增从外植体组织分离的干细胞群的方法。该方法优选包括在培养基中培养组织至少一个收获周期的步骤。收获周期通常包括以下步骤:在外植体细胞的培养基中保持组织,持续足以获得半汇合干细胞培养物的时间段,并收集这些细胞用于最终的干细胞群。优选地,在每个收获周期之前,组织被最低程度的操作。
在具体实施方案中,本发明涉及从外植体组织获得原代衍生干细胞的扩增群的体外方法,所述方法包括:
a.洗涤和最低程度操作外植体组织;
b.在具有培养基第一培养表面中在体外培养所述外植体组织以建立原代衍生干细胞的培养物;
c.从培养基收集原代衍生干细胞;和
d.将外植体组织转移到具有培养基的第二培养表面;
e.用外植体组织重复步骤b、c和d至少一次,从而获得从外植体组织原代衍生的扩增的群。
优选地,这些方法包括洗涤和最低程度操作外植体组织。优选地,在每个收获周期之前,外植体组织不被酶促或化学处理或解剖。优选地,从外植体组织获得原代衍生的干细胞的扩增群的方法还包括在生长培养基上传代原代衍生的干细胞并从生长培养基收集干细胞的步骤。传代可以在每个收获周期多次或结束后进行。
所述方法可以进一步包括传代外植的细胞并在生长培养基中培养传代的细胞。在一些方面,在培养基中至少一个收获周期培养组织包括最初在含有少于5%血清的培养基中培养组织,然后在含有15%血清的培养基中培养组织。在含有少于5%血清的培养基中培养组织的持续时间为3天。在含有15%血清的培养基中培养组织的持续时间为7天。在一些实施方案中,培养基和生长培养基是dmem,例如,培养基是dmem-f12和/或生长培养基是dmem-低葡萄糖(dmem-lowglucose)。在一些方面,生长培养基不补充非必需氨基酸。在一些实施方案中,外植的细胞和/或传代细胞具有稳定的核型。
本发明还涉及干细胞的治疗组合物,其包含从至少第一收获周期获得的外植的细胞。在一些实施方案中,组合物包含从前3个收获周期、前65个收获周期或前100个收获周期获得的外植的细胞。在一些方面,组合物包含从至少第1代(p1)、从p1至p3或从p1至p50获得的传代的细胞。在一些实施方案中,组合物可以在冷冻条件下储存,而在组合物解冻后不改变其干细胞标记物的表达。
在本发明的方法和组合物的一些实施方案中,干细胞是神经嵴(neuronalcrest)或外周神经系统起源。在一些方面,干细胞是成体或产后干细胞、非胚胎干细胞。在一些实施方案中,组织是牙髓或产后组织,例如脐带或胎盘。
附图说明
图1描述了从脐带(uc)分离干细胞的示意图。a组描述了5cm的uc块的分离。b组描述了uc组织的洗涤;c组描述了切成小块的uc组织。d组表示已经机械地转移到基础培养基中并在塑料培养板中培养的小块uc组织。e组显示培养表面上cd105阳性的干细胞的粘附。f组描述了间充质干细胞的培养。
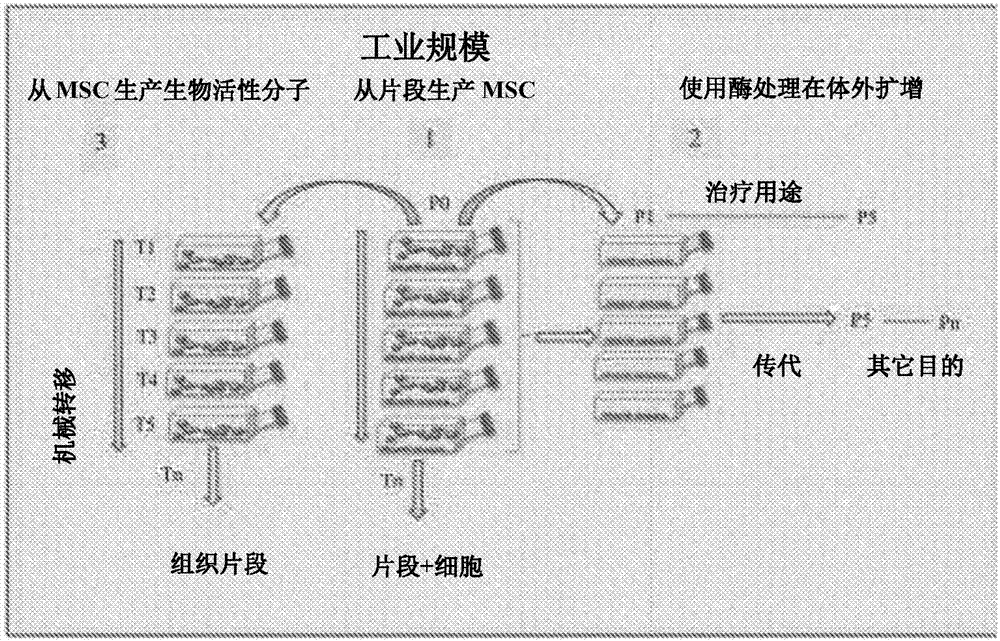
图2描述了以工业规模培养干细胞的方法。
图3描述了在生物技术、医学、药物、美容、营养等中使用的干细胞分离和机械转移的非酶方法。
图4描述了不同供体的片段的分离和这些片段在相同瓶中的共培养。从不同的hla匹配、部分匹配和/或不匹配的供体获得的片段可以在相同的瓶中培养以生产msc,只要它们不表现任何免疫反应的信号。
图5描述了“一次”干细胞分离的组织来源冷冻保存的常规方法(a)和本申请的组织来源冷冻保存(b)。
图6描述了正常msc与ep或ips细胞之间的主要差异。
图7描述了hidpsc在两种类型的生长培养基中的生长。
图8描述了idpsc腹膜内接种对感染mtb和牛分枝杆菌菌株(mbovis)的c57bl/6小鼠的肺细胞产生细胞因子的作用。
图9描述了通过荧光显微镜可视化的用于角膜移植的idpsc中的角膜缘(limbal)干细胞标记物的免疫染色。比例尺表示50μm的距离。a组描述了idpsc中p63的核定位。b组是a组与dapi核染色的合并图像。c组描述了整联蛋白β1的膜定位。d组描述了波形蛋白、中间丝的检测。e组描述了连接蛋白43的检测,其是细胞膜蛋白。f组描述了abcg2(一种浆膜蛋白)的检测。g组描述了k3/12的弱阳性免疫染色。h组描述了阴性对照。
图10描述了在idpsc中角膜缘干细胞标记物(abcg2、连接蛋白43和k12)的表达的rt-pcr分析。泳道1是100bp指示(ladder)。泳道2是人角膜组织样本。泳道3是人角膜缘组织样本。第4行是idpsc样本。
图11描述了牙髓中干细胞的龛。a组,齿由冠部和根部组成。b组显示了牙髓的血管周围的龛(a)和神经丛(b)的位置。c组描述了在血管周围的龛(a)和神经丛(b)中巢蛋白的免疫染色。d组显示了成牙质细胞下(subodontoblastic)的龛(c)和无细胞和富含细胞的区域(d)的位置。e组和f组分别描述了在成牙质细胞下的龛(c)和无细胞和富含细胞区域(d)中巢蛋白的免疫染色。
图12描述了脐带中神经干细胞的推定龛。a组显示了羊膜上皮中神经丝(nf)的表达。b组显示了静脉中低亲和力神经生长因子(p75或cd271)的表达。c组显示了从推定的神经干细胞龛分离的干细胞的β-iii微管蛋白的阳性免疫染色。d组显示从推定的神经干细胞龛分离的干细胞的gfap的阳性免疫染色。
图13描述了脐带组织中不同的干细胞龛中转录因子比如oct3/4、nanog和sox2的表达。白色箭头描述了表达这些转录因子的细胞,而黑色箭头描述了缺乏这些转录因子表达的细胞。
具体实施方式
美国专利申请公开号2004/0136967公开了从脐带基质获得干细胞的方法,包括从脐带的外植体分离细胞或者酶促游离(freeing)细胞,例如胶原酶或胰蛋白酶。然后用另一载玻片覆盖组织切片并在完全培养基中培养。因此,现有技术没有教导外植体培养物的可重复使用。
与现有技术的扩大干细胞群的方法不同,本发明使用多次收获周期来增加培养的干细胞群。还不同于现有技术的方法,起源组织不被解剖(dissected),而是机械地切片或磨碎用以重复的收获周期。组织应该被最低程度操作。收获周期是指将组织转移到新鲜培养表面以建立例如干细胞的新的外植体培养物的周期。在本发明中,收获的周期(用于干细胞来源的组织转移)可以通过多次周期重复,例如1至100次转移。多次收获周期建立多次原代培养以提供具有独特特征的干细胞的大规模供应。换言之,本发明提供了稳健的大规模培养干细胞的方法,其通过重复原代培养的从最低程度操作的组织来源收获的干细胞,所述最低程度操作的组织来源通过相同组织的重复收获周期。出乎意料地,重复收获周期导致在每个周期中获得的粘附细胞的数量增加。并且令人惊奇的是,我们发现通过重复的外植体培养,通过非酶促收获暴露于含有抗生素/抗真菌剂和补充剂的培养生长培养基的最低程度操作的组织块,我们可以多次启动(initiate)第0代(p0)条件,由于在整个重复收获周期提供足够的干细胞数量期间低代次的核型稳定性,提高了安全性的机会。
在现有技术中,对于非治疗应用,主要是诊断目的,部分已知最低程度操作组织的方法。特别地,这些目的包括培养器官(例如角膜)和组织外植体,用于有毒的、致癌的、治疗物质、生物制剂的作用评估或敏感性测试。最近,我们已经公开了这种技术用于分离牙髓干细胞的应用。这种方法的主要目的是保持培养的组织结构,例如,实质和间质在其结构和功能方面是保守的。因此,在培养期间的在体外的器官类似于在体内的器官功能。此外,我们还利用培养外植体的方法用于从器官的块分离细胞,这以前在现有技术中被用于诊断、筛选方法:分离的组织被分离,在无菌条件下最低程度处理,切成小块并放置在含有培养基的细胞培养皿中。
外植体培养是这样一种培养,即其中细胞留在其周围细胞外基质中,其更准确地模拟在体内干细胞/祖细胞从龛(niches)迁移到外部环境,并且祖细胞迁移出组织并粘附于培养皿的表面。两种方法具有相似之处,因为它们保留了环境和干细胞龛。利用该方法用于干细胞扩增是有益的,因为在培养时间下的组织经受缺氧。然而,培养基提供了足够的营养供应以支持最低程度操作的组织的外植体细胞和组织内的细胞保持在有活力的状态。与现有技术的教导相比,这种益处是意想不到的,并且在保持干细胞的未分化状态方面提供了巨大的优势。因此,作为小器官和外植体的牙髓的机械转移是连续地分离干细胞以便在分离后保持其原始表征不变的策略。牙髓是松散的结缔组织,并且该器官可以在重量上变化。dp更轻,趋于流动而没有粘附和细胞长出(outgrowth)。为了加速髓组织对基底的附着,可以使用无菌盖玻片,甚至提供更多的缺氧条件。使用盖玻片是在多次收获周期期间在组织干细胞龛内诱导缺氧的另一个实例。
在一个实施方案中,本发明涉及从最低程度操作的组织通过至少一个收获周期在体外扩增至原代衍生和传代的干细胞扩增群的方法。在每个收获周期期间,外植体细胞的数量是用经典的酶促衍生的干细胞扩增分离的细胞数目的至少两倍。收获周期的数量可以是至少三个周期(h1-h3),至少65个周期(h1-h65)或高达100个收获周期(h1-h100)。每个收获周期包括在具有抗生素的补充的培养基中,将最低程度操作的组织片段(例如非酶促地或化学处理的,未解剖的)保持至外植体细胞,持续足以获得半汇合的干细胞培养物的时间段,并收集这些细胞用于最终干细胞群。这一代培养的细胞是收获周期的原代(p0)。
所述方法包括洗涤最低程度操作的组织,并在体外保持组织至至少一次收获,使得干细胞,比如各种类型的间充质多能和多能干细胞附着于培养表面或珠。来自收获周期的每个外植体培养物为p0,并且作为随后传代或生物反应器系统的来源。外植体培养可以传代多达50代。细胞可以酶促地或非酶促地传代。在重复的收获周期中,组织和细胞可以保持在含有抗微生物溶液的生长因子和/或dmem补充的生长培养基中。在一些方面,通过使用本发明的方法从一块组织获得的干细胞的总数是通过经典的酶促衍生的干细胞扩增可以获得的干细胞的数量的至少三倍。在一些实施方案中,通过本发明的方法获得的干细胞的总数量比通过经典的酶促衍生的干细胞扩增获得的干细胞的数量至少3到至少65倍高。因此,本发明涉及与使用相同组织来源的经典方法相比,增加从单一或混合供体的组织/液体来源获得的干细胞数量的方法。
组织可以在建立外植体培养物之前冷冻保存。来自外植体培养和随后代次的粘附细胞也可以被冷冻保存。冷冻后的细胞活力大于50-90%,这使得它们适合于工业和治疗应用。在该方法的一些方面,根据描述的方法,治疗组合物获得自多次收获周期期间获得的混合的冷冻-解冻的间充质干细胞的批次。该批次可以包括来自混合的收获周期或相同收获周期的混合代次的细胞。用于研究和临床应用的一批细胞可以是包含来自不同收获周期的p0细胞的组合物。
在另一个实施方案中,本发明涉及一种细胞治疗的方法,包括将根据本发明的方法分离的批次注射和/或植入人或动物的需要治疗的位置。该批细胞可以来自不同收获周期和代次直接冷冻库存(stock)的混合细胞。该批次细胞也可以在施用至人或动物之前从冷冻库存的细胞培养。在一些实施方案中,测试该批次细胞的靶向治疗所需的功能参数,比如释放的相关因子和/或测试的相关的生物标记物。根据本发明的方法分离的最终干细胞群表达间充质标记物。最终的干细胞群还可以表达多能标记物、神经外胚层标记物和/或周细胞标记物。在一些实施方案中,优选地,最终干细胞群表达间充质和一些多能标记物,尤其是神经外胚层、上皮标记物和/或周细胞标记物。在一些实施方案中,优选地,干细胞缺乏mhcii类表达,比如hlaii(hla-dr)表达。对于神经元应用,优选的cns因子、测试的标记物,比如β微管蛋白、sox2或sox10;或神经元生长因子和肽,比如gdnf,ngf和bdnf。该批细胞可以具有测试的其分化能力。在一些方面,本发明还涉及用于细胞治疗的缓冲的治疗组合物,其中ph不是酸性和细胞毒性。
本发明的一个实施方案还涉及从最低程度操作组织在体外在扩增培养中获得原代干细胞的方法,其中所述组织不被化学品或酶处理或解剖完全分解。这些方法包括:从在细胞培养室中的所述组织连续地、周期性地或间歇地收获粘附的干细胞,所述组织保持在含有抗微生物溶液的补充的生长培养基中,并从第一次收获传代培养的细胞,重复步骤。在优选的实施方案中,重复收获步骤,并且对于每个收获步骤,将传代步骤重复至少一次、两次或三次。多次收获周期后培养物中产生的干细胞可以混合,并通过相关生物标记物的质量控制测试功能性。这些细胞可以在治疗使用之前冷冻。
表1比较了从本发明的方法和现有技术的方法获得的细胞的数量。
表1.从不同来源获得的成体间充质干细胞的比较:
在一个实施方案中,本发明的方法包括用抗生素和缓冲溶液从组织洗涤过量的血液,并对收集的组织进行消毒。组织还被机械地分解(dissociated)成组织片段,其中组织被切割成无显微镜眼睛可见的块。例如,组织片段应当大于1mm。将组织片段暴露于补充的培养基中。在优选的实施方案中,补充的培养基基于dmem,更优选dmemf12。在几次培养后,当粘附的细胞达到半汇合时,将组织片段转移到新平板用于另一个收获周期。
在一些实施方案中,每个收获周期后的干细胞(p0)培养多代次,以扩增培养物中干细胞的数量。干细胞可以传3代、15代或50代,同时保持正常核型。在一些实施方案中,在每个重复周期中,干细胞获得自前述重复周期的来自几个供体或来自相同供体的几次取样(例如来自相同供体的乳牙牙髓,通过外科手术的不同组织取样)混合的组织块。来源组织可以新鲜获得,或者在细胞库、冷冻保存后解冻获得。
在外植体收获之后,未分化或分化的细胞的酶促分裂/传代可以在现有技术中已知的经典细胞培养条件下,或在生物反应器条件下,在无菌塑料瓶、玻璃器皿、无菌袋、微载体(micro-carries)、微纤维反应器等中进行。
在一个实施方案中,可以通过机械转移、营养不足和缺氧条件加速干细胞扩增的方法。在一些实施方案中,该方法包括以下步骤:
(i)获得产后、年轻或成体间充质细胞和前体干细胞,其来自最低程度操作的组织(优选分离自含有神经外胚层起源的干细胞龛的细胞和周细胞的组织,或/和上皮来源的组织,即牙组织、产后组织,比如胎盘、羊膜、脐带;毛囊和外胚层(under-extodermal)来源的脂肪组织等。
(ii)其中所述组织基本上被测试活力,用抗生素、抗真菌条件保持,基本上清除血液,机械分离成小块(优选约0.5-5mm以便暴露于补充培养基)。
(iii)来自最低程度操作的组织的所述干细胞集落形成是在非酶促地/机械方法下进行的。
(iv)所述集落形成基本上通过机械操作和培养基暴露以及细胞粘附于塑料器皿、珠或粘附表面诱导,其中所述集落起始发生在缺氧/饥饿压力条件、低血清培养基5%或更低下约第3天或更晚,或在正常培养生长条件下更晚起始。
(v)重复多次收获提供具有高细胞数量的低传代数的用途,其中每个收获发生新的原代培养,其中在最初收获后组织尺寸减小,从随后收获的细胞收获压力和持续时间比最初原代收获培养快约两倍。
(vi)从这种最低程度操作的组织获得的所述细胞与起源于模拟体内条件的多干细胞龛在体内和体外表达相同的标记物。
(vii)所述集落形成的术语(term)可以在压力-缺氧条件下被基本上诱导和加速至少两倍,所述压力-缺氧条件可以由化学压力例如缺氧、机械压力例如在盖玻片压力;或/和补充饥饿下诱导,优选血清5%和更少(参见下面的实施例ms2)。
(viii)来自多次重复收获的干细胞的原代收获的集落形成单位后被酶促或非酶促传代后,暴露于适于细胞增殖的条件,其中在优选实施例中,血清补充为约15%,或进行生长因子和额外的补充。如果使用本领域公知的低葡萄糖dmem以加速缺氧/压力条件,所述增殖可以显著加速(参见下面的实施例ms3)。
用于细胞治疗的移植可在缓冲的药物组合物、水凝胶、植入支架内进行,具有或不具有额外的活性添加剂或协同的药物或生物成分等。
所获得的细胞可以通过单次但优选重复的施用方式通过全身或/和局部施用途径施用细胞来用于各种细胞治疗应用,如下面实施例中所示的,在狗中多发性硬化治疗等治疗中,其中通过2-3次,优选>2次,3次或4次重复注射细胞获得最佳结果,以改善症状。在更优选的方法中,应当在疾病的相对早期阶段施用细胞,其中过程的可逆性可以通过细胞治疗调节。
以上列出的优势将被转化为多种客户利益:
·预期允许稳健和可重复的工业规模生产明确定义和表征的干细胞
·来自早期代次的高质量的起始材料用于未来生物反应器扩增
·cellavita细胞的独特的蛋白表达模式允许简单和标准化的表征
图3显示了干细胞分离和片段的机械转移的非酶促方法的可能应用。细胞或分离的片段可以被冻干、沉淀、提取和纯化或进一步加工以分离生物活性分子。这些分子可以用作新的未知分子的测序和合成的模板,以产生用于美容(aesthetic)(恢复青春(rejuvenation))外部和内部使用的“混合物(cocktails)”或治疗霜剂。另一方面,msc可用于治疗各种疾病、损伤和创伤,如干细胞治疗。这些细胞还可以用作在癌症治疗中递送各种治疗或自杀分子的载体。这些细胞还可用于遗传和组织工程,具有或不具有支持物(或支架)的器官打印,以及与多种疾病中的其它类型的细胞和/或干细胞组合。
然而,我们发现分离的干细胞的最终群可以同时表现相似但也明确的特性,如表2所示,根据干细胞龛,其可以影响干细胞特性。
表2.uc和dp组织中的不同的干细胞龛
虽然表2显示了两种组织的实例,但是本公开不限于这些组织,并且可以包括任何类型的组织:
a.衍生自胚胎外胚层和神经嵴(舌下和下颌下组织、唾液和泪腺、牙乳头、齿叶(dentallamina)、hertwig上皮根鞘、牙囊、牙周韧带、皮肤、胎记等);
b.衍生自胚外中胚层(卵黄囊和羊膜、脐带、胎盘和胎膜);和
c.衍生自胚胎中胚层(脂肪组织和骨髓)。
所有这些组织可以用作片段的来源,其可以根据图1和2加工,并根据图3和4使用。
在a,b,c中描述的,但不限于这些实例,从不同组织来源获得的每种干细胞类型可用于各种目的和多种适应症的细胞治疗和再生医学。同时,一旦这些干细胞具有不同的胚胎起源,它们可以组合使用以便在治疗性治疗中提供增强的或协同的或补充的作用。示例性组织来源包括:脐带、牙/牙周组织、下颌组织、粘膜组织、胃肠组织、胎盘、羊水或羊膜囊、神经元组织(例如含有神经或神经胶质细胞的组织)、从神经嵴或周围神经系统起源、肌肉、骨、皮肤(真皮、表皮、含角质细胞或含黑素细胞的组织、包皮、整形手术或活检衍生的皮肤)、脂肪的/脂肪组织、血液和其它体液、乳腺组织、内皮组织、睾丸组织、卵巢、心脏组织、肝脏、骨组织、肺组织,唾液腺组织、胰腺组织、心脏和脾衍生的组织。
干细胞分离和机械转移的非酶促方法也可以扩展到具有最低程度变异的其它组织,比如脐带(图1)。例如,从脐带(uc)分离5cm的块(图1a),用无菌溶液如生理盐水溶液洗涤以便除去过量的血液过量(图1b)。将最初的uc组织块切成甚至更小的组织片段用于培养(c组),并机械地转移到基础培养基中(图1d)。基础培养基可以对于每种类型的干细胞是特异性的,其具体制剂取决于组织起源。cd105阳性的干细胞从组织迁移以粘附到培养表面上(图1e)。在图1f中,可观察到起源自小uc组织片段的间充质干细胞(msc)的培养物。
该方法可以容易地转化为工业规模培养,如图2所示。从uc分离多个片段并置于各种培养瓶中用于msc生产。在体外扩增后,使用酶处理这些细胞,其可以用于细胞治疗和用于其它目的。uc片段的几种机械转移可以提供大量的msc,其可以用于生产生物活性分子。
本发明的另一个实施方案涉及包括根据该方法分离的干细胞对患者的治疗方案。该方案开始于全身施用干细胞以保护性地调节患者的免疫和炎症状态。当通过测量患者中的细胞因子和免疫球蛋白和其它免疫调节因子(例如用血液测试)证实抗炎免疫调节作用时,可以将干细胞的局部植入用于再生细胞疗法。
功能测定可以包括协同因子的表征,比如例如疾病的抗炎、抗微生物和再生,所述疾病可以具有感染的风险,比如医院感染、伤口愈合,比如糖尿病性溃疡、手术伤口等。
在优选的实施方案中,使用细胞疗法治疗的cellavita技术应当与患者接受的其它可接受的医学和支持治疗辅助进行,但是辅助药物的体外相容性应当被测试为不抑制细胞免疫调节和再生潜能。例如,抗生素、类固醇、抗增殖和免疫抑制剂可以是有毒的,或抑制细胞活性,因此该方案应当调整到相关的治疗方案。
本公开、其方面和实施方案不限于本文公开的具体材料类型、成分、方法或其它实施例。现有技术中已知的许多额外的材料类型、成分、方法和规程被预期用于来自本公开的特定实施方案。因此,例如,尽管公开了特定的实施方案,但是这样的实施方案和实施方案的成分可以包括任何与预期的操作一致的成分、模型、类型、材料、单元、版本、数量和/或如现有技术中已知的用于这样的系统和实施方案成分。
词语“示例性”、“实施例”或其各种形式,在本文中用于表示用作实施例、实例或说明。在本文中被描述为“示例性”或“实施例”的任何方面或设计不一定被解释为相对于其它方面或设计是优选的或有利的。此外,提供实施例仅用于清楚和理解的目的,并且不意味着以任何方式限制或约束所公开的主题或本公开的相关部分。应当理解,可以表现不同范围的大量附加或替代实施例,但为了简洁的目的已经省略。
虽然本公开包括许多不同形式的实施方案,但是在附图中示出并且在本文中将详细描述特定的实施方案,应理解本公开被认为是所公开的方法和系统的原理的实施例,并不旨在将所公开的概念的广泛方面限于所示的实施例。
实施例
实施例1:牙髓的活力
maheral-atariabou-asi等人(2011)声称根据kerkis等人,2006的出版物从牙髓(dp)提取的多能细胞的起源不能是牙髓,因为5至7岁的患者不具有牙髓,而是所谓的牙胚,所述牙胚是未分化细胞的聚集体,其将形成具有不同部分的未来的牙齿:牙釉质、牙髓、牙龈(gum)、牙骨质、骨、血管和神经。然而,cordeiro等人(2008)发现人类脱落的乳牙的干细胞在体内产生dp样组织。作者将在人牙切片内制备的生物可降解支架中种植的shed移植到免疫缺陷小鼠中。他们观察到由这些细胞形成组织,证明了与生理的dp很类似的结构和细胞性。透射电子显微镜和免疫组织化学对牙本质涎蛋白的超微结构分析表明shed在体内分化为成牙质细胞样细胞。值得注意的是,shed也分化为内皮样细胞,如通过用稳定地转导有lacz的shed工程化的组织中,含血的血管壁内衬细胞的β-半乳糖苷酶染色所证明的。这项工作表明脱落的乳牙构成用于dp组织工程的干细胞的可行来源。
基于提取时髓的颜色,我们在细胞培养之前筛选有活力的髓。脱落的牙齿应具有红色的髓,作为细胞活力的指示。红色指示直到取出的时候髓接受血流。如果髓是灰色的,则可能流入髓的血流已被损害。因此,干细胞可能坏死并且不再具有恢复活力(arora等人,2009)。
在接受由伦理委员会(在欧洲的helsinki,在美国的ibr)批准的知情同意之后,在含有50%青霉素/链霉素溶液(100单位/ml青霉素,100单位/ml链霉素)和50%磷酸盐缓冲生理盐水(pbs)的无菌溶液中重复洗涤来自健康受试者的新鲜提取的乳牙,进行髓提取和培养实例,再次洗涤新鲜提取的髓,并在补充有3-20%胎牛血清(fbs)、100单位/ml青霉素、100单位/ml链霉素的dp组织保持培养基中进行髓的活力测试。一旦从最低程度操作的牙髓开始集落形成并且细胞附着于塑料细胞培养表面,在重复随后的收获周期之前进行安全性测试(无菌性测试和支原体测试),和或通过传代来扩增细胞和或冷冻保存组织或细胞。
如下进行髓提取和培养:
·在含有50%青霉素/链霉素溶液(100单位/ml青霉素,100单位/ml链霉素)和50%磷酸盐缓冲生理盐水(pbs)的无菌溶液中重复洗涤来自健康受试者的新鲜提取的乳牙。
·采用无菌针头从牙齿取出dp。在含有3%青霉素/链霉素的溶液中洗涤新鲜提取的髓。在补充有15%胎牛血清(fbs,hyclone)、100单位/ml青霉素、100单位/ml链霉素、2mml-谷氨酰胺和2mm非必需氨基酸的dp组织保持培养基中进行髓的活力测试。
·一旦牙髓附着到塑料细胞培养表面,将从5天到两周观察到集落形成。
·进行的安全测试为无菌测试和支原体测试。
实施例2:来自牙髓外植体的干细胞
在最近的牙起源衍生的干细胞综述中(deepaponnaiyan2014)详细描述的牙组织的不同实例描述了来自牙组织的干细胞的生物标志物的不同表征,比如牙髓干细胞、牙周韧带干细胞、来自人脱落的乳牙的干细胞,来自根尖乳头的牙囊祖细胞干细胞,口腔骨膜干细胞和最近的来自牙龈结缔组织的。引人注目地,大多数标记物在不同的牙干细胞之间是相似的,但是notch和cd29标记物在来自各种牙源的干细胞之间的表达不同。通过我们公开的方法获得的细胞的体外和体内标记物表达是相同的。
实施例3:来自在早期代次重复收获的畸胎瘤形成人idpsc
畸胎瘤形成是确定细胞比如胚胎干细胞(es细胞)或诱导多能干细胞(ips细胞)的多能性的重要工具。我们的研究使用与gropp等人(2012)最近公开的规程类似的用于评估细胞畸胎瘤形成能力的一致规程。当106个细胞(gropp等人使用105个细胞)的小鼠es细胞和人ips细胞用人工基底膜(matrigel)皮下共移植到免疫缺陷小鼠中时,我们的方法显示高度可重复和有效。在100%的情况下,我们观察到大量动物中的畸胎瘤形成,甚至在移植后直到6个月的随访检查中。我们使用这种方法进行其它成体间充质干细胞(msc)的生物安全性分析,比如衍生自乳牙的牙髓、脐带、脂肪组织等的那些。除了这里描述的实验系统,使用也部分表达多能标记物的来自骨髓、肝、卵黄囊、尿囊和羊水的狗胎儿msc验证了该方法。
畸胎瘤测定方法:
监测肿瘤的发展4个月(~16周)。对于多能细胞,畸胎瘤的组织学标准是发生衍生自所有三个胚层的组织。该分析由病理学家进行。对于成体/间充质干细胞,考虑在细胞注射部位的正常组织完整性的任何类型或任何变化。
结果
实验系统:
a.小鼠胚胎干细胞
b.小鼠3t3成纤维细胞
c.永久的小鼠细胞系balbc3t3细胞系,克隆a31
d.人类ips-idpsc
e.人类es细胞
f.人类idpsc
我们在多种研究中使用前述方法来表征由我们建立的不同小鼠es细胞系多能性,以及确认在第25代以上的es细胞多能性,以及用于表征从小鼠es细胞系获得的亚克隆(sukoyan等人,2002;carta等人,2006;kerkis等人,2007;lavagnolli等人,2007;hayshi等人,2010)。此外,在我们小组最近的出版物中(
实验系统:
a.在早期(n=10)和晚期代次(n=10)的idpsc三种不同的原代培养物
b.人原代成纤维细胞
我们先前报道了idpsc群的分离,其在至少前25代期间,其表达胚胎干细胞标记物oct-4、nanog、ssea-3、ssea-4、tra-1-60和tra-1-81以及几种间充质干细胞标记物,同时保持正常核型和干细胞扩增的表征速率。此外,在化学上确定的培养条件下,这些细胞可以在体外诱导,经历一致的分化成平滑和骨骼肌、神经元、软骨和骨。尽管idpsc具有小尺寸,并且在细胞质中细胞器缺乏(paucity),但它们与表现典型间充质/成纤维细胞样形态的天然多能细胞不同(lizier等人,2012)。idpsc是间充质的,与es和ips细胞不同,其是上皮的(图5)。msc细胞和es细胞(或ips细胞)之间的主要区别是msc正在迁移、可塑(plastic)和锚定(anchoring)。msc细胞合成细胞外基质并且是无细胞连接的细胞。
肿瘤形成与上述es和ips细胞有关,但与正常msc无关。idpsc由具有可变数量的(1-25%的细胞)表达多能标记物的干细胞的msc群体组成(kerkis等人,2006;lizier等人,2012)。该规程适用于idpsc的群体,尤其是使用的细胞数方面,其基于20%的idpsc表达的多能标记物计算。评估5×106的idpsc的致畸性,而不是测试106细胞。虽然在所有研究的器官中发现idpscdna的存在,但没有观察到肿瘤形成或任何形态学改变(lizier等人)。
实施例4:主要干细胞标记物的表达模式不受牙髓(dp)的冷冻保存影响
由于来自脐带、羊膜囊、胎盘的新生细胞和来自牙髓的年轻干细胞的不成熟性,当与它们的成体对照物(骨髓和脂肪组织)相比时,它们显示出更高的增殖能力和更高的扩增率。因此,来自脐带、羊膜囊、胎盘和牙髓的细胞可以冷冻保存以备将来用作同种异体治疗(组织片段和/或细胞最初冷冻保存在冰箱(约-80℃)随后在液氮中(约-196℃))。
我们发现,在冷冻保存和解冻外植体之前和之后,收获的细胞保持相同模式的因子表达。我们已经显示衍生自最低程度操作的外植体的细胞,其来自冷冻保存前和后的牙髓和脐带的重复收获长期培养,表达间充质标记物(即cd73、cd105、cd90)不表达造血标记物(即cd34、cd45),并且不表达至少一种组织相容性标记物——hla-dr,并且保持核型稳定性,保持多能性(与诱导因子比如视黄酸、bmp-tnf、ngf等培养,分化成不同的谱系包括神经元)。
例如,从乳牙中取出的牙髓冷冻保存,然后解冻以分离idpsc。将idpsc培养至收获0/3代(h0p3),并通过facs分析与来自新鲜dp的相同代次的hidpsc进行比较。发现两种细胞的表达模式相似(表3)。两种idpsc对于间充质标记物(cd105、cd73、cd90)的表达是阳性的,并且对于造血标记物cd45和组织相容性标记物hla-dr的表达是阴性的。
表3.牙髓冷冻保存对hidpsc表型表征的影响
实施例5:生长培养基对hidpsc生长速率的影响
为了评价用于培养hidpsc的最佳生长条件,在两种类型的生长培养基中接种等量细胞并培养5天(表4)。发现在第2类型的生长培养基中的细胞生长速率更高(两倍)(图7),因此由dmem-低葡萄糖和补充有15%fbs组成的生长培养基对于细胞培养是更优的。
表4.所使用的各种培养基的组成
实施例5:从牙髓(dp)起始细胞的萌发(sprouting)
在组织培养物中保持dp的两个相等片段。片段之一在15%fbs存在下培养,另一个在血清饥饿压力条件(4.8%fbs)下培养。在4.8%fbs存在下,从dp起始细胞萌发为3天,在15%fbs存在下为7天。因此,血清饥饿可以减少髓转移所需的时间。因此,在血清饥饿培养基存在下,从牙髓起始细胞萌发更快。
实施例7:晚期群(lp)hidpsc的旁分泌作用包括抗炎、免疫调节和抗微生物特性
msc通过旁分泌机制辅助并通过抗炎和免疫调节机制调节再生环境。
idpsc腹膜内接种对静脉注射感染毒力不同的结核分枝杆菌(mycobacteriatuberculosis,mtb)和牛分枝杆菌(mycobacteriaumbovis)菌株的c57bl/6小鼠的肺细胞的细胞因子产生的作用。如bioplex测试数据所示,idpsc接种降低受感染的肺细胞产生的细胞因子(图8)。在感染高毒力mtb菌株的肺中观察到促炎症(tnf-α,il-1b,mip-2,il-17)和抗炎(il-10)细胞因子的强烈降低。ifn-g和kc产生的降低较不明显。仅在接种idpsc的小鼠中观察到il-4产生的诱导。
肿瘤坏死因子(tnf,cachexin或恶液质素,以前称为肿瘤坏死因子α或tnfα)是参与全身性炎症的脂肪因子。tnf的主要作用是调节免疫细胞。tnf是内源性热原,能够诱导发热、凋亡性细胞死亡、恶病质、炎症并抑制肿瘤发生和病毒复制,并通过il1和il6生产细胞对败血症应答。tnf生产的调节异常涉及多种人类疾病,包括阿尔茨海默氏病。
干扰素γ(ifnγ或ifn-g)是二聚化可溶性细胞因子,其是干扰素类ii型的唯一成员,被称为免疫干扰素,一种对于针对病毒和细胞内细菌感染和肿瘤控制的先天性和适应性免疫关键的细胞因子。ifnγ是巨噬细胞的重要活化剂。异常ifnγ表达与许多炎性和自身免疫性疾病相关。ifnγ在免疫系统中的重要性部分来自ifnγ直接抑制病毒复制的能力,并且最重要的是来自ifnγ的免疫刺激和免疫调节作用。
白细胞介素17是通过增加各种组织中的趋化因子生产以将单核细胞和中性粒细胞募集到炎症位点、在迟发型反应中充当有效调节子的细胞因子,类似于ifnγ。白细胞介素17是响应于细胞外病原体侵入免疫系统并诱导病原体细胞基质破坏的促炎细胞因子。
白细胞介素-1β(il-1β)也称为分解代谢物,是一种炎症响应的重要调节子的细胞因子蛋白。它参与多种细胞活动,包括细胞增殖、分化和凋亡。il-17最显著的作用是其参与诱导和调节促炎症响应。il-17通常与过敏反应相关。
白细胞介素-10(il-10),也称为人细胞因子合成抑制因子(csif),是一种抗炎细胞因子。il-10是在免疫调节和炎症中具有多效作用的细胞因子。
趋化因子配体2(cxcl2)也称为巨噬细胞炎症蛋白2(mip-2),在调节中性粒细胞炎症响应中起主要作用,并且是败血症发展中的调节子。kc和巨噬细胞炎症蛋白-2(mip-2)是具有c-x-c基序的趋化因子,其在外科损伤后在皮肤中表现出不同的时间模式的表达。kc和mip-2是中性粒细胞的化学引诱物。已知两种趋化因子在广谱的急性和慢性炎性情况中表达,并且被认为是确保炎症反应的性质和量级的关键决定因素。
实施例8:通过最低程度操作的组织的多次收获(lp)获得的hidpsc的稳健的免疫调节能力
当衍生自单核细胞衍生的树突细胞衍生的共培养时,idpsc呈现免疫调节作用。共培养导致lint增殖的能力降低(50%和更多),并诱导lint分化,使lintcd4+/foxp3+/il10+和lintcd4+/foxp3+/ifn-γ+细胞的比例显著增加。在idpsc中没有观察到hla-dr表达的作用。
我们已经证明,尽管从最低程度操作的组织重复收获获得的干细胞(cellavita方法)表达大多数多能标记物,但是这些干细胞注射到小鼠中不形成畸胎瘤。
表5.各种哺乳动物物种之间的hidpsc处理的比较,使用新鲜培养衍生的细胞模型和形成冷冻保存的髓和细胞的细胞模型
实施例9:牙髓中新神经丛和成牙质细胞下干细胞龛的发现
早期神经元标记物巢蛋白的免疫染色显示神经丛、血管周围的龛和成牙质细胞下的龛之间的表达差异。神经丛中的细胞的巢蛋白呈阳性(图11c,b),而血管周围的龛中的细胞的巢蛋白呈阴性(图11c,a)。尽管无细胞和富含细胞区域的龛的巢蛋白是阴性的(图11d,d和11f,d),但是成牙质细胞下的龛中的细胞的巢蛋白也是阳性的(图11d,c和11e,c)
实施例10:脐带中干细胞的推定的龛的发现
通过检测神经丝(nf)和低亲和力神经生长因子受体(p75或cd271)检测脐带中的推定干细胞龛。nf是在神经元中发现的10nm的丝或中间丝。cd271是神经营养因子的两种受体类型之一,其是刺激神经元细胞存活和分化的蛋白质生长因子家族。在脐带组织中发现nf和cd271。nf的表达主要在羊膜上皮区域(图12a),而cd271的表达主要在静脉和动脉中(图12b)。从这些龛分离的干细胞能够进行神经元分化,在体外表达β-iii微管蛋白和胶质纤维酸性蛋白(gfap)(图12c和12d)。
实施例11:脐带中多能干细胞或未成熟前体龛的发现
检测到转录因子,比如oct3/4、nanog和sox2的表达。使用核定位在wj、静脉(ve)、动脉(ar)的细胞中以及在定位于羊膜上皮(aep)的基底层(bl)的细胞中,适当地检测到oct3/4蛋白表达(图13)。检测到少量核中表达所有这三种标记物的细胞,类似于多能干细胞。因此,我们可以预测分离了来自ve或ar和来自脐带的aep的少量“真正的”多能干细胞。这些细胞也可以在其它相关组织中发现,比如胎盘和衍生自胚外中胚层的组织。
实施例12:安全性测定-在几次机械转移后干细胞龛的体外培养
众所周知,干细胞位于人体的特定解剖部位,称为干细胞龛(scn)。每个组织具有其自己的scn,其对从周围微环境接受的信号响应,做出关于干细胞的命运的决定。然而,关于干细胞在其天然龛内的原位生物学表征,以及这些表征如何可以在msc的体外培养过程中改变了解甚少。例如,细胞可以停止表达一些蛋白质或开始表达新的蛋白质。这个问题是非常重要的,因为它可以在临床研究中具有深远的影响,并可以解释在再生医学中干细胞的成功和/或失败。体外培养的细胞缺乏组织结构以及基于组织环境(细胞-细胞相互作用,特定因子的分泌等)的所有功能。这就是为什么细胞系可以失去组织特异性功能,并可能获得与scn中的细胞完全不同的分子表型。对scn中干细胞标记物的表达模式的研究对于评价干细胞安全是必要的,尤其是其在人中的使用。
在表6中,我们总结了通过免疫组织化学测定体内分离后细胞的组织样本所获得的数据,以及使用几种机械转移培养的细胞的体外样本的流式细胞术所获得的数据。表6证明在体内和体外尤其是牙髓中,由干细胞表达的大多数标记物,在机械转移后获得的分离细胞中没有显示标记物的缺失。在脐带组织的机械转移后获得的细胞证实在表达c-kit和波形蛋白的细胞频率中的微小差异。虽然这些结果来自从牙髓和脐带组织分离的干细胞,但是这种安全性测定可以用于来自其它来源的干细胞,甚至用于液相中的细胞。
表6.在体内和体外来自牙髓和脐带组织的干细胞中主要干细胞标记物的表达模式。+++表示高数量的细胞,++表示中等数量的细胞,+表示低数量的细胞。
除非另有定义,否则本文中的所有技术和科学术语具有与本发明所属领域的普通技术人员通常理解的相同的含义。尽管在本发明的实践或测试中可以使用与本文所述的那些类似或等同的任何方法和材料,但是本文描述了优选的方法和材料。引用的所有出版物,专利和专利出版物为了所有目的通过引用整体并入本文。
本文所讨论的出版物仅仅是为了在本申请的申请日之前的公开提供的。本文中的任何内容都不应被解释为承认本发明不具有借助于先前发明的先前发表的权利。
应当理解,本公开的发明不限于所描述的特定方法、规程和材料,因为这些可以变化。还应当理解,本文所使用的术语仅用于描述特定实施方案的目的,并且不旨在限定本发明的范围,本发明的范围将仅由所附权利要求限定。
本领域技术人员将认识到或能够使用不超过常规实验来确定与本文所述的本发明的具体实施例的许多等同物。这样的等同物旨在被所附权利要求所涵盖。
参考文献
1.chenvc等人.(2012)“scalablegmpcompliantsuspensionculturesystemforhumanescells”stemcellres.2012may;8(3):388-402.
2.schirmaier等人:scale-upofadiposetissue-derivedmesenchymalstemcellproductioninstirredsingle-usebioreactorsunderlow-serumconditions.eng.lifesci.2014,14,292–303,
3.vallee等人:adipose-tissueengineering:takingadvantageofthepropertiesofhumanadipose-derivedstem/stromalcells.patholbiol(paris).2009jun;57(4):309-17
4.nekanti等人:optimizationandscale-upofwharton'sjelly-derivedmesenchymalstemcellsforclinicalapplications.stemcellresearch(2010)5,244–254.
5.brookeg,rossettit,pelekanosr,等人:manufacturingofhumanplacenta-derivedmesenchymalstemcellsforclinicaltrials.brjhaematol.144:571–579.2008.
6.govindasamy等人:humanplateletlysatepermitsscale-upofdentalpulpstromalcellsforclinicalapplications.cytotherapy,2011;earlyonline,1–13
7.hanley等人:efficientmanufacturingoftherapeuticmesenchymalstromalcellswiththeuseofthequantumcellexpansionsystem;cytotherapy,2014-08-01,volume16,issue8,1048-1058
8.liziernf,kerkisa,gomescm,heblingj,oliveiracf,caplanai,kerkisi.scaling-upofdentalpulpstemcellsisolatedfrommultipleniches.plosone.2012;7(6):e39885;kerkisi,caplanai.stemcellsindentalpulpofdeciduousteeth.tissueengpartbrev.2012apr;18(2):129-38.
9.maheral-atariabou-asinúriacasalsfarrélluísginertarridalluísginertarridaeduardoferréspadrópatentapplicationtitle:pluripotentstemcellsobtainedfromdentalpulphttp://www.faqs.org/patents/app/20110236356
10.cordeiromm,dongz,kanekot,zhangz,miyazawam,shis,smithaj,
11.arorav,arorap,munshiak.bankingstemcellsfromhumanexfoliateddeciduousteeth(shed):savingforthefuturejclinpediatrdent33(4):289–294,2009
12.sukoyanma,kerkisay,mellomr,kerkisie,visintinja,pereiralv.establishmentofnewmurineembryonicstemcelllinesforthegenerationofmousemodelsofhumangeneticdiseases.brazjmedbiolres.2002may;35(5):535-42.
13.cartal,pereiral,arteaga-solise,lee-arteagasy,lenartb,starcherb,merkelca,sukoyanm,kerkisa,hazekin,keenedr,sakaily,ramirezf.fibrillins1and2performpartiallyoverlappingfunctionsduringaorticdevelopment.jbiolchem.2006mar24;281(12):8016-23
14.kerkisa,fonsecasa,serafimrc,lavagnollitm,abdelmassihs,abdelmassihr,kerkisi.invitrodifferentiationofmalemouseembryonicstemcellsintobothpresumptivespermcellsandoocytes.cloningstemcells.2007winter;9(4):535-48.
15.lavagnollitm,fonsecasa,serafimrc,pereiravs,santosej,abdelmassihs,kerkisa,kerkisi.presumptivegermcellsderivedfrommousepluripotentsomaticcellhybrids.differentiation.2009sep-oct;78(2-3):124-30
16.hayashima,guerreirojr,cassolaac,liziernf,kerkisa,camargoac,kerkisi.long-termcultureofmouseembryonicstemcell-derivedadherentneurospheresandfunctionalneurons.tissueengpartcmethods.2010dec;16(6):1493-502.
17.limabl,santosej,fernandesgr,merkelc,mellomr,gomesjp,soukoyanm,kerkisa,massironism,visintinja,pereiralvanewmousemodelformarfansyndromepresentsphenotypicvariabilityassociatedwiththegeneticbackgroundandoveralllevelsoffbn1expression.plosone.2010nov30;5(11):e14136.
18.
19.deepaponnaiyan--dodentalstemcellsdepictdistinctcharacteristics?—establishingtheir“phenotypicfingerprint”dentresj(isfahan).2014mar-apr;11(2):163–172
20.brito等人.,imbalanceofp75ntr/trkbproteinexpressioninhuntington’sdisease:implicationforneuroprotectivetherapies.celldeathanddisease(2013)4,e595;doi:10.138/cddis.2013.116
21.crisostomopr,wangm,wairiukogm,等人.highpassagenumberofstemcellsadverselyaffectsstemcellactivationandmyocardialprotection.shock.2006;26:575-80.
22.digirolamocm,stokesd,colterd,等人.propagationandsenescenceofhumanmarrowstromalcellsinculture:asimplecolony-formingassayidentifiessampleswiththegreatestpotentialtopropagateanddifferentiate.brjhaematol.1999;107:275-81.
23.muragliaa,canceddar,quartor.clonalmesenchymalprogenitorsfromhumanbonemarrowdifferentiateinvitroaccordingtoahierarchicalmodel.jcellsci.2000;113:1161-6.
24.rubiod,garcia-castroj,martinmc,等人.spontaneoushumanadultstemcelltransformation.cancerres.2005;65:3035-9.
25.stenderupk,justesenj,clausenc,kassemm.agingisassociatedwithdecreasedmaximallifespanandacceleratedsenescenceofbonemarrowstromalcells.bone.2003;33:919-26.
- 还没有人留言评论。精彩留言会获得点赞!