一种芒果乙烯受体MiERS1基因的制作方法
【
技术领域:
】本发明涉及生物
技术领域:
,具体涉及一种芒果乙烯受体miers1基因。
背景技术:
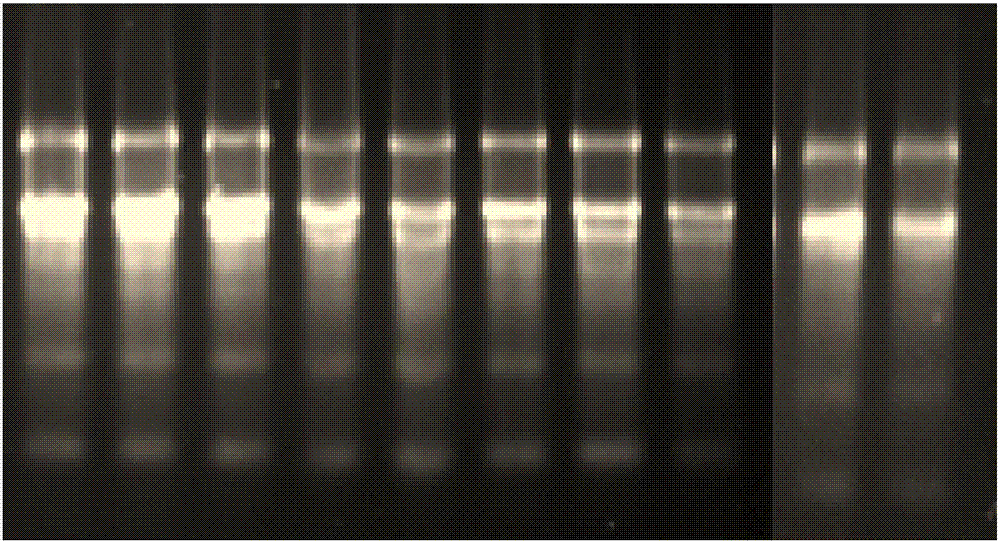
:芒果(mangniferaindical.)是著名的热带水果,风味独特,肉质嫩滑,营养价值高,有“热带果王”的美誉,在世界热带水果产量中仅次于香蕉位居第二,在世界生产与贸易中均占有重要地位。近年来,全球芒果年产量达到2700万吨。芒果属于典型的呼吸跃变型果实,采后果实成熟过程中有明显的呼吸跃变,果实达到完熟时呼吸强度最大,之后随着果实组织衰老逐渐降低。芒果采后对乙烯非常敏感,跃变开始之前,组织内乙烯浓度极低,在即将发生跃变之前,乙烯明显上升,引起呼吸跃变(张劲,等.《6个芒果品种品质特性评价研究》.食品科技,2011.)。芒果采收正值夏季,采后不耐贮,易腐烂,代谢旺盛,果实容易后熟而变黄、变软,低温贮藏中极易受到冷害,导致果皮快速褐变,温度较高时又易发生病害,从而降低芒果的食用价值,制约芒果生产发展。乙烯(ethylene)是一种重要的植物激素,几乎所有的高等植物都能合成乙烯,并在植物整个生长发育过程中起作用。种子萌发、器官衰老与脱落、果实成熟、环境胁迫和病原反应等过程都受乙烯的调控,特别是在果蔬成熟和衰老过程中乙烯起独特的调控作用,但乙烯需要通过信号转导途径与乙烯受体结合,才能对植物的诸多生理过程产生影响。乙烯受体是乙烯信号转导途径中的关键信号传递因子,是乙烯的应答顺利进行所必需(jiang和fu,2002;李丽秀和赖钟雄,2013)。通过控制乙烯受体的表达可以影响乙烯信号转导途径,使植物成熟和采后寿命等相关变化得以延缓或中断。乙烯受体基因家族这类乙烯信号转导途径中关键因子的克隆及表达特性的确定,对研究果实发育成熟有着重要的意义。乙烯受体成员在果实成熟过程中可能具有多种调控模式,不同类型果实的乙烯受体基因的表达是复杂多样的。乙烯受体在植物中由多基因家族所编码,根据其编码蛋白结构不同,可分为etr1和etr2家族两大类。etr1家族又可分为etr1亚类和ers1亚类,它们主要区别在于是否存在反应调节器结构域。乙烯受体基因处于乙烯信号传导其他基因座的最上游。ers1最早在酵母中通过与etr1交叉杂交分离得到,研究表明,由于ers1在c端缺少信号接收域,因此与下游负调控因子ctr1的结合能力比etr1略低。另外ers1的表达受乙烯诱导,而etr1则不受。有报道称ers1表达量的增加能够提高组织对乙烯的敏感性。这些现象虽然与乙烯负调控模型矛盾,但可能是由于乙烯与受体结合后需要一定的时间来释放乙烯,因此会促进组织生成更多的受体来结合乙烯,同时不断合成新的受体会激活ctr1,在乙烯水平下降时会适当地减弱乙烯反应。最近发现ers1具有双重的生物特性,不仅能像其它受体一样抑制乙烯信号,而且能够在依赖etr1的条件下促进乙烯反应,因为ers1需要与其它受体共同协作才能负调控乙烯信号,它与etr1结合形成异源复合体后会在一定程度上降低etr1的活性,如果过量表达ers1则会对etr1功能产生抑制,从而增强乙烯反应。通过调节乙烯受体基因ers1的表达水平调控果实成熟衰老进程,是实现利用生物技术延缓果实后熟软化的一个重要途径。乙烯受体由多基因家族编码,它们在结构和基因时空表达上各有特点。目前在拟南芥和水稻中均分离了5个乙烯受体基因,在模式植物的基础上,乙烯受体在果实中的研究也取得了一些进展。番茄果实存在至少6个乙烯受体成员,苹果可能具有5个,梨果实中含有4个,研究者又相继从香蕉、冬枣、柑橘、柿等果实中分离得到了多个乙烯受体同源基因。由此可见,每一种水果内可能同时存在多个乙烯受体基因,而随着研究的深入,可能每一种果实中将会有更多的乙烯受体基因不断被克隆。且乙烯受体基因在不同植物中可能存在着不同的表达模式。通过基因工程等技术手段调控乙烯受体基因的功能,从而降低果实对乙烯的敏感性,是延缓果实成熟衰老进程的有效途径之一,这已在番茄果实中得到了证实。目前,虽然对于芒果果实乙烯释放以及乙烯反应已经有很多报道,但是有关芒果果实成熟衰老进程中乙烯信号转导途径的研究,尤其是乙烯受体的表达与调控机制尚不清楚,还未克隆出芒果果实乙烯受体基因家族成员。技术实现要素:本发明的发明目的在于:针对上述问题,提供一种芒果乙烯受体miers1基因及其编码的蛋白质。为了实现上述目的,本发明采用的技术方案如下:一种芒果乙烯受体miers1基因,所述基因名称为miers1,所述miers1的核苷酸序列如序列表中seqidno:1所示。一种芒果乙烯受体蛋白,所述蛋白名称miers1蛋白,所述miers1蛋白是如下述1)或2)所述的蛋白质:1)其氨基酸序列由序列表seqidno:2所示;2)由序列表seqidno:1所述的碱基序列编码。一种表达载体,包含权利要求1所述基因miers1。本发明的芒果乙烯受体基因miers1来源是芒果(mangniferaindical.),为漆树科芒果属的栽培品种台农1号,产地广西百色。基因miers1的碱基序列如序列表中seqidno:1所示;共2171碱基序列,1个多聚腺苷酸尾巴和1个有1890个碱基序列的开放解码区。基因miers1编码的蛋白质含有629个氨基酸,其氨基酸序列由序列表seqidno:2所示。miers1蛋白分子总量为80518.8,等电点pi6.34,5’不翻译区和3’不翻译区序列分别位于总长度的102和109核苷酸。负电荷和正电荷的氨基酸残基总数分别为77和71,分子式为c3595h5734n992o1046s29,总原子数为11396,脂肪系数103.73,不稳定系数38.96,亲水性为0.025,说明它是一个稳定的疏水蛋白。基因miers1与ncbi数据库中已报道序列未找到重合的。经nnpredict软件预测分析miers1蛋白的二级结构,结构显示:含有α-螺旋、β-转角和无规则卷曲3种结构,其中miers1蛋白α-螺旋所占的比例分别为43.45%;β-转角占7.52%;无规则卷曲占29.94%。故miers1蛋白二级结构主要以α-螺旋为主。含有上述基因的重组载体、转基因细胞系或重组菌也属于本发明的保护范围之内。扩增上述基因的全长或其任一片段的引物对也属于本发明的保护范围之内。一种权利要求1所述芒果乙烯受体miers1基因的应用,在芒果调控成熟过程和/或延长芒果保质期中的应用。一种权利要求1所述芒果乙烯受体miers1基因的应用,在抑制芒果乙烯受体表达中的应用。一种延长芒果保质期的方法,运用乙烯抑制剂作用于权利要求1所述基因miers1。优化的,所述乙烯抑制剂选自1-甲基环丙烯、1-戊基环丙烯、1-已基环丙烯、1-丙基环丙烯和1-辛基环丙烯中的任一种或几种。通过运用乙烯抑制剂针对性的作用于本发明乙烯受体基因miers1或基因miers1编码的蛋白质,可以实现阻断乙烯合成,达到减少芒果内乙烯量的目的,从而达到延长芒果保质期的目的。同理,在芒果成熟前,也可以使用化化学物质或生物物质作用于本发明乙烯受体基因miers1或基因miers1编码的蛋白质,可以实现加速乙烯合成,促进芒果提前成熟,从而达到调节芒果成熟过程目的。综上所述,由于采用了上述技术方案,本发明的有益效果是:本发明从芒果中分离克隆得到了芒果乙烯受体基因miers1,通过乙烯抑制剂作用于乙烯受体基因miers1可以显著抑制乙烯受体基因miers1的表达,从而减少乙烯释放量和延缓乙烯释放高峰。本发明为芒果采后保持品质,延缓衰老提供了一种新的手段。为芒果采后保藏提供一种新的方法。本发明所述基因编码序列短,易于克隆和基因工程操作,所需材料易于获取。【附图说明】图1实施例1中1.1rna的提取方法rna电泳图;图2实施例1下1.6半定量rt-pcr分析rna电泳检测图;电泳图顺序依次为:空白组处理组0、2、4、6、8天,1-mcp处理组0、2、4、6、8天。图3实施例1下2.3荧光定量pcr检测中样品的样品的gadph扩增曲线;图4实施例1下2.3荧光定量pcr检测中样品的miers1扩增曲线;图5实施例1下2.3荧光定量样品的gadph溶解曲线;图6实施例1下2.3荧光定量样品的miers1溶解曲线;图7实施例2中miers1的三级结构预测图;图8实施例2中miers1的蛋白系统进化树构建图;图9实施例3中1-mcp处理对miers1基因表达的影响结果图;图10实施例3中1-mcp处理对芒果25℃贮藏条件下乙烯产生速率的影响;图11实施例3中1-mcp处理对芒果4℃贮藏条件下乙烯产生速率的影响。【具体实施方式】下面结合实施例对本发明作进一步说明。下述实施例中,如无特殊说明,均为常规方法。本发明的完成所依赖的遗传资源为:芒果(mangniferaindical.)是漆树科芒果属的栽培品种台农1号,产地广西。实施例1芒果乙烯受体基因miers1提取本发明中,总rna的提取和cdna合成总rna的提取参照bioteke(北京百泰克生物技术有限公司生产)通用植物总rna提取试剂盒(离心柱型)说明书操作,并保存于-80℃冰箱。cdna第一链合成参照revertaidtmfirst-strandsynthesissystem说明书操作,合成的cdna用于保守区克隆、3′race及orf克隆。5′racecdna第一链的合成参照transscriptⅱfirst-strandcdnasynthesissupermix试剂盒进行,cdna第一链的合成时加入0.2μl2pmol/0.2μl5′etr特异引物gsp:5′-gctgccatcttcaagcct-3′;cdna第一链合成产物纯化、加同聚物尾及cdna第二链的合成均参照5′racesystemforrapidamplificationofcdnaends,version2.0的试剂盒。1.1rna的提取方法1)匀浆处理:取100mg芒果组织在液氮中迅速研磨成粉末,加入500μlbufferrls,立即涡旋剧烈震荡混匀。2)然后于4℃12,000rpm(~13,400×g)离心2分钟。3)将上清液转入已装入收集管的过滤柱(spincolumnsfs)中,4℃12,000rpm离心1分钟,小心吸取收集管中的上清并转移至新的rnase-free离心管(自备)中,枪头尽量避免接触收集管中的细胞碎片沉淀。4)缓慢加入0.5倍上清体积的无水乙醇,混匀(此时可能会出现沉淀),将得到的溶液和沉淀一起转入已装入收集管的吸附柱(spincolumnsrm)中。4℃12,000rpm离心1分钟,弃废液,将吸附柱重新放回收集管中。5)向吸附柱rm中加入350μlbufferrw1,4℃12,000rpm离心1分钟,弃废液,将吸附柱重新放回收集管中。6)配制dnasei混合液:取52μlrnase-freewater,向其中加入8μl10×reactionbuffer和20μldnasei(1u/μl),混匀,配制成终体积为80μl的反应液。7)向吸附柱中直接加入80μldnasei混合液,20-30℃孵育15分钟。8)向吸附柱rm中加入350μlbufferrw1,4℃12,000rpm离心1分钟,弃废液,将吸附柱重新放回收集管中。9)向吸附柱rm中加入500μlbufferrw2,4℃12,000rpm离心1分钟,弃废液,将吸附柱重新放回收集管中。10)重复步骤9。11)4℃12,000rpm离心2分钟。12)将吸附柱rm装入新的rnase-freecentrifugetubes(1.5lml)中,向吸附膜的中间部位悬空滴加30-50μllrnase-freewater,室温放置2分钟,4℃12,000rpm离心1分钟,得到的rna溶液保存在-70℃。rna电泳图详见图1。1.2引物设计以电子克隆得到的2171bp的cdna序列为基础,利用dnaman6.0和primer5.0软件设计引物,由上海生工生物工程技术有限公司合成。基因克隆引物信息1.3目的片段的扩增以合成的cdna第一链为模板,p1f、p1r为引物进行保守区扩增,用3′gsp与3p、3np为上下游引物进行3′rt-pcr2轮扩增;以5′racecdna为模板,用5′gsp与3p、3np为上下游引物进行5′race扩增。pcr反应体系为:cdna模板1μl,2×taqpcrmastermix12.5μl,上游引物1μl,下游引物1μl,加ddh2o9.5μl至总体积25μl。pcr反应程序如下:94℃预变性3min,94℃变性1min,tm℃退火1min,延伸tmin,72℃再延伸10min结束反应,35个循环。1.4目的片段的回收、连接转化及测序用dna回收试剂盒回收目的片段,将目的片段与pmd18-t载体连接并转化dh5α感受态细胞,转化产物涂在含有1μlamp:1mllb的平板中培养,12h后挑起单菌落进行重组子菌液鉴定,阳性克隆子送到北京六合华大基因科技股份有限公司测序。1.5序列分析用genbank上的blast检索已有序列与分析其保守结构域,采用dnaman6.0和primer5.0软件进行引物设计、序列拼接及翻译氨基酸。用expasy数据库中的protparam工具和protscale工具预测分析推测蛋白质理化性质,蛋白质跨膜结构预测与分析采用tmpred(http://www.ch.embnet.org/software/tmpred_form.html),磷酸化位点预测用netphos2.0(http://www.cbs.dtu.dk/services/netphos),利用mega5.0构建系统进化树。利用ncbi网站的cd-search服务(http://www.ncbi.nlm.nih.gov/structure/cdd/wrpsb.cgi)分析芒果果实磷脂酶dα氨基酸序列的保守结构域。1.6半定量rt-pcr分析对25℃条件下分别贮藏0、2、4、6、8、10天的芒果进行取样。4℃条件下分别贮藏0、3、6、9、12、15、18和21天的芒果进行取样。总rna的提取和反转录方法同1.1。1)主要仪器和试剂德国analytikjenaqtowere2.2荧光定量pcr仪引物由生工生物工程(上海)股份有限公司合成。rna提取试剂盒:康为世纪生物科技公司的ominiplantrnakit(dnasei)(cw2598s)反转录试剂:全式金transscriptone-stepgdnaremovalandcdnasynthesissupermix(#at311)定量pcr试剂:诺唯赞aceqqpcrsybrgreenmastermix6×dnaloadingdye,10×tae(400mmtris-acetateand10mmedta,ph8.0),等其他试剂均购于生工生物工程(上海)股份有限公司。2)内参基因详细信息no.genesymbol基因信息(如accession#)引物是否设计合成1gadph是3)目的基因信息no.genesymbol基因信息(如accession#)引物是否设计合成1ers1是4)引物序列5)rna提取5.1实验试剂康为世纪生物科技公司的ominiplantrnakit(dnasei)5.2详细操作步骤具体操作步骤详见说明书。5.3rna电泳检测结果见图2。附图中,电泳图顺序依次为:电泳图顺序依次为:空白组处理组0、2、4、6、8天,1-mcp处理组0、2、4、6、8天。2.2.反转录2.2.1实验试剂transscriptone-stepgdnaremovalandcdnasynthesissupermix(北京全式金生物技术有限公司)。2.2.2cdna第一链合成和gdna去除1.将rna模板、primermix、2×tsreactionmix、rt/rienzymemix、gdnaremover和rnase-freewater溶解并置于冰上备用。2.根据以下表格配置反应体系,总体积为20μl。试剂20μl反应体系终浓度rnatemplate4μl1μg-5μgprimermix1μl2×tsreactionmix10μlrt/rienzymemix1ulgdnaremover1ulrnase-freewaterto20ul3.先将rna模板、引物与rnase-freewater混匀,65℃孵育5分钟,冰浴2分钟,然后再加入其它反应组分。4.轻轻混匀,25℃10分钟,50℃15分钟,85℃5秒失活。反应结束后,短暂离心,置于冰上冷却。5.逆转录产物直接用于荧光定量pcr反应。2.3.荧光定量pcr检测2.3.1实验样本将cdna样品稀释10倍作为模板上机检测。2.3.2实时定量pcr试验材料及仪器定量pcr试剂:aceqqpcrsybrgreenmastermix定量pcr仪:德国analytikjenaqtowere2.22.3.3pcr反应步骤配制反应混合液pcr循环条件仪器的操作完成上述步骤后,把加好样品的96孔板放在analytikjenaqtowere2.2荧光定量pcr仪中进行反应。实验结果上述实验结果,样品的样品的gadph扩增曲线见图3,样品的miers1扩增曲线见图4,样品的gadph溶解曲线见图5,样品的miers1溶解曲线见图6。实施例2芒果乙烯受体基因miers1的验证或表达特性分析用protparam工具分析miers1核苷酸推导的氨基酸序列,结果表明:miers1蛋白分子总量为80518.8,等电点pi6.34,负电荷和正电荷的氨基酸残基总数分别为77和71,分子式为c3595h5734n992o1046s29,总原子数为11396,脂肪系数103.73,不稳定系数38.96,亲水性为0.025,推测该蛋白为一个稳定的疏水蛋白。protscale分析亲水性和疏水性预测表明:miers1最大疏水性值为+4.500,最小亲水性值为-4.500,并且有20处最大疏水峰,值在+2.000以上;14个亲水峰,值在-2.000以下。其疏水性氨基酸明显多于亲水性氨基酸,且软件预测结果为疏水性,故推测蛋白整条肽链呈现疏水性。经netphos2.0软件分析miers1蛋白磷酸化位点,miers1肽链中的ser、thr和tyr均有可能发生磷酸化,分之在0.5以上的氨基酸位点有89个,其中,含有ser磷酸化位点有35个,分别位于肽链第8、9、10、14、24、26、78、112、124、125、146、187、217、237、301、316、348、398、399、420、421、435、441、467、481、485、517、523、570、572、575、608、638、653、660处;thr磷酸化位点有29个,分别位于肽链第1、6、7、41、107、120、170、174、188、193、197、198、238、252、256、267、416、426、483、611、621、647、663处;tyr磷酸化位点有12个,分别位于肽链第305、484、511、657、704处。经tmpred推测,miers1蛋白是跨膜蛋白,蛋白miers1的n末端在氨基酸54-75,85-105,114-141及607-625处有跨膜区域。经signaip4.0serve分析miers1蛋白,结果表明在整个肽链上不含有信号肽,推测蛋白均为非分泌蛋白。利用coils程序分析miers1蛋白的卷曲螺旋结构,以window14,21和28为参数,按照几率>50%可形成螺旋的原则,结果表明:miers1蛋白在氨基酸150-180及340-380之间形成卷曲螺旋结构的可能性很大。蛋白的保守结构域含有gaf结构域、hiska结构域、hatpase_c结构域及rec结构域。经nnpredict软件预测分析miers1蛋白质的二级结构,结构显示:miers1蛋白含有α-螺旋、β-转角和无规则卷曲3种结构,其中α-螺旋所占的比例分别为43.45%;β-转角占7.52%;无规则卷曲占29.94%。故miers1蛋白二级结构主要以α-螺旋为主。经swiss-model工具对miersl蛋白的三维结构进行预测,预测结果如图7所示。miers1蛋白系统进化树构建如图8所示。实施例31-甲基环丙烯抑制基因miers1表达对芒果衰老影响研究1.1-mcp处理对miers1表达的影响1-mcp即1-甲基环丙烯(1-methylcyclopropene,1-mcp)是一种新型乙烯受体抑制剂,具有无毒无味,使用方便,成本低廉,安全高效等优点。取1-mcp处理新采摘的芒果样品,按常规方法进行处理,处理时间分别为0、2、4、6、8、10、12、14、16天,同时设置空白对照组。每个时间点到之后,分别取芒果样品按实施例1和2方法提取miers1基因,检测miersl基因的表达水平。结果表明,经1-mcp处理后miersl表达水平显著降低,且于4-10天降低最为明显。结果如图9所示。2.1-mcp处理对芒果中乙烯释放量的影响取新采摘的广西芒果台农1号,分别于25℃和4℃条件下贮藏,采用5μl/l的1–mcp处理芒果0-21天后,检测芒果内乙烯释放量和按实施例1方法检测miersl表达水平。同时设置空白组。实验结果表明,贮藏25℃和4℃贮藏条件下,1-mcp处理有利于延缓芒果内miers1表达水平,延缓和降低了乙烯释放高峰和呼吸速率,从而对维持果实品质和延长果实采后寿命有显著的影响。其中在25℃下,经1-mcp处理的芒果乙烯释放量均低于未处理的,释放高峰也延后;而在4℃时,经1-mcp处理的芒果乙烯释放量在16天前均低于未处理的,16天后反而高于未处理的芒果样品,且释放高峰也延后达5个天以上。结果图10为1-mcp处理对芒果25℃贮藏条件下乙烯产生速率的影响;图11为1-mcp处理对芒果4℃贮藏条件下乙烯产生速率的影响。可以理解,因为1-戊基环丙烯、1-已基环丙烯、1-丙基环丙烯和1-辛基环丙烯均具有1-甲基环丙烯的基础结果环丙烯,只是取代基发生变化,属于类似物,在化学上也属于结构相似的类似物,在化学实验中常可以替换使用,故本发明中,也可以采用1-戊基环丙烯、1-已基环丙烯、1-丙基环丙烯和1-辛基环丙烯中的任一种取代1-甲基环丙烯进行试验,也可同时选择1-戊基环丙烯、1-已基环丙烯、1-丙基环丙烯、1-辛基环丙烯和1-甲基环丙烯中的两种或多种的混合液进行试验,均可达到本发明目的,不影响本发明的实施。上述说明是针对本发明较佳可行实施例的详细说明,但实施例并非用以限定本发明的专利申请范围,凡本发明所提示的技术精神下所完成的同等变化或修饰变更,均应属于本发明所涵盖专利范围。sequencelisting<110>广西壮族自治区农业科学院农产品加工研究所<120>一种芒果乙烯受体miers1基因<160>2<170>patentinversion3.3<210>1<211>2171<212>dna<213>人工序列<400>1acatggggacgtgtaacttgaacaagcagctctcttgcggagagcttagaatggaaagct60cagattaatcgctcattctccgatccataaggcaattgttcaatgatatcttgtgattgc120attgatacacaattggcccaggatgaactcctggtgaaatatcagtatatctcagatgta180ttgattgcccttgcgtatttctccattcccttggagcttatatatttcgtgaagaagtca240gcattctttccatatagatgggtgctaatgcagttcggtgcttttattattctttgtgga300gcaacccacttcataagcttgtggacgttctctctgcactccaaagccgtcgctgtggtg360atgactgtcgcaaagagttcctgtgctctagtgtcatgtctaactgctttgctacttgtt420tacatcattcctgatcttttgagtgtcagaactcgggaattattcctgaagaacagggct480gaagagcttgaccgggagatgggccttatccttactcaagaagaaactggaaggcatgtt540agaatgctgactcatgaaattaggagcacactcgatagagatacaattttaagaaccact600cttgttgagctaggtaggaccttaggcttggaggaatgtgcattgtggatgccttcaaga660actggtatcactttagaactttctcacactctgaaccatcaaatacaagttggatctacc720gttcctagaaaccttcctataattaatgacatctttggcactgctcgagcaaccttttta780ccctataactgcccactggcagggaccagacctgttgtgggaagacataggccttatgag840ggtgtcgctgttcgggtacctcttttacattactcaaatttccaaatcagcgattggcct900gacttctctgcaaaaagctatgcagtcatggttctgatcctccctactgacagtggtaga960aaatggagagatcatgagttggaacttgtggaagttgtagcagaccaggttgctgttgct1020ctttctcatgctgcaattctggaacaatctatgcgggctcgtgatcagctcttgcaacag1080aatgttgctttagatttggctcgacaagaggcagaaaaggcaatccatgctcgaaatgat1140ttccgcgctgtaatgaaccatgaaatgaggactctgatgcatgcagttattgcattgtct1200tcacttctcttggagacagaattaagtccagatcaaagggcaatgatagagacagttcta1260aagagtagtaacctcttgacaacacttgttgatgatgttttggatctttcccgacttgaa1320gatggtagtctagaattagaaattggaccattcaatcttcacgaagttttcagagaggtc1380attaatttgataaaacctattgcatctataaagaagttatccatgaatataaatatggcg1440ccagattcaccaacatactcaattggcgacaagaaacgactgatgcaaactattttgaac1500attgtgggtaatgctgtaaagttcacaaaggagggttatgttttgattaaagcttctgtt1560gcaaatccagagtcttcaagagactggcgacctgaatttcaccctgcgtcatgcaatggc1620cacttctaccttcgagtacaggttactgattctggctgtggtattctgccacaagacatt1680ccatttgtctttaccagatttgttcaatctcgaagtggctccaacagaagtacaggtgga1740gcaggacttggacttgccattagcagaaggtttgtcaatctcatggcagggcacatttgg1800attgagagtgaaggtcttgacaaaggtagcacagtaacatttcttgtccaacttgggata1860tgcaccacacaccaagctgtacccaaaggtcgagtaaatcatggaagtgcagatctctct1920ggacataaacaagtatttagagacaccgatgaggtagctttatccaaaacacgataccaa1980agaagcctttaggtcacagtgacagcagatgcagcattcagggaccagcttattcaggat2040atgatttttcctcatattcaagtaaaagctgaaggccgaaatcccggtgttacattatta2100catagagaagaatgaaaccaatataattatgaattagattttattataaactgagtaatg2160attaatctatg2171<210>2<211>629<212>prt<213>人工序列<400>1metilesercysaspcysileaspthrglnleualaglnaspgluleu151015leuvallystyrglntyrileseraspvalleuilealaleualatyr202530pheserileproleugluleuiletyrphevallyslysseralaphe354045pheprotyrargtrpvalleumetglnpheglyalapheileileleu505560cysglyalathrhispheileserleutrpthrpheserleuhisser65707580lysalavalalavalvalmetthrvalalalyssersercysalaleu859095valsercysleuthralaleuleuleuvaltyrileileproaspleu100105110leuservalargthrarggluleupheleulysasnargalagluglu115120125leuaspargglumetglyleuileleuthrglnglugluthrglyarg130135140hisvalargmetleuthrhisgluileargserthrleuaspargasp145150155160thrileleuargthrthrleuvalgluleuglyargthrleuglyleu165170175gluglucysalaleutrpmetproserargthrglyilethrleuglu180185190leuserhisthrleuasnhisglnileglnvalglyserthrvalpro195200205argasnleuproileileasnaspilepheglythralaargalathr210215220pheleuprotyrasncysproleualaglythrargprovalvalgly225230235240arghisargprotyrgluglyvalalavalargvalproleuleuhis245250255tyrserasnpheglnileserasptrpproasppheseralalysser260265270tyralavalmetvalleuileleuprothraspserglyarglystrp275280285argasphisgluleugluleuvalgluvalvalalaaspglnvalala290295300valalaleuserhisalaalaileleugluglnsermetargalaarg305310315320aspglnleuleuglnglnasnvalalaleuaspleualaargglnglu325330335alaglulysalailehisalaargasnasppheargalavalmetasn340345350hisglumetargthrleumethisalavalilealaleuserserleu355360365leuleugluthrgluleuserproaspglnargalametilegluthr370375380valleulysserserasnleuleuthrthrleuvalaspaspvalleu385390395400aspleuserargleugluaspglyserleugluleugluileglypro405410415pheasnleuhisgluvalphearggluvalileasnleuilelyspro420425430ilealaserilelyslysleusermetasnileasnmetalaproasp435440445serprothrtyrserileglyasplyslysargleumetglnthrile450455460leuasnilevalglyasnalavallysphethrlysgluglytyrval465470475480leuilelysalaservalalaasnprogluserserargasptrparg485490495progluphehisproalasercysasnglyhisphetyrleuargval500505510glnvalthraspserglycysglyileleuproglnaspileprophe515520525valphethrargphevalglnserargserglyserasnargserthr530535540glyglyalaglyleuglyleualaileserargargphevalasnleu545550555560metalaglyhisiletrpileglusergluglyleuasplysglyser565570575thrvalthrpheleuvalglnleuglyilecysthrthrhisglnala580585590valprolysglyargvalasnhisglyseralaaspleuserglyhis595600605lysglnvalpheargaspthraspgluvalalaleuserlysthrarg610615620tyrglnargserleu625当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 一种二化螟Bt蛋白受体ALP4基因及其应用
- 一种二化螟Bt蛋白受体ALP5基因及其应用
- 水稻ABA受体OsPYL3基因、其编码蛋白及其应用
- 检测表皮生长因子受体基因位点突变的荧光定量pcr试剂盒的制作方法
- 抗人雌激素受体单克隆抗体及抗人孕激素受体单克隆抗体的制备及应用的制作方法
- 子宫癌遗传检测试剂盒的制作方法
- 一种利用雌激素受体α基因预测优秀冰雪运动员身高的细胞生物学方法
- T1r味觉受体及其编码基因的制作方法
- 放射性配基N<sub>2</sub>S<sub>2</sub>-Memantine和脑受体显像剂<sup>99m</sup>Tc-Memantine的油水分配系数测定方法
- 猕猴雌二醇受体β基因片段及其应用的制作方法