巨桉EgrZFP6在提高植物在渗透胁迫下适应胁迫的用途的制作方法
本发明属于植物生物
技术领域:
,具体涉及巨桉egrzfp6在提高植物在渗透胁迫下适应胁迫的用途。
背景技术:
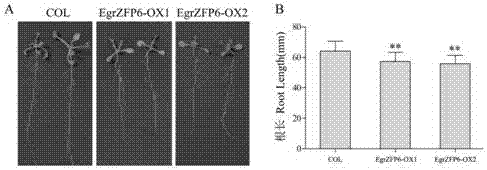
:低温、干旱等非生物胁迫会制约植物生长和发育,对农林业生产有严重影响。研究植物抗逆生理和分子机制对提高植物非生物逆境胁迫抗性具有重要意义(dosreisetal.,2012;roychoudhuryetal.,2015)。为了应对逆境胁迫,植物在长期进化过程中形成了一定的响应机制,植物感受到逆境信号后,通过相应基因调控,改变一系列代谢过程,产生应对胁迫的响应,如提高可溶性糖含量,增加抗渗透胁迫物质如脯氨酸、甜菜碱等,进而提高植物抗逆性(takabe,2012)。在植物抗逆分子响应机制中,作为基因开关的转录因子(transcriptionfactor,tf)发挥重要作用。植物中转录因子数量庞大,相当一部分成员与逆境调控相关,如bzip、wrky、ap2/erebp、myb和nac等,近年来锌指类转录因子(zincfingerprotein:zfp)也被证明在植物逆境胁迫响应中发挥重要作用(gujjaretal.,2014;reddyetal.,2013)。根据zfp转录因子具有的半胱氨酸(c)和组氨酸(h)残基数量和位置,可将锌指蛋白转录因子分为c2h2,c2h,c2c2,c2hcc2c2,c2c2c2等类型。其中c2h2型锌指蛋白是植物中研究较多、功能较为明确的一类锌指蛋白。其锌指结构域由约30个氨基酸组成,包含两个半胱氨酸和两个组氨酸,以及一段植物所特有的高度保守序列(qalggh)(kuboetal.,1998;黄骥等,2004)。c2h2型锌指蛋白在植物中成员众多,拟南芥(arabidopsisthaliana)中发现176个成员(englbrechtetal.,2004),水稻(oryzasativa)基因组数据显示水稻上有182个(agarwaletal.,2007)。c2h2型锌指蛋白广泛参与植物生长、发育和代谢,以及植物对低温,高盐和干旱等非生物逆境的响应(agnieszkaand2012)。水稻oszfp1作为负调控因子,能抑制盐胁迫相关基因表达,转基因植株对盐胁迫抗性降低。同时,其逆境响应还受脱落酸(aba)影响,暗示其可能参与aba依赖的逆境响应过程(kongetal.,2004)。水稻另外一个c2h2型锌指蛋白转录因子基因oszfp245的超表达则能够提高植株对低温、干旱和氧化逆境的抗性(huangetal.,2009)。矮牵牛(petuniahybrid)基因zpt2-3也可被低温、干旱和重金属等非生物胁迫诱导表达,其转基因植株干旱耐受力有效提高(suganoetal.,2003)。这些表明c2h2型锌指蛋白转录因子能够在植物非生物逆境胁迫响应中发挥作用。目前植物c2h2型锌指蛋白研究主要集中在模式植物中,在林木中研究较少。巨桉(eucalyptusgrandis)作为世界三大用材树种之一,在我国南方各省栽培广泛,为我国林业产业发展作出重要贡献。但其对低温敏感,不耐干旱、盐渍,这严重限制其栽培范围的扩大;低温、干旱等不良天气条件还经常对桉树生产造成损失。因此,研究其非生物逆境胁迫响应分子机制对桉树抗逆分子育种具有重要意义。技术实现要素:为了解决上述问题,本发明提供巨桉c2h2型锌指结构蛋白转录因子egrzfp6在提高植物在渗透胁迫下适应胁迫的用途。在本发明一个实施方案中,在植物中过表达巨桉c2h2型锌指结构蛋白转录因子egrzfp6,在渗透胁迫下改变植株根构型,提高植物在渗透胁迫下适应胁迫的用途。其中,所述改变植物根构型包括增加侧根数量以及促进侧根的伸长生长。其中,所述的逆境胁迫包括但不限于干旱、高盐等渗透胁迫。另一方面,本发明还提供一种提高植物在渗透胁迫下的适应性的方法,向目标植物转入巨桉c2h2型锌指结构蛋白转录因子egrzfp6,并使其在目标植物体内进行过量表达。其中,所述的渗透胁迫包括但不限于干旱,高盐等。另一方面本发明还提供一种在渗透胁迫下改变植株根构型的方法,其特征在于,向目标植物转入巨桉c2h2型锌指结构蛋白转录因子egrzfp6,并使其在目标植物体内进行过量表达。在本发明一个实施方案中,将转入所述egrzfp6基因的目标植物进行干旱诱导,进一步促进侧根增加和伸长,改变植株根构型。其中,所述改变植物根构型包括增加侧根数量以及促进侧根的伸长生长。其中,可采用本领域已知的所有方法向目标植物转入巨桉c2h2型锌指结构蛋白转录因子egrzfp6,包括但不限于,农杆菌介导、基因枪等。本发明构建35s::egrzfp6超表达载体,采用花序侵染法进行拟南芥遗传转化;和对照相比,egrzfp6超表达拟南芥转化植株在peg(1g·l-1以上)处理下,能大幅度促进侧根增加和伸长,改变植株根构型。附图说明图1所示为巨桉egrzfp6蛋白序列与其他植物同源蛋白序列比对。锌指结构域,l-box和ear基序用加粗黑线标示。meszat11:木薯manihotesculenta(oay41302.1);mtrzat11:蒺藜苜蓿medicagotruncatula(xp_013468307.1);grazat11:雷蒙德氏棉gossypiumraimondii(xp_012483278.1);luszat11:亚麻linumusitatissimum(xp_002533000);atzat11:拟南芥arabidopsisthaliana(aec09397.1)图2egrzfp6的亚细胞定位。图3所示为野生型和超表达egrzfp6拟南芥转基因株系鉴定。col:野生型;egrzfp6-ox1和egrzfp6-ox2:超表达株系。图4所示为野生型和超表达egrzfp6拟南芥转基因株系生长7天后表型(a)和根长差异(b)。图5所示为盐胁迫下野生型和超表达egrzfp6基因拟南芥表型。图6所示为模拟干旱胁迫下野生型和超表达egrzfp6基因拟南芥。具体实施方式以下实例用于说明本发明,但不用来限制本发明的范围。实施例11.材料与方法1.1材料巨桉幼苗生长在浙江农林大学苗圃地。所选试验材料为长势一致的6个月苗龄幼苗。拟南芥野生型为col。实验处理均在snijders微气候控制生长箱(mc1000,荷兰)中进行。培养条件为,拟南芥:白天24℃15h,夜间22℃9h;巨桉:白天25℃15h,夜间22℃12h。光照强度150μmol·m-2s-1,相对湿度(rh)70%。1.2egrzfp6蛋白序列分析在巨桉数据库phytozome(http://www.phytozome.net/search.php)下载egrzfp6全长序列,设计引物,进行全长序列测序验证后,将其编码蛋白序列在https://blast.ncbi.nlm.nih.gov/blast.cgi进行blast比对,选择5个与该蛋白序列相似程度比较高的不同物种蛋白序列,利用clastalx1.83进行多重比对。同时,用cdd在线软件(https://www.ncbi.nlm.nih.gov/structure/cdd/wrpsb.cgi)分析蛋白序列保守域。1.3egrzfp6亚细胞定位设计引物(表1),pcr扩增去掉终止密码子的egrzfp6开放阅读框序列,以pcambia1300为载体骨架,利用多克隆位点处的kpni和xbai酶切位点,构建egrzfp6-gfp融合蛋白表达载体。提取质粒,金粉包埋后轰击洋葱(alliumcepa)表皮细胞。激光共聚焦显微镜扫描成像。1.4egrzfp6超表达载体构建、异源转化拟南芥和超表达拟南芥低温、盐和peg处理超表达载体骨架为含35s启动子的pcambia1301,egrzfp6全长序列扩增引物见表1,利用多克隆位点处的kpni和xbai酶切位点,双酶切法构建35s::egrzfp6超表达载体。提取质粒,选取健壮盛花期拟南芥采用农杆菌蘸花法转化拟南芥(许红梅等,2010)。收获侵染植株种子,于含20mg·l-1潮霉素的1/4ms培养基上(hygromycin)进行筛选,阳性植株继续繁殖,收获,筛选,pcr检测,直到获得t3代转基因纯合株系。提取10天苗龄的野生型和两个超表达egrzfp6拟南芥纯合株系rna,逆转录后,设计引物(表1)以atactin为内参基因进行半定量rt-pcr,鉴定egrzfp6在转基因株系中的表达情况。野生型和超表达egrzfp6拟南芥种子均匀播种在1/2ms培养基上,4℃暗处理24h后置于生长箱中生长。一周后,比较和分析野生型和超表达株系表型变化;参考zhao等(2016)的方法,将10天苗龄的幼苗于-8℃低温处理3天,然后正常培养条件下缓苗3天,拍照观察表型,统计植株死亡率;盐(nacl)处理梯度为0mmol·l-1,50mmol·l-1和100mmol·l-1;使用peg6000模拟干旱处理,处理梯度为1g·l-1,5g·l-1和9g·l-1,种子发芽后,于生长箱中生长一周,观察表型并拍照(nikon,d7000)。1.5rna提取,cdna合成及基因定量表达分析巨桉rna提取参考ctab+trizol法(王亚红等,2010),拟南芥rna提取参照trizol法(金美芳,2004)。cdna反转录试剂盒由康为世纪公司提供,定量荧光染料sybr由takara公司(大连,中国)提供。设计引物(表1)用bio-radcfx96定量rt-pcr系统(biorad,美国)以及系统自带的bio-radcfxmanager(ver1.5.5.34)软件进行实验和结果分析。表1相关引物序列引物名称引物序列(5′-3′)egrzfp6-sgfp-fcggggtacctcaacgacattctcttcagcaegrzfp6-sgfp-rgctctagacaggagcaaggcatctatct35s-egrzfp6-fcggggtaccgaaaaggcaccccacaaa35s-egrzfp6-rgctctagacctaaactcagtcggtccaaaatactin-rt-ftgcccatcgggtaattcatagttcatactin-rt-rcctcatgccatcctccgtcttegrzfp6-rt-fatcccaagatgcacgagtgctcegrzfp6-rt-rcggaccaaccacgaaaatctca2结果与分析2.1egrzfp6编码蛋白结构分析egrzfp6编码蛋白含有2个锌指结构域,对该基因蛋白序列进行保守结构域搜索,并将其与其他不同植物中的同源蛋白比对分析结果表明,该基因编码蛋白含有2个高度保守的锌指蛋白结构域,且2个锌指结构域核心序列均为qalggh,该序列是植物锌指蛋白所特有的(takatsuji,1999)。另外,egrzfp6蛋白序列还分别含有1个乙烯响应元件结合因子相关双性抑制子(erfassociatedamphiphilicrepression:ear)基序:dlnltp和1个l-box基序(图1)。2.2egrzfp6亚细胞定位结果分析构建egrzfp6-gfp表达载体,利用基因枪轰击洋葱表皮的方法对egrzfp6蛋白进行亚细胞定位分析,结果表明该基因表达的蛋白定位于细胞核中(图2)。其可能是作为转录因子在细胞核内发挥基因表达调控作用。2.3超表达egrzfp6转基因拟南芥株系鉴定半定量rt-pcr分析结果(图3)表明,野生型col中没有检测到egrzfp6表达,在两个转基因纯合株系egrzfp6-ox1和egrzfp6-ox2中egrzfp6具有明显表达(图3)。2.4超表达egrzfp6基因拟南芥株系正常条件及低温、高盐和peg处理下表型分析超表达egrzfp6拟南芥2个纯合株系egrzfp6-ox1和egrzfp6-ox2,播种7天后和对照(col)相比,根长生长明显受到抑制(图4a)。2个转基因株系的根长分别为野生型的89.4%和87.0%(图4b)。盐处理下,egrzfp6超表达拟南芥株系与野生型拟南芥随盐浓度提高,主根、侧根生长都表现为受盐抑制,但受抑制程度相对于野生型有所减弱,表示超表达egrzfp6在一定程度上能够提高植株对盐胁迫的耐受性(图5)。在peg处理中,1g·l-1以上的浓度即能诱导egrzfp6超表达拟南芥株系侧根大量发生,同时侧根伸长作用也明显被促进。但伸长和对照相比,在peg处理下并没有受到明显抑制,表示超表达egrzfp6株系根长生长对渗透胁迫的抑制作用敏感性下降(图6),9g·l-1时侧根数目增多为野生型3.2倍以上,5g·l-1侧根长度为野生型的4.0倍左右。以上所述仅是本发明的优选实施方式,应当指出,对于本
技术领域:
的普通技术人员来说,在不脱离本发明技术原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。sequencelisting<110>浙江农林大学<120>巨桉egrzfp6在提高植物在渗透胁迫下适应胁迫的用途<130><160>8<170>patentinversion3.3<210>1<211>30<212>dna<213>人工序列<400>1cggggtacctcaacgacattctcttcagca30<210>2<211>28<212>dna<213>人工序列<400>2gctctagacaggagcaaggcatctatct28<210>3<211>27<212>dna<213>人工序列<400>3cggggtaccgaaaaggcaccccacaaa27<210>4<211>29<212>dna<213>人工序列<400>4gctctagacctaaactcagtcggtccaaa29<210>5<211>24<212>dna<213>人工序列<400>5tgcccatcgggtaattcatagttc24<210>6<211>21<212>dna<213>人工序列<400>6cctcatgccatcctccgtctt21<210>7<211>22<212>dna<213>人工序列<400>7atcccaagatgcacgagtgctc22<210>8<211>22<212>dna<213>人工序列<400>8cggaccaaccacgaaaatctca22当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1