双特异性嵌合抗原受体c-Met/PD-1scFv-CAR-T及其构建方法和应用与流程
本发明属生物技术领域,涉及抗原受体c-met/pd-1scfv-car-t及其构建方法,具体涉及双特异性嵌合抗原受体c-met/pd-1scfv-car-t及其构建方法和应用。本发明利用基因工程技术获得含双特异性重组嵌合抗原受体的融合基因,感染t细胞后使t细胞表达该双特异性嵌合抗原受体。该双特异性嵌合抗原受体t细胞能够识别肿瘤细胞表面的c-met及阻断激活t细胞表面的抑制性受体pd-1,避免激活t细胞被诱导凋亡,保持持续激活能力,从而特异性杀伤c-met阳性的肿瘤细胞,可应用于实体肿瘤的免疫治疗。
背景技术:
现有技术公开了正常的免疫反应过程中,未致敏t细胞至少需要两种信号的刺激才能够被激活并产生特异性免疫应答;其中第一个信号是t细胞借助t细胞受体(tcr)识别与mhc-i或ii类分子结合的抗原,通过tcr/cd3复合体传递抗原识别信号;第二个信号是以cd28为主的t细胞表面受体分子识别抗原提呈细胞(apc)或其他细胞表面的共刺激分子,传递协同刺激信号。研究表明,肿瘤特异性t细胞在机体清除肿瘤细胞的免疫反应中发挥主要作用,同时肿瘤细胞通过抑制参与t细胞识别及抗原反应的分子表达、降低免疫原性等途径产生免疫逃逸,从而使机体的免疫系统不能发挥清除肿瘤细胞的作用。
有研究发现,以肿瘤特异性单克隆抗体的单链可变区(scfv)取代tcr的α、β链可变区,并将scfv直接和共刺激分子(如cd28、cd137等)以及t细胞信号传导区域cd3ζ连接,形成嵌合抗原受体(car)表达于t细胞表面,可通过scfv识别肿瘤特异性抗原,直接产生t细胞活化的第一、第二信号,促进t细胞活化、增殖,从而特异性杀伤肿瘤细胞。该过程主要依赖car-t细胞表面的scfv对肿瘤抗原的特异性识别,免疫反应的特异性和杀伤性较强。
目前,car-t细胞在血液系统恶性肿瘤治疗中取得显著疗效,但在实体瘤的临床应用上却面临着诸多挑战。主要障碍之一是由于实体肿瘤的免疫抑制微环境,即通过上调抑制性受体来抑制肿瘤浸润性淋巴细胞(til)和基因工程修饰t细胞的功能,特别是免疫抑制性受体pd-1的上调。pd-1是目前研究的最多的一个免疫抑制性受体,其表达在t细胞和前体b细胞表面,属于免疫球蛋白超家族的一员。pd-l1是目前已发现的pd-1主要配体,研究发现pd-l1在多种肿瘤细胞膜表面的表达水平升高,肿瘤细胞巧妙地利用pd-l1去结合t细胞表面的pd-1受体来传递抑制性信号,从而导致免疫逃逸。
有临床前研究和临床试验均指出,采用pd-l1或pd-1阻断抗体来增强癌症患者的抗肿瘤免疫应答可作为一种新型的肿瘤免疫治疗手段,并且能够获得持续性的临床应答。此外,有研究提示,car-t细胞治疗联合抗pd-1抗体可以大大增强car-t细胞的抗肿瘤效应,那么在car-t细胞膜表面表达pd-1-scfv-car融合受体,即采用pd-1scfv封闭car-t细胞表面的pd-1受体,从而避免pd-l/pd-1的抑制信号,增强car-t细胞治疗的效用?利用抗c-metscfv和抗pd-1scfv构建的双特异性嵌合抗原受体hc-metscfv-linker-pd-1scfv-cd8α-cd28-cd137-cd3ζ表达于t细胞表面,可使t细胞特异性杀伤c-met阳性的肿瘤细胞,同时封闭t细胞表面的抑制性受体pd-1,使t细胞持续激活,因此双特异性嵌合抗原受体修饰的t细胞有望打破过继传输t细胞治疗中的pd-l/pd-1的免疫抑制信号轴,在实体肿瘤免疫治疗领域具有广阔的应用前景。
基于现有技术的现状,本申请的发明人拟提供双特异性嵌合抗原受体c-met/pd-1scfv-car-t及其构建方法和用于实体肿瘤的免疫治疗中的用途。
技术实现要素:
本发明的目的是要解决现有技术存在的技术问题,提供一种双特异性嵌合抗原受体c-met/pd-1scfv-car-t及其构建方法和用于实体肿瘤的免疫治疗中的用途。
本发明构建了一种抗hc-metscfv-pd-1scfv的嵌合抗原抗受体(hc-metscfv-linker-pd-1scfv-cd8α-cd28-cd137-cd3ζ),表达该嵌合抗原受体的t细胞具有其独特性,经实验证实其对肿瘤细胞特异性杀伤功能,具有临床治疗中的应用前景。
本发明利用基因工程技术获得一种含双特异性重组嵌合抗原受体hc-metscfv-pd-1scfv-cd8α-cd28-cd137-cd3ζ的融合基因,将该基因片段插入慢病毒表达载体系统,通过对慢病毒进行包装获得含病毒的上清,感染t细胞,使t细胞表达该双特异性嵌合抗原受体。这种双特异性嵌合抗原受体t细胞(car-t)能够识别肿瘤细胞表面的c-met及阻断激活t细胞表面的抑制性受体pd-1,避免激活t细胞被诱导凋亡,保持持续激活能力,从而特异性杀伤c-met阳性的肿瘤细胞,可应用于实体肿瘤的免疫治疗。
更具体的,本发明采用的技术方案是:双特异性嵌合抗原受体hc-metscfv-linker-pd-1scfv-cd8α-cd28-cd137-cd3ζ,所述嵌合抗原受体由抗人c-met及pd-1单克隆抗体轻链和重链可变区hc-metscfv、pd-1scfv、人cd8α铰链区、人cd28跨膜区和胞内区、人cd137及cd3ζ胞内区结构串联构成;所述双特异性嵌合抗原受体的氨基酸序列如序列表中seqidno.1所示;
所述双特异性嵌合抗原受体的核酸序列如sqeidno.2所示;
所述双特异性嵌合抗原受体含有针对人c-met抗原的单链抗体hc-metscfv,其氨基酸序列如seqidno.3所示,单链抗体的重链和轻链之间用15个氨基酸的连接短肽连接;
所述嵌合抗原受体含有针对人c-met抗原的单链抗体hc-metscfv,其核酸序列如seqidno.4所示;
所述嵌合抗原受体含有针对人pd-l1的受体pd-1胞外结合区,其氨基酸序列如seqidno.5所示;
所述嵌合抗原受体含有针对人pd-l1的受体pd-1胞外结合区,其核酸序列如seqidno.6所示;
所述嵌合抗原受体的氨基末端含有一个cd8α信号肽,其氨基酸序列如序列表中的seqidno.7所示;
所述嵌合抗原受体的氨基末端含有一个cd8α信号肽,其核酸序列如序列表中的seqidno.8所示。
本发明进一步提供了所述的双特异性抗原受体hc-metscfv-linker-pd-1scfv-cd8α-cd28-cd137-cd3ζ在制备嵌合抗原受体t细胞及其在肿瘤免疫治疗中的应用。
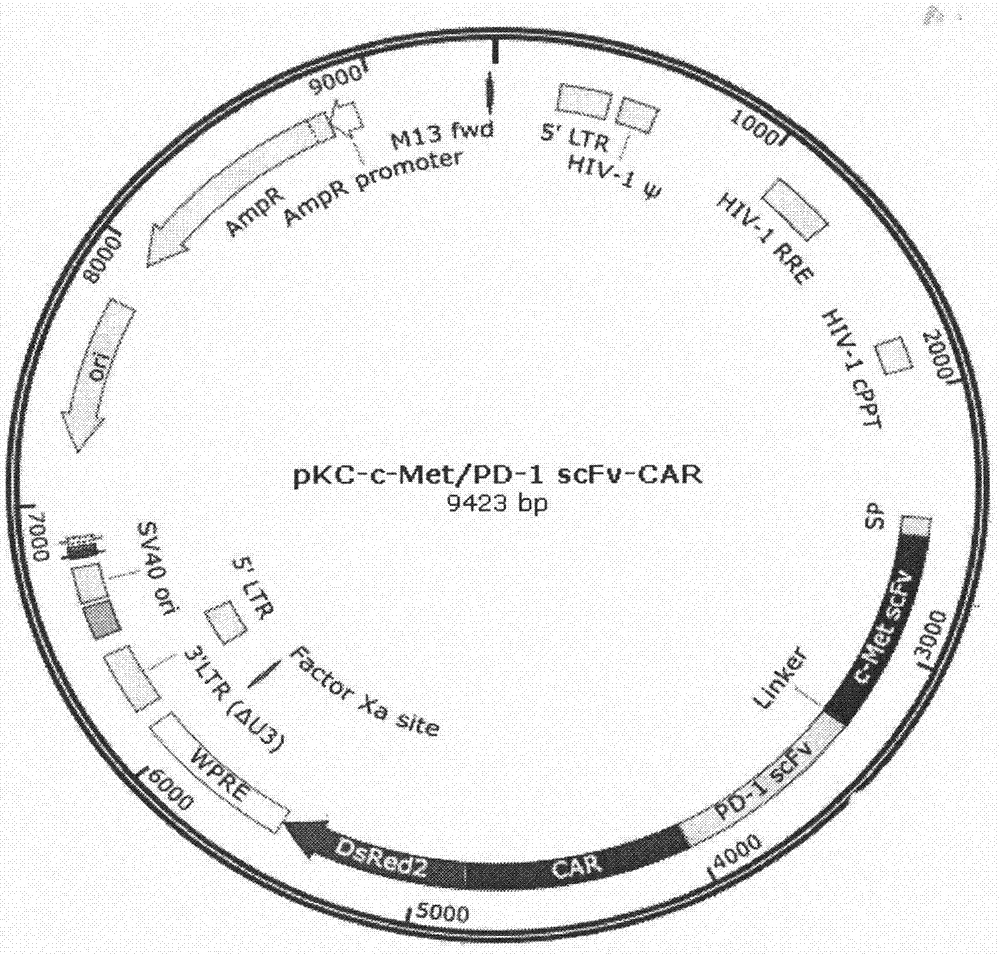
本发明采用抗人c-metscfv及pd-1scfv氨基酸序列,将其进行密码子优化,从ncbigenbank数据库中搜索到人cd8α信号肽基因、人cd8α铰链区基因、人cd28跨膜区和胞内区基因、cd137以及cd3ζ胞内区基因序列信息,全基因合成cd8α信号肽、cd8α-cd28-cd137-cd3ζ基因片段,然后通过重组酶连接成双特异性嵌合抗原受体hc-metscfv-pd-1scfv-cd8α-cd28-cd137-cd3ζ,插入到慢病毒表达载体pkc-ef1α-ds-red(pkc-空载体)中,构建抗人c-met/pd-1scfv-car表达载体(如图1所示);外送测序完全符合设计(如图2所示);利用该质粒与慢病毒的包装质粒vsvg和r-8.91在293t细胞中包装病毒,感染jurkat细胞(如图3所示);使jurkat细胞表达该嵌合抗原受体。获得的car-jurkat细胞在体外与c-met及pd-l1阳性的肿瘤细胞mkn45共培养,elisa方法检测共培养上清中的细胞因子水平(如图4所示);
本发明的嵌合抗原受体用于修饰t淋巴细胞,修饰后的t细胞(car-t细胞)用于制备实体肿瘤免疫治疗制剂,尤其是能用于表面c-met及pd-l1阳性的肿瘤治疗制剂;所述的双特异性嵌合抗原受体hc-metscfv-pd-1scfv-cd8α-cd28-cd137-cd3ζ。在肿瘤的免疫治疗中具有广阔的应用前景,
附图说明
图1为pkc-car慢病毒表达载体示意图。
图2为pkc-car慢病毒表达载体部分测序峰值图。
图3为荧光显微镜下观察慢病毒感染48h后jurkatt细胞的红色荧光(red)表达情况。
图4为car-t实验组、对照组细胞上清中il-2浓度比较。
图5为在不同效靶比例情况下,car-t实验组与对照组细胞毒性比较。
图6为car-t细胞对裸鼠移植瘤生长的影响。
具体实施方式
下面结合附图和具体实施方式对本发明作进一步详细说明:
实施例1:hc-metscfv-pd-1scfv-cd8α-cd28-cd137-cd3ζ基因序列的确定
从美国国立医学图书馆网站数据库中搜索到人pd-1胞外段基因、人cd8α信号肽基因、人cd8α铰链区基因、人cd28跨膜区和胞内区基因、cd137以及cd3ζ胞内区基因序列;抗c-met及pd-1单链抗体(抗c-metscfv及pd-1scfv)基因序列来自本实构建的单克隆抗体序列,将其进行密码子优化,以保证在编码氨基酸序列不变的情况下更适合t细胞的表达,各基因序列信息如sequencelisting(序列表seqidno.1-8)所示;
将上述基因序列依次按人cd8α信号肽基因、抗人c-metscfv、抗人pd-1scfv、car(包括人cd8α铰链区基因、人cd28跨膜区和胞内区基因、人cd137及cd3ζ胞内区基因序列)进行连接,在各序列连接处引入不同的酶切位点,最终形成完整的hc-metscfv-pd-1scfv-car基因序列信息。
实施例2
构建hc-metscfv-pd-1scfv-cd8α-cd28-cd137-cd3ζ表达质粒
叠合pcr合成基因片段:
利用叠合pcr合成c-metscfv、pd-1scfv、car序列,在各序列连接处引入不同的酶切位点,利用重组酶连成完整的c-metscfv-pd-1scfv-car序列,克隆到pkc-ef1α-ds-red慢病毒表达载体(pkc-空载体)中,获得抗人c-metscfv-pd-1scfv-car表达质粒(pkc-car质粒)和含有该质粒的dh5α菌液,质粒信息如图1所示;
重组质粒测序:
将重组质粒送上海美吉生物医药科技有限公司进行测序,将测序结果与拟合成的hc-metscfv-pd-1scfv-car序列比对以证实序列正确,测序引物pkc-ef1α-ds-red载体上的通用测序引物:
sense:5’-gactcagccggctct-3’
anti-sense:5’-ctcgccctcgatctc-3’
部分测序结果如图2所示。
实施例3:目的质粒和包装质粒的大量抽提
将pkc-car质粒、pkc-空载体质粒、以及vsvg和r-8.91包装质粒的菌株在lb培养液中大量培养,以碱裂解法大量提取质粒(北京天根生化科技有限公司无内毒素质粒提取试剂盒),以备转染。
1)将菌液按1∶1000的比例加入100ml含氨苄青霉素的lb培养液中,37℃恒温孵箱220rpm震摇12-16h;
2)取少量菌液测a600吸光度,达0.4左右停止震摇;
3)向吸附柱cp6中加入2.5ml的平衡液bl,8000rpm离心2min,倒掉收集管中的废液,将吸附柱重新放回收集管中;
4)将菌液于4℃8000rpm离心3min,尽量吸除上清,收集细菌沉淀;
5)加入8ml溶液p1(确定已加入rnasea),彻底悬浮细菌细胞沉淀;
6)加入8ml溶液p2,立即温和地上下颠倒6-8次,使菌体充分裂解,室温放置5min;
7)加入8ml溶液p4,立即温和地上下颠倒6-8次,充分混匀,至溶液出现白色分散絮状沉淀,室温放置10min,8000rpm离心10min;使白色沉淀离至管底,将全部溶液小心地倒入过滤器cs1中,慢慢推动推柄过滤,滤液收集在干净的50ml的离心管中;
8)向滤液中加入0.3倍滤液体积的异丙醇,上下颠倒混匀后转移到吸附注cp6中;
9)室温8000rpm离心2min,倒掉收集管中的废液,将吸附柱cp6重新放回收集管中;
10)向吸附柱cp6中加入10ml漂洗液pw(请先检查是否已加入无水乙醇),8000rpm离心2min,弃掉收集管中的废液,将吸附柱重新放回收集管中;
11)重复操作步骤10;
12)向吸附柱cp6中加入3ml无水乙醇,室温8000rpm离心2min,倒掉废液;
13)将吸附柱cp6重新放回收集管中,8000rpm离心5min,将吸附柱中残余的漂洗液去除;
14)在无菌超净内将吸附柱cp6置于一个新的50ml收集管中,相吸附膜中间部位悬空滴加2ml洗脱缓冲液tb,室温放置5min,然后8000rpm离心2min,将50ml离心管中的洗脱液全部分装入2个干净的1.5ml离心管,-20℃保存备用;
15)将取2μldna溶液紫外分光光度法测质粒dna浓度和a260/a280,并同时进行琼脂糖凝胶电泳分析质粒的质量。
实施例4:慢病毒的包装
细胞处理:转染前24h用胰酶消化、收集处于对数生长期的第4-12带293t细胞,将细胞转移入10cm培养皿中,细胞在10ml含有10%fbs、1%ps的dmem培养基中生长,置37℃5%co2培养箱进行培养24h,达60-70%汇合率时进行转染;
转染体系配置:
1)于1.5ml灭菌ep管内加入50μl无血清opti-mem培养基,再加入10μg质粒(pkc-car目的质粒或pkc-空载质粒∶vsvg∶r-8.91=4∶3∶3),充分混匀,室温静置5min;另一支1.5灭菌离心管中加入50μl无血清opti-mem培养基及30μl转染试剂lipofectamine2000(lipo2000∶总dna(μg)=2∶1),充分混匀,室温静置5min,然后将两管中溶液混匀,室温静置20min;
2)将原培养皿中培养液小心吸出,加入8ml无血清opti-mem培养基;
3)将上述质粒-lipo2000混合液加入10cm培养皿中,小心混匀;
4)将培养皿放入37℃5%co2培养12-16h,加入2mlfbs,继续培养;
5)转染24h后于荧光显微镜下观察293t细胞转染后red荧光表达情况,分别于转染后48h、2h收集细胞培养上清,室温3000rpm离心15min,收集上清;
6)用0.45μm滤器过滤病毒上清,分别获得pkc-空载和pkc-car病毒原液。
实施例5jurkatt细胞的培养、感染和扩增
1)孵苏本室冻存的t细胞来源的jurkatt细胞,传4代以后接种至24孔板,向培养24h的jurkat细胞中每孔加入终浓度为4μg/ml的polybrene预处理4h,然后加入pkc-空载体或pkc-car病毒浓缩液200μl,置于培养箱培养,24h内不换液;
2)感染24h后进行第二次感染,将细胞1000rpm离心10min,小心吸去上清,加入新鲜含10%fbs的rpmi1640培养液重悬后调整细胞浓度至1×106/ml,用polybrene预处理4h后加入200μlpkc-car病毒浓缩液继续培养;
3)继续培养96h后,用荧光显微镜观察jurkatt细胞的red红色荧光表达情况并照相。结果显示jurkat感染率达50%左右(如图3所示)。
实施例6
体外共培养测定car-jurkatt细胞分泌细胞因子及对肿瘤细胞杀伤能力实验
elisa法测定细胞因子水平:
elisa试剂盒(购自r&d公司),按试剂盒说明书操作。
1)将来自于c、d、e组培养36h的上清从-20℃取出,室温融化;
2)将浓缩洗涤液、稀释液、标准品、密封板条从冰箱中取出室温预温;
3)分别将浓缩洗涤液和稀释液用双蒸水稀释至工作浓度;
4)制备标准品:加入500ml稀释液至冻干标准品中,自然溶解至少20min,混匀,稀释成2000、1000、500、250、125、62.5、31.25、15.6和0pg/ml;
5)将样品和不同浓度的标准品加入相应孔中,100μl/孔,用封板胶纸封住反应孔,37℃孵育2h;
6)洗板5次,甩干板内液体;
7)将相应抗体平衡至室温,加入100μl/孔,用封板胶纸封住反应孔,37℃孵育1h;
8)洗板5次,甩干板内液体;
9)将显色底物平衡至室温,每孔加入100μl,37℃避光孵育30min;
10)将终止液平衡至室温,加入终止液50μl/孔,混匀后30min内酶标仪450nm(检测波长)和570nm(校正波长)波长测量吸光度值;
11)绘出4参数(4-pl)线性标准曲线,通过样品的od值查出其浓度。结果显示双特异性嵌合抗原受体修饰的jurkat细胞能够明显促进il-2及ifn-γ的分泌(如图4所示)。
cytotox96非放射性细胞毒性法检测car-jurkatt细胞对mkn45细胞的杀伤活性:
试剂盒购自promega公司,按说明书要求操作。
1)靶细胞制备:800rpm离心5min收集对数生长期mkn45细胞,用含5%fbs的rpmi1640调整细胞浓度至2×106/ml;
2)效应细胞制备:1000rpm离心10min收集感染后一周的car-jurkat,用含5%fbs的rpmi1640调整细胞浓度至2×106/ml。
3)在u底96孔板设置下列对照组和实验组(三复孔):
4)按不同效:靶比例(2∶1,1∶2,1∶4,1∶8和1∶16)加效应细胞和靶细胞,均用含5%fbs的rpmi1640补足体积为200μl/孔;
5)200g离心96孔平板4min,置37℃,5%co2,饱和湿度培养箱培养7h,提前45min在靶细胞最大释放孔加10μl细胞裂解液(10x),200g离心平板4min;
6)ldh活性测定;
7)ctl活性计算:计算各组(3复孔)平均吸光度(a490),按操作步骤计算ctl裂解靶细胞的百分率,结果显示双特异性嵌合抗原受体修饰的jurkat细胞杀伤靶细胞活性明显强于对照组(如图5所示)。
实施例7体内car-t细胞抑制肿瘤生长的能力实验
采用免疫磁珠分离人外周血中cd3+t细胞
1)选取健康志愿者,从肘静脉抽取20ml外周全血,首先分离出pbmc;
2)将分离得到的pbmc用含10%fbs的rpmi1640培养基重悬,并计数细胞浓度;
3)用macs缓冲液洗涤pbmc一次,1000rpm离心10min沉淀细胞,弃上清;
4)加入适量的anti-human-cd3免疫磁珠(20ul/107个pbmc))混匀,4℃孵育15min;
5)加入适量macs缓冲液,1000rpm离心10min,沉淀细胞用macs缓冲液重悬;
6)取出minimacs分离器,将ms分离柱放入分离器中,并用适量macs缓冲液洗涤ms分离柱;
7)将步骤(5)中的细胞悬液缓慢加入ms分离柱内,进行细胞分选,是未被cd3免疫磁珠标记的阴性细胞首先流出;
8)用适量macs缓冲液洗涤分离柱三次,每次都使柱子中的缓冲液流尽;
9)准备一支新的15ml离心管上,从minimacs分离器上取下ms分离柱,放置新的离心管上;
10)加入1mlmacs缓冲液于分离柱上,快速将滞留的细胞完全洗脱下来,重复三次,cd3免疫磁珠标记的阳性细胞最后全部洗脱下来;
11)收集的细胞离心后去上清,用含10%fbs的rpmi1640培养基重悬细胞,细胞计数用于后续实验。
t淋巴细胞的激活、扩增与感染实验
cd3+t淋巴细胞被病毒感染前需要激活与扩增,按cd3/cd28免疫激活磁珠说明书进行,具体操作步骤如下:
1)将上述分离得到的cd3+t细胞计数,调整浓度为2×106/ml,每孔总体积为500μl种植于24孔板中;
2)每孔加入25μl免疫磁珠,以获得磁珠与细胞比为1∶1;
3)加入重组人il-2,使其终浓度为30u/ml;
4)细胞每天检查,观察细胞的生长状况。每2-3天更换培养基,并重新加入重组人il-2;
5)当发现细胞生长密度较大时,收集细胞离心,重悬后计数并将密度调整为1×106/ml,传代至6孔板中继续培养;
6)培养1周后收集细胞,加入无血清的rpmi1640培养基重悬,加入6孔板中,总体积为2ml,用8μg/mlpolybrene预处理细胞4h;
7)4h后将适量浓缩慢病毒悬液缓慢均匀地加入6孔板中,24h后更换培养基;
8)每天使用荧光显微镜观察t细胞状态,在同一视野下用红色荧光及白光拍照,统计细胞感染效率,在感染第5天收集各组t细胞于离心管中,1000rpm离心5min;
9)弃上清后用无菌pbs重悬,清洗一遍,细胞活性鉴定经台盼蓝染色置于显微镜下观察计数;
10)最后用pbs制成1×108/ml的悬液用于输注裸鼠。
动物实验:
建立裸鼠移植瘤模型
1)6-8周龄的雄性裸小鼠饲养于学校动物房spf饲养室,移植肿瘤前至少提前1周购于上海斯莱克实验动物有限公司;
2)mkn45细胞用高糖dmem培养基(10%fbs,1%ps),37℃、5%co2培养箱中培养;
3)将处于对数生长期的mkn45细胞用0.25%胰酶消化、中和、离心,丢弃离心管上清;
4)用无菌pbs重悬细胞,1000rpm离心5min,丢弃上清,加入无菌pbs重选细胞并计数,调整浓度为5×107/ml的悬液;
5)用碘伏消毒裸鼠右侧背部皮肤,然后用75%的乙醇擦拭背部,等待片刻让酒精挥发,在每只裸鼠右侧背部皮下注射100μl的细胞悬液,轻轻压迫针孔防止细胞溢出,然后放回笼中继续培养;
6)每周观察小鼠2次,称体重、测肿块长度和宽度。计算肿瘤体积v=1/2×长×宽的平方。
car-t细胞的输注:
1)提前计算并准备car-t细胞,测量并计算裸鼠移植瘤体积,当生长达到50mm3左右时(大约注射后1周),将裸小鼠随机分成3组,每组5只;
2)将car-t细胞重悬于无菌pbs,通过尾静脉输注小鼠体内。第一组为对照组输入1×107t细胞;第二组为实验组1输入1×107pkc-c-met-car-t细胞;第三组为实验组2输入1×107pkc-c-met/pd-1-car-t细胞;
3)然后继续每周观察2次裸鼠饮食、排便及精神状态,称体重,测肿瘤长度与宽度,共6周;
4)水合氯醛麻醉裸鼠,剥离肿瘤组织并解剖裸鼠,统计各组数据进行分析,结果显示双特异性嵌合抗原受体修饰的t细胞能够显著抑体内肿瘤的生长(如图6所示)。
综上所述,本发明的内容并不局限在上述的实施例中,相同领域的有识之士可以在本发明的技术指导思想之内轻易提出其他的实施例,但这种实施例都包括在本发明的范围之内。
- 还没有人留言评论。精彩留言会获得点赞!