一种烟草尼古丁含量调控基因NtCLC‑b及其克隆方法与应用与流程
本发明属于遗传工程技术领域,进一步属于涉及烟草尼古丁的合成及调控相关基因,具体涉及一种烟草尼古丁含量的调控基因ntclc-b及其克隆方法与应用。
背景技术:
研究烟草尼古丁的代谢调控是非常有意义的一项工作,通过基因调控可以提供不同尼古丁含量的烟草品种,为烟草商业生产个性化尼古丁烟草产品提供原材料。尼古丁对人体有很强的生理刺激作用,是烟草商业性使用的物质基础。许多世界顶级烟草公司如菲利普莫里斯,帝国烟草、日本烟草、英美烟草等公司都投入巨资对烟草尼古丁的代谢途径、调控机制进行研究。
尼古丁是一种吡啶类生物碱,主要存在于茄科烟草属(nicotiana)植物中,是烟草体内的一种重要的次生代谢产物。烟草尼古丁的合成和转运受到多个因素的调控,目前已经鉴别和克隆出来了一些尼古丁合成途径中的关键基因,如qpt、pmt、mpo、jaz、myc2a等。
从分子生物学角度对尼古丁的合成代谢途径研究尚未完全透彻。通过氯离子通道调控尼古丁合成基因,从而影响尼古丁含量的研究未见报道。尼古丁调控基因对于烟草商业生产至关重要,目前大多数的尼古丁合成基因相关专利都掌握在国外烟草公司手中。因此,研究尼古丁合成途径相关调控基因对中国烟草企业改良烟草产品中尼古丁含量具有重要意义。
技术实现要素:
本发明的第一目的在于提供一种烟草尼古丁含量调控基因ntclc-b,所述基因编码一种调控烟草尼古丁含量的多肽,所述多肽包含的氨基酸序列如seqid:no.2所示。
本发明的第二目的在于提供所述烟草尼古丁含量调控基因ntclc-b包含的核苷酸序列,所述核苷酸序列如seqid:no.1所示。
本发明的第三目的在于提供一种所述烟草尼古丁含量调控基因ntclc-b的克隆方法,包括以下步骤:
(1)烟草叶片cdna合成:提取烟草rna,反转录得到第一链cdna;
(2)ntclc-b基因的pcr扩增:以烟草叶片cdna为模板,根据ntclc-b基因序列设计引物,进行pcr扩增,回收和纯化pcr扩增产物,并测序。
本发明的第四目的在于提供一种所述烟草尼古丁含量调控基因ntclc-b的应用,即在烟草植株体内过量表达所述ntclc-b基因,可降低烟草中尼古丁的含量。
本发明的第五目的在于提供一种所述烟草尼古丁含量调控基因ntclc-b的重组载体。
本发明的第六目的在于提供一种所述烟草尼古丁含量调控基因ntclc-b的表达盒。
本发明的第七目的在于提供一种所述烟草尼古丁含量调控基因ntclc-b的转基因细胞系。
本发明的第八目的在于提供一种所述烟草尼古丁含量调控基因ntclc-b的重组菌。
附图说明
图1ntclc-b的pcr产物电泳图;
图中,m-分子量标记;1-pcr产物;
图2中间载体pdonr-zeo图;
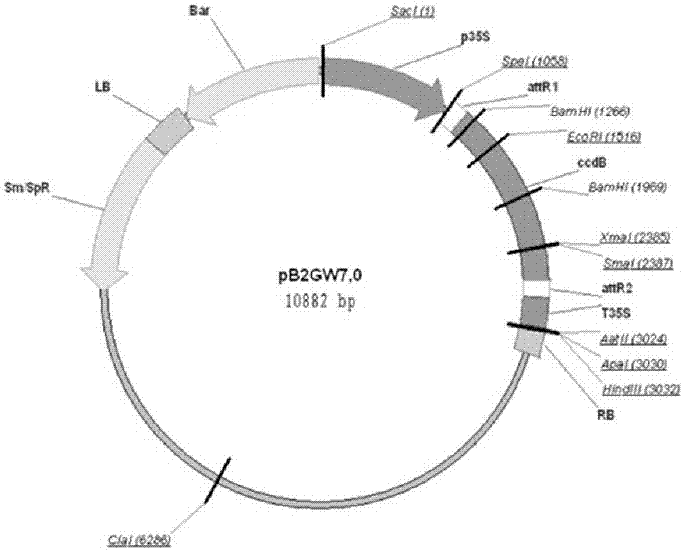
图3ntclc-b基因植物表达载体pb2gw7图;
图4转ntclc-b基因过表达载体的烟草株系基因表达水平柱状图;
图中,oe-1为过表达1株系,oe-2为过表达2株系,k326为野生型烟株对照;
图5ntclc-b基因过表达烟草植株的尼古丁含量。
图中,oe-1为过表达1株系,oe-2为过表达2株系,k326为野生型烟株对照。
具体实施方式
下面结合附图对本发明作进一步的说明,但不以任何方式对本发明加以限制,基于本发明教导所做的任何变换或替换,均属于本发明的保护范围。
本发明所述的烟草尼古丁含量调控基因ntclc-b编码一种调控烟草尼古丁含量的多肽,所述多肽包含的氨基酸序列如seqid:no.2所示。所述多肽还可能为由seqidno:2所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加而形成,且具有调控烟草尼古丁含量的衍生多肽。所述一个或几个氨基酸残基的取代和/或缺失和/或添加是指不超过10个氨基酸残基的取代和/或缺失和/或添加。
本发明所述烟草尼古丁含量调控基因ntclc-b包含的核苷酸序列如seqid:no.1所示。或者在高严谨条件下可与序列表中seqidno:1限定的dna序列杂交的核苷酸序列;或者与序列表中seqidno:1限定的dna序列具有70%以上同源性,且编码相同功能蛋白质的dna序列。
本发明所述烟草尼古丁含量调控基因ntclc-b的克隆方法包括以下步骤:
(1)烟草叶片cdna合成:提取烟草rna,反转录得到第一链cdna;
(2)ntclc-b基因的pcr扩增:以烟草叶片cdna为模板,根据ntclc-b基因序列设计引物,进行pcr扩增,回收和纯化pcr扩增产物并测序。
作为本发明的一种优选,所述引物为:
正向引物:5’-atggaggagccaactcgattagtag-3’;
反向引物:5’-tcagttcccctttttaccgctttttg-3’。
本发明所述烟草尼古丁含量调控基因ntclc-b的应用为在烟草植株体内过量表达所述ntclc-b基因,可降低烟草中尼古丁的含量。可通过多种方法实现过量表达ntclc-b基因,如:植物病毒载体介导基因过表达的方法、农杆菌介导转化过表达载体、优化修改基因编码匡、优化基因启动子等方法。本发明所述的过量表达基因的方法并不限于上述几种方法,只要能过量表达ntclc-b即可。
本发明的ntclc-b基因在构建到植物表达载体中时,在其转录起始核苷酸前可加上任何一种增强启动子或诱导型启动子。为了便于对转基因植物细胞或植物进行鉴定及筛选,可对所使用的载体进行加工,如加入植物可选择性标记(gus基因、荧光素酶基因等)或具有抗性的抗生素标记物(庆大霉素,卡那霉素等)。被转化的植物宿主既可以是单子叶植物,也可以是双子叶植物,如:烟草、水稻、小麦、玉米、黄瓜、番茄、杨树、草坪草或苜蓿等。携带有本发明ntclc-b基因的表达载体可通过使用ti质粒、ri质粒、植物病毒载体、直接dna转化、显微注射、电导、农杆菌介导等常规生物学方法转化植物细胞或组织,并将转化的植物经组织培育成植株。
含有本发明所述ntclc-b基因的重组载体、表达盒、转基因细胞系、和重组菌等基因工程产品均属于本发明的保护范围。
本发明的烟草尼古丁调控相关蛋白及其编码基因为农作物尤其是烟草尼古丁含量育种提供基因与技术的支持。
本发明中,在烟草植株内过表达烟草内源ntclc-b,会使烟草烟叶的尼古丁含量明显降低,在植物尼古丁育种领域具有广阔的应用前景,其经济效益潜力巨大。
下面将结合具体实施例,对本发明进行进一步的说明。
实施例中所有的植物组织材料取自烤烟(nicotianiatabacum)品种‘k326’及过表达ntclc-b基因‘k326’植株。烟草植株的生长和发育阶段都是在人工气候室中,并保持生长温度在22-25℃之间,以尽量减少外界环境因素对烟草尼古丁合成过程中的影响。实验选取的烟草材料为旺长期,发育表型相近的非转基因烟株及转基因烟株。采取非转基因烟株及转基因烟株的上部、中部及下部叶片。对于另一组,对非转基因烟株及转基因烟株进行打顶处理,然后采取非转基因烟株及转基因烟株的上部、中部及下部叶片。将这些烟草材料杀青以进行尼古丁含量检验。
实施例1:克隆ntclc-b基因
以烟草叶片cdna为模板,根据烟草基因组数据库信息设计引物,进行ntclc-b基因的pcr扩增,得到pcr扩增产物。设计引物如下所示:
正向引物:5’-atggaggagccaactcgattagtag-3’;
反向引物:5’-tcagttcccctttttaccgctttttg-3’。
pcr反应体系和扩增条件如表1所示。
表1pcr反应体系与条件
将扩增获得的pcr产物在0.8%的琼脂糖凝胶电泳,凝胶电泳结果如图1所示。电泳结束后,采用qiagen公司pcr产物纯化试剂盒,按照产品说明回收纯化所述的pcr产物,并送invitrogen测序,验证序列结果。
实施例2:植物转化载体构建
以实施例1中ntclc-b全长片段为模板,用含有gateway接头序列的引物进行pcr扩增,扩增产物经pcr产物纯化后,经过bp反应插入到invitrogen公司pdonr-zeo载体中。将构建好的bp反应载体通过lr反应将ntclc-b片段置换到pb2gw7过表达载体中。具体步骤如下:
(1)根据烟草基因组中筛选出的ntclc-b基因序列设计引物,并根据gateway系统中的bp反应要求,在正向引物前加上5’-gggacaagtttgtacaaaaaagcaggctgc-3’序列,在反向引物前加5’-ggggaccactttgtacaagaaagctgggtc-3’序列。得到gateway反应引物序列如下:
ntclc-b_f:
5’-ggggacaagtttgtacaaaaaagcaggctgcatggaggagccaactcgattagtag-3’;
ntclc-b_r:
5’-ggggaccactttgtacaagaaagctgggtctcagttcccctttttaccgctttttg-3’。
(2)pcr反应均采用phusion高保真聚合酶进行pcr克隆。
pcr反应体系与条件与实施例1相同。
(3)bp反应:
(a)在200μl离心管中准备8μl的反应体系,包括:1-7μl的attb-pcr产物(约15~150ng,质量浓度≥10ng/μl)、1μl150ng/μl的pdonr-zeo载体和适量的te缓冲液(ph8.0),在室温下混匀;
(b)将bpclonase™ii酶混合物在冰上静置2min融化,轻轻震荡2次,混匀待用;
(c)向(1)准备的样品中加入2μl的bpclonase™ii酶混合物,轻轻地将体系混匀;
(d)将bpclonase™ii酶混合物放回到-20°c或者-80°c保存;
(e)将反应体系放在25°c温浴1h;
(f)向反应体系中加入1μl的蛋白酶k溶液,轻轻震荡,然后将样品放在37°c温浴10min,以便终止bp反应;
(g)将混合液转化大肠杆菌后,取转化菌液涂在含zeacin抗性的lb平板上,挑取菌落至含相应抗生素培养基溶液中摇菌培养,确认后提取阳性克隆的pdonr-zeocin质粒(图2)备用。
(4)lr反应:
(a)在200μl离心管中准备8μl的反应物,包括:1-7μl的获得的pdonr-zeocin质粒(50-150ng)、1μl150ng/μl的pb2gw7载体和适量的te缓冲液(ph8.0),在室温下混匀;
(b)将lrclonase™ii酶混合物静置在冰上2min融化,轻轻震荡2次以混匀;
(c)加入2μl的lrclonase™ii酶混合物,轻轻震荡将体系混匀;
(d)将lrclonase™ii酶混合物放回到-20℃或者-80℃冰箱保存;
(e)将反应体系放在25℃温浴反应1h;
(f)向反应体系中加入1μl的蛋白酶k溶液以终止lr反应,轻轻震荡后,将样品放在37℃静置10min;得到pb2gw7重组载体。
实施例3:农杆菌介导的烟草转化及转基因植株的鉴定
(1)冻融法转化农杆菌
将1μg(200ng/μl)pb2gw7重组载体加入到100μl感受态农杆菌lba4404中,混匀后在冰上静置5min,放入液氮中冷冻5min,然后从液氮中取出,放入37℃水浴锅中水浴5min,再在冰上静置5min后,加入500μllb溶液,在28℃、充分震荡条件下恢复培养4h,最后将菌液均匀涂抹于选择性平板培养基上,28℃下培养48h。
(2)叶盘法转化烟草品种k326。
具体方法如下:
(a)无菌条件下,将烟草k326种子放入ep管中用无菌水冲洗2-3次;
(b)于75%的酒精中浸泡30-60s;
(c)再用0.1%的升汞处理5min,最后用无菌水冲洗5次;
(d)播种于ms培养基上,培养在云南烟草农业科学研究院组织培养室中,暗培养4d,25℃光照培养20-30d。
(e)待烟草苗长至3-5cm时(20-30d),取顶芽放于ms+ba0.2mg/l(壮芽,使其快速成长)培养基上,继代培养。
(f)继代培养14天后(有小叶片即可),取叶片,大小1cm×1cm,切去叶柄,叶片表面及叶边缘划伤,放入ms+ba1.0mg/lph6.0-6.5的预培养培养基上,正面朝下紧贴培养基放置,于黑暗条件下预培养2-3d。
(g)然后取出预培养的叶片或茎段,放入农杆菌侵染液中进行侵染。侵染前一天晚上,摇菌农杆菌2瓶。将2ml离心管装满菌液,4000r/min离心5min,用悬菌液清洗两次。以1:10比例(10ml悬菌液放1管1.5ml菌体)放入悬菌液,加入as25mg/l(40ml中加40μlas)不断摇晃侵染液,使其与叶片及茎段切口处充分接触,10min后,取出,放于灭过菌的干燥滤纸上吸干菌液。
其中,农杆菌侵染液的制备方法如下:
1)取-80℃冰箱保存的已被转化的农杆菌,划平板培养,lb固体平板中加入50mg/lspec和50mg/lrif;
2)挑取单菌斑到含50mg/lspec和50mg/lrif的5mllb液体培养基中,放入摇床中28℃、200r/min培养过夜(12-16h);
3)保存菌种,750μl菌液加入灭过菌的甘油250μl,-80℃冰箱保存备用。
4)摇菌,lb液体培养基10ml加spec(所需浓度50mg/l)10μl、rif(所需浓度50mg/l)10μl及菌液10μl,28℃、200r/min过夜培养(12-16h)。
5)当菌液浓度达到od600=1.5左右时,取2ml菌液加入到离心管中,4000r/min离心5min;
6)倒掉上清液,吸1ml新的ms液体培养基,重悬农杆菌,4000r/min离心5min。
7)重复步骤(6)1次;
8)用1ml的ms液体培养基重悬菌后,再加入到40ml的ms的液体培养基中(含40μl25mg/l的as),即为侵染液。放置2h以上,再侵染。
200ml悬菌液的配制如下:
20×大量元素10ml
200×有机元素1ml
200×铁盐1ml
200×微量元素1ml
蔗糖5.6g
(h)将叶片及茎段放回到预培养基上,28℃黑暗条件下共培养2-3天,至叶片切口周围有微菌斑形成;
(i)洗菌,取出共培养的烟草叶片及茎段,用添加500mg/lcef的无菌水冲洗5次,第一次放置于摇床摇30min,后面每次5min,以洗去外植体表面的农杆菌;
(j)取出后,用滤纸吸干,转移到烟草诱芽培养基上,诱芽培养基为ms+ba1.0mg/l+hyg25mg/l+cef500mg/lph5.8;过2周观察,如果发现不长菌,则降低cef浓度。如果长菌,则继续保持cef浓度。
(k)每2周更换1次培养基,直至长出不定芽(一般情况为2周)。切下再生的小苗(1cm左右),转入继代培养基ms+ba0.2-0.1mg/l+hyg25mg/l+cef500mg/lph5.8;
(l)至小苗长至2cm长时(有小芽即可),转接入生根培养基ms+naa0.2-0.1mg/l上,24士1℃、12h光照、1500lx培养3周左右,长出粗壮根系。
(m)待根生长至2-3cm。苗高7-10cm左右时,移出三角瓶洗去根部培养基,移栽于花盆中,温室培养。
(3)得到稳定的转基因株系
采用qiagen公司dna提取试剂盒,提取转基因烟草幼苗的基因组dna,设计basta抗性基因引物进行pcr扩增,筛选阳性植株,检测到25株阳性植株。
检测引物为:
basta_f:5’-acaagcacggtcaacttcc-3’;
basta_r:5’-actcggccgtccagtcgta-3’。
提取野生型植株和25株转ntclc-b的t0代植株的总rna,进行实时荧光定量pcr(qrt-pcr)分析,内参基因为26s,分析不同株系的表达情况。选取表达量最高的2株植株oe-1和oe-2(图4)。单株收取种子分别播种,用basta抗生素筛选tl代植株的分离情况,如此重复直至t3代获得遗传稳定的转基因株系。
ntclc-bqrt引物:
ntclc-b_qrt_f:5’-gctcactttgatagctgccc-3’;
ntclc-b_qrt_r:5’-aagcgccaatgtgaaccaat-3’。
26s内参基因引物:
26s_f:5’-gaagaaggtcccaagggttc-3’;
26s_r:5’-tctccctttaacaccaacgg-3’。
实施例4:测定烟叶中尼古丁含量
依据标准yc/t160-2002检测中烟草材料的尼古丁含量。选取的烟草材料为旺长期,发育表型相近的非转基因烟株及转基因烟株为处理对象,以野生型烟草k326为对照。取5株非转基因烟株及转基因烟株的上部、中部及下部叶片。对于另一组,对5株非转基因烟株及转基因烟株进行打顶处理,然后采取非转基因烟株及转基因烟株的上部、中部及下部叶片。
用5%乙酸水溶液萃取烟草样品,萃取液中的总植物碱(以尼古丁计)与对氨基苯磺酸和氯化氰反应,氯化氰由氰化钾和氯胺t在线反应产生。反应产物用比色计在460nm测定。
主要仪器设备:连续流动分析仪(美国api)(德国sealaa3)(法国alliance)。
配置试剂:
brij35溶液(聚乙氧基月桂醚):水中加5滴22%brij35,搅拌均匀。
缓冲溶液a:称取2.35g氯化钠(nacl),7.60g硼酸钠(na2b4o3·10h2o),用水溶解,然后转入1l容量瓶中,加入1mlbrij35,用蒸馏水稀释至1l。使用前用定性滤纸过滤。
缓冲溶液b:称26g磷酸氢二钠(na2hpo4),10.4g柠檬酸[coh(cooh)(ch2cooh)2·h2o],7g对氨基苯磺酸(nh2c6h4so3h),用水溶解,然后转入1l容量瓶中,加入1mlbrij35,用蒸馏水稀释至1l。使用前用定性滤纸过滤。
氯胺t溶液(n-氯-4-甲基苯硫酰胺钠盐)[ch3c6h4so2n(na)cl·3h2o]:取8.65g氯胺t,溶于水中,然后转入500ml的容量瓶中,用水定容至刻度。使用前用定性滤纸过滤。
0.22mol/lnaoh缓冲液:naoh8.8g,na2hpo426.0g,c6h8o7•h2o(一水柠檬酸)10.4g,用水溶解并定容至1000ml。
对氨基苯磺酸缓冲液:称取c6h7no3s(对氨基苯磺酸)7g,na2hpo426.0g,c6h8o7•h2o(一水柠檬酸)10.4g,用水溶解并定容至1000ml。
氯胺t:称取氯胺t1.2g,用纯水溶解定容至100ml,用棕色试剂瓶保存。
氰化钾:kcn0.4g,用纯水溶解定容至100ml。
naco3溶液:10gnaco3,蒸馏水溶解并定容至1000ml。
分析步骤:称取0.3g烟样于150ml三角瓶或塑料瓶中(精确至0.0001g);加入50ml5%乙酸溶液盖上塞子;在普通摇床上振荡萃取30min,转速控制170r/min,用滤纸过滤上机。(如样品液浓度超出工作标准液的浓度范围,则应稀释)。
结果的计算与表述:
以干基计的总植物碱的含量,由下面公式得出:
式中:
c——样品液总植物碱的仪器观测值,单位为mg/ml;
v——萃取液的体积,单位为ml;
m——试料的质量,单位为mg;
w——试样的水分含量,单位为%。
以两次测定的平均值作为测定结果,结果精确至0.01%。
试验结果表明:oe-1的尼古丁含量约为对照的一半,而oe-2的尼古丁含量约为对照的25%(图5)。打顶与未打顶烟株的上中下部烟叶尼古丁含量都表现出明显的降低,说明过表达ntclc-b基因能够有效降低烟叶中的尼古丁含量。
序列表
<110>云南省烟草农业科学研究院
<120>一种烟草尼古丁含量调控基因ntclc-b及其克隆方法与应用
<130>2017
<160>2
<170>siposequencelisting1.0
<210>1
<211>2361
<212>dna
<213>nicotianasp.
<400>1
atggaggagccaactcgattagtagaagaagcaacgattaataacatggacggacaacag60
aatgaagaagaaagagatccagagagcaattcactgcatcagcctcttctcaagagaaac120
agaacactatcatccagtccatttgccttggttggagctaaggtctcccacatcgaaagt180
ttggattatgagatcaacgagaatgatctcttcaagcatgactggagaaggagatctaga240
gttcaagtattacagtatgtgttcttgaaatggacactggcatttttggtcggcctgctt300
acaggagttacagccaccctcatcaatcttgcaatcgaaaacatggctggttacaaactt360
cgagctgttgtgaactatatcgaggatagaaggtaccttatgggatttgcatattttgcg420
ggtgctaattttgtgctcactttgatagctgcccttctctgcgtgtgctttgcacctact480
gctgcagggcctggaattcctgaaatcaaagcttatctcaacggtgtagatactcccaat540
atgtatggagcaaccacactttttgtcaagatcattggaagcattgcagcagtttctgct600
agcttagaccttggaaaagaagggccattggttcacattggcgcttgctttgcttcctta660
ctaggtcaaggtggtccagataattaccggctcaggtggcgttggctccgttacttcaac720
aacgatcgggacaggcgagatcttatcacatgtgggtcatcatcaggtgtgtgtgctgct780
ttccgttctccagtaggtggtgtcctatttgctttagaggaagtggcaacatggtggaga840
agtgcactcctctggagaactttcttcagcacggcagttgtggtggtgatactgagggcc900
ttcattgaatactgcaaatctggcaactgtggactttttggaagaggagggcttatcatg960
tttgatgtgagtggtgtcagtgttagctaccatgttgtggacatcatccctgttgtagtg1020
attggaatcataggcggacttttgggaagcctctacaatcatgtcctccacaaaattctg1080
aggctctacaatctgatcaacgagaagggaaaactacataaggttcttctcgctctgagt1140
gtctcccttttcacctccatttgcatgtatggacttccttttttggccaaatgcaagcct1200
tgtgatccatcacttcccgggtcttgtcctggtactggagggacaggaaacttcaagcag1260
ttcaactgcccagacggctattacaatgatcttgctactcttctccttacaaccaacgat1320
gatgcagtccgaaacattttctccataaacactcccggtgaattccaagttatgtctctt1380
attatctacttcgttctgtattgcatattgggactcatcacttttgggattgctgtgcca1440
tctggtctcttccttccaatcatcctcatgggttcagcttatggtcgcttgcttgccatt1500
gccatgggatcttatacaaaaattgatccagggctgtatgcggttctcggagcagcttcc1560
cttatggctggttcaatgagaatgactgtttctctttgcgtcatatttcttgagctaaca1620
aacaatcttctccttctgccaataacaatgctggttcttctaattgccaaaagtgtagga1680
gactgcttcaacctaagtatttatgaaataatattggagctgaaaggtctacctttcctg1740
gatgccaacccggagccatggatgagaaatatcactgctggtgagcttgctgatgtaaag1800
ccaccagtagttacactctgtggagttgagaaggtgggacgtatcgtagaggccttgaag1860
aacaccacatataacggattccctgtcgtcgatgaaggagtagtgccaccggtgggtctg1920
ccagttggggcaactgaattgcacggtcttgtcctaagaactcaccttcttttggttctc1980
aagaaaaagtggttccttcatgaaagacggaggacagaggagtgggaagtgagagagaaa2040
ttcacctggattgatttagctgagaggggcggtaagatcgaagatgtgttagttacaaag2100
gatgaaatggagatgtatgtcgatttgcatcccctgactaacacaaccccttatactgtg2160
gtagaaagcttgtcagtggctaaggcaatggtgcttttcaggcaggtggggctccgccac2220
atgctcattgtacccaaataccaagcagcaggggtatctcctgtggtgggaatcttgacc2280
aggcaagacttgagagcccacaacattttgagtgtcttccctcatctggagaagtcaaaa2340
agcggtaaaaaggggaactga2361
<210>2
<211>786
<212>prt
<213>nicotianasp.
<400>2
metglugluprothrargleuvalgluglualathrileasnasnmet
151015
aspglyglnglnasngluglugluargaspprogluserasnserleu
202530
hisglnproleuleulysargasnargthrleuserserserprophe
354045
alaleuvalglyalalysvalserhisilegluserleuasptyrglu
505560
ileasngluasnaspleuphelyshisasptrpargargargserarg
65707580
valglnvalleuglntyrvalpheleulystrpthrleualapheleu
859095
valglyleuleuthrglyvalthralathrleuileasnleualaile
100105110
gluasnmetalaglytyrlysleuargalavalvalasntyrileglu
115120125
aspargargtyrleumetglyphealatyrphealaglyalaasnphe
130135140
valleuthrleuilealaalaleuleucysvalcysphealaprothr
145150155160
alaalaglyproglyileprogluilelysalatyrleuasnglyval
165170175
aspthrproasnmettyrglyalathrthrleuphevallysileile
180185190
glyserilealaalavalseralaserleuaspleuglylysglugly
195200205
proleuvalhisileglyalacysphealaserleuleuglyglngly
210215220
glyproaspasntyrargleuargtrpargtrpleuargtyrpheasn
225230235240
asnaspargaspargargaspleuilethrcysglysersersergly
245250255
valcysalaalapheargserprovalglyglyvalleuphealaleu
260265270
glugluvalalathrtrptrpargseralaleuleutrpargthrphe
275280285
pheserthralavalvalvalvalileleuargalapheileglutyr
290295300
cyslysserglyasncysglyleupheglyargglyglyleuilemet
305310315320
pheaspvalserglyvalservalsertyrhisvalvalaspileile
325330335
provalvalvalileglyileileglyglyleuleuglyserleutyr
340345350
asnhisvalleuhislysileleuargleutyrasnleuileasnglu
355360365
lysglylysleuhislysvalleuleualaleuservalserleuphe
370375380
thrserilecysmettyrglyleupropheleualalyscyslyspro
385390395400
cysaspproserleuproglysercysproglythrglyglythrgly
405410415
asnphelysglnpheasncysproaspglytyrtyrasnaspleuala
420425430
thrleuleuleuthrthrasnaspaspalavalargasnilepheser
435440445
ileasnthrproglyglupheglnvalmetserleuileiletyrphe
450455460
valleutyrcysileleuglyleuilethrpheglyilealavalpro
465470475480
serglyleupheleuproileileleumetglyseralatyrglyarg
485490495
leuleualailealametglysertyrthrlysileaspproglyleu
500505510
tyralavalleuglyalaalaserleumetalaglysermetargmet
515520525
thrvalserleucysvalilepheleugluleuthrasnasnleuleu
530535540
leuleuproilethrmetleuvalleuleuilealalysservalgly
545550555560
aspcyspheasnleuseriletyrgluileileleugluleulysgly
565570575
leupropheleuaspalaasnprogluprotrpmetargasnilethr
580585590
alaglygluleualaaspvallysproprovalvalthrleucysgly
595600605
valglulysvalglyargilevalglualaleulysasnthrthrtyr
610615620
asnglypheprovalvalaspgluglyvalvalproprovalglyleu
625630635640
provalglyalathrgluleuhisglyleuvalleuargthrhisleu
645650655
leuleuvalleulyslyslystrppheleuhisgluargargargthr
660665670
gluglutrpgluvalargglulysphethrtrpileaspleualaglu
675680685
argglyglylysilegluaspvalleuvalthrlysaspglumetglu
690695700
mettyrvalaspleuhisproleuthrasnthrthrprotyrthrval
705710715720
valgluserleuservalalalysalametvalleupheargglnval
725730735
glyleuarghismetleuilevalprolystyrglnalaalaglyval
740745750
serprovalvalglyileleuthrargglnaspleuargalahisasn
755760765
ileleuservalpheprohisleuglulysserlysserglylyslys
770775780
glyasn
785
- 还没有人留言评论。精彩留言会获得点赞!