一种基于TDR基因的转基因水稻不育系的培育方法与流程
本发明涉及分子生物学核酸化学技术领域,特别涉及一种水稻遗传工程不育系的制备方法。
背景技术
水稻是是世界上近一半的人口的主食,除可食用外,还可以酿酒、制糖和作为工业原料,人类对水稻的需求很大。多年来的生产实践表明,杂交水稻一般可比常规稻增产20%以上,因此杂交水稻彰显着巨大的增产潜力。
杂交水稻的发展依赖于杂交水稻不育系的培育。我国杂交水稻的研究始于上个世纪60年代,70年代开始大规模被种植。第一代杂交水稻是以核质互作雄性不育系为遗传工具的三系法,第二代杂交水稻是以光温敏雄性不育系为遗传工具的两系法,“三系法”和“两系法”杂交育种技术对粮食增产贡献巨大,但也存在问题。三系法中可利用的具有优良形状的父母本有限,配组不自由,野败保持系频率较低。
生产上现有的不育系间遗传差异小,杂交种育性稳定性不够,抵御逆境能力较差。而“两系法”中光温敏雄性不育系的育性受外界温度控制,易导致光温敏不育系自交结实,制种失败。因此开发新一代不育系对杂交水稻的发展至关重要。
技术实现要素:
本发明所要解决的技术问题在于,提供了一种基于tdr基因的转基因水稻不育系的培育方法。本发明将水稻普通核不育基因tdr与小麦花粉致死基因ki连锁,同时以橙色荧光蛋白tagrfp基因作为报告基因,对水稻tdr突变体进行基因改造,获得f1代杂合体。f1代杂合体在自交结实过程中遵循孟德尔分离定律,产生的后代既有能保持三连锁基因的杂合体,又有无育性的不育系。
为解决上述技术问题,本发明提供了一种基于tdr基因的转基因水稻不育系的培育方法,包括以下步骤:
a、tagrfp基因表达盒的获得;
b、小麦花粉致死基因ki的扩增;
c、水稻tdr盒的获得;
d、花粉特异性启动子pg47的扩增;
e、各基因表达元件的连接。
所述步骤c、水稻tdr盒的获得,进一步的包括:
提取水稻植株的基因组dna,以基因组dna为模板,以f2/r2为上下游引物进行完整tdr基因表达盒的扩增,扩增过程中分别在上游引物和下游引物中引入smai和nael酶切位点,同时在nael后方添加drai酶切位点,其扩增的水稻tdr基因包含其上游的启动子和下游的终止子,同时包含所有的外显子和内含子。
所述步骤a、tagrfp基因表达盒的获得,进一步包括:
以含有tagrfp基因完整表达盒的质粒为模板,以f1/ri为上下游引物进行扩增,扩增过程中分别在上下游引物中引入aaui和kpni位点。
所述步骤b、小麦花粉致死基因ki的扩增,进一步包括:
以水稻基因组dna为模板,以f3/r3为上下游引物进行小麦花粉致死基因ki的扩增,在上下游引物中引入drai酶切位点;其扩增的致死基因ki包含所有的外显子和内含子。
所述步骤d、花粉特异性启动子pg47的扩增,进一步包括:
以水稻基因组dna为模板,以f4/r4为上下游引物进行花粉特异性启动pg47扩增,在上下游引物中分别引入kpni和smai酶切位点。
所述步骤e、各基因表达元件的连接,进一步包括:
第一步,将tagrfp基因表达元件连接到pcambia1300载体上;
第二步,将完整的水稻tdr基因引入第一步中已经引入tagrfp基因表达元件的pcambia1390载体上;
第三步,将小麦花粉致死基因ki连入到已经连有tagrfp基因表达元件和水稻tdr基因的pcambia1390双元载体上;
第四步,将水稻花粉特异性启动pg47连接到前三步已经连接上tagrfp基因表达元件、水稻tdr基因和小麦花粉致死基因ki的复合载体上。
为解决上述技术问题,本发明又提供了一种水稻普通核不育tdr突变体的利用方法,应用水稻tdr基因表达盒构建如前述任一项方法制备的水稻不育系。
为解决上述技术问题,本发明另提供了一种如前述任一项方法在水稻遗传育种中的应用。
本发明有益效果包括:本发明将水稻普通核不育基因tdr与小麦花粉致死基因ki连锁,同时以橙色荧光蛋白tagrfp基因作为报告基因,对水稻tdr突变体进行基因改造,获得f1代杂合体。f1代杂合体在自交结实过程中遵循孟德尔分离定律,产生的后代既有能保持三连锁基因的杂合体,又有无育性的不育系。
附图说明
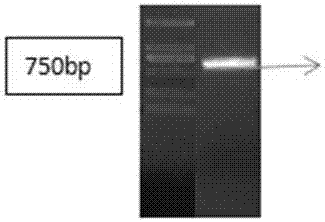
图1:tagrfp基因完整表达盒pcr扩增凝胶电泳检测图;
图2:水稻tdr基因表达盒pcr扩增凝胶电泳检测图;
图3:小麦花粉致死基因ki的扩增凝胶电泳检测图;
图4:花粉特异性启动子pg47的扩增凝胶电泳检测图;
图5:植物表达载体基因连锁、转录方向和酶切位点图谱
图6:转基因植株结出的稻穗照片。
具体实施方式
下面结合实施例详述本发明。为使本发明的目的、技术方案及优点更加清楚、明确,以下参照附图并举实施例对本发明进一步详细说明,但本发明并不局限于这些实施例。
本发明是将水稻普通核不育基因tdr与小麦花粉致死基因ki连锁,同时以橙色荧光蛋白tagrfp基因作为报告基因,对水稻tdr突变体进行基因改造,获得f1代杂合体。f1代杂合体在自交结实过程中遵循孟德尔分离定律,产生的后代既有能保持三连锁基因的杂合体,又有无育性的不育系,即所谓的第三代不育系。
为解决上述技术问题,本发明提供了一种基于tdr基因的转基因水稻不育系的培育方法,包括以下步骤:
a、tagrfp基因表达盒的获得;
b、小麦花粉致死基因ki的扩增;
c、水稻tdr盒的获得;
d、花粉特异性启动子pg47的扩增;
e、各基因表达元件的连接。
所述步骤c、水稻tdr盒的获得,进一步的包括:
提取水稻植株的基因组dna,以基因组dna为模板,以f2/r2为上下游引物进行完整tdr基因表达盒的扩增,扩增过程中分别在上游引物和下游引物中引入smai和nael酶切位点,同时在nael后方添加drai酶切位点,其扩增的水稻tdr基因包含其上游的启动子和下游的终止子,同时包含所有的外显子和内含子。
所述步骤a、tagrfp基因表达盒的获得,进一步包括:
以含有tagrfp基因完整表达盒的质粒为模板,以f1/r1为上下游引物进行扩增,扩增过程中分别在上下游引物中引入aaui和kpni位点。
所述步骤b、小麦花粉致死基因ki的扩增,进一步包括:
以水稻基因组dna为模板,以f3/r3为上下游引物进行小麦花粉致死基因ki的扩增,在上下游引物中引入drai酶切位点;其扩增的致死基因ki包含所有的外显子和内含子。
所述步骤d、花粉特异性启动子pg47的扩增,进一步包括:
以水稻基因组dna为模板,以f4/r4为上下游引物进行花粉特异性启动pg47扩增,在上下游引物中分别引入kpni和smai酶切位点。
所述步骤e、各基因表达元件的连接,进一步包括:
第一步,将tagrfp基因表达元件连接到pcambia1300载体上;
第二步,将完整的水稻tdr基因引入第一步中已经引入tagrfp基因表达元件的pcambia1390载体上;
第三步,将小麦花粉致死基因ki连入到已经连有tagrfp基因表达元件和水稻tdr基因的pcambia1390双元载体上;
第四步,将水稻花粉特异性启动pg47连接到前三步已经连接上tagrfp基因表达元件、水稻tdr基因和小麦花粉致死基因ki的复合载体上。
为解决上述技术问题,本发明又提供了一种水稻普通核不育tdr突变体的利用方法,应用水稻tdr基因表达盒构建如前述任一项方法制备的水稻不育系。
为解决上述技术问题,本发明另提供了一种如前述任一项方法在水稻遗传育种中的应用。
1、水稻tdr基因表达盒的获得
提取水稻武运粳7号植株的基因组dna,以基因组dna为模板,以f2/r2为上下游引物进行完整tdr基因表达盒的扩增,扩增过程中分别在上游引物和下游引物中引入smai和nael酶切位点,同时在nael后方添加drai酶切位点,其扩增的水稻tdr基因包含其上游的启动子和下游的终止子,同时包含所有的外显子和内含子。
2、tagrfp基因表达盒的获得
以实验室保存的含有tagrfp基因完整表达盒的质粒为模板,以f1/r1为上下游引物进行扩增,扩增过程中分别在上下游引物中引入aaui和kpni位点。表达盒总长度为696bp,通过pcr扩增和琼脂糖凝胶电泳检测,初步获得了tagrfp基因完整表达盒。
3、小麦花粉致死基因ki的扩增
以水稻基因组dna为模板,以f3/r3为上下游引物进行小麦花粉致死基因ki的扩增,在上下游引物中引入drai酶切位点。其扩增的致死基因ki包含所有的外显子和内含子。
4、花粉特异性启动子pg47的扩增
以水稻基因组dna为模板,以f4/r4为上下游引物进行花粉特异性启动pg47扩增,在上下游引物中分别引入kpni和smai酶切位点。
5、各基因表达元件的连接
第一步,利用上下游引物中引入的aaui和kpn1酶切位点和pcambia1390双元载体上aaui和kpn1酶切位点,将tagrfp基因表达元件连接到pcambia1390载体上。第二步,连接tdr基因,利用上下游引物中引入的smal和nael酶切位点和pcambia1390双元载体上的smal和nael酶切位点,将完整的水稻tdr基因引入第一步中已经引入tagrfp基因表达元件的pcambia1390载体上,同时在nael后方添加drai酶切位点。第三步,利用上下游都引入的drai酶切位点,将小麦花粉致死基因ki通过单酶切位点连入到已经连有tagrfp基因表达元件和水稻tdr基因的pcambia1390双元载体上。最后一步,利用上下游引物中引入kpni和smai酶切位点和pcambia1390双元载体上的kpni和smai酶切位点,将水稻花粉特异性启动pg47连接到前三步已经连接上tagrfp基因表达元件、水稻tdr基因和小麦花粉致死基因ki的复合载体上。
表1:各基因扩增到的引物、引物中添加的酶切位点
以上所述,仅是本发明的几个实施例,并非对本发明做任何形式的限制,虽然本发明以较佳实施例揭示如上,然而并非用以限制本发明,任何熟悉本专业的技术人员,在不脱离本发明技术方案的范围内,利用上述揭示的技术内容做出些许的变动或修饰均等同于等效实施案例,均属于本发明技术方案保护范围内。
- 还没有人留言评论。精彩留言会获得点赞!