抗体及其制造方法、以及提升抗体的热稳定性的方法与流程
本发明涉及抗体及其制造方法。另外,本发明涉及提升抗体的热稳定性的方法。
背景技术:
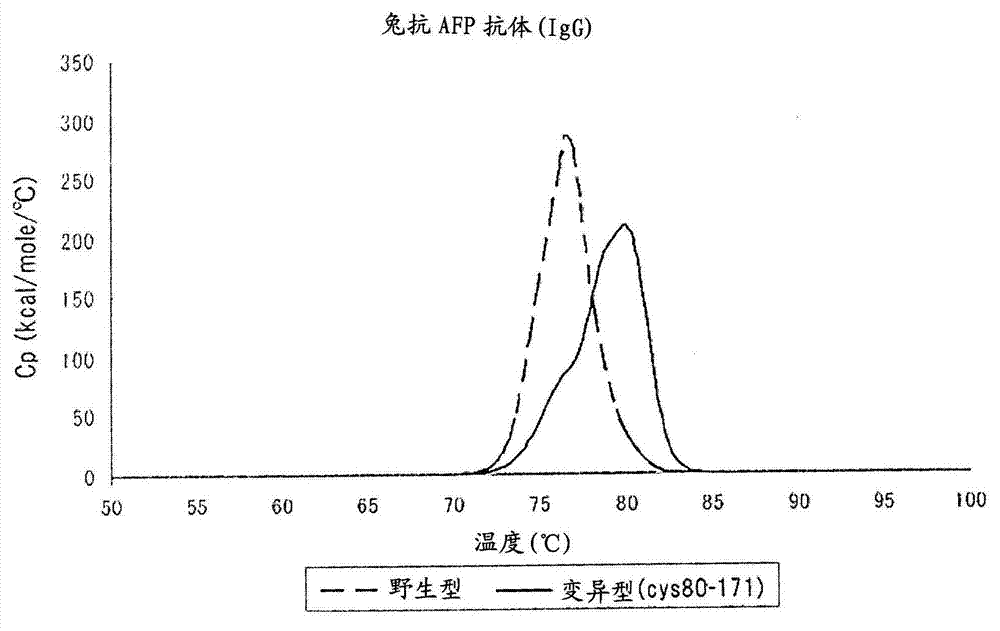
:以往已知通过向抗体的氨基酸序列导入变异而使该抗体对于抗原的亲和性改变的技术。但是,即使由变异的导入而对于抗原的亲和性如期望被改变,也有抗体的热稳定性同时降低的情况。由于抗体的热稳定性与抗体的保存稳定性或耐凝集性相关,从而作为抗体医药开发中的指标之一使用。抗体的热稳定性多受抗体的分子结构的影响。关于抗体的分子结构,在例如非专利文献1中,对兔igg抗体的抗原结合性片段(fab)进行结晶化解析而知悉兔抗体的轻链有特有的二硫键。更具体而言,在非专利文献1中记载了在兔抗体的轻链中的基于kabat法的第80及第171位存在不存在于人抗体及小鼠抗体中的2个半胱氨酸残基,上述的兔抗体在这些半胱氨酸残基间形成特有的二硫键。已知二硫键一般影响蛋白质分子的结构。但是,非专利文献1公开了兔抗体的fab和人-兔的嵌合抗体的fab结构极其类似,兔抗体的fab的整体结构及稳定性与嵌合抗体的fab无异。如上所述,由于在人抗体的轻链中的基于kabat法的第80及第171位不存在半胱氨酸残基,在非专利文献1的人-兔的嵌合抗体中不存在兔抗体特有的二硫键。即,非专利文献1提示,兔抗体特有的二硫键不贡献于抗体的结构及稳定性。【现有技术文献】【非专利文献】【非专利文献1】araih.等,crystalstructureofaconformation-dependentrabbitiggfabspecificforamyloidprefibrillaroligomers,biochimbiophysacta.vol.1820,p.1908-1914,2012【发明的概要】【发明要解决的课题】如上所述,由于抗体的热稳定性如保存稳定性及耐凝集性一样,是关于抗体的品质的要素,期望提升抗体的热稳定性的技术的确立。一般而言,由于蛋白质分子内的二硫键有热稳定性相关的可能性,本发明人关注于兔抗体特有的二硫键。进而,对各种兔抗体的热稳定性进行实际测定的结果,令人惊讶地发现,与非专利文献1的提示相反,兔抗体特有的二硫键贡献于热稳定性的提升。还发现,在兔抗体之中也有轻链的第80及第171位无半胱氨酸残基的抗体,及将所述抗体的轻链的第80及第171个氨基酸残基取代为半胱氨酸,则热稳定性提升。还发现,来源于兔以外的哺乳动物的抗体也通过进行同样的取代而热稳定性提升。【用于解决课题的手段】从而,本发明提供一种抗体,其特征在于,在基于kabat法的可变区的第80个氨基酸残基和基于kabat法的恒定区的第171(在eu指数的第171)的氨基酸残基不是半胱氨酸的抗体中,该可变区的第80个氨基酸残基和该恒定区的第171个氨基酸残基被取代为半胱氨酸。另外,本发明提供一种抗体的制造方法,其包括:对于基于kabat法的可变区的第80个氨基酸残基和基于kabat法的恒定区的第171个氨基酸残基不是半胱氨酸的抗体,将该可变区的第80个氨基酸残基和该恒定区的第171个氨基酸残基取代为半胱氨酸的工序,及回收在该取代工序中得到的抗体的工序。再者,本发明提供提升抗体的热稳定性的方法,其特征在于,在基于kabat法的可变区的第80个氨基酸残基和基于kabat法的恒定区的第171个氨基酸残基不是半胱氨酸的抗体中,将该可变区的第80个氨基酸残基和该恒定区的第171个氨基酸残基取代为半胱氨酸。【发明的效果】本发明提供与进行半胱氨酸取代之前的原来的抗体比,热稳定性提升的抗体。【附图说明】【图1】是显示将兔抗α胎蛋白(afp)抗体的野生型及变异型(cys80-171)的热稳定性用示差扫描热量计(dsc)测定之时的解析峰的坐标图。“变异型(cys80-171)”表示在野生型的抗体中的可变区的第80个氨基酸残基及恒定区的第171个氨基酸残基被取代为半胱氨酸的抗体。可变区及恒定区的氨基酸残基的编号基于kabat法。【图2】是显示将人抗人上皮生长因子受体2类型(her2)抗体(fab)的野生型及其变异型(cys80-171)的热稳定性用dsc测定之时的解析峰的坐标图。【图3a】是显示将小鼠抗胰岛素抗体的野生型及变异型(cys80-171)的热稳定性用dsc测定之时的解析峰的坐标图。【图3b】是显示将小鼠抗胰岛素抗体(fab)的野生型及变异型(cys80-171)的热稳定性用dsc测定之时的解析峰的坐标图。【图3c】是显示将小鼠抗胰岛素抗体(fab)的野生型及变异型(cys108-171)的热稳定性用dsc测定之时的解析峰的坐标图。“变异型(cys108-171)”表示在野生型的抗体中的可变区的第108个氨基酸残基及恒定区的第171个氨基酸残基被取代为半胱氨酸的抗体。【图4】是显示将兔抗afp抗体的野生型及变异型(cys80-171)对抗原的亲和性用elisa法分析的结果的坐标图。【图5】是显示将人抗her2抗体(fab)的野生型及变异型(cys80-171)对抗原的亲和性用elisa法分析的结果的坐标图。【图6】是显示将小鼠抗胰岛素抗体的野生型及变异型(cys80-171)对抗原的亲和性用elisa法分析的结果的坐标图。【具体实施方式】[1.热稳定性提升的抗体]本实施方式的抗体的特征在于,在基于kabat法的可变区的第80个氨基酸残基和基于kabat法的恒定区的第171个氨基酸残基不是半胱氨酸的抗体中,该可变区的第80个氨基酸残基和该恒定区的第171个氨基酸残基被取代为半胱氨酸。这样的氨基酸残基的取代的结果,抗体的热稳定性与进行取代之前的原来的抗体相比提升。即,本实施方式的抗体是以热稳定性提升的方式人工改变的抗体(以下,也称为“改变抗体”)。在本说明书中,“基于kabat法”是指抗体的可变区的氨基酸残基的编号根据由kabat等的编号方案(参照kabate.a.等,sequencesofproteinsofimmunologicalinterest,5thed.publichealthservice,nationalinstitutesofhealth,bethesda,md(1991),nihpublicationno.91-3242),抗体的恒定区的氨基酸残基的编号根据由kabat等的eu指数。eu指数在由kabat等的上述文献中记载。以下,将基于kabat法的可变区的第80个氨基酸残基和基于kabat法的恒定区的第171个氨基酸残基不是半胱氨酸的抗体也称为“原来的抗体”。另外,以下将基于kabat法的可变区的第80个氨基酸残基也简称为“第80个氨基酸残基”,将基于kabat法的恒定区的第171个氨基酸残基也简称为“第171个氨基酸残基”。在原来的抗体中的第80及第171个氨基酸残基只要是半胱氨酸以外的残基,就也可为任何氨基酸残基。基于kabat法的可变区的第80个氨基酸残基、及基于kabat法的恒定区的第171个氨基酸残基存在于轻链及重链的两方。在本实施方式中,原来的抗体只要是在轻链及重链之任一方或两方中的上述的位置的氨基酸残基不是半胱氨酸的抗体即可。在优选的实施方式中,原来的抗体是在轻链中的第80个氨基酸残基、及在该轻链中的第171个氨基酸残基不是半胱氨酸的抗体。当将轻链中的第80及第171个氨基酸残基取代为半胱氨酸时,认为在这些半胱氨酸残基间、即轻链的可变区和该轻链的恒定区之间形成二硫键。此二硫键相当于上述的兔抗体特有的二硫键。原来的抗体也可为识别任何抗原的抗体。另外,原来的抗体只要是第80及第171个氨基酸残基不是半胱氨酸,就可为天然的抗体,也可为人工制出的抗体。作为人工制出的抗体,例如,可举出向天然的抗体的氨基酸序列导入变异(氨基酸残基的取代、附加或删除)的抗体、嵌合抗体、人源化抗体等。原来的抗体只要是第80及第171个氨基酸残基不是半胱氨酸,就也可为来源于任何动物的抗体。作为这样的动物,优选哺乳动物,例如可举出人、小鼠、兔、大鼠、猪、绵羊、山羊、骆驼、牛、马等。在它们之中也优选人、小鼠及兔。本实施方式的改变抗体与原来的抗体来源于相同的动物。在优选的实施方式中,原来的抗体是其基因的碱基序列公知的或能确认该碱基序列的抗体。具体而言,是在公知的数据库中公开了抗体基因的碱基序列的抗体,或可得到产生该抗体的杂交瘤的抗体。作为这样的数据库,可举出例如由nationalcenterforbiotechnologyinformation(ncbi)提供的genebank等。当有产生原来的抗体的杂交瘤时,抗体基因的碱基序列可通过由公知的方法,从该杂交瘤取得抗体基因,对其碱基序列进行测序而得到。本实施方式的改变抗体与原来的抗体比较而热稳定性提升。抗体的热稳定性一般可由测定伴随热应激而变性的抗体的量或比率的方法评价。这样的测定方法本身是在现有技术中公知的,例如,可举出利用示差扫描热量计(dsc)、cd光谱、荧光光谱、傅里叶变换红外分光光度计(ftir)等的测定。在本实施方式中,抗体的热稳定性优选由从利用dsc的测定得到的信息评价。作为这样的信息,例如可为tm(热容量是最大值之时的温度),也可为解析峰本身。在本实施方式中,用dsc测定的改变抗体的tm值比原来的抗体的tm值高。例如,用dsc测定的改变抗体的tm值与原来的抗体的tm值比而高至少约1℃,优选高至少约2℃,更优选高至少约3℃。在原来的抗体中的第80及第171个氨基酸残基向半胱氨酸的取代几乎不影响对于抗原的亲和性。从而,本实施方式的改变抗体与原来的抗体结合于相同的抗原,对于该抗原的亲和性也与原来的抗体是同程度。抗体对于抗原的亲和性可由elisa法等的免疫学测定法评价,也可由抗原抗体反应中的动力学参数(结合速度常数、解离速度常数及解离常数)评价。动力学参数可由表面等离子体共振(spr)技术取得。在本实施方式中,原来的抗体及改变抗体的类可为igg、iga、igm、igd及ige之任一者,但优选为igg。igg的亚类不特别限定,也可为igg1、igg2、igg3及igg4之任一者。在本实施方式中,原来的抗体及改变抗体的轻链的种类优选为κ链。在本实施方式中,原来的抗体只要是有进行了被半胱氨酸的取代的第80及第171个氨基酸残基,就也可处于抗体片段的形态。另外,改变抗体只要是含取代为半胱氨酸的第80及第171位,就也可处于抗体片段的形态。作为这样的抗体片段,可举出例如fab、f(ab’)2、fab’、fd等。在它们之中,也特别优选fab。本实施方式的改变抗体的使用法与原来的抗体比无特别变化。改变抗体可与原来的抗体同样地在各种试验及研究、或者抗体医药等中利用。另外,本实施方式的改变抗体也可被在现有技术中公知的标记物质等修饰。[2.抗体的制造方法]由本实施方式的抗体的制造方法(以下,也简称为“制造方法”)可得到上述的本实施方式的改变抗体。在本实施方式的制造方法中,首先,对于基于kabat法的可变区的第80个氨基酸残基和基于kabat法的恒定区的第171个氨基酸残基不是半胱氨酸的抗体,将该可变区的第80个氨基酸残基和该恒定区的第171个氨基酸残基取代为半胱氨酸。在本实施方式的制造方法中,基于kabat法的可变区的第80个氨基酸残基和基于kabat法的恒定区的第171个氨基酸残基不是半胱氨酸的抗体与上述的“原来的抗体”相同。在原来的抗体中的第80及第171个氨基酸残基向半胱氨酸的取代可由dna重组技术及其他分子生物学技术等公知的方法进行。例如,当有产生原来的抗体的杂交瘤时,如后述的实施例1所示,使用从该杂交瘤提取的rna,由逆转录反应及race(cdna末端的快速扩增)法,各自合成编码轻链的多核苷酸及编码重链的多核苷酸。例如,当将原来的抗体的轻链中的第80及第171个氨基酸残基取代为半胱氨酸时,通过将编码轻链的多核苷酸使用用于将第80及第171个氨基酸残基取代为半胱氨酸的引物而由pcr法扩增,得到了编码在第80及第171位有半胱氨酸的轻链的多核苷酸。通过将得到的多核苷酸与编码原来的抗体的重链的多核苷酸一同整合到在现有技术中公知的表达用载体,得到了含编码本实施方式的改变抗体的多核苷酸的表达载体。编码轻链的多核苷酸及编码重链的多核苷酸可整合到1个表达载体,也可分别整合到2个表达载体。表达载体的种类不特别限定,可为哺乳动物细胞用表达载体,也可为大肠杆菌用表达载体。通过将得到的表达载体转导或转染到适当的宿主细胞(例如,哺乳动物细胞或大肠杆菌)中,可得到改变抗体。当无产生原来的抗体的杂交瘤时,由例如kohler及milstein,nature,vol.256,p.495-497,1975中记载的方法等的公知的方法制作产生抗体的杂交瘤即可。或者,也可使用从用指定的抗原免疫的小鼠或兔等的动物的末梢血或脾脏取得的rna。当使用从末梢血或脾脏取得的rna时,如后述的参考例所示,也可从该rna合成cdna,从得到的cdna制作fab噬菌体库。可使用此库,由噬菌体展示法等取得编码作为原来的抗体的fab的多核苷酸。通过将得到的多核苷酸由如上述一样的pcr法扩增,可得到作为本实施方式的改变抗体的第80及第171个氨基酸残基被半胱氨酸取代的fab。接下来,在本实施方式的制造方法中,回收在上述的取代工序中得到的抗体。例如,将表达改变抗体的宿主细胞溶解于含适当的增溶剂的溶液而使抗体游离到该溶液中。当上述的宿主细胞将改变抗体分泌到培养基中时,回收培养上清。游离的抗体可在亲和性层析法等现有技术中由公知的方法回收。例如,当制造的改变抗体是igg时,可由使用蛋白a或g的亲和性层析法回收。也可根据需要,将回收的抗体由凝胶过滤等的现有技术中公知的方法纯化。由本实施方式的制造方法得到的抗体的详细与对于上述的改变抗体而所述的同样。由本实施方式的制造方法得到的抗体与原来的抗体比较而热稳定性提升。抗体的热稳定性及其测定方法的详细与对于本实施方式的改变抗体而所述的同样。如上所述,在原来的抗体中的第80及第171个氨基酸残基向半胱氨酸的取代几乎不影响对于抗原的亲和性。从而,由本实施方式的制造方法得到的改变抗体与原来的抗体结合于相同的抗原,对于该抗原的亲和性也与原来的抗体是同程度。抗体对于抗原的亲和性及其评价方法的详细细节与对于本实施方式的改变抗体而所述的同样。[3.提升抗体的热稳定性的方法]由提升本实施方式的抗体的热稳定性的方法(以下,也简称为“方法”)可提升基于kabat法的可变区的第80个氨基酸残基和基于kabat法的恒定区的第171个氨基酸残基不是半胱氨酸的抗体的热稳定性。本实施方式的方法作为对象的抗体与上述的“原来的抗体”相同。在本实施方式的方法中,通过对于原来的抗体,将基于kabat法的可变区的第80个氨基酸残基和基于kabat法的恒定区的第171个氨基酸残基取代为半胱氨酸而提升热稳定性。在原来的抗体中的第80及第171个氨基酸残基的取代可如上所述由dna重组技术及其他分子生物学技术等公知的方法进行。例如,当有产生原来的抗体的杂交瘤时,与对于本实施方式的制造方法而所述的同样地可取得含编码改变抗体的多核苷酸的表达载体。进而,将得到的表达载体转导或转染到适当的宿主细胞中而可取得表达上述抗体的宿主细胞。在本实施方式的方法中,认为由向原来的抗体中的第80及第171个氨基酸残基导入半胱氨酸在该半胱氨酸残基间形成二硫键而抗体的热稳定性提升。另外,如上所述,在原来的抗体中的第80及第171个氨基酸残基向半胱氨酸的取代几乎不影响对于抗原的亲和性。从而,将本实施方式的方法,例如,由变异的导入,对于抗原的亲和性提升,但如果应用于热稳定性降低的变异抗体,原样维持该抗体对于抗原的亲和性而提升热稳定性变得可能。在本发明的范围内包括编码本实施方式的热稳定性提升的抗体或其片段的单离并且纯化的多核苷酸。编码本实施方式的改变抗体的片段的单离并且纯化的多核苷酸优选编码在基于kabat法的第80位置含半胱氨酸的可变区及在基于kabat法的第171位置含半胱氨酸的恒定区。在本发明的范围中也包括含上述多核苷酸的载体。载体是为了转导或转染而设计的多核苷酸构建物。载体的种类不特别限定,可从表达载体、克隆载体、病毒载体等现有技术中公知的载体适宜选择。另外,在本发明的范围内也包括含该载体的宿主细胞。宿主细胞的种类不特别限定,可从真核细胞、原核细胞、哺乳动物细胞等适宜选择。接下来,将本发明由实施例详细地说明,但本发明不限定于这些实施例。【实施例】【参考例:在兔fab中的二硫键和热稳定性的关联】如上所述,在非专利文献1中提示兔抗体特有的二硫键(形成在轻链的第80及第171位的半胱氨酸残基间)不贡献于稳定性。但是,基于蛋白质分子内的二硫键有热稳定性相关的可能性的假设,本发明人对兔抗体的fab的热稳定性进行了实际测定。(1)兔fab克隆的取得从用afp免疫的兔的末梢血取得淋巴细胞,从该淋巴细胞提取mrna而合成cdna。将得到的cdna使用用于克隆抗体基因的公知的引物进行扩增,制作fab噬菌体库。使用得到的库,由公知的fab噬菌体展示法及生物淘选(langim,barbascf3rd,schleefrr.,recombinantrabbitfabwithbindingactivitytotype-1plasminogenactivatorinhibitorderivedfromaphage-displaylibraryagainsthumanα-granules,(1996)gene172(2):295-8及philippam.o'brien,robertaitken,antibodyphagedisplay,(2002)methodsinmolecularbiologyvolumeno.178参照)得到3种兔抗afp抗体的fab克隆。将这些克隆以下称为“a1”、“2-2-37”及“c3”。基于各fab克隆的氨基酸序列而研究形成在fab分子中的二硫键的数。得知克隆a1与人抗体及小鼠抗体同样,是有6个二硫键的fab(以下,也称为“ss6个型”)。ss6个型的fab在轻链的可变区内有1个二硫键,在重链的可变区内有1个二硫键,在轻链的恒定区内有1个二硫键,在重链的恒定区内有2个二硫键,及在轻链的恒定区和重链的恒定区之间有1个二硫键。得知克隆2-2-37是有7个二硫键的fab(以下,也称为“ss7个型”)。克隆2-2-37与非专利文献1中记载的兔fab同样,在轻链的可变区的第80位及该轻链的恒定区的第171位有半胱氨酸残基。即,ss7个型的fab除了ss6个型的fab的6个二硫键之外,在轻链的可变区和该轻链的恒定区之间有二硫键。得知克隆c3是有8个二硫键的fab(以下,也称为“ss8个型”)。ss8个型的fab除了ss7个型的fab的7个二硫键之外,在重链的可变区内有二硫键。(2)热稳定性的测定由凝胶过滤,将得到的含各fab克隆的溶液的溶剂取代为在用示差扫描热量计(dsc)的测定中使用的缓冲液(磷酸缓生理盐水:pbs)。凝胶过滤的条件如下所述。[凝胶过滤的条件]缓冲液pbs(ph7.4)使用的柱superdex200increase10/300(gehealthcare公司)柱体积(cv)24ml样品体积500μl流速1.0ml/min溶出量1.5cv级分体积500μl将含各fab克隆的级分用pbs稀释而调制样品(终浓度5μm)。使用microcalpeaq-dsc(malverninstrumentsltd公司)而测定各fab克隆的tm值。测定条件如下所述。[dsc测定条件]样品量400μl测定范围30℃~100℃升温速度1℃/分(3)结果由dsc测定取得的各fab克隆的tm值示于表1。【表1】克隆类型tm(℃)a1ss6个型742-2-37ss7个型81c3ss8个型812-2-37及c3的tm值比a1高7℃。另外,2-2-37和c3是相同的tm值。这些表明,在ss7个型的fab中的轻链的可变区的第80半胱氨酸残基和该轻链的恒定区的第171半胱氨酸残基之间的二硫键贡献于热稳定性。【实施例1:改变抗体的制作】通过向兔抗体(ss6个型)、人抗体及小鼠抗体各自导入变异至在ss7个型的兔抗体形成特有的二硫键,探讨改善各抗体的热稳定性与否。(1)各抗体的基因的取得(1.1)兔抗体的基因的取得通过将在上述的参考例中取得的兔抗afp抗体的fab克隆a1的基因整合到含编码兔抗体的fc区域的基因的质粒dna,取得含兔抗afp抗体的基因的质粒dna。(1.2)人抗体的基因的取得向genscript日本株式会社委托人抗her2单克隆抗体(曲妥珠单抗)的基因的合成而取得含人抗her2抗体的基因的质粒dna。(1.3)小鼠抗体的基因的取得如下所述,取得小鼠抗胰岛素抗体的基因。[试剂]isogen株式会社日本基因smarter(注册商标)race5'/3'试剂盒clontech公司10×a-attachmentmix东洋纺株式会社pcdna(商标)3.4topo(注册商标)ta克隆试剂盒thermofisher公司competenthighdh5α东洋纺株式会社qiaprepspinminiprep试剂盒qiagen公司kodplusneo东洋纺株式会社ligationhighver.2东洋纺株式会社(i)从产生抗体的杂交瘤提取总rna在抗原中使用人胰岛素由kohler及milstein,nature,vol.256,p.495-497,1975中记载的方法制作产生小鼠抗人胰岛素抗体的杂交瘤。将此杂交瘤的培养物(10ml)以1000rpm离心5分钟处理之后,除去上清。将得到的细胞用isogen(1ml)溶解,于室温静置5分钟。向其中添加氯仿(200μl),搅拌15秒钟之后,于室温静置3分钟。进而,将其以12000×g于4℃离心处理10分钟,回收含rna的水相(500μl)。向回收的水相添加异丙醇(500μl)而混合。将得到的混合物于室温静置5分钟之后,以12000×g于4℃离心处理10分钟。除去上清,向得到的沉淀物(总rna)添加70%乙醇(1ml),以7500×g于4℃离心处理10分钟。除去上清,使rna风干,溶解于无rna酶的水(20μl)。(ii)cdna的合成使用在上述(i)中得到的各总rna而调制以下的组成的rna样品。[rna样品]总rna(500ng/μl)1μlrt引物1μl去离子水1.75μl合计3.75μl将调制的rna样品于72℃加热3分钟之后,于42℃温育2分钟。进而,向rna样品添加12μmsmarteriia寡核苷酸(1μl)而调制cdna合成用样品。使用此cdna合成用样品而调制以下的组成的逆转录反应液。[逆转录反应液]5×first-strand缓冲液2μl20mmdtt1μl10mmdntpmix1μlrna酶抑制剂0.25μlsmartscribert(100u/μl)1μlcdna合成用样品4.75μl合计10μl使调制的逆转录反应液于42℃反应90分钟。进而,将反应液于70℃加热10分钟,添加n-[三(羟甲基)甲基]甘氨酸-edta(50μl)。使用得到的溶液作为cdna样品而调制以下组成的5'race反应液。[5'race反应液]10×pcr缓冲液5μldntpmix5μl25mmmg2so43.5μlcdna样品2.5μl10×universalprimermix5μl3'-引物1μlkodplusneo(1u/μl)1μl纯化水27μl合计50μl将调制的5'race反应液在下述的反应条件下供于race反应。再者,下述的“y”对于轻链是90秒钟,对于重链是150秒钟。[反应条件]将于94℃经2分钟、于98℃经10秒钟、于50℃经30秒钟、及于68℃经y秒进行30个循环、及于68℃经3分钟。使用在上述的反应中得到的5’race产物而调制以下组成的溶液。使该溶液于60℃反应30分钟而向5’race产物的末端附加腺嘌呤。5’race产物9μl10×a-attachmentmix1μl合计10μl使用得到的腺嘌呤附加产物及pcdna(商标)3.4topo(注册商标)ta克隆试剂盒而调制以下的组成的ta克隆反应液。将该反应液于室温温育10分钟而将腺嘌呤附加产物克隆到pcdna3.4中。[ta克隆反应液]腺嘌呤附加产物4μl盐溶液1μlpcdna3.41μl合计6μl(iii)转化、质粒提取及序列的确认将在上述(ii)中得到的ta克隆样品(3μl)添加到dh5α(30μl)中,在冰上静置30分钟之后,将混合物于42℃加热45秒钟而进行热休克。再次、在冰上静置2分钟之后,将全量涂布到含有氨苄西林的lb板上。将该板于37℃温育16小时。将板上的单集落取到含有氨苄西林的lb液体培养基中,于37℃振荡培养16小时(250rpm)。将培养物以5000×g离心5分钟处理而回收大肠杆菌的转化体。从回收的大肠杆菌使用qiaprepspinminiprep试剂盒提取质粒。具体的操作根据该试剂盒附带的手册而进行。将得到的质粒的碱基序列使用pcdna3.4载体引物确认。如上取得含小鼠抗胰岛素抗体的基因的质粒dna。(2)各抗体的变异型的基因的取得(2.1)引物的设计及pcr基于各抗体的基因的碱基序列而设计用于将轻链中的可变区的第80个氨基酸残基及恒定区的第171个氨基酸残基变异为半胱氨酸的引物。另外,为了比较,设计用于将小鼠抗胰岛素抗体的轻链中的可变区的第108个氨基酸残基及恒定区的第171个氨基酸残基变异为半胱氨酸的引物。抗体分子的结构上、轻链的第108及第171个氨基酸残基间的距离与可形成二硫键的轻链的第80及第171个氨基酸残基间的距离接近。以下表示各引物的碱基序列。[兔抗afp抗体的引物][可变区]for:5’-tgtgaagatgctgccacttattac-3’seqidno:1rev:5’-cgtcacgccactgatggtga-3’seqidno:2[恒定区]for:5’-tgtacctacagcctgagcagcac-3’seqidno:3rev:5’-gtcttcggggctctgcggtg-3’seqidno:4[人抗her2抗体的引物][可变区]for:5’-tgtgaagacttcgccacgtattac-3’seqidno:5rev:5’-ctgcagagagctgatcgtcagg-3’seqidno:6[恒定区]for:5’-tgtacgtacagcctgagttccacc-3’seqidno:7rev:5’-gtcttttgaatcttgttcggtcacgg-3’seqidno:8[小鼠抗胰岛素抗体的引物][可变区(第80)]for:5’-tgtgaagatgctgccacttattac-3’seqidno:9rev:5’-ctccatgctgctgattgtgag-3’seqidno:10[可变区(第108)]for:5’-tgtgctgatgctgcaccaactgtatc-3’seqidno:11rev:5’-tctgatttccagcttggtgcc-3’seqidno:12恒定区for:5’-tgtacctacagcatgagcagcac-3’seqidno:13rev:5’-gtctttgctgtcctgatcag-3’seqidno:14以含各抗体的基因的质粒dna作为模板使用而调制以下的组成的pcr反应液。[pcr反应液]10×pcr缓冲液5μl25mmmg2so43μl2mmdntpmix5μl正向引物1μl反向引物1μl质粒dna(40ng/μl)0.5μlkodplusneo(1u/μl)1μl纯化水33.5μl合计50μl将调制的pcr反应液在下述的反应条件下供于pcr反应。[反应条件]将于98℃经2分钟、于98℃经10秒钟、于54℃经30秒钟、及于68℃经4分钟进行30个循环、及于68℃经3分钟。向得到的pcr产物(50μl)添加2μl的dpni(10u/μl)而使pcr产物片段化。使用dpni处理的pcr产物而调制以下的组成的连接反应液。将该反应液于16℃温育1小时而进行连接反应。[连接反应液]dpni处理结束pcr产物2μlligationhighver.25μlt4多核苷酸激酶1μl纯化水7μl合计15μl(2.2)转化、质粒dna提取及序列的确认将连接反应后的溶液(3μl)添加到dh5α(30μl)中,与上述(1.3)同样地得到大肠杆菌的转化体。从得到的大肠杆菌使用qiaprepspinminiprep试剂盒提取质粒dna。将得到的各质粒dna的碱基序列使用pcdna3.4载体引物进行确认。以下,以这些质粒dna作为哺乳动物细胞表达用质粒使用。(3)在哺乳动物细胞的表达[试剂]expi293(商标)细胞invitrogen公司expi293(商标)expression培养基invitrogen公司expifectamine(商标)293转染试剂盒invitrogen公司(3.1)转染将expi293细胞在5%co2气氛下、于37℃振荡培养(150rpm)而使增殖。准备对应于样品数的数的30ml的细胞培养物(3.0×106个细胞/ml)。使用编码各抗体的野生型及变异型的质粒dna而调制以下的组成的dna溶液,静置5分钟。[dna溶液]轻链质粒溶液相当于15μg的量(μl)重链质粒溶液相当于15μg的量(μl)opti-mem(商标)适量(ml)合计1.5ml调制以下的组成的转染试剂,静置5分钟。expifectamine试剂80μl质粒溶液1420μl合计1.5ml将调制的dna溶液及转染试剂混合而静置20分钟。将得到的混合液(3ml)添加到细胞培养物(30ml)中,在5%co2气氛下、于37℃振荡培养20小时(150rpm)。20小时后,向各培养物添加各自150μl及1.5ml的expifectamine(商标)转染增强子1及2,在5%co2气氛下、于37℃振荡培养6天(150rpm)。(3.2)抗体的回收及纯化将各细胞培养物以3000rpm离心5分钟处理而回收培养上清。在培养上清中,含从转染的expi293(商标)细胞分泌的各抗体。将得到的培养上清再次以15000×g离心10分钟处理而回收上清。将得到的上清使用hitrapproteinahp柱(gehealthcare公司)进行纯化。将得到的溶液使用superdex200increase10/300gl柱(gehealthcare公司)进行进一步纯化而取得抗体溶液。对于纯化的具体的操作根据各柱的附带文书而进行。(4)结果得到兔抗afp抗体及人抗her2抗体的野生型和作为它们的改变抗体的变异型(cys80-171)。另外,得到小鼠抗胰岛素抗体的野生型和作为其改变抗体的变异型(cys80-171)及变异型(cys108-171)。以下表示各抗体的变异型的详细内容。表2显示兔抗afp抗体的野生型(克隆a1)及其变异型的轻链(κ)的氨基酸序列。在以下的表中,“变异型(cys80-171)”表示在野生型的抗体中的可变区的第80个氨基酸残基及恒定区的第171个氨基酸残基被取代为半胱氨酸的抗体。加下划线部表示可变区,用□框住的残基表示基于kabat法的可变区的第80个氨基酸残基及恒定区的第171个氨基酸残基。【表2】兔抗afp抗体的野生型的轻链(seqidno:15)兔抗afp抗体的变异型(cys80-171)的轻链(seqidno:16)表3显示人抗her2抗体的野生型(曲妥珠单抗)及其变异型的轻链(κ)的氨基酸序列。加下划线部表示可变区,用□框住的残基表示基于kabat法的可变区的第80个氨基酸残基及恒定区的第171个氨基酸残基。【表3】人抗her2抗体的野生型的轻链(seqidno:17)人抗her2抗体的变异型(cys80-171)的轻链(seqidno:18)小鼠抗胰岛素抗体的变异型(cys80-171)是在野生型抗体的轻链(κ)中的可变区的第80丙氨酸残基及恒定区的第171丝氨酸残基被取代为半胱氨酸的抗体。另外,小鼠抗胰岛素抗体的变异型(cys108-171)是在野生型抗体的轻链(κ)中的可变区的第108丙氨酸残基及恒定区的第171丝氨酸残基被取代为半胱氨酸的抗体。【实施例2:改变抗体的热稳定性】探讨在实施例1中制作的各抗体的野生型及它们的变异型的热稳定性。(1)热稳定性的测定由凝胶过滤将实施例1中得到的含各抗体的溶液的溶剂取代为pbs(ph7.4)。将含各抗体的级分用pbs稀释而调制样品(终浓度5μm)。使用microcalpeaq-dsc(malverninstrumentsltd公司)而测定各抗体的tm值。凝胶过滤及dsc测定的条件与参考例中所述的条件相同。(2)结果由dsc测定取得的各抗体的tm值示于以下的表4~7。另外,各抗体的解析峰示于图1、图2及图3a~3c。【表4】兔抗afp抗体(igg)tm(℃)野生型76.6变异型(cys80-171)80.0【表5】人抗her2抗体(fab)tm(℃)野生型80.9变异型(cys80-171)84.4【表6】小鼠抗胰岛素抗体(igg)tm(℃)野生型76.4变异型(cys80-171)80.1【表7】小鼠抗胰岛素抗体(fab)tm(℃)野生型77.3变异型(cys80-171)80.6变异型(cys108-171)75.2如表4~7所示,兔抗体、人抗体及小鼠抗体的变异型(cys80-171)均变得与各抗体的野生型比,tm值高3℃以上。从而表明,通过在基于kabat法的可变区的第80个氨基酸残基及恒定区的第171个氨基酸残基不是半胱氨酸的抗体中,将这些氨基酸残基取代为半胱氨酸,可提升抗体的热稳定性。其中,在非专利文献1中记载了由于在兔抗体中在轻链的第80及第171位的磺基丙氨酸残基间形成二硫键,从而在各抗体的变异型(cys80-171)中也同样地形成二硫键的可能性高。从而提示,半胱氨酸残基的导入所导致的热稳定性的提升因在ss7个型的兔抗体中形成特有的二硫键。一方面,小鼠抗体的变异型(cys108-171)变得与野生型比,tm值低2℃以上。如上所述,轻链的第108及第171个氨基酸残基处于抗体分子的结构上、接近于轻链的第80及第171个氨基酸残基的位置。但是表明,即使将轻链的第108及第171个氨基酸残基取代为半胱氨酸,热稳定性也不提升,反倒降低。【实施例3:改变抗体对于抗原的亲和性】将在实施例1中制作的各抗体的野生型及它们的变异型对抗原的亲和性由elisa法探讨。(1)由elisa法的测定(1.1)抗原及检测用抗体作为兔抗afp抗体的抗原,使用afp蛋白质(leebiosolutions公司、货号:105-11)。作为人抗her2抗体的抗原,使用her2蛋白质(r&dsystems公司、货号:1129-er)。作为小鼠抗胰岛素抗体的抗原,使用humulinr注100单位(elililly公司)。将在实施例1中制作的各抗体用含有1%bsa的pbs阶段性地稀释而得到浓度不同的多种抗体溶液。(1.2)测定将各抗原用pbs(ph7.4)稀释而调制各抗原的溶液。将各抗原的溶液添加到maxisorp(商标)平底板(thermofisher公司)的孔中,于4℃静置一晚。除去抗原溶液,将封闭溶液(含有1%bsa的pbs)添加到各孔中而封闭。在除去封闭溶液之后,向各孔添加各100μl各抗体溶液,将抗原抗体反应于室温进行1小时。除去抗体溶液,向各孔添加清洗液(含有1%bsa的pbs)而清洗孔。清洗后,对应于检测用抗体的种类,添加hrp标记抗兔fc抗体、hrp标记抗人fc抗体或hrp标记抗小鼠fc抗体的溶液而于室温进行抗原抗体反应。除去抗体溶液,向各孔添加清洗液(含有1%bsa的pbs)而清洗孔。清洗后,将abst底物(thermofisher公司)的溶液添加到各孔中,对在450nm处的吸光度进行测定。(2)结果如图4~6所示,兔抗体、人抗体及小鼠抗体之任一者的变体对于抗原的亲和性均与各抗体的野生型大致相同。从而表明,基于kabat法的可变区的第80个氨基酸残基及恒定区的第171个氨基酸残基向半胱氨酸的取代几乎不影响对于抗原的亲和性。从而提示,通过将基于kabat法的可变区的第80个氨基酸残基及恒定区的第171个氨基酸残基取代为半胱氨酸,可不影响对于抗原的亲和性而提升抗体的热稳定性。【比较例:改变其他部位的抗体的热稳定性】探讨当将野生型的抗体中的基于kabat法的可变区的第80个氨基酸残基及恒定区的第171个氨基酸残基的附近的氨基酸残基取代为半胱氨酸时,抗体的热稳定性是否也提升。(1)改变其他部位的抗体的制作与实施例1同样地,将小鼠抗胰岛素抗体的野生型中的基于kabat法的可变区的选自第79~81位的1个氨基酸残基和基于kabat法的恒定区的选自第169~171位的1个氨基酸残基取代为半胱氨酸而制作小鼠抗胰岛素抗体的变异型。另外,将人her2抗体的野生型中的基于kabat法的可变区的第80个氨基酸残基和基于kabat法的恒定区的170或第171个氨基酸残基取代为半胱氨酸而制作人her2抗体的变异型。(2)热稳定性的测定与实施例2同样地对各抗体的野生型及它们的变异型的热稳定性进行测定。结果示于表8及9。【表8】小鼠抗胰岛素抗体(fab)tm(℃)野生型77.3变异型(cys80-171)80.6变异型(cys79-169)77.2变异型(cys79-170)76.6变异型(cys79-171)76.5变异型(cys80-169)76.1变异型(cys80-170)75.1变异型(cys81-169)74.9变异型(cys81-170)76.1变异型(cys81-171)74.4【表9】人抗her2抗体(fab)tm(℃)野生型80.9变异型(cys80-171)84.4变异型(cys80-170)78.7如表8及9所示,在变异型(cys80-171)中,热稳定性比野生型提升,但在其他变异型中,热稳定性比野生型降低。这些结果表明,当将野生型抗体中的基于kabat法的可变区的第80个氨基酸残基及恒定区的第171个氨基酸残基的附近的氨基酸残基取代为半胱氨酸时,抗体的热稳定性不提升。序列表<110>sysmexcorporation<120>抗体及其制造方法、以及提升抗体的热稳定性的方法<130>17-028cn<160>18<170>patentinversion3.5<210>1<211>24<212>dna<213>人工序列<220><223>引物<400>1tgtgaagatgctgccacttattac24<210>2<211>20<212>dna<213>人工序列<220><223>引物<400>2cgtcacgccactgatggtga20<210>3<211>23<212>dna<213>人工序列<220><223>引物<400>3tgtacctacagcctgagcagcac23<210>4<211>20<212>dna<213>人工序列<220><223>引物<400>4gtcttcggggctctgcggtg20<210>5<211>24<212>dna<213>人工序列<220><223>引物<400>5tgtgaagacttcgccacgtattac24<210>6<211>22<212>dna<213>人工序列<220><223>引物<400>6ctgcagagagctgatcgtcagg22<210>7<211>24<212>dna<213>人工序列<220><223>引物<400>7tgtacgtacagcctgagttccacc24<210>8<211>26<212>dna<213>人工序列<220><223>引物<400>8gtcttttgaatcttgttcggtcacgg26<210>9<211>24<212>dna<213>人工序列<220><223>引物<400>9tgtgaagatgctgccacttattac24<210>10<211>21<212>dna<213>人工序列<220><223>引物<400>10ctccatgctgctgattgtgag21<210>11<211>26<212>dna<213>人工序列<220><223>引物<400>11tgtgctgatgctgcaccaactgtatc26<210>12<211>21<212>dna<213>人工序列<220><223>引物<400>12tctgatttccagcttggtgcc21<210>13<211>23<212>dna<213>人工序列<220><223>引物<400>13tgtacctacagcatgagcagcac23<210>14<211>20<212>dna<213>人工序列<220><223>引物<400>14gtctttgctgtcctgatcag20<210>15<211>215<212>prt<213>穴兔(oryctolaguscuniculus)<400>15gluleuvalleuthrglnthrproserservalseralaalavalgly151015glythrvalthrileasncysglnalaserglnservalserasnleu202530leualatrptyrglnglnlysproglyglnproprolysleuleuile354045tyrglyalaserasnleugluserglyvalproserargphearggly505560serglyserglythrglnphethrleuthrileserglyvalthrala65707580gluaspalaalathrtyrtyrcysglnserglytyrtyrserthrgly859095alailethrpheglylysglythrlysleugluilelysargasppro100105110valalaproservalleuleupheproproserlysglugluleuthr115120125thrglythralathrilevalcysvalalaasnlysphetyrproser130135140aspilethrvalthrtrplysvalaspglythrthrglnglnsergly145150155160ilegluasnserlysthrproglnserprogluaspasnthrtyrser165170175leuserserthrleuserleuthrseralaglntyrasnserhisser180185190valtyrthrcysgluvalvalglnglyseralaserproilevalgln195200205serpheasnargglyaspcys210215<210>16<211>215<212>prt<213>人工序列<220><223>改变抗体<400>16gluleuvalleuthrglnthrproserservalseralaalavalgly151015glythrvalthrileasncysglnalaserglnservalserasnleu202530leualatrptyrglnglnlysproglyglnproprolysleuleuile354045tyrglyalaserasnleugluserglyvalproserargphearggly505560serglyserglythrglnphethrleuthrileserglyvalthrcys65707580gluaspalaalathrtyrtyrcysglnserglytyrtyrserthrgly859095alailethrpheglylysglythrlysleugluilelysargasppro100105110valalaproservalleuleupheproproserlysglugluleuthr115120125thrglythralathrilevalcysvalalaasnlysphetyrproser130135140aspilethrvalthrtrplysvalaspglythrthrglnglnsergly145150155160ilegluasnserlysthrproglnserprogluaspcysthrtyrser165170175leuserserthrleuserleuthrseralaglntyrasnserhisser180185190valtyrthrcysgluvalvalglnglyseralaserproilevalgln195200205serpheasnargglyaspcys210215<210>17<211>214<212>prt<213>智人(homosapiens)<400>17aspileglnmetthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnaspvalasnthrala202530valalatrptyrglnglnlysproglylysalaprolysleuleuile354045tyrseralaserpheleutyrserglyvalproserargphesergly505560serargserglythraspphethrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysglnglnhistyrthrthrpropro859095thrpheglyglnglythrlysvalgluilelysargthrvalalaala100105110proservalpheilepheproproseraspgluglnleulyssergly115120125thralaservalvalcysleuleuasnasnphetyrproarggluala130135140lysvalglntrplysvalaspasnalaleuglnserglyasnsergln145150155160gluservalthrgluglnaspserlysaspserthrtyrserleuser165170175serthrleuthrleuserlysalaasptyrglulyshislysvaltyr180185190alacysgluvalthrhisglnglyleuserleuprovalthrlysser195200205pheasnargglyglucys210<210>18<211>214<212>prt<213>人工序列<220><223>改变抗体<400>18aspileglnmetthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnaspvalasnthrala202530valalatrptyrglnglnlysproglylysalaprolysleuleuile354045tyrseralaserpheleutyrserglyvalproserargphesergly505560serargserglythraspphethrleuthrileserserleuglncys65707580gluaspphealathrtyrtyrcysglnglnhistyrthrthrpropro859095thrpheglyglnglythrlysvalgluilelysargthrvalalaala100105110proservalpheilepheproproseraspgluglnleulyssergly115120125thralaservalvalcysleuleuasnasnphetyrproarggluala130135140lysvalglntrplysvalaspasnalaleuglnserglyasnsergln145150155160gluservalthrgluglnaspserlysaspcysthrtyrserleuser165170175serthrleuthrleuserlysalaasptyrglulyshislysvaltyr180185190alacysgluvalthrhisglnglyleuserleuprovalthrlysser195200205pheasnargglyglucys210当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1