一种β-半乳糖苷酶、其编码基因及其应用的制作方法
本发明涉及一种β-半乳糖苷酶、其编码基因及其应用,属于生物技术领域。
背景技术:
低聚半乳糖是一种功能性低聚糖,作为一种新型的食品添加剂,广泛的应用于食品行业。目前虽然已经有商品化的低聚半乳糖出售,但是价格昂贵。我国的低聚半乳糖生产未成规模,其核心技术瓶颈是缺乏高效制备低聚半乳糖的酶。因此,挖掘性能优良的工业用酶,对推动低聚半乳糖的产业化至关重要。
β-半乳糖苷酶(β-galactosidase,ec.3.2.1.23),全称为β-d-半乳糖苷半乳糖水解酶,商品名为乳糖酶,往往具有催化乳糖水解和转糖苷两种功能。β-半乳糖苷酶的转糖苷功能,可以以乳糖为底物,合成低聚半乳糖。但是绝大部分β-半乳糖苷酶的转糖苷活性较低,合成低聚半乳糖的效率很低。例如,中国发明专利申请201610579268.4中的β-半乳糖苷酶的低聚半乳糖转化率仅为9.5%;中国发明专利申请201610580177.2中的β-半乳糖苷酶的低聚半乳糖转化率仅为11.5%。已知较好的合成低聚半乳糖的酶来自bacilluscirulans菌,其转化率为48.3%。
有机溶剂催化最独特的优势是促使水解酶转向合成反应。其原因是反应中添加了有机溶剂后,水活度下降,会更有利于进行合成反应。β-半乳糖苷酶是一个既有水解能力又有合成能力(转糖苷)的酶,因此,理论上在有机溶剂中实施β-半乳糖苷酶催化合成低聚半乳糖,有可能会提高合成效率。但是有机溶剂对酶有毒性,绝大部分的β-半乳糖苷酶不能耐受有机溶剂,因此也就无法实现在有机溶剂存在时高效合成低聚半乳糖。
技术实现要素:
针对上述现有技术中所存在的问题,本发明提供了一种β-半乳糖苷酶、其编码基因及其应用。本发明提供的β-半乳糖苷酶具有有机溶剂耐受性,能够在有机溶剂中高效催化合成低聚半乳糖。
本发明β-半乳糖苷酶,其氨基酸序列如seqidno:1所示,或者与所述seqidno:1序列有92%以上的相似性或同源性的氨基酸序列。
编码所述β-半乳糖苷酶的基因,其核苷酸序列如seqidno:2所示。该序列已经经过序列优化,能在大肠杆菌中有效表达出有酶活力的β-半乳糖苷酶。含有上述β-半乳糖苷酶的基因的表达载体为pet32a;含有上述表达载体的重组表达宿主为大肠杆菌bl21(de3)。
用于表达所述β-半乳糖苷酶的重组表达质粒,所述重组表达质粒包含编码所述β-半乳糖苷酶的基因。
本发明产生所述β-半乳糖苷酶的重组菌株,所述重组菌株含有上述重组表达质粒。
本发明β-半乳糖苷酶的应用,是在有机溶剂体系中高效催化合成低聚半乳糖。具体包括如下步骤:
以乳糖为底物,以ph值6.0-7.5的磷酸盐缓冲液为溶剂配制浓度为300-500g/l的乳糖溶液,向反应体系中添加有机溶剂以及β-半乳糖苷酶,40-60℃下反应合成低聚半乳糖。
所述有机溶剂为乙二醇或丁二醇,有机溶剂的添加体积为磷酸盐缓冲液体积的10-20%,优选为20%。
所述β-半乳糖苷酶的添加比例为10u/ml。
本发明β-半乳糖苷酶高效催化合成低聚半乳糖的反应条件优选为:乳糖浓度400g/l,酶量10u/ml,磷酸盐缓冲液ph值6.5,反应温度50℃,反应时间10h。
本发明β-半乳糖苷酶在没有有机溶剂的环境中也能有效合成低聚半乳糖。
本发明β-半乳糖苷酶催化合成低聚半乳糖,低聚半乳糖的最高产物转化效率达到55.8±0.8%,是无有机溶剂时的1.7倍,因此具有很好的工业应用价值。
附图说明
图1是本发明β-半乳糖苷酶在50℃时的温度稳定性曲线。
图2是本发明β-半乳糖苷酶合成低聚半乳糖的最适ph曲线。
图3是本发明β-半乳糖苷酶合成低聚半乳糖的最适温度曲线。
图4是本发明β-半乳糖苷酶在无有机溶剂时合成低聚半乳糖的hplc图谱。
图5是本发明β-半乳糖苷酶在存在乙二醇时合成低聚半乳糖的hplc图谱。
具体实施方式
实施例1:β-半乳糖苷酶的基因和蛋白序列
之前的研究中实验室保存过一株黄热厌氧芽孢杆菌(anoxybacillusfavithermus)ne3,采用酚-氯仿法抽提总dna。基于genebank中公布的来源anoxybacillus属的β-半乳糖苷酶的序列,设计正向引物为cccggatccatgggggagcgaaacatg,反向引物为cccgagctctttaactccatgattcatg。通过pcr扩增,获得一条长度为1.9kb条带。将该条带测序后,获得编码该基因的蛋白质序列如seqidno:1所示。ncbi序列比对结果表明,该蛋白序列与编号为wp_032100326的β-半乳糖苷酶具有90%的序列相似性。将获得的野生型的基因序列克隆到大肠杆菌中进行重组表达,未见重组蛋白。因此,委托上海生工生物有限公司优化了核苷酸序列,人工合成了基因序列如seqidno:2所示。该序列在大肠杆菌中成功获得了有活性的表达。
实施例2:β-半乳糖苷酶的重组表达和蛋白质纯化
人工合成的β-半乳糖苷酶基因,直接克隆在表达载体pet32a中。质粒转化大肠杆菌bl21(de3)中,培养至od600为0.6-0.8,加1mm的诱导物iptg,37℃诱导表达6h。离心收集菌体,超声破碎细胞。采用镍亲和层析柱纯化重组蛋白。sds-page检测蛋白纯度。
实施例3:β-半乳糖苷酶在50℃的温度稳定性
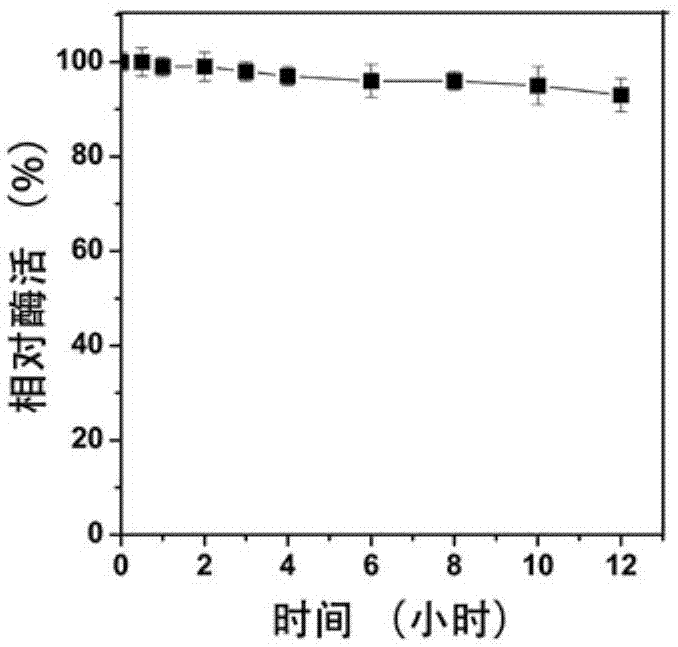
本案例中的β-半乳糖苷酶是在50℃进行应用,因此检测了该酶50℃时的温度稳定性。β-半乳葡萄糖苷酶的水解活性和转糖苷活性测定方法不同。虽然水解活性的高低与转糖苷活性高低之间没有直接关联,但是可以用水解活性来衡量酶的稳定性。水解活性的测定以onpg为底物,一个标准反应体系包含10μl适当稀释的纯酶,30μl100mm的onpg和260μlna2hpo4-kh2po4缓冲液(50mm,ph6.5)。在55℃反应5分钟,加等体积(300ul)1mna2co3溶液终止反应。冷却后在od415测吸光值。以不加酶的样品为空白对照。温度稳定性的测定方法为,用na2hpo4-kh2po4缓冲液(50mm,ph6.5)适当稀释纯化的β-半乳糖苷酶,然后将酶孵育在50℃下,间隔一定时间取样,然后加入底物onpg等在55℃反应10min,测残留酶活。以未孵育的样品组酶活作为100%。获得的温度稳定性曲线如图1所示。该β-半乳糖苷酶在50℃的温度稳定性非常好,孵育12小时后,依然保留了93%以上的酶活。
实施例4:β-半乳糖苷酶的有机溶剂耐受性
不同的有机溶剂的毒性不同,通常认为可以用极性系数logpow值(溶剂在辛醇/水双相系统中的分配系数的对数值)表征有机溶剂的毒性。选择logpow值从3.5到-1.93的12种有机溶剂加入标准反应体系中,有机溶剂的终浓度分别为10%(v/v)、15%(v/v)和20%(v/v)。室温放置1小时后,在55℃反应10min,测残留酶活。以无添加的样品组的酶活(201.5±4.5u/mg)定义为100%。获得的β-半乳糖苷酶的有机溶剂耐受性的结果如表1所示。反应体系中含有10%有机溶剂时,β-半乳糖苷酶对这12种有机溶剂均表现出了非常优良的耐受性,即使是最差的正己醇也保持了58%以上的活性。有机溶剂的浓度提高到20%时,β-半乳糖苷酶对9种有机溶剂依然保留了一半以上酶活,尤其是丁二醇、乙二醇、甲苯、正己烷和环己烷这5种有机溶剂,β-半乳糖苷酶保持了92%以上的酶活。这些数据表明该β-半乳糖苷酶是一个优良的有机溶剂耐受酶,在有机溶剂催化中具有一定的应用价值。
表1有机溶剂对β-半乳糖苷酶酶活的影响
实施例5:β-半乳糖苷酶在无有机溶剂时合成低聚半乳糖
本案例中的β-半乳糖苷酶具有转糖苷活性,可以催化乳糖合成低聚半乳糖。测定了合成低聚半乳糖反应时的最适ph和最适反应温度。低聚半乳糖的合成量用高效液相色谱hplc检测。色谱条件为:安捷伦1260hplc色谱仪,色谱柱aminexhpx-87c(300mm×7.8mm),蒸发光散射检测器,流动相为超纯水流速为0.6ml/min,柱温80℃。
在最适ph6.5下测定温度对低聚半乳糖合成的影响,温度范围为30到65℃。在50℃测定不同ph值对低聚半乳糖合成的影响。缓冲液为50mmna2hpo4-柠檬酸缓冲液(ph4.0到5.0),na2hpo4-kh2po4缓冲液(ph5.0到8.0)和甘氨酸-naoh缓冲(ph8.0到10.0)。用这些不同的缓冲液配制配置400g/l的乳糖溶液。然后按照10u/ml的比例加入纯化的β-半乳糖苷酶,在转速为100rpm的恒温摇床中震荡反应10h后,沸水浴5分钟使酶失活,离心去除蛋白。上清液进行hplc检测。获得的最适ph是6.5(图2),最适反应温度是50℃(图3)。最适反应条件下,乳糖合成低聚半乳糖的转化率为32±0.3%。hplc的检测图见图4,主要产物是半乳三塘、半乳四糖和半乳五糖。
实施例6:β-半乳糖苷酶在存在有机溶剂时合成低聚半乳糖
本案例中的β-半乳糖苷酶在存在有机溶剂时,合成低聚半乳糖的效率显著增强。综合考虑了β-半乳糖苷酶的有机溶剂耐受性和有机溶剂自身的毒性后,选择使用乙二醇或丁二醇。用na2hpo4-kh2po4缓冲液(50mm,ph6.5)配置400g/l的乳糖溶液,按照20%(v/v)的比例加入乙二醇或丁二醇,按照10u/ml的比例加入纯化的β-半乳糖苷酶。为了防止有机溶剂的挥发,反应在能严格密封的亨盖特厌氧试管中进行。试管放置于转速为100rpm的50℃恒温摇床中震荡反应10h后,沸水浴5分钟使酶失活,离心去除蛋白。上清液进行hplc检测。添加乙二醇时的低聚半乳糖的转化率最高,达到55.8±0.8%,主要产物依然是半乳三塘、半乳四糖和半乳五糖,但是可以明显看到半乳四糖和半乳五糖合成的量增多(图5)。添加丁二醇时的低聚半乳糖的转化率略低,为48.5±0.9%。由此可见,有机溶剂的添加提高1.7倍的转化率,使低聚半乳糖的酶法制备具有了很好的工业应用价值。
seqidno:1
<β-galactosidasefromanoxybacillusfavithermus>
myigvdyypeqwpkemieediqgmkalganavrigefswarlmqqtenidfsffdeiidklkenglsimfgtptatfpawlarkhpvdlskdeygrvrvfggrrqycfnsktyrlyhpniteqlvthykdhlaivawqidnefghedmcycenchkefqqwlqekykhinalneaygtifwgqtyndfseipmptktitthnpsllldyarfmsfsvnrfahemtniikkhkphqqvttnvsggffnkwfdheenvrlmdfvswdnypvwdggldepvspaaiamshdfnrgllgrnywimeqlmgaqghemigvlprpnqakmwsyqafahgctnmlyfqwrgmdrgaeqftegivdhsnmrgrkykevqsvgshiptlefglqspiqadiavlfdyeniwslrfqpqskafdfttellrlkapfyrlnanidvipvsrdfstykvlvipvlqiidetlaeklkmfarsgglpifsfrtgtknkqnnihfkhilpahvsdlvgayiheveslagrhvpivgeghhkgrkamcsvwrdllepttrnvlyryddpfyplaaitenqygdgnvyyigtfidgdvlndkpkymkqkhniwhvesdegvevyrrvcdggvdmfllnhtdqekqielfegldlkpylsqivki
seqidno:2
atgtacatcggtgtggattattatccggaacagtggccgaaagaaatgattgaagaagatattcagggcatgaaagcactgggtgccaatgccgtgcgtattggcgaattttcttgggcacgtctgatgcagcagaccgaaaatattgattttagtttctttgacgagatcatcgataaactgaaagaaaatggtctgagcattatgtttggcaccccgaccgccacctttccggcatggctggcccgtaaacatccggtggatctgagtaaagatgaatatggtcgcgtgcgtgtttttggcggtcgtcgtcagtattgctttaatagcaaaacctatcgcctgtatcatccgaatattaccgaacagctggtgacccattataaagatcatctggccattgtggcatggcagattgataatgaatttggtcatgaagatatgtgttactgtgaaaattgtcataaggaatttcagcagtggctgcaggaaaaatataaacatattaacgccctgaacgaagcctatggtacaattttctggggtcagacctataatgattttagcgaaattccgatgccgaccaaaaccattaccacccataatccgagtctgctgctggattatgcccgttttatgagttttagtgtgaatcgctttgcacatgaaatgaccaatattattaagaagcacaagccgcatcagcaggttaccaccaatgtgagtggtggctttttcaataagtggtttgatcatgaagagaatgtgcgtctgatggattttgttagttgggataattatccggtgtgggatggcggcctggatgaaccggtgagcccggcagccattgcaatgagtcatgattttaatcgcggcctgctgggtcgtaattattggattatggaacagctgatgggtgcccagggccatgaaatgattggtgtgctgccgcgtccgaatcaggcaaaaatgtggagctatcaggcatttgcacatggttgcaccaatatgctgtattttcagtggcgcggtatggatcgtggtgccgaacagtttaccgaaggtattgtggatcatagtaatatgcgcggccgtaaatataaagaagttcagagcgtgggtagtcatattccgaccctggaatttggtctgcagagtccgattcaggcagatattgcagttctgtttgattatgaaaacatctggagtctgcgctttcagccgcagagtaaagcctttgattttaccaccgaactgctgcgtctgaaagcaccgttttatcgtctgaatgccaatattgatgttattccggtgagtcgcgattttagtacctataaagtgctggttattccggttctgcagattattgatgaaaccctggcagaaaaactgaaaatgtttgcccgtagcggtggtctgccgatttttagttttcgtaccggcaccaaaaataagcagaataatattcatttcaagcacatcctgccggcccatgttagtgatctggttggtgcctatattcatgaagttgaaagtctggcaggccgccatgttccgattgtgggcgaaggccatcataaaggtcgcaaagccatgtgtagcgtgtggcgcgatctgctggaaccgaccacccgtaatgttctgtatcgctatgatgatccgttttatccgctggcagcaattaccgaaaatcagtatggcgatggtaatgtttattatattggtacattcatcgacggtgacgtgctgaatgataaaccgaaatatatgaaacagaagcataacatttggcatgtggaaagtgatgaaggcgtggaagtgtatcgccgtgtgtgtgatggcggcgttgatatgtttctgctgaatcataccgatcaggaaaaacagattgaactgtttgaaggcctggatctgaaaccgtatctgagtcagattgtgaaaatttaa
序列表
<110>安徽大学
<120>一种β-半乳糖苷酶、其编码基因及其应用
<160>2
<170>siposequencelisting1.0
<210>1
<211>1947
<212>dna
<213>anoxybacillusfavithermus
<400>1
atgtacatcggtgtggattattatccggaacagtggccgaaagaaatgattgaagaagat60
attcagggcatgaaagcactgggtgccaatgccgtgcgtattggcgaattttcttgggca120
cgtctgatgcagcagaccgaaaatattgattttagtttctttgacgagatcatcgataaa180
ctgaaagaaaatggtctgagcattatgtttggcaccccgaccgccacctttccggcatgg240
ctggcccgtaaacatccggtggatctgagtaaagatgaatatggtcgcgtgcgtgttttt300
ggcggtcgtcgtcagtattgctttaatagcaaaacctatcgcctgtatcatccgaatatt360
accgaacagctggtgacccattataaagatcatctggccattgtggcatggcagattgat420
aatgaatttggtcatgaagatatgtgttactgtgaaaattgtcataaggaatttcagcag480
tggctgcaggaaaaatataaacatattaacgccctgaacgaagcctatggtacaattttc540
tggggtcagacctataatgattttagcgaaattccgatgccgaccaaaaccattaccacc600
cataatccgagtctgctgctggattatgcccgttttatgagttttagtgtgaatcgcttt660
gcacatgaaatgaccaatattattaagaagcacaagccgcatcagcaggttaccaccaat720
gtgagtggtggctttttcaataagtggtttgatcatgaagagaatgtgcgtctgatggat780
tttgttagttgggataattatccggtgtgggatggcggcctggatgaaccggtgagcccg840
gcagccattgcaatgagtcatgattttaatcgcggcctgctgggtcgtaattattggatt900
atggaacagctgatgggtgcccagggccatgaaatgattggtgtgctgccgcgtccgaat960
caggcaaaaatgtggagctatcaggcatttgcacatggttgcaccaatatgctgtatttt1020
cagtggcgcggtatggatcgtggtgccgaacagtttaccgaaggtattgtggatcatagt1080
aatatgcgcggccgtaaatataaagaagttcagagcgtgggtagtcatattccgaccctg1140
gaatttggtctgcagagtccgattcaggcagatattgcagttctgtttgattatgaaaac1200
atctggagtctgcgctttcagccgcagagtaaagcctttgattttaccaccgaactgctg1260
cgtctgaaagcaccgttttatcgtctgaatgccaatattgatgttattccggtgagtcgc1320
gattttagtacctataaagtgctggttattccggttctgcagattattgatgaaaccctg1380
gcagaaaaactgaaaatgtttgcccgtagcggtggtctgccgatttttagttttcgtacc1440
ggcaccaaaaataagcagaataatattcatttcaagcacatcctgccggcccatgttagt1500
gatctggttggtgcctatattcatgaagttgaaagtctggcaggccgccatgttccgatt1560
gtgggcgaaggccatcataaaggtcgcaaagccatgtgtagcgtgtggcgcgatctgctg1620
gaaccgaccacccgtaatgttctgtatcgctatgatgatccgttttatccgctggcagca1680
attaccgaaaatcagtatggcgatggtaatgtttattatattggtacattcatcgacggt1740
gacgtgctgaatgataaaccgaaatatatgaaacagaagcataacatttggcatgtggaa1800
agtgatgaaggcgtggaagtgtatcgccgtgtgtgtgatggcggcgttgatatgtttctg1860
ctgaatcataccgatcaggaaaaacagattgaactgtttgaaggcctggatctgaaaccg1920
tatctgagtcagattgtgaaaatttaa1947
<210>2
<211>648
<212>prt
<213>anoxybacillusfavithermus
<400>2
mettyrileglyvalasptyrtyrprogluglntrpprolysglumet
151015
ileglugluaspileglnglymetlysalaleuglyalaasnalaval
202530
argileglygluphesertrpalaargleumetglnglnthrgluasn
354045
ileasppheserphepheaspgluileileasplysleulysgluasn
505560
glyleuserilemetpheglythrprothralathrpheproalatrp
65707580
leualaarglyshisprovalaspleuserlysaspglutyrglyarg
859095
valargvalpheglyglyargargglntyrcyspheasnserlysthr
100105110
tyrargleutyrhisproasnilethrgluglnleuvalthrhistyr
115120125
lysasphisleualailevalalatrpglnileaspasngluphegly
130135140
hisgluaspmetcystyrcysgluasncyshislysglupheglngln
145150155160
trpleuglnglulystyrlyshisileasnalaleuasnglualatyr
165170175
glythrilephetrpglyglnthrtyrasnaspphesergluilepro
180185190
metprothrlysthrilethrthrhisasnproserleuleuleuasp
195200205
tyralaargphemetserpheservalasnargphealahisglumet
210215220
thrasnileilelyslyshislysprohisglnglnvalthrthrasn
225230235240
valserglyglyphepheasnlystrppheasphisglugluasnval
245250255
argleumetaspphevalsertrpaspasntyrprovaltrpaspgly
260265270
glyleuaspgluprovalserproalaalailealametserhisasp
275280285
pheasnargglyleuleuglyargasntyrtrpilemetgluglnleu
290295300
metglyalaglnglyhisglumetileglyvalleuproargproasn
305310315320
glnalalysmettrpsertyrglnalaphealahisglycysthrasn
325330335
metleutyrpheglntrpargglymetaspargglyalagluglnphe
340345350
thrgluglyilevalasphisserasnmetargglyarglystyrlys
355360365
gluvalglnservalglyserhisileprothrleuglupheglyleu
370375380
glnserproileglnalaaspilealavalleupheasptyrgluasn
385390395400
iletrpserleuargpheglnproglnserlysalapheaspphethr
405410415
thrgluleuleuargleulysalaprophetyrargleuasnalaasn
420425430
ileaspvalileprovalserargasppheserthrtyrlysvalleu
435440445
valileprovalleuglnileileaspgluthrleualaglulysleu
450455460
lysmetphealaargserglyglyleuproilepheserpheargthr
465470475480
glythrlysasnlysglnasnasnilehisphelyshisileleupro
485490495
alahisvalseraspleuvalglyalatyrilehisgluvalgluser
500505510
leualaglyarghisvalproilevalglygluglyhishislysgly
515520525
arglysalametcysservaltrpargaspleuleugluprothrthr
530535540
argasnvalleutyrargtyraspaspprophetyrproleualaala
545550555560
ilethrgluasnglntyrglyaspglyasnvaltyrtyrileglythr
565570575
pheileaspglyaspvalleuasnasplysprolystyrmetlysgln
580585590
lyshisasniletrphisvalgluseraspgluglyvalgluvaltyr
595600605
argargvalcysaspglyglyvalaspmetpheleuleuasnhisthr
610615620
aspglnglulysglnilegluleuphegluglyleuaspleulyspro
625630635640
tyrleuserglnilevallysile
645
- 还没有人留言评论。精彩留言会获得点赞!