拟南芥At4g36920基因在调控植物原花色素生物合成及反刍动物抗臌胀病的应用的制作方法
本发明属于基因工程应用的技术领域,具体为拟南芥at4g36920基因在调控植物原花色素生物合成及反刍动物抗臌胀病的应用。
背景技术:
原花色素(proanthocyanidins)是植物中具有重要活性的类黄酮次生代谢产物。不仅在种子休眠、寿命和萌发的调控中起着重要作用,而且还参与调控植物的生物和非生物逆境胁迫(debeaujonetal.,2003);棕色棉是目前应用最广泛的天然彩色棉,其纤维中的棕色色素就是原花色素(xiaoetal.,2014;yanetal.,2018);紫花苜蓿中原花色素含量高于干重的2%时,可以有效地防止牛、羊等反刍动物鼓胀病的发生(verdieretal.,2012);除此之外,还具有抗氧化、抗炎和抗癌等有益于人类健康的功效(dixonetal.,2005)。因此,原花色素生物合成和调控方面的研究,成为植物次生代谢领域内的一个重要研究方向。
原花色素主要在植物的种皮、叶、花、果实以及树皮等部位积累,其含量受各种因素的影响(dixonetal.,2005)。拟南芥中原花色素主要在种皮中积累,其生物合成起始于双受精后1到2天的珠孔区域,随后在内种皮中朝着合点的方向逐渐积累,直到大概5到6天时在合点区域的生物合成结束(debeaujonetal.,2003;lepiniecetal.,2006)。因此,更准确地说,原花色素在拟南芥的内种皮、珠孔和合点部位积累。目前,原花色素的生物合成途径已经比较清楚。但是,原花色素生物合成的调控是一个复杂的过程,其调控的分子机制还有很多问题亟待回答。
目前的研究表明,原花色素的生物合成主要受到mbw三元复合物(r2r3-myb(tt2/myb5)、bhlh(tt8/gl3/egl3)和wd40(ttg1))的调控。tt2-tt8-ttg1三元复合物是原花色素生物合成的主要正调控蛋白复合体,而种皮特异表达的tt2基因是原花色素积累的关键因子。当在拟南芥中过量表达tt2基因时,原花色素的含量没有增加(nesietal.,2001),进一步同时过量表达tt2和pap1时也只在一些特异的细胞中检测到原花色素(sharmaetal.,2005)。一些研究试图增加紫花苜蓿等的原花色素含量(verdieretal.,2012;escarayetal.,2014;yuanandgrotewold,2015;lietal.,2016),结果不理想,远远达不到干重2%的水平,因此不能够有效地防止牛、羊等反刍动物鼓胀病的发生。而与之形成鲜明对比的是,与原花色素共用合成途径的花青素的生物合成也主要受到mbw三元复合物(r2r3-myb(pap1/pap2/myb113/myb114)、bhlh(tt8/gl3/egl3)和wd40(ttg1))的调控,在拟南芥中过量表达pap1时,几乎全株植物都是紫色,积累大量花青素(borevitzetal.,2000),目前通过基因工程已经获得了富集花青素的西红柿、蒺藜苜蓿和紫花苜蓿等(butellietal.,2008;peeletal.,2009)。zhao等人和li等人认为无法获得比较好的原花色素的改良效果的最大障碍是原花色素生物合成和调控非常复杂,仍然有很多问题亟待回答(zhaoetal.,2010;lietal.,2016)。根据现有技术的研究结果,我们推测可能存在未知的关键基因负调控tt2或与之相关的三元复合物的功能,从而抑制了过量表达tt2基因促进原花色素积累的功能,导致过量表达tt2基因时不能够富集原花色素。
技术实现要素:
为了解决现有技术中仍不明确原花色素调控合成的缺陷,本发明提供了拟南芥at4g36920基因在调控植物原花色素生物合成及反刍动物抗臌胀病的应用,实现的目的为,发现拟南芥at4g36920基因在调控植物原花色素生物合成中的用途,为改良植物原花色素生物合成和代谢提供重要的理论支撑,从而得到原花色素含量高的植物品种,进一步提高反刍动物抗臌胀病的能力。
为了实现上述目的,本发明提供了拟南芥at4g36920基因在调控植物原花色素生物合成及反刍动物抗臌胀病的应用,本发明通过利用分子生物学、遗传学和代谢物定量分析的手段,确定拟南芥at4g36920基因(ap2)调控原花色素生物合成途径中的靶基因和关键转录因子,发现了拟南芥at4g36920基因(ap2)负调控植物原花色素的生物合成,通过抑制该基因的表达,增加植物原花色素的积累,以此将该基因应用在提高反刍动物抗臌胀病的用途。
本发明采用上述技术方案,有益效果包括:原花色素在植物的生长发育和逆境胁迫中起着重要的调控作用,对人类健康有诸多益处。诸多现有技术研究暗示存在未知的关键基因负调控tt2或与之相关的mbw三元复合物的功能,进而负调控原花色素的积累,但什么基因参与其中,及其调控的分子机理仍然不清楚。本发明利用转录组学和反向遗传学的方法,发现ap2突变体种皮颜色比野生型更深,积累更多的原花色素;ap2m3(编码的蛋白与ap2的相同,但其mrna不能够被mirna172结合)过量表达的种皮除珠孔和合点之外显著变浅,原花色素含量大幅下降;突变体和过量表达均不影响黄酮醇和花青素的积累。这些结果表明,ap2在珠孔和合点之外的种皮中特异地负调控原花色素的生物合成。本发明将利用分子生物学、遗传学和代谢物定量分析的手段,确定ap2调控原花色素生物合成途径中的靶基因和关键转录因子,阐明ap2调控原花色素生物合成的遗传网络,为改良植物原花色素生物合成和代谢提供重要的理论支撑。
附图说明
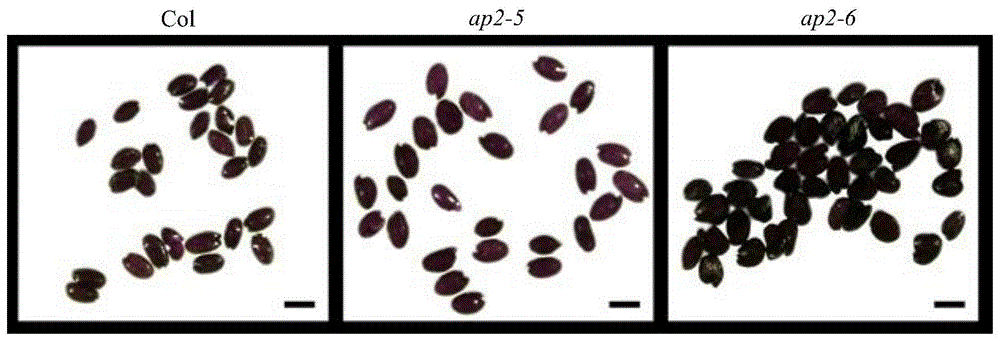
图1为ap2-5、ap2-6、野生型拟南芥成熟种子种皮经过dmaca染色后的图片。
具体实施方式
以下结合实施例对本发明作出进一步的说明。本发明中没有特殊说明的操作方法都是现有技术,没有特殊说明的操作条件也都是常温常压下,试剂均为市售自购。
实施例一:本发明提供的拟南芥at4g36920基因(ap2)在调控植物原花色素生物合成及反刍动物抗臌胀病的应用,拟南芥at4g36920基因(ap2)负调控植物原花色素的生物合成,通过抑制该基因的表达,增加植物原花色素的积累,以此将该基因应用在提高反刍动物抗臌胀病的用途。
本发明采用如下方法确认了拟南芥at4g36920基因(ap2)对于植物原花色素的生物合成所起到的负调控作用:
(1)获得了候选基因的纯合突变体:
根据原花色素在受精后1-6天积累的特点,我们获得了受精后0-6天荚果的基因表达数据,结合arabidopsisefpbrowser中的数据,我们筛选了近百个在种皮中特异表达或在种皮中表达较高的候选基因。然后,从abrc购买了突变体,鉴定并获得了纯合突变体。
(2)突变体表型分析:
我们采用反向遗传学的方法,分析候选基因纯合突变体的表型。由于原花色素生物合成途径中关键结构基因,如chs、chi和f3h和调控基因如tt2、tt8和tt16等的突变均导致种皮颜色变浅,我们首先分析了种皮颜色的变化。发现ap2-5(弱突变体)和ap2-6(强突变体)突变体种皮颜色比野生型变深。
因此ap2的突变可能增加了原花色素的积累,暗示ap2可能负调控种皮中原花色素的积累。由于原花色素和花青素的生物合成途径的绝大部分是相同的,既然ap2可能负调控原花色素的生物合成,它是否也调控花青素的生物合成。我们观察种子萌发后4天幼苗下胚轴的花青素积累情况,发现ap2-5和ap2-6突变体与野生型的表型相似。因此,推测ap2可能并不调控花青素的生物合成。
(3)ap2调控的类黄酮代谢产物分析
由于dmaca特异地与原花色素反应,dmaca染色后颜色的深浅与原花色素的含量成正比。我们进一步采用dmaca染色成熟的突变体种子,发现ap2-5和ap2-6的种子种皮染色后比野生型颜色更深(图1)。如图1所示,ap2-5和ap2-6的成熟种子种皮经过dmaca染色后比野生型颜色加深。bar为0.5mm。进一步表明ap2突变体种皮颜色变深,可能是由于ap2基因突变导致种皮中原花色素的积累增加,ap2很可能负调控原花色素的生物合成。
现有技术的研究表明,mirna172结合到ap2基因的mrna上抑制其翻译成蛋白,而ap2m3编码的蛋白与ap2的相同,但其mrna不能够被mirna172结合抑制其翻译。因此我们构建了ap2、ap2m3以及ap2m3融合myc标签的过量表达载体,转化拟南芥并获得纯合植株。首先,我们发现ap2m3和ap2m3融合myc标签的过量表达种子的种皮颜色显著变浅,而ap2过量表达种子的种皮颜色与野生型相似。然后,采用dmaca对成熟的突变体种子进行染色,发现染色3小时后,已经有22%的ap2-6种子的种皮颜色明显变深;当染色6小时后,已经有21%的ap2-5和51%的ap2-6种子的种皮颜色明显变深,而此时野生型仍然没有显著地被染色;当染色12小时后,ap2-5和ap2-6种子的种皮显著染色的比例分别超过94%和96%,而此时只有35%的野生型被显著染色;当染色24小时后,ap2-5和ap2-6均被显著染色,而野生型也只67%的被显著染色。染色12小时后,过量表达ap2基因的种子种皮被染色的比例竟然比野生型还高,但显著低于突变体;而其它染色时间,过量表达和野生型之间没有显著差异。总体来说,过量表达与野生型之间没有显著差异。现有技术的研究显示,mirna172结合到ap2基因的mrna上抑制其翻译成蛋白,使得35s::ap2的蛋白水平与野生型相比没有显著升高。因此,我们过量表达ap2基因的蛋白水平也可能与野生型相似,从而导致过量表达的种皮中原花色素的积累与野生型相似。除了珠孔和合点外,过量表达ap2m3和ap2m3融合myc标签的种子种皮在不同的染色时间均不能被显著染色。
最后,我们进一步定量测定了成熟种子中原花色素的含量,发现突变体中原花色素的积累显著地增加,而过量表达ap2m3的种子中原花色素含量大幅度地下降。这些结果清晰地表明,ap2基因在珠孔和合点之外的种皮中显著地抑制原花色素的生物合成。
由于ap2负调控原花色素的生物合成,它是否也会调控其它类黄酮(黄酮醇和花青素)的积累呢?我们观察发现,4天的幼苗中突变体与野生型的花青素积累没有明显变化,定量测定了4天幼苗和17天植株中的花青素含量,发现ap2-5、ap2-6突变体和ap2m3过量表达的花青素含量与野生型也没有显著差异。进一步表明ap2基因不调控花青素的生物合成。
由于黄酮醇主要在种皮和幼苗中积累,我们采用hplc的方法分析了黄酮醇在成熟种子和6天幼苗中的含量,发现ap2-5、ap2-6突变体和ap2m3过量表达植株的黄酮醇含量与野生型并没有显著差异。这些结果表明,ap2基因也不调控黄酮醇的生物合成。综上所述,ap2基因不调控黄酮醇和花青素的积累,而特异地在珠孔和合点之外的种皮中负调控原花色素的生物合成。
(4)为培育具有更高含量的原花色素,具有抗鼓胀病品质的植物新品种提供了可能。
本研究利用转录组学和反向遗传学的方法,发现ap2突变体种皮颜色比野生型更深,积累更多的原花色素,ap2基因负调控原花色素的积累;ap2m3(编码的蛋白与ap2的相同,但其mrna不能够被mirna172结合)过量表达的种皮除珠孔和合点之外显著变浅,原花色素含量大幅下降;突变体和过量表达均不影响黄酮醇和花青素的积累。这些结果表明,ap2在珠孔和合点之外的种皮中特异地负调控原花色素的生物合成。利用基因工程的手段,抑制ap2基因的表达,从而增加原花色素的积累,为培育具有更高含量的原花色素和具有抗鼓胀病品质的植物新品种提供了可能。
以上所述仅为本发明的优选实施例而已,并不用于限制本发明,对于本领域的技术人员来说,本发明可以有各种更改和变化。凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
- 还没有人留言评论。精彩留言会获得点赞!