一种四环素类抗生素降解基因tet(X)的优化及其在真核表达系统中的应用的制作方法
本发明属于基因工程技术领域,具体涉及一种四环素类抗生素降解基因tet(x)的优化及其在真核表达系统中的应用。
背景技术:
四环素类抗生素包括四环素、土霉素、金霉素等,是一类广谱抗生素,被广泛应用于人类及动物疾病的防治,同时也被作为饲料添加剂应用于禽畜养殖业和水产养殖业中。四环素类抗生素结构复杂,含并四苯骨架,属于难生物降解物质,很容易在环境中富集,在动物体内,四环素仅有部分被吸收并发生代谢,大部分以活性形式(母体或代谢产物)随粪便和尿液排出体外,进入生态环境。目前,四环素类抗生素在家畜和人群中均使用广泛,因其较低的吸收率导致四环素类抗生素的残留问题十分严峻,四环素类抗生素的残留对环境、家畜和人类健康发展均造成重大威胁。当前,降解四环素类抗生素的方式主要有以下两种:一是生物降解,包括微生物降解和植物降解;二是非生物降解,如水解、氧化降解及光降解等。
近期,四环素降解基因tet(x)新亚型的发现,为解决四环素的残留问题提供一些新思路。tet(x)可编码一种依赖于黄素的单加氧酶,该酶在o2、nadph、mgcl2的参与下可以高效降解四环素类抗生素。
随着分子生物学的发展,通过基因工程的方法,以微生物为生物容器来合成功能蛋白具有广阔的前景。而在众多的表达宿主中,原核表达产量低,且有可能形成包涵体,不利于功能蛋白的分离纯化,且会增加分离纯化的成本。毕赤酵母是最近迅速发展的一种真核表达宿主,营养要求不高,适合于高密度发酵的大规模生产方式,其分泌自身的蛋白很少,有利于表达产物的分离和纯化。
因此,通过分子生物学手段和遗传工程技术,有望通过基因tet(x)和毕赤酵母构建一种可降解四环素类抗生素的工程酵母,从而为四环素类抗生素的降解提供了一种新的途径。
技术实现要素:
为了克服上述现有技术的不足,本发明的首要目的是提供一种编码四环素类抗生素降解酶的优化基因tet(x)*。
本发明的第二个目的是提供一种能分泌四环素类抗生素降解酶tet(x)的工程酵母。
本发明的第三个目的是提供一种能分泌四环素类抗生素降解酶tet(x)的工程酵母的构建方法。
本发明的第四个目的是提供上述工程酵母在降解四环素类抗生素中的应用。
为实现上述目的,本发明所采用的技术方案为:
本发明提供一种编码四环素类抗生素降解酶的优化基因tet(x)*,该基因由tet(x)基因优化而来,优化后的tet(x)*适用于真核表达系统,该优化基因具有如seqidno:2所示的核苷酸优化序列。
tet(x)是一种依赖于黄素的单加氧酶,该酶在o2、nadph、mgcl2的参与下可以高效降解四环素类抗生素。本发明的编码四环素类抗生素降解酶的优化基因tet(x)*是在tet(x)原序列(如seqidno:1所示)的基础上进行优化后得到的,优化后的tet(x)*基因在真核表达体系中具有更高的表达水平。
优化的原则为:(1)替换表达宿主细胞的稀有密码子,尽可能使用宿主在蛋白翻译时的高频密码子;(2)为防止mrna提前终止转录,优化时避免产生at富含区;(3)尽可能提升序列的gc含量,有利于外源基因tet(x)*的表达;(4)避免优化序列时产生ecori、noti、sali酶切位点;(5)替换使mrna不稳定的序列和结构。
本发明还提供一种能分泌四环素类抗生素降解酶tet(x)的工程酵母,该工程酵母含有上述的优化基因tet(x)*。
四环素降解基因tet(x)可编码一种依赖于黄素的单加氧酶,该酶在o2、nadph、mgcl2的参与下可以高效降解四环素类抗生素。毕赤酵母是最近迅速发展的一种真核表达宿主,营养要求不高,适合于高密度发酵的大规模生产方式,其分泌自身的蛋白很少,有利于表达产物的分离和纯化。将从tet(x)优化后得到的tet(x)*基因导入到毕赤酵母中构建工程酵母,可以为四环素类抗生素的降解提供新的途径。
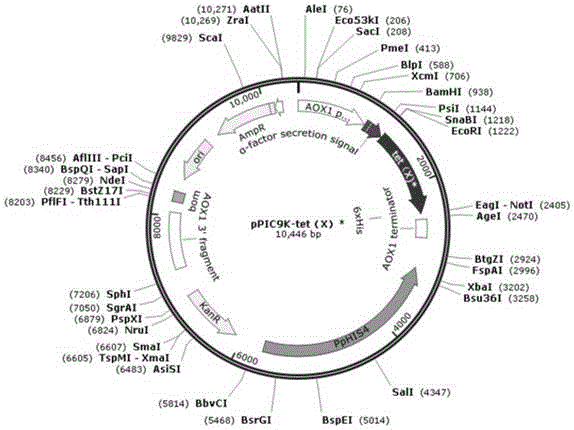
本发明还提供一种能分泌四环素类抗生素降解酶tet(x)的工程酵母的构建方法,以上述优化基因tet(x)*为目的基因,以ppic9k为载体,以毕赤酵母gs115为宿主,通过基因工程手段获得。
毕赤酵母gs115是真核表达系统常用的宿主,有着众多的技术优势。同原核表达相比,gs115可以对外源基因编码的蛋白质进行加工、折叠以及蛋白质修饰,且gs115染色体可以整合多个外源基因的拷贝数,从而在甲醇的严格诱导下能表达出更高水平的蛋白质。本发明选用ppic9k质粒作为载体主要有以下几个方面的考量:(1)ppic9k载体的his4可与毕赤酵母gs115的缺陷型his4发生同源重组,从而使酵母获得自身合成氨基酸的能力,便于使用无氨基酵母氮源的培养基筛选转化子;(2)ppic9k载体上的分泌肽信号可以诱导其信号肽下游的蛋白质分泌至宿主体外,而gs115自身分泌的胞外蛋白较少,故发酵液中的蛋白质主要为外源基因所编码的蛋白质,从而使构建得到的工程酵母表达系统具有初步纯化蛋白的效果。
优选的,上述工程酵母的构建方法,包括以下步骤:
s1、将优化基因tet(x)*插入到ppic9k质粒中,构建重组质粒;
s2、将毕赤酵母gs115制备成gs115感受态细胞(gs115感受态现制现用,避免冻融);
s3、将步骤s1的重组质粒转化到步骤s2的gs115感受态细胞中,筛选阳性转化子(电转完后,30℃静止培养1h后再涂布筛选)得到工程酵母。
优选的,在上述构建方法中,步骤s1的重组质粒在转化前先进行线性化处理,得到线性化重组质粒后再将其转化到步骤s2的gs115感受态细胞中。因为ppic9k-tet(x)*的完全线性化使得其更容易与gs115的染色体组上的基因发生同源重组。
具体的,上述线性化处理方法为:采用sali限制性内切酶对重组质粒进行单酶切。更优选的,步骤s1的重组质粒在线性化处理前需先转化到感受态细胞dh5α中进行验证和表达,以验证重组质粒是否构建成功,并使其满足后续的浓度需求。
优选的,在上述构建方法中,步骤s2的gs115感受态细胞在制备过程中使用了1m的dtt(二硫苏糖醇)。以提高感受态细胞的电转效率。同时,gs115感受态在制备时菌量要足够,制备后的感受态较粘稠,涂布至平板时可见一层薄膜。
优选的,在上述构建方法中,步骤s3采用电转化法将重组质粒转化到gs115感受态细胞中,电转化需要足够高的电压和足够长的放电时间,因此,本发明的电转化所采用的电击电压为1500-2000v,放电时间为4-6ms。
优选的,在上述构建方法中,重组质粒的构建方法为:将优化后的tet(x)*基因插入到ppic9k载体的ecori酶切位点和noti酶切位点之间。酶切位点的选择原则是:选择目的基因上没有,而质粒的多克隆位点上有的酶切位点,同时要避免切出平末端,并保证该酶切位点仅存在于多克隆位点区,避免切割质粒骨架其他开放阅读框。
优选的,在上述构建方法中,步骤s3所述筛选阳性转化子的原则为:先使用不含g418抗性的md空白板(即基础葡萄糖培养基,minimaldextrosemedium)筛选阳性转化子,待转化子在md空白板上长出后再用含g418的md筛选多拷贝的转化子,以得到功能更优的工程酵母。
具体的,高拷贝转化子在筛选时,先使用不含g418的md空白板筛选阳性转化子,待转化子在空白板上长出后再用含g418的md筛选多拷贝的转化子。gs115生长速度慢,需要足够的时间来积累g418的抗性,避免在获得足够抗性前就被杀死。
本发明还提供上述工程酵母在降解四环素类抗生素中的应用。
与现有技术相比,本发明的有益效果是:
本发明在tet(x)基因原序列的基础上经过优化得到一种编码四环素类抗生素降解酶的优化基因tet(x)*,并首次使用ppic9k载体将优化后的tet(x)*基因稳定的转入到毕赤酵母gs115中,构建得到工程酵母gs115-ppic9k-tet(x)*,该工程酵母具有表达tet(x)蛋白的能力。通过工程酵母的表达效果试验可见,构建后的工程酵母gs115-ppic9k-tet(x)*,其外源基因表达的蛋白质在nadph、mg2+、o2的参与下表现出生物功能,对四环素类抗生素具有高效的降解作用。
附图说明
图1为tet(x)*优化序列插入位点图谱;
图2为转化子dh5α-ppic9k-tet(x)*的电泳图谱;m:dl2000marker;1-9:转化子;10:阴性对照;
图3为线性化重组载体ppic9k-tet(x)*和线性化空载体ppic9k的sali单酶切电泳图谱;m:dl10000marker;1:线性化重组载体ppic9k-tet(x)*;2:线性化空载体ppic9k;
图4为gs115-ppic9k-tet(x)*转化子和gs115-ppic9k转化子在md培养基上的单菌落;
图5为高拷贝转化子的形成机制;
图6为gs115-ppic9k-tet(x)*转化子在含g418的md平板上的生长情况;
图7为转化子gs115-ppic9k-tet(x)*和转化子gs115-ppic9k的基因组重组分析图谱;m:dl5000marker;1-10:gs115-ppic9k-tet(x)*;11-15:gs115-ppic9k;16:阴性对照;
图8为工程酵母gs115-ppic9k-tet(x)*对四环素类抗生素的降解效果;a组为4ug/mleravacycline的降解试验;b组为16ug/mltetracycline的降解试验;1:药物;2:药物+发酵液;3:药物+nadph、mg2+、o2;4、药物+发酵液+nadph、mg2+、o2。
具体实施方式
下面对本发明的具体实施方式作进一步说明。在此需要说明的是,对于这些实施方式的说明用于帮助理解本发明,但并不构成对本发明的限定。此外,下面所描述的本发明各个实施方式中所涉及的技术特征只要彼此之间未构成冲突就可以相互组合。
下述实验例中所使用的试验方法如无特殊说明,均为常规方法;所使用的材料、试剂等,如无特殊说明,为可从商业途径得到的试剂和材料。
实施例1构建大肠杆菌dh5α-ppic9k-tet(x)*转化子
(1)基因tet(x)的序列优化
由于真核生物和原核生物在蛋白质的翻译过程中对密码子的偏好性不同。为了提高tet(x)在毕赤酵母gs115中的表达水平,首先对tet(x)的原序列(originaldna,如seqidno:1所示)进行优化,此外,为方便后续研究,在终止密码子前添加6xhis标签序列,得到最优序列,即优化后的降解基因tet(x)*(optimizeddna,如seqidno:2所示)。
(2)优化基因tet(x)*的载体连接以及感受态细胞的转化
将优化后的基因tet(x)*插入到ppic9k载体的ecori酶切位点和noti酶切位点之间(见图1),得到重组质粒ppic9k-tet(x)*。然后将感受态细胞dh5α冰浴5-10min使其冻融,加入500ng重组质粒ppic9k-tet(x)*,轻轻混匀,继续冰浴30min,随后42℃水浴热激60s,冰浴3min,再加入900ulsob,37℃、180rpm培养1h,将培养液稀释10-2倍后涂布于含amp100(浓度为100ug/ml的氨苄青霉素)的药板上37℃培养16-18h,待菌落长出后随机挑取9个单菌落对插入片段进行pcr扩增(以单克隆为dna模板)。
pcr扩增的引物为:上游引物(pic9k-f)的核苷酸序列如seqidno:3所示,下游引物(pic9k-r)的核苷酸序列如seqidno:4所示。
pic9k-f(5’-3’):aggatccaaacgatgagatttc;
pic9k-r(5’-3’):cgaggatgacgatgagcgcat。
pcr扩增的体系(25μl)为:双纯水19.375μl,10×pcrbuffer2.5μl,dntps2μl,上游引物0.5μl,下游引物0.5μl,rtaqdna聚合酶0.125μl。
pcr扩增的反应条件为:94℃预变性5min;94℃变性45s,58℃退火45s,72℃延伸2min,共30个循环;72℃终延伸10min;4℃保存。
pcr结束后,pcr产物经1.2%琼脂糖凝胶电泳鉴定,均扩增出一条约1900bp的条带(见图2),表明重组质粒ppic9k-tet(x)*成功转化至感受态细胞dh5α中,得到转化子dh5α-ppic9k-tet(x)*。将阳性转化子保存于30%甘油肉汤中,置于-80℃备用。
实施例2构建工程酵母gs115-ppic9k-tet(x)*
(1)重组质粒ppic9k-tet(x)*的线性化
复苏dh5α-ppic9k-tet(x)*,参照天根质粒小提试剂盒的说明书提取dh5α-ppic9k-tet(x)*的质粒,使用sali限制性内切酶(takara)对浓度及纯度合格的质粒进行单酶切,同时以未插入优化序列tet(x)*的ppic9k为空载对照。
sali单酶切体系为:dna1μg,10×h(缓冲液)2μl,sali1μl,加双纯水至总体积为20μl。
37℃过夜酶切后,参照takaradna片段纯化试剂盒说明书回收酶切产物,并将回收后的产物进行1%琼脂糖凝胶电泳,剩余产物-80℃保存备用。胶图(图3)显示,ppic9k-tet(x)*重组质粒经单酶切后得到一条约10446bp的条带,ppic9k经酶切后得到一条约9276bp的条带,表明ppic9k-tet(x)*重组质粒已线性化。
(2)gs115感受态的制备
从-80℃低温冰箱中取出gs115(甘油保菌),用ypd固体培养基复苏,挑取单菌落接种于4mlypd液体培养基中,30℃、220rpm过夜培养,吸取1ml过夜培养液至含50mlypd液体培养基的锥形瓶中,30℃、220rpm培养至od600=1.3左右,收集菌液于50ml离心管中,冰浴20min,4℃、1500g离心5min,弃上清液后加入20ml新鲜的ypd液体培养基和0.5ml1m的dtt,30℃、220rpm培养20min,4℃、1500g离心5min,弃上清液,加40ml预冷的双纯水洗涤菌体沉淀,4℃、1500g离心5min,加30ml预冷的双纯水洗涤菌体沉淀,4℃、1500g离心5min,加4ml预冷的1m山梨醇洗涤菌体沉淀,4℃、1500g离心5min,加200μl预冷的1m山梨醇重悬菌体沉淀,分装,每管80μl,得到gs115感受态细胞,置于冰上备用,多余的感受态-80℃保存。
(3)电转线性化ppic9k-tet(x)*重组质粒
取2-3μg线性化ppic9k-tet(x)*重组质粒与感受态细胞混匀,同时以ppic9k空载为对照,混匀后继续冰浴5min,吸取混合液加至预冷的0.2cm电转杯,轻轻拍打铺平杯底,排除气泡,使用bio-rad电转仪电击,电压1800v,电转时间以4-6ms为宜,取出电转杯加入1ml预冷的1m山梨醇吹打混匀,将混悬液吸至1.5ml离心管中,30℃静止培养1h后涂布md平板,平板放置在30℃恒温培养箱培养3-4d可见md培养基上长出白色单菌落(见图4),即gs115-ppic9k-tet(x)*转化子。
(4)高拷贝转化子的筛选
为提高tet(x)蛋白的表达量,选用载体质粒上kanr基因赋予gs115的g418抗性进行转化子筛选。随着插入到宿主gs115染色体组中的kanr越多,宿主对g418的抗性就越高,同时随着kanr一起以表达盒形式插入的tet(x)*拷贝数也越多(多拷贝转化子形成机制如图5所示),因此分别选用含0.2mg/ml、0.5mg/ml、1mg/ml、2mg/ml、3mg/ml、4mg/ml、6mg/ml、8mg/mlg418的md平板筛选转化子。
用无菌牙签从电转后涂布的md平板上挑取一个转化子,依次点接种于含0.2mg/ml、0.5mg/ml、1mg/ml、2mg/ml、3mg/ml、4mg/ml、6mg/ml、8mg/mlg418的md平板同一位置,最后再接种于空白的md平板上;换一根牙签重新挑取一个转化子重复上述接种操作;接种好的md平板置于30℃下培养4-5d。
共计挑取52个转化子,52个转化子均为多拷贝,0.2mg/ml、0.5mg/ml、1mg/ml、2mg/ml、3mg/ml、4mg/ml、6mg/ml平板均生长良好,8mg/ml平板上出现明显的生长差异(见图6)。选取10个8mg/ml平板生长良好的转化子和电转后直接涂布md平板上的5个空载进一步分析,参照天根酵母基因组提取试剂盒说明书提取上述15个菌株的全基因组,使用pcr分析这15株菌的基因整合情况,筛除muts(甲醇利用缓慢型)的菌株。
pcr扩增的引物为:上游引物(aox1-f)的核苷酸序列如seqidno:5所示,下游引物(aox1-r)的核苷酸序列如seqidno:6所示。
aox1-f(5’-3’):gactggttccaattgacaagc;
aox1-r(5’-3’):gcaaatggcattctgacatcc。
pcr扩增的体系(25μl)为:双纯水18.375μl,10×pcrbuffer2.5μl,dntps2μl,上游引物0.5μl,下游引物0.5μl,rtaqdna聚合酶0.125μl,dna模板1μl。
pcr扩增的条件为:94℃预变性5min;94℃变性45s,58℃退火45s,72℃延伸2min,共30个循环;72℃终延伸10min;4℃保存。
pcr产物经1.2%琼脂糖凝胶电泳鉴定,如图7所示,ppic9k-tet(x)*转化子均扩增出两条带,一条为编码aox1基因的片段(约2200bp),一条为ppic9k-tet(x)*上外源基因的片段(约1600bp)。ppic9k空载转化子均扩增出两条带,一条为编码aox1基因的片段(约2200bp),一条为ppic9k上的片段(约500bp)。表明外源基因正确插入到gs115的染色体组中,未破坏宿主自身的aox1基因,工程酵母gs115-ppic9k-tet(x)*构建成功。用15%甘油肉汤保存gs115-ppic9k-tet(x)*和gs115-ppic9k阳性转化子,-80℃备用。
实施例3工程酵母gs115-ppic9k-tet(x)*的表达效果验证
以嗜热菌为受试菌,采用打孔法分别进行eravacycline(依拉瓦环素)降解试验和tetracycline(四环素)降解试验,每个试验均分为4组:1、药物组;2、药物+发酵液组;3、药物+nadph、mg2+、o2组;4、药物+发酵液+nadph、mg2+、o2组。以上4组均在37℃孵育5h后吸取20ul体系滴在涂布有嗜热菌的对应孔径内,60℃正放过夜培养后观察抑菌圈大小。
结果如图8所示,在上述四组试验组中,只有药物+发酵液+nadph、mg2+、o2组没有出现明显的抑菌圈,说明在nadph、mg2+、o2的参与下,构建后的工程酵母发酵液对四环素类抗生素具有降解的作用,使嗜热菌表现出一定的生长活性;原因是构建后的工程酵母的外源基因得到了有效的表达,能向外分泌依赖于黄素的单加氧酶tet(x),该酶在o2、nadph、mgcl2的参与下可以高效降解四环素类抗生素。可见,本发明构建的工程酵母gs115-ppic9k-tet(x)*,其外源基因表达的蛋白质在nadph、mg2+、o2的参与下表现出生物功能,对四环素类抗生素具有高效的降解作用。
以上对本发明的实施方式作了详细说明,但本发明不限于所描述的实施方式。对于本领域的技术人员而言,在不脱离本发明原理和精神的情况下,对这些实施方式进行多种变化、修改、替换和变型,仍落入本发明的保护范围内。
序列表
<110>华南农业大学
<120>一种四环素类抗生素降解基因tet(x)的优化及其在真核表达系统中的应用
<141>2020-01-06
<160>6
<170>siposequencelisting1.0
<210>2
<211>1158
<212>dna
<213>2ambystomalateralexambystomajeffersonianum
<400>2
atgagcaataaagaaaaacaaatgaatttacttagtgataagaacgttgcaataattggt60
ggtggacccgttggactgactatggcaaaattattacagcaaaacggcatagacgtttca120
gtttacgaaagagacaacgaccgagaggcaagaatttttggtggaacccttgacctacac180
aaaggttcaggtcaggaagcaatgaaaaaagcgggattgttacaaacttattatgactta240
gccttaccaatgggtgtaaatattgctgatgaaaaaggcaatattttatccacaaaaaat300
gtaaagcccgaaaatcgatttgacaatcctgaaataaacagaaatgacttaagggctatc360
ttgttgaatagtttagaaaacgacacggttatttgggatagaaaacttgttatgcttgaa420
cctggtaagaagaagtggacactaacttttgagaataaaccgagtgaaacagcagatctg480
gttattattgccaatggtggaatgtctaaagtaagaaaatttgttaccgacacggaagtt540
gaagaaacaggtactttcaatatacaagccgatattcatcatccagaggtgaactgtcct600
ggattttttcagctatgcaatggaaaccggctaatggctgctcatcaaggtaatttatta660
tttgcgaatcctaataataatggtgcattgcattttggaataagttttaaaacacctgat720
gaatggaaaaaccaaacgcaggtagattttcaaaacagaaatagtgtcgttgattttctt780
ctgaaagaattttccgattgggacgaacgctacaaagaactgattcgtgtgacatcatct840
tttgtagggttagcgacacgaatatttcccttaggtaagtcttggaaaagtaagcgtcca900
ttacccataacgatgattggagatgctgctcatttgatgcctccttttgcaggacaaggc960
gtaaacagcgggttgatggatgccttgatattgtcggataatctgaccaatgggaaattt1020
aacagcattgaagaggctattgaaaattatgaacagcaaatgtttatctatggcaaagaa1080
gcacaagaagaatcaactcaaaacgaaattgaaatgtttaaacccgactttacgtttcag1140
caattgttaaatgtataa1158
<210>2
<211>1176
<212>dna
<213>2ambystomalateralexambystomajeffersonianum
<400>2
atgtccaacaaggagaagcaaatgaacttgttgtctgataagaacgttgctattattggt60
ggtggtccagttggtttgaccatggctaagttgttgcaacaaaacggtattgacgtttca120
gtttacgaaagagacaacgacagagaagctagaatttttggtggtactttggatttgcat180
aagggttctggtcaggaagctatgaagaaggctggattgttgcagacctattatgatttg240
gctttgccaatgggtgttaacattgctgatgagaagggtaatattttgtctactaagaac300
gttaagccagaaaacagatttgataacccagagattaacagaaacgacttgagagctatt360
ttgttgaactccttggaaaacgatactgttatctgggatagaaagttggttatgttggag420
ccaggtaagaaaaagtggacattgacttttgaaaacaagccatctgagacagctgatttg480
gttatcattgctaacggtggtatgtctaaggttagaaagtttgttactgacactgaagtt540
gaagaaacaggtacttttaacattcaggctgacatccatcatccagaagttaattgtcca600
ggtttttttcagttgtgtaatggtaacagattgatggctgctcaccaaggaaacttgttg660
tttgctaacccaaacaacaacggtgctttgcactttggtatttcttttaagactccagat720
gagtggaaaaaccagactcaagttgactttcaaaatagaaactctgttgttgatttcttg780
ttgaaggaattttctgactgggatgagagatacaaggaattgattagagttacctcttct840
tttgttggattggctactagaatttttccattgggtaagtcttggaagtctaagagacct900
ttgccaattactatgattggtgatgctgctcatttgatgccaccatttgctggtcaaggt960
gttaactctggtttgatggatgctttgattttgtctgataatttgactaacggtaagttt1020
aactctatcgaagaggctattgaaaattacgagcagcaaatgtttatctatggtaaggaa1080
gctcaagaggagtctactcaaaacgagattgaaatgttcaagccagactttacttttcaa1140
caattgttgaatgttcaccatcaccatcatcactaa1176
<210>3
<211>22
<212>dna
<213>2ambystomalateralexambystomajeffersonianum
<400>3
aggatccaaacgatgagatttc22
<210>4
<211>21
<212>dna
<213>2ambystomalateralexambystomajeffersonianum
<400>4
cgaggatgacgatgagcgcat21
<210>5
<211>21
<212>dna
<213>2ambystomalateralexambystomajeffersonianum
<400>5
gactggttccaattgacaagc21
<210>6
<211>21
<212>dna
<213>2ambystomalateralexambystomajeffersonianum
<400>6
gcaaatggcattctgacatcc21
- 还没有人留言评论。精彩留言会获得点赞!