一种双酶级联催化合成(R)-3-氨基-1-丁醇的方法
本发明属于生物
技术领域:
,具体涉及一种双酶级联催化合成(r)-3-氨基-1-丁醇的方法,尤其涉及一种利用醇脱氢酶和胺脱氢酶级联催化合成手性(r)-3-氨基-1-丁醇的方法。
背景技术:
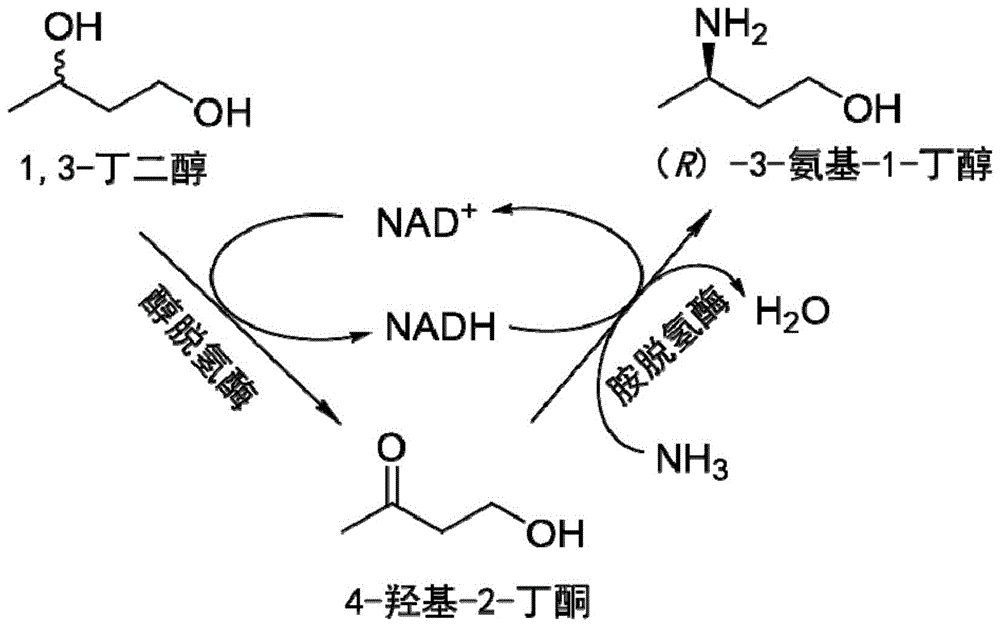
:手性胺醇化合物是许多生物活性分子的结构单元,是重要的医药和精细化工中间体,其中(r)-3-氨基-1-丁醇是合成抗艾滋病整合酶抑制剂度鲁特韦(dolutegravir)的重要前体,并且还可衍生为β-内酰胺,用于合成青霉烯类抗生素。(r)-3-氨基-1-丁醇主要是通过化学法合成,酶法合成的报道较少。1995年gertzmann等以手性(r)-丙氨酸为原料,经氨基保护后,以重氮甲烷增长碳链变为β-氨基酸酯,脱保护后还原得到目标产物。1977年kinas等用巴豆酸酯与(r)-苯乙胺反应生成具有两个手性中心的一组差向异构体,通过硅胶柱层析分离后得到单一的异构体,然后经过酯还原、脱苄基得到(r)-3-氨基-1-丁醇;该方法步骤较少且原料易得,是一种有希望实现工业化生产的方法,但是存在以下问题,由于第一步反应选择性较差,得到几乎等量的两个対映异构体,造成分离纯化比较困难,常采用色谱柱法分离,洗脱剂用量大、损失大、效率低,同时由于使用了价格昂贵的lialh4作为还原剂,原材料成本也显著上升,该方法不宜规模化工业化生产。1998年,tatsuya等报道了一种合成(r)-3-氨基-1-丁醇的方法,但该方法中原料的价格昂贵,反应条件苛刻。目前,(r)-3-氨基-1-丁醇的合成方法中,化学法需要高温、高压和金属催化剂、反应条件苛刻、污染大、安全系数低,收率为60%~70%。生物法反应条件温和,但转化率低。因此,寻找新的绿色方法来合成(r)-3-氨基-1-丁醇,对促进现有化学工艺的绿色升级和加强对环境的保护具有重要的科学价值和社会意义。1,3-丁二醇是一种重要的化工原料,具有易于获得、价格低等特点,用于(r)-3-氨基-1-丁醇的合成原料,具有很好的经济效益。技术实现要素:本发明的目的是如何合成手性(r)-3-氨基-1-丁醇。本发明首先保护合成手性(r)-3-氨基-1-丁醇的方法。本发明所保护的合成手性(r)-3-氨基-1-丁醇的方法,具体可为方法一,可包括如下步骤:(a)以1,3-丁二醇为底物,经酶a催化反应生成4-羟基-2-丁酮;(b)以步骤(a)生成的4-羟基-2-丁酮为底物,经酶b催化反应生成手性(r)-3-氨基-1-丁醇;所述酶a选自如下任一种:醇脱氢酶、醇脱氢酶突变体;所述酶b选自如下任一种:胺脱氢酶、胺脱氢酶突变体。本发明所保护的合成手性(r)-3-氨基-1-丁醇的方法,具体可为方法二,可包括如下步骤:以1,3-丁二醇为底物,经酶a和酶b催化反应,生成手性(r)-3-氨基-1-丁醇;所述酶a选自如下任一种:醇脱氢酶、醇脱氢酶突变体;所述酶b选自如下任一种:胺脱氢酶、胺脱氢酶突变体。上述任一所述醇脱氢酶可为如下(a1)-(a42)中任一。(a1)来源于短小乳杆菌(lactobacillusbreviskb290)的醇脱氢酶adh1,氨基酸序列的accessionnumber为ban05992.1。(a2)来源于高温厌氧杆菌(thermoanaerobacterbrockii)的醇脱氢酶adh2,氨基酸序列的accessionnumber为caa46053.1。(a3)来源于固氮弧菌属(aromatoleumaromaticumebn1)的醇脱氢酶adh3,氨基酸序列的accessionnumber为cai07428.1。(a4)来源于链霉菌(streptomycescoelicolor)的醇脱氢酶adh4,氨基酸序列的accessionnumber为q9kym4。(a5)来源于枯草芽孢杆菌(bacillussubtilis)的醇脱氢酶adh5,氨基酸序列的accessionnumber为wp_003234015.1。(a6)来源于肺炎克雷白杆菌(klebsiellapneumoniae)的醇脱氢酶adh6,氨基酸序列的accessionnumber为wp_015958558.1。(a7)来源于葡萄糖杆菌(gluconobacteroxydans)的醇脱氢酶adh7,氨基酸序列的accessionnumber为wp_011253549.1。(a8)来源于赖氏菌属(leifsoniasp.s749)的醇脱氢酶adh8,氨基酸序列的accessionnumber为bad99642.1。(a9)来源于赤红球菌(rhodococcusruber)的醇脱氢酶adh9,氨基酸序列的accessionnumber为q8klt9。(a10)来源于高加索酸奶乳杆菌(lactobacilluskefiridsm20587)的醇脱氢酶adh10,氨基酸序列的accessionnumber为aap94029.1。(a11)来源于威吉利热厌氧杆菌(thermoanaerobacterwiegeliirt8.b1)的醇脱氢酶adh11,氨基酸序列的accessionnumber为aem77517.1。(a12)来源于乳杆菌(lactobacilluszymaedsm19395)的醇脱氢酶adh12,氨基酸序列的accessionnumber为krl08675.1。(a13)来源于酒球菌(oenococcusalcoholitolerans)的醇脱氢酶adh13,氨基酸序列的accessionnumber为kgo31568.1。(a14)来源于愈伤热球菌(thermococcusguaymasensis)的醇脱氢酶adh14,氨基酸序列的accessionnumber为adv18977.1。(a15)来源于肉毒梭菌(clostridiumbotulinum)的醇脱氢酶adh15,氨基酸序列的accessionnumber为wp_003399463.1。(a16)来源于致黑脱硫肠状菌(desulfotomaculumnigrificans)的醇脱氢酶adh16,氨基酸序列的accessionnumber为wp_003542410.1。(a17)来源于一氧化碳嗜热窦菌(thermosinuscarboxydivorans)的醇脱氢酶adh17,氨基酸序列的accessionnumber为wp_007290608.1。(a18)来源于烷嗜热厌氧杆菌(thermoanaerobactermathranii)的醇脱氢酶adh18,氨基酸序列的accessionnumber为wp_013150923.1。(a19)来源于梭菌属(clostridium)的醇脱氢酶adh19,氨基酸序列的accessionnumber为wp_013239134.1。(a20)来源于厚壁菌门细菌(firmicutesbacteriumcag:137)的醇脱氢酶adh20,氨基酸序列的accessionnumber为cdb31037.1。(a21)来源于拜氏梭菌(clostridiumbeijerinckii)的醇脱氢酶adh21,氨基酸序列的accessionnumber为wp_026889046.1。(a22)来源于梭菌属(clostridium)的醇脱氢酶adh22,氨基酸序列的accessionnumber为wp_039771361.1。(a23)来源于乙草甲烷菌(methanosarcinaacetivorans)的醇脱氢酶adh23,氨基酸序列的accessionnumber为wp_048065256.1。(a24)来源于淡色甲烷菌(methanosarcinalacustris)的醇脱氢酶adh24,氨基酸序列的accessionnumber为wp_048125376.1。(a25)来源于甲烷八叠球菌(methanosarcinathermophila)的醇脱氢酶adh25,氨基酸序列的accessionnumber为wp_048166386.1。(a26)来源于西洋甲烷菌(methanosarcinasiciliae)的醇脱氢酶adh26,氨基酸序列的accessionnumber为wp_048172170.1。(a27)来源于德氏葡萄球菌(desnuesiellamassiliensis)的醇脱氢酶adh27,氨基酸序列的accessionnumber为wp_055668884.1。(a28)来源于卡尔德氏杆菌(caldanaerobactersubterraneus)的醇脱氢酶adh28,氨基酸序列的accessionnumber为kuk09008.1。(a29)来源于钟氏梭菌(clostridiumljungdahlii)的醇脱氢酶adh29,氨基酸序列的accessionnumber为wp_063556461.1。(a30)来源于拜氏梭菌(clostridiumbeijerinckii)的醇脱氢酶adh30,氨基酸序列的accessionnumber为wp_065417405.1。(a31)来源于未培养梭菌(unculturedclostridiumsp.)的醇脱氢酶adh31,氨基酸序列的accessionnumber为sci81347.1。(a32)来源于丝状孢子梭菌(clostridiumtaeniosporum)的醇脱氢酶adh32,氨基酸序列的accessionnumber为wp_069679756.1。(a33)来源于脱硫弧菌(desulfovibriolitoralis)的醇脱氢酶adh33,氨基酸序列的accessionnumber为wp_072695399.1。(a34)来源于脱硫肠状菌(desulfotomaculumputei)的醇脱氢酶adh34,氨基酸序列的accessionnumber为wp_073236284.1。(a35)来源于拜氏梭菌(clostridiumbeijerinckii)的醇脱氢酶adh35,氨基酸序列的accessionnumber为wp_077844196.1。(a36)来源于梭状芽胞杆菌(clostridiumpuniceum)的醇脱氢酶adh36,氨基酸序列的accessionnumber为wp_077846831.1。(a37)来源于二醇梭菌(clostridiumdiolis)的醇脱氢酶adh37,氨基酸序列的accessionnumber为wp_087701616.1。(a38)来源于匙形梭菌(clostridiumcochlearium)的醇脱氢酶adh38,氨基酸序列的accessionnumber为wp_089865926.1。(a39)来源于笃斯梭菌(clostridiumuliginosum)的醇脱氢酶adh39,氨基酸序列的accessionnumber为wp_090093236.1。(a40)来源于梭菌属细菌(clostridialesbacteriumsk-y3)的醇脱氢酶adh40,氨基酸序列的accessionnumber为wp_094550774.1。(a41)来源于近平滑假丝酵母(clostridiumbotulinum)的醇脱氢酶adh41,氨基酸序列的accessionnumber为wp_096043277.1。(a42)在(a1)-(a41)中任一所限定的蛋白质的n端和/或c端连接标签后得到的融合蛋白。上述任一所述胺脱氢酶可为如下(b1)-(b10)中任一。(b1)来源于嗜热脂肪土芽孢杆菌(geobacillusstereothermophilus)的胺脱氢酶amdh1,氨基酸序列为seqidno.1。(b2)来源于嗜热脂肪芽孢杆菌(bacillusstereothermophilus)的胺脱氢酶amdh2,氨基酸序列为seqidno.2。(b3)来源于嗜冷芽孢八叠球菌(sporosarcinapsychrophila)的胺脱氢酶amdh3,氨基酸序列为seqidno.3。(b4)来源于球形赖氨酸芽孢杆菌(lysinibacillussphaericus)的胺脱氢酶amdh4,氨基酸序列为seqidno.4。(b5)来源于西伯利亚微杆菌(exiguobacteriumsibiricum)的胺脱氢酶的胺脱氢酶amdh5,氨基酸序列为seqidno.5。(b6)来源于普通高温放线菌(thermoactinomycesintermedius)的胺脱氢酶amdh6,氨基酸序列为seqidno.6。(b7)来源于温泉热碱芽孢杆菌(caldalkalibacillusthermarum)的胺脱氢酶amdh7,氨基酸序列为seqidno.7。(b8)来源于糖莱斯氏菌(laceyellasacchari)的胺脱氢酶amdh8,氨基酸序列为seqidno.8。(b9)来源于栗褐芽孢杆菌(bacillusbadius)的胺脱氢酶amdh9,氨基酸序列为seqidno.9。(b10)在(b1)-(b9)中任一所限定的蛋白质的n端和/或c端连接标签后得到的融合蛋白。上述任一所述胺脱氢酶的突变体可为(c1)或(c2)。(c1)将seqidno.3所示的胺脱氢酶amdh3的第114位和第291位依次突变为缬氨酸残基和半胱氨酸残基。(c2)将seqidno.3所示的胺脱氢酶amdh3的第111位、第114位、第294位依次突变为苯丙氨酸残基、缬氨酸残基和半胱氨酸残基。上述任一所述的方法中,所述酶a和所述酶b均是以粗酶液、粗酶粉、纯酶或全细胞的形式发生催化作用的。上述任一所述的方法中,所述经酶a和酶b催化反应可为经酶a和酶b共表达全细胞催化反应。上述任一所述的方法中,所述粗酶液、粗酶粉和/或纯酶均按照包括如下步骤的方法制备得到:在宿主细胞中表达所述酶a和/或所述酶b,得到重组细胞;裂解所述重组细胞获得所述粗酶液、粗酶粉或纯酶。上述任一所述的方法中,所述全细胞均按照包括如下步骤的方法制备得到:在宿主细胞中表达所述酶a和/或所述酶b,得到的重组细胞即为所述全细胞。所述重组细胞是按照包括如下步骤的方法制备获得的:向所述宿主细胞到导入能够表达所述酶a和/或所述酶b的核酸分子,经诱导培养后获得表达所述酶a和/或所述酶b的所述重组细胞。所述“能够表达所述酶a和/或所述酶b的核酸分子”是通过重组载体的形式导入到所述宿主细胞中的。所述重组载体可为携带有所述酶a和/或所述酶b的编码基因的细菌质粒、噬菌体、酵母质粒或逆转录病毒包装质粒。所述宿主细胞可为原核细胞或低等真核细胞。所述原核细胞可为细菌。所述低等真核细胞可为酵母细胞。所述细菌可为大肠杆菌。所述重组载体具体为将所述醇脱氢酶和/或所述胺脱氢酶或其突变体的编码基因替换pet28a载体的酶切位点ncoⅰ和xhoⅰ之间的小片段后得到的重组质粒。上述任一所述的方法中,所述催化反应的温度可为25~40℃(具体如30℃)。所述催化反应的时间可为4~48h(具体如24h)。上述任一所述的方法中,当所述酶a以全细胞、粗酶液、粗酶粉或纯酶的形式进行步骤(a)时,催化反应在如下(d1)所示缓冲液中进行。当所述酶b以全细胞、粗酶液、粗酶粉或纯酶的形式进行步骤(b)时,催化反应在如下(d2)所示缓冲液中进行。当酶a和酶b共表达全细胞催化反应时,催化反应在如下(d2)所示缓冲液中进行。(d1)浓度为50~100mm,ph值为6.5~8.5的磷酸盐缓冲液。(d2)浓度为50mm~4m,ph值为7.0~9.5的氯化铵/氨水缓冲液。(d1)具体可为浓度为100mm,ph值为8.0的磷酸盐缓冲液。(d2)具体可为浓度为1m,ph值为8.0的氯化铵/氨水缓冲液。在本发明的一个实施例中,所述宿主细胞具体为大肠杆菌,更加具体的为e.colibl21(de3)。相应的,所述诱导培养为向培养体系中加iptg至终浓度0.1-0.5mm(具体如0.1mm),20-37℃(具体如20℃)诱导培养12-24h(具体如16h)。在本发明中,所述酶a的辅酶具体可为氧化型辅酶i(即nad+)或氧化型辅酶ii(即nadp+)。所述酶b的辅酶具体为nadh或nadph。所述1,3-丁二醇在所述反应体系中浓度可为1-100mm(如20mm)。所述重组细胞在所述反应体系中的浓度可为50-500g/l(如100g/l)。所述粗酶液或粗酶粉在所述反应体系中的浓度可为10-50g/l(如20g/l)。所述纯酶在所述反应体系中的浓度可为0.1-2g/l(如0.5g/l)。所述nad+在所述反应体系中的浓度可为0.1-2.0mm(如1.0mm)。所述醇脱氢酶进行步骤(a)所述的催化反应体系组成如下:浓度为100mm磷酸盐缓冲液(ph值为8.0),终浓度为1-10mm的1,3-丁二醇,终浓度为1mm的nad+或nadp+,终浓度为2%的丙酮(v/v),终浓度为10-40g/l的所述醇脱氢酶的粗酶液或粗酶粉。所述胺脱氢酶或其突变体进行步骤(b)所述的催化反应体系组成如下:浓度为1m的氯化铵/氨水缓冲液(ph值为8.5),终浓度为1-20mm的4-羟基-2-丁酮,终浓度为2g/l的gdh粗酶粉,终浓度为100mm的葡萄糖,终浓度为1mm的nad+,终浓度为100g/l的所述胺脱氢酶或其突变体的全细胞。当醇脱氢酶(或醇脱氢酶突变体)和胺脱氢酶(或胺脱氢酶突变体)全细胞催化反应的组成如下:浓度为1m的氯化铵/氨水缓冲液(ph值为8.0)、终浓度为20mm的1,3-丁二醇、终浓度为1mm的即nad+、终浓度为1g/l的溶菌酶、终浓度为6u/ml的dnasei(脱氧核糖核酸酶)、终浓度为200g/l醇脱氢酶(或醇脱氢酶突变体)和胺脱氢酶(或胺脱氢酶突变体)共表达全细胞。本发明还保护酶系统或其相关产品。所述酶系统包括上述任一所述酶a和所述酶b。所述相关产品可为能够表达所述酶系统中各酶的核酸分子,或含有所述核酸分子的表达盒、重组载体、重组菌或转基因细胞系。所述酶系统或其相关产品在合成手性(r)-3-氨基-1-丁醇中的应用也属于本发明的保护范围。本发明所提供的合成手性(r)-3-氨基-1-丁醇的方法中,存在辅因子自循环系统。所述辅因子再生系统为所述酶a催化1,3-丁二醇氧化成4-羟基-2-丁酮,nad+被还原成nadh;所述酶b催化4-羟基-2-丁酮生成(r)-3-氨基-1-丁醇,nadh被氧化成nad+,生成的nad+重新参与到1,3-丁二醇生成4-羟基-2-丁酮的氧化。本发明提供了一条全新的绿色生物合成路线,以廉价的1,3-丁二醇为原料,通过双酶共表达催化合成手性(r)-3-氨基-1-丁醇。本发明操作方便,具有产物光学纯度高等优点,在生物催化制备手性(r)-3-氨基-1-丁醇具有较好的工业应用前景。附图说明图1为醇脱氢酶偶联胺脱氢酶制备(r)-3-氨基-1-丁醇的反应示意图。图2为醇脱氢酶的粗酶粉催化1,3-丁二醇生成4-羟基-2-丁酮反应的gc检测图谱。图3为胺脱氢酶制备(r)-3-氨基-1-丁醇液相色谱hplc检测结果。e:混旋3-氨基-1-丁醇标准品液相色谱结果;f:样品(r)-3-氨基-1-丁醇液相色谱结果。图4为醇脱氢酶偶联胺脱氢酶制备(r)-3-氨基-1-丁醇液相色谱hplc检测结果。g:混旋3-氨基-1-丁醇标准品液相色谱结果;h:样品(r)-3-氨基-1-丁醇液相色谱结果。具体实施方式下面结合具体实施方式对本发明进行进一步的详细描述,给出的实施例仅为了阐明本发明,而不是为了限制本发明的范围。以下提供的实施例可作为本
技术领域:
普通技术人员进行进一步改进的指南,并不以任何方式构成对本发明的限制。下述实施例中的实验方法,如无特殊说明,均为常规方法,按照本领域内的文献所描述的技术或条件或者按照产品说明书进行。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。以下实施例中的定量试验,均设置三次重复实验,结果取平均值。下述实施例中,如无特殊说明,序列表中各核苷酸序列的第1位均为相应dna/rna的5′末端核苷酸,末位均为相应dna/rna的3′末端核苷酸。下述实施例中,adh1为来源于短小乳杆菌(lactobacillusbreviskb290)的醇脱氢酶,氨基酸序列的accessionnumber为ban05992.1;adh2为来源于高温厌氧杆菌(thermoanaerobacterbrockii)的醇脱氢酶,氨基酸序列的accessionnumber为caa46053.1;adh3为来源于固氮弧菌属(aromatoleumaromaticumebn1)的醇脱氢酶,氨基酸序列的accessionnumber为cai07428.1;adh4为来源于链霉菌(streptomycescoelicolor)的醇脱氢酶,氨基酸序列的accessionnumber为q9kym4;adh5为来源于枯草芽孢杆菌(bacillussubtilis)的醇脱氢酶,氨基酸序列的accessionnumber为wp_003234015.1;adh6为来源于肺炎克雷白杆菌(klebsiellapneumoniae)的醇脱氢酶,氨基酸序列的accessionnumber为wp_015958558.1;adh7为来源于葡萄糖杆菌(gluconobacteroxydans)的醇脱氢酶,氨基酸序列的accessionnumber为wp_011253549.1;adh8为来源于赖氏菌属(leifsoniasp.s749)的醇脱氢酶,氨基酸序列的accessionnumber为bad99642.1;adh9为来源于赤红球菌(rhodococcusruber)的醇脱氢酶,氨基酸序列的accessionnumber为q8klt9;adh10为来源于高加索酸奶乳杆菌(lactobacilluskefiridsm20587)的醇脱氢酶,氨基酸序列的accessionnumber为aap94029.1;adh11为来源于威吉利热厌氧杆菌(thermoanaerobacterwiegeliirt8.b1)的醇脱氢酶,氨基酸序列的accessionnumber为aem77517.1;adh12为来源于乳杆菌(lactobacilluszymaedsm19395)的醇脱氢酶,氨基酸序列的accessionnumber为krl08675.1;adh13为来源于酒球菌(oenococcusalcoholitolerans)的醇脱氢酶,氨基酸序列的accessionnumber为kgo31568.1;adh14为来源于愈伤热球菌(thermococcusguaymasensis)的醇脱氢酶,氨基酸序列的accessionnumber为adv18977.1;adh15为来源于肉毒梭菌(clostridiumbotulinum)的醇脱氢酶,氨基酸序列的accessionnumber为wp_003399463.1;adh16为来源于致黑脱硫肠状菌(desulfotomaculumnigrificans)的醇脱氢酶,氨基酸序列的accessionnumber为wp_003542410.1;adh17为来源于一氧化碳嗜热窦菌(thermosinuscarboxydivorans)的醇脱氢酶,氨基酸序列的accessionnumber为wp_007290608.1;adh18为来源于烷嗜热厌氧杆菌(thermoanaerobactermathranii)的醇脱氢酶,氨基酸序列的accessionnumber为wp_013150923.1;adh19为来源于梭菌属(clostridium)的醇脱氢酶,氨基酸序列的accessionnumber为wp_013239134.1;adh20为来源于厚壁菌门细菌(firmicutesbacteriumcag:137)的醇脱氢酶,氨基酸序列的accessionnumber为cdb31037.1;adh21为来源于拜氏梭菌(clostridiumbeijerinckii)的醇脱氢酶,氨基酸序列的accessionnumber为wp_026889046.1;adh22为来源于梭菌属(clostridium)的醇脱氢酶,氨基酸序列的accessionnumber为wp_039771361.1;adh23为来源于乙草甲烷菌(methanosarcinaacetivorans)的醇脱氢酶,氨基酸序列的accessionnumber为wp_048065256.1;adh24为来源于淡色甲烷菌(methanosarcinalacustris)的醇脱氢酶,氨基酸序列的accessionnumber为wp_048125376.1;adh25为来源于甲烷八叠球菌(methanosarcinathermophila)的醇脱氢酶,氨基酸序列的accessionnumber为wp_048166386.1;adh26为来源于西洋甲烷菌(methanosarcinasiciliae)的醇脱氢酶,氨基酸序列的accessionnumber为wp_048172170.1;adh27为来源于德氏葡萄球菌(desnuesiellamassiliensis)的醇脱氢酶,氨基酸序列的accessionnumber为wp_055668884.1;adh28为来源于卡尔德氏杆菌(caldanaerobactersubterraneus)的醇脱氢酶,氨基酸序列的accessionnumber为kuk09008.1;adh29为来源于钟氏梭菌(clostridiumljungdahlii)的醇脱氢酶,氨基酸序列的accessionnumber为wp_063556461.1;adh30为来源于拜氏梭菌(clostridiumbeijerinckii)的醇脱氢酶,氨基酸序列的accessionnumber为wp_065417405.1;adh31为来源于未培养梭菌(unculturedclostridiumsp.)的醇脱氢酶,氨基酸序列的accessionnumber为sci81347.1;adh32为来源于丝状孢子梭菌(clostridiumtaeniosporum)的醇脱氢酶,氨基酸序列的accessionnumber为wp_069679756.1;adh33为来源于脱硫弧菌(desulfovibriolitoralis)的醇脱氢酶,氨基酸序列的accessionnumber为wp_072695399.1;adh34为来源于脱硫肠状菌(desulfotomaculumputei)的醇脱氢酶,氨基酸序列的accessionnumber为wp_073236284.1;adh35为来源于拜氏梭菌(clostridiumbeijerinckii)的醇脱氢酶,氨基酸序列的accessionnumber为wp_077844196.1;adh36为来源于梭状芽胞杆菌(clostridiumpuniceum)的醇脱氢酶,氨基酸序列的accessionnumber为wp_077846831.1;adh37为来源于二醇梭菌(clostridiumdiolis)的醇脱氢酶,氨基酸序列的accessionnumber为wp_087701616.1;adh38为来源于匙形梭菌(clostridiumcochlearium)的醇脱氢酶,氨基酸序列的accessionnumber为wp_089865926.1;adh39为来源于笃斯梭菌(clostridiumuliginosum)的醇脱氢酶,氨基酸序列的accessionnumber为wp_090093236.1;adh40为来源于梭菌属细菌(clostridialesbacteriumsk-y3)的醇脱氢酶,氨基酸序列的accessionnumber为wp_094550774.1;adh41为来源于近平滑假丝酵母(clostridiumbotulinum)的醇脱氢酶,氨基酸序列的accessionnumber为wp_096043277.1;amdh1为来源于嗜热脂肪土芽孢杆菌(geobacillusstereothermophilus)的胺脱氢酶,氨基酸序列为seqidno.1;amdh2为来源于嗜热脂肪芽孢杆菌(bacillusstereothermophilus)的胺脱氢酶,氨基酸序列为seqidno.2;amdh3为来源于嗜冷芽孢八叠球菌(sporosarcinapsychrophila)的胺脱氢酶,氨基酸序列为seqidno.3;amdh4为来源于球形赖氨酸芽孢杆菌(lysinibacillussphaericus)的胺脱氢酶,氨基酸序列为seqidno.4;amdh5为来源于西伯利亚微杆菌(exiguobacteriumsibiricum)的胺脱氢酶的胺脱氢酶,氨基酸序列为seqidno.5;amdh6为来源于普通高温放线菌(thermoactinomycesintermedius)的胺脱氢酶,氨基酸序列为seqidno.6;amdh7为来源于温泉热碱芽孢杆菌(caldalkalibacillusthermarum)的胺脱氢酶,氨基酸序列为seqidno.7;amdh8为来源于糖莱斯氏菌(laceyellasacchari)的胺脱氢酶,氨基酸序列为seqidno.8;amdh9为来源于栗褐芽孢杆菌(bacillusbadius)的胺脱氢酶,氨基酸序列为seqidno.9;amdh3-m1为将seqidno.3所示的胺脱氢酶amdh3的第114位和第291位依次突变为缬氨酸残基和半胱氨酸残基,得到的胺脱氢酶突变体;amdh3-m2为将seqidno.3所示的胺脱氢酶amdh3的第111位、第114位、第294位依次突变为苯丙氨酸残基、缬氨酸残基和半胱氨酸残基,得到的胺脱氢酶突变体。2×high-fidelitymastermix为擎科生物科技有限公司的产品,货号为tp001。1,3-丁二醇和4-羟基-2-丁酮均为上海阿拉丁生化科技股份有限公司的产品,货号依次为b111018和h106330。混旋3-氨基-1-丁醇为上海韶远试剂,货号为sy030187。本发明的发明人经过大量实验,建立了醇脱氢酶和胺脱氢酶级联催化合成手性(r)-3-氨基-1-丁醇的方法。醇脱氢酶偶联胺脱氢酶制备(r)-3-氨基-1-丁醇的反应示意图见图1。实施例1、表达醇脱氢酶的工程菌和表达胺脱氢酶的工程菌的制备分别将醇脱氢酶和胺脱氢酶的编码基因送到金开瑞生物科技有限公司合成(根据需要以大肠杆菌为宿主进行密码子优化),将合成的基因连接于各种表达载体上构建而成。所述的表达载体为本领域常规的各种载体。本发明所述载体具体为pet28a,将pet28a的酶切识别位点ncoⅰ和xhoⅰ之间的dna小片段替换为全基因合成后的相关酶的编码基因,得到重组表达载体。将上述重组表达载体转化至合适的微生物宿主中。所述宿主微生物为本领域常规的各种宿主微生物,只要能满足上述重组表达载体稳定的自行复制以及醇脱氢酶或胺脱氢酶基因可被有效表达即可。其中,较佳的所述宿主微生物为大肠杆菌(escherichiacoli),优选大肠杆菌bl21(de3),将前述重组表达载体转入e.colibl21(de3)中,可得到本发明的基因工程菌株。利用定点突变方法得到醇脱氢酶基因的突变体,然后按照上述步骤,获得表达醇脱氢酶突变体的相应的基因工程菌株。利用定点突变方法得到胺脱氢酶基因的突变体,然后按照上述步骤,获得表达胺脱氢酶突变体的相应的基因工程菌株。实施例2、醇脱氢酶和胺脱氢酶共表达的工程菌的制备将醇脱氢酶(或醇脱氢酶突变体)和胺脱氢酶(或胺脱氢酶突变体)的编码基因分别进行全基因合成(根据需要以大肠杆菌为宿主进行密码子优化),将合成的基因连接于各种表达载体上构建而成。所述的表达载体为本领域常规的各种载体。本发明所述载体具体为pet28a。首先将pet28a的酶切识别位点ncoⅰ和xhoⅰ之间的dna小片段替换为全基因合成后的醇脱氢酶的编码基因,得到中间载体;然后采用重组克隆试剂盒(南京诺唯赞生物科技股份有限公司,货号c112-02)将全基因合成后胺脱氢酶的编码基因连接到醇脱氢酶的编码基因之后,得到重组表达质粒。将上述重组表达质粒转化至合适的微生物宿主中。所述宿主微生物为本领域常规的各种宿主微生物,只要能满足上述重组表达质粒稳定的自行复制以及醇脱氢酶基因和胺脱氢酶基因可被有效表达即可。其中,较佳的所述宿主微生物为大肠杆菌(escherichiacoli),优选大肠杆菌bl21(de3),将前述重组表达质粒转入e.colibl21(de3)中,即可得到本发明的基因工程菌株。实施例3、醇脱氢酶和胺脱氢酶的表达或共表达及全细胞、粗酶的制备1、将实施例1构建的重组表达载体或实施例2构建的重组表达质粒转入大肠杆菌bl21(de3)感受态细胞中,获得重组细胞。2、挑取转化子至含有50μg/ml卡那霉素的5mllb液体培养基中,37℃、220rpm震荡过夜12h,再按照体积百分含量1%接种量接种到含有50μg/ml卡那霉素的tb液体培养基中,37℃培养至od600nm为0.7时,加入终浓度为0.1mmol/l的iptg,20℃、220rpm诱导表达12h后,4℃、4000rpm离心10min,收集沉淀菌体(即全细胞),并将收集的菌体用磷酸盐缓冲液(50mmol/l,ph7.4)重悬,得到菌体悬液;之后在冰浴条件下超声破碎菌体细胞,得到破碎后的样品(即为粗酶液)。3、将粗酶液于4℃、8000rpm离心30min,收集上清,所得上清液置于-80℃冷冻后再用真空干燥仪冷冻干燥,获得粗酶粉。实施例4、醇脱氢酶或醇脱氢酶突变体催化1,3-丁二醇生成4-羟基-2-丁酮用实施例3制备的醇脱氢酶或醇脱氢酶突变体的粗酶粉催化1,3-丁二醇生成4-羟基-2-丁酮。在反应体系中依次加入浓度为100mm磷酸盐缓冲液(ph值为8.0)、终浓度为5mm的1,3-丁二醇、终浓度为1mm的nad+或nadp+、终浓度为2%的丙酮(v/v)和终浓度为10-40g/l的醇脱氢酶或醇脱氢酶突变体的粗酶粉。将该反应体系在30℃的条件下反应24h后,对产物进行气相色谱(gc)检测。气相色谱(gc)的检测条件如下:色谱柱:rtx-1(25×0.32mmid);升温程序:80℃,保留3分钟;5℃/min升温到120℃,保留4分钟;分流比:20:1;色谱柱流量:1ml/min;进样量:2μl;运行时间:15分钟。醇脱氢酶的粗酶粉催化1,3-丁二醇生成4-羟基-2-丁酮反应的gc检测结果图谱见图2(a为混旋1,3-丁二醇标准品气相色谱结果,b为4-羟基-2-丁酮标准品气相色谱结果,c为混旋1,3-丁二醇和4-羟基-2-丁酮标准品气相色谱结果,d为样品底物1,3-丁二醇和中间产物4-羟基-2-丁酮的气相色谱结果)。计算转化率,检测结果如表1所示。结果表明,醇脱氢酶和醇脱氢酶突变体均能够催化1,3-丁二醇生成4-羟基-2-丁酮,转化率为8-99%。表1.醇脱氢酶和醇脱氢酶突变体催化1,3-丁二醇生成4-羟基-2-丁酮的转化率结果名称转化率(%)名称转化率(%)adh143adh2246adh299adh239adh352adh2412adh460adh2587adh555adh2613adh654adh2760adh759adh2811adh860adh2954adh951adh3015adh1057adh3112adh1131adh3220adh1216adh3325adh1340adh3440adh1499adh3575adh1512adh3651adh1692adh3769adh1714adh3811adh1899adh3946adh198adh409adh2015adh4112adh2115注:转化率=a1/a2×100%;a1:气相色谱分析获得的样品1,3-丁二醇或4-羟基-2-丁酮峰面积值;a2:液相色谱分析获得的标准品1,3-丁二醇或4-羟基-2-丁酮的峰面积值。实施例5、胺脱氢酶或胺脱氢酶突变体全细胞催化4-羟基-2-丁酮生成(r)-3-氨基-1-丁醇用实施例3制备的胺脱氢酶或胺脱氢酶突变体全细胞催化4-羟基-2-丁酮生成(r)-3-氨基-1-丁醇。1、在反应体系中依次加入浓度为1m的氯化铵/氨水缓冲液(ph值为8.5)、终浓度为20mm的4-羟基-2-丁酮、终浓度为2g/l的gdh粗酶粉、终浓度为100mm的葡萄糖、终浓度为1mm的nad+和终浓度为100g/l的胺脱氢酶或胺脱氢酶突变体的全细胞。将该反应体系在30℃的条件下反应24h,得到反应液。2、将300μl0.4mol/l硼酸缓冲液(ph9.5)、150μl超纯水、200μl衍生剂和100μl反应液充分混匀并静置2分钟,然后12000rpm离心10分钟,收集上清并用滤膜过滤。衍生剂:将0.343g邻苯二甲醛、5ml无水乙醇、0.147gn-乙酰-l半胱氨酸和适量0.4mol/l硼酸缓冲液(ph9.5)混合,然后用0.4mol/l硼酸缓冲液(ph9.5)定容至25ml,避光备用。3、对滤液进行hplc检测。hplc检测条件为:agilentsb-aqc18柱(4.6*250mm,5μm);检测波长334nm;柱温:35℃,流速:1ml/min,上样量:10μl;梯度洗脱程序如表2所示。表2.hplc的梯度洗脱程序时间(min)流动相(甲醇)%流动相(0.05mol/l乙酸钠)%0-6(不包括6)30706-7(包括6,不包括7)30707-15(包括7,不包括15)455515-15.5(包括15,不包括15.5)455515.5-20(包括15.5和20)3070注:表中的%表示体积百分含量。胺脱氢酶催化还原4-羟基-2-丁酮生成(r)-3-氨基-1-丁醇反应的hplc检测结果图谱如图3所示(e为混旋3-氨基-1-丁醇标准品液相色谱结果,f为样品(r)-3-氨基-1-丁醇液相色谱结果)。计算转化率,部分hplc检测结果如表3所示。结果表明,胺脱氢酶或其突变体全细胞可以不对称还原催化4-羟基-2-丁酮生成(r)-3-氨基-1-丁醇,转化率为5-36%,立体选择性大于99%(r)。表3.胺脱氢酶及其突变体全细胞催化4-羟基-2-丁酮生成(r)-3-氨基-1-丁醇的转化率结果名称转化率(%)ee(%)amdh136>99(r)amdh220>99(r)amdh323>99(r)amdh3-m120>99(r)amdh3-m229>99(r)amdh426>99(r)amdh523>99(r)amdh6<5>99(r)amdh7<5>99(r)amdh8<5>99(r)amdh9<5>99(r)注:转化率=a1/a2×100%;a1:液相色谱分析获得的(r)-3-氨基-1-丁醇峰面积值;a2:液相色谱分析获得的标准品(r)-3-氨基-1-丁醇的峰面积值;ee(立体选择性)=(as-ar)/(as+ar)×100%;as:液相色谱分析获得的(s)-3-氨基-1-丁醇的峰面积值;ar:液相色谱分析获得的(r)-3-氨基-1-丁醇的峰面积值。实施例6、醇脱氢酶(或醇脱氢酶突变体)和胺脱氢酶(或胺脱氢酶突变体)共表达全细胞催化1,3-丁二醇生成(r)-3-氨基-1-丁醇用实施例3制备的醇脱氢酶(或醇脱氢酶突变体)和胺脱氢酶(或胺脱氢酶突变体)全细胞催化1,3-丁二醇生成(r)-3-氨基-1-丁醇。1、在反应体系中依次加入浓度为1m的氯化铵/氨水缓冲液(ph值为8.0)、终浓度为20mm的1,3-丁二醇、终浓度为1mm的即nad+、终浓度为1g/l的溶菌酶、终浓度为6u/ml的dnasei(脱氧核糖核酸酶)、终浓度为200g/l醇脱氢酶(或醇脱氢酶突变体)和胺脱氢酶(或胺脱氢酶突变体)共表达全细胞。将该反应体系在30℃的条件下反应24h,得到反应液。2、将50μl反应液、50μl1mol/l氯化铵/氨水缓冲液(ph8.5)和500μl乙腈充分混匀,然后过滤。3、将20μl步骤2得到的滤液、20μlmarfey’s试剂(浓度为14mmol/l,溶剂为乙腈)、36μlnahco3(浓度为1mol/l)和100μldmso混匀,40℃、1000rpm反应2h后,加入40μlhcl(浓度为1mol/l)终止反应,得到反应液。4、将步骤3得到的反应液进行hplc检测。hplc检测条件为:zorbaxsb-c18柱(4.6×150mm,5μm);检测波长340nm;柱温:25℃;流速:1ml/min;上样量:10μl;流动相a:超纯水(0.1%三氟乙酸),流动相b:甲醇(0.1%三氟乙酸)。洗脱程序:60%a/40%b;40%b保留6分钟;在9分钟内增加b到60%,保留3分钟;在2分钟内减少b到40%,保留5分钟。醇脱氢酶(或醇脱氢酶突变体)和胺脱氢酶(或胺脱氢酶突变体)共表达全细胞催化1,3-丁二醇生成(r)-3-氨基-1-丁醇的hplc检测结果图谱如图4所示。计算转化率,部分hplc检测结果如表4所示。结果表明,醇脱氢酶(或醇脱氢酶突变体)和胺脱氢酶(或胺脱氢酶突变体)可以共表达全细胞形式催化1,3-丁二醇生成(r)-3-氨基-1-丁醇,转化率为9-38%,立体选择性大于99%(r)。表4.醇脱氢酶(或醇脱氢酶突变体)和胺脱氢酶(或胺脱氢酶突变体)共表达全细胞催化1,3-丁二醇生成(r)-3-氨基-1-丁醇的转化率结果名称转化率(%)ee(%)adh4-amdh118>99(r)adh4-amdh29>99(r)adh4-amdh315>99(r)adh4-amdh420>99(r)adh14-amdh132>99(r)adh14-amdh216>99(r)adh14-amdh328>99(r)adh14-amdh435>99(r)adh18-amdh135>99(r)adh18-amdh218>99(r)adh18-amdh326>99(r)adh18-amdh438>99(r)注:转化率=a1/a2×100%;a1:液相色谱分析获得的(r)-3-氨基-1-丁醇峰面积值;a2:液相色谱分析获得的标准品(r)-3-氨基-1-丁醇的峰面积值;ee(立体选择性)=(as-ar)/(as+ar)×100%;as:液相色谱分析获得的(s)-3-氨基-1-丁醇的峰面积值;ar:液相色谱分析获得的(r)-3-氨基-1-丁醇的峰面积值。<110>中国科学院天津工业生物技术研究所<120>一种双酶级联催化合成(r)-3-氨基-1-丁醇的方法<160>9<170>patentinversion3.5<210>1<211>429<212>prt<213>artificialsequence<400>1metgluleuphelystyrmetgluthrtyrasptyrgluglnvalleu151015phecysglnasplysgluserglyleulysalaileilealailehis202530aspthrthrleuglyproalaleuglyglythrargmettrpmettyr354045asnserglugluglualaleugluaspalaleuargleualaarggly505560metthrtyrserasnalaalaalaglyleuasnleuglyglyglylys65707580thrvalileileglyaspproarglysasplysasnglualametphe859095argalapheglyargpheileglnglyleuasnglyargtyrilethr100105110alagluaspvalglythrthrvalalaaspmetaspileiletyrgln115120125gluthrasptyrvalthrglyileserproglupheglysersergly130135140asnproserproalathralatyrglyvaltyrargglymetlysala145150155160alaalalysglualapheglyseraspserleugluglylysvalval165170175alavalglnglyvalglyasnvalalatyrhisleucysarghisleu180185190hisglugluglyalalysleuilevalthraspileasnlysgluval195200205valalaargalavalgluglupheglyalalysalavalaspproasn210215220aspiletyrglyvalglucysaspilephealaprocysalaleugly225230235240glyileileasnaspglnthrileproglnleulysalalysvalile245250255alaglyseralaleuasnglnleulysgluproarghisglyaspile260265270ilehisglumetglyilevaltyralaproasptyrvalileasnala275280285glyglyvalileasnvalalaaspgluleutyrglytyrasnargglu290295300argalametlyslysilegluglniletyraspasnileglulysval305310315320phealailealalysargaspasnileprothrtyrvalalaalaasp325330335argmetalaglugluargilegluthrmetarglysalaalasergln340345350pheleuglnasnglyhishisileleuserargargproargproleu355360365thralaalaargalaglyleuargargalaaspaspglyglythrthr370375380thrmetglngluglnlyspheargileleuthrileasnproglyser385390395400thrserthrlysileglyvalphegluasngluargalailealaser405410415lyslysargseralathrargalaglyalaseralaile420425<210>2<211>366<212>prt<213>artificialsequence<400>2metthrleugluilepheglutyrleuglulystyrasptyrglugln151015valvalphecysglnasplysgluserglyleulysalaileileala202530ilehisaspthrthrleuglyproalaleuglyglythrargmettrp354045thrtyraspsergluglualaalailegluaspalaleuargleuala505560lysglymetthrtyrserasnalaalaalaglyleuasnleuglygly65707580alalysthrvalileileglyaspproarglysasplyssergluala859095metpheargalaleuglyargtyrileglnglyleuasnglyargtyr100105110ilethralagluaspvalglythrthrvalaspaspmetaspileile115120125hisglugluthraspphevalthrglyileserproserpheglyser130135140serglyasnproserprovalthralatyrglyvaltyrargglymet145150155160lysalaalaalalysglualapheglythraspasnleugluglylys165170175valilealavalglnglyvalglyasnvalalatyrhisleucyslys180185190hisleuhisalagluglyalalysleuilevalthraspileasnlys195200205glualavalglnargalavalgluglupheglyalaseralavalglu210215220proasngluiletyrglyvalglucysaspiletyralaprocysala225230235240leuglyalathrvalasnaspgluthrileproglnleulysalalys245250255valilealaglyseralaleuasnglnleulysgluasparghisgly260265270aspileilehisglumetglyilevaltyralaproasptyrvalile275280285asnalaglyglyvalileasnvalalaaspgluleutyrglytyrasn290295300arggluargalaleulysargvalgluseriletyraspthrileala305310315320lysvalilegluileserlysargaspglyilealathrtyrvalala325330335alaaspargleualaglugluargilealaserleulysasnserarg340345350serthrtyrleuargasnglyhisaspileileserargarg355360365<210>3<211>364<212>prt<213>artificialsequence<400>3metgluilephelystyrmetgluhisglnasptyrgluglnleuval151015ilecysglnasplysalaserglyleulysalaileilealailehis202530aspthrthrleuglyproalaleuglyglythrargmettrpthrtyr354045alaserglugluglualailegluaspalaleuargleualaarggly505560metthrtyrserasnalaalaalaglyleuasnleuglyglyglylys65707580thrvalileileglyasnprolysthrasplysasnaspglumetphe859095argalapheglyargtyrilegluglyleuasnglyargtyrilethr100105110alagluaspvalglythrthrglualaaspmetaspleuileasnleu115120125gluthrasptyrvalthrglythrseralaglyalaglysersergly130135140asnproserprovalthralatyrglyiletyrtyrglymetlysala145150155160alaalalysglualapheglyaspaspserleualaglylysthrval165170175alavalglnglyvalglyasnvalalatyralaleucysglutyrleu180185190hisglugluglyalalysleuileilethraspileasnglugluala195200205valglnargalavalaspalapheglyalathralavalglyileasn210215220gluiletyrserglnglualaaspilephealaprocysalaleugly225230235240alaileileasnaspgluthrileproglnleulysalalysvalile245250255alaglyseralaleuasnglnleulysgluthrarghisglyaspleu260265270ilehisglumetglyilevaltyralaproasptyrvalileasnser275280285glyglyvalileasnvalalaaspgluleuaspglytyrasnargglu290295300argalaleulysargvalgluglyiletyraspvalileglylysile305310315320phealaileserlysargaspasnileprothrtyrvalalaalaasp325330335argmetalaglugluargilealaargvalalaasnthrargserthr340345350pheleuglnasnglulysservalleuserargarg355360<210>4<211>362<212>prt<213>artificialsequence<400>4metgluilephelystyrmetglulystyrasptyrgluglnleuval151015phecysglnaspglualaserglyleulysalavalilealailehis202530aspthrthrleuglyproalaleuglyglyalaargmettrpthrtyr354045alaserglugluasnalavalgluaspalaleuargleualaarggly505560metthrtyrserasnalaalaalaglyleuasnleuglyglyglylys65707580thrvalileileglyaspprophelysasplysasngluglumetphe859095argalaleuglyargpheileglnglyleuasnglyargtyrilethr100105110alagluaspvalglythrthrvalthraspmetaspleuilehisglu115120125gluthrasptyrvalthrglyileserproalapheglysersergly130135140asnproserprovalthralatyrglyvaltyrargglymetlysala145150155160alaalalysglualapheglysergluserleugluglyleulysile165170175servalglnglyleuglyasnvalalatyrlysleucysglutyrleu180185190hisasngluglyalalysleuvalvalthraspileasnglnalaala195200205ileaspargvalvalasnasppheaspalailealavalalaproasp210215220gluiletyralaglngluvalaspilepheserprocysalaleugly225230235240alaileleuasnaspgluthrileproglnleulysalalysvalile245250255alaglyseralaleuasnglnleulysaspserarghisglyaspphe260265270leuhisgluleuglyilevaltyralaproasptyrvalileasnala275280285glyglyvalileasnvalalaaspgluleutyrglytyrasnargglu290295300argalaleulysargvalaspglyiletyraspserileglulysile305310315320phealaileserlysargaspglyileprothrtyrvalalaalaasn325330335argleualaglugluargilealaargvalalalysserargsergln340345350pheleulysasnglulysasnileleuhis355360<210>5<211>374<212>prt<213>artificialsequence<400>5metvalgluthrasnvalglualaargpheserilephegluthrmet151015alametgluasptyrgluglnvalvalphecyshisasplysvalser202530glyleulysalaileilealailehisaspthrthrleuglyproala354045leuglyglyleuargmettrpasntyralaseraspgluglualaleu505560ileaspalaleuargleualalysglymetthrtyrserasnalaala65707580alaglyleuasnleuglyglyglylysalavalileileglyaspala859095lysthrglnlysserglualaleupheargalapheglyargtyrval100105110glnserleuasnglyargtyrilethralagluaspvalasnthrthr115120125valalaaspmetasptyrilehismetgluthraspphevalthrgly130135140valserproalapheglyserserglyasnproserprovalthrala145150155160tyrglyvaltyrargglymetlysalaalaalalysgluvaltyrgly165170175thraspserleuglyglylysthrvalalaileglnglyvalglyasn180185190valalapheasnleucysarghisleuhisglugluglyalalysleu195200205ilevalthraspileasnglnaspalaleuargargalaglugluala210215220pheglyalaleuvalvalglyproaspgluiletyrservalaspala225230235240aspilephealaprocysalaleuglyalathrleuasnaspgluthr245250255ileproglnleulysvallysileilealaglyalaalaleuasngln260265270leulysgluasparghisglyaspmetleuglngluargglyileleu275280285tyrthrproaspphevalileasnalaglyglyvalileasnvalala290295300aspgluleuaspglytyrasnarggluargalametlyslysvalglu305310315320leuvaltyraspalavalalalysvalilegluilealalysargasp325330335hisleuprothrtyrargalaalaglulysmetalaglugluargile340345350alathrmetglyseralaargserglnpheleuargargasplysasn355360365ileleuglyserarggly 370<210>6<211>366<212>prt<213>artificialsequence<400>6metlysilepheasptyrmetglulystyrasptyrgluglnleuval151015metcysglnasplysgluserglyleulysalaileilecysilehis202530valthrthrleuglyproalaleuglyglymetargmettrpthrtyr354045alaserglugluglualailegluaspalaleuargleuglyarggly505560metthrtyrserasnalaalaalaglyleuasnleuglyglyglylys65707580thrvalileileglyaspproarglysasplysasnglualametphe859095argalaleuglyargpheileglnglyleuasnglyargtyrilethr100105110alagluaspvalglythrthrvalgluaspmetaspileilehisglu115120125gluthrargtyrvalthrglyvalserproalapheglysersergly130135140asnproserprovalthralatyrglyvaltyrargglymetlysala145150155160alaalalysglualapheglyaspaspserleugluglylysvalval165170175alavalglnglyvalglyhisvalalatyrgluleucyslyshisleu180185190hisasngluglyalalysleuilevalthraspileasnlysgluasn195200205alaaspargalavalglnglupheglyalagluphevalhisproasp210215220lysiletyraspvalglucysaspilephealaprocysalaleugly225230235240alaileileasnaspgluthrilegluargleulyscyslysvalval245250255alaglyseralaleuasnglnleulysglugluarghisglylysmet260265270leugluglulysglyilevaltyralaproasptyrvalileasnala275280285glyglyvalileasnvalalaaspgluleuleuglytyrasnargglu290295300argalametlyslysvalgluglyiletyrasplysileleulysval305310315320phegluilealalysargaspglyileprosertyrleualaalaasp325330335argmetalaglugluargileglumetmetarglysthrargserthr340345350pheleuglnaspglnargasnleuileasnpheasnasnlys355360365<210>7<211>370<212>prt<213>artificialsequence<400>7metserthrvalthrpheaspglnilesergluhisgluglnvalmet151015phecysasnaspprohisthrglyleulysalaileilealailehis202530asnthrthrleuglyproalaleuglyglycysargmetleuprotyr354045lysserglugluglualaleuthraspvalleuargleuserlysgly505560metthrtyrsercysvalalaalaaspvalasppheglyglyglylys65707580alavalileileglyaspproarglysasplysthrprogluleuphe859095argalapheglyglnphevalglnserleuasnglyargphetyrthr100105110glythraspmetglythrthrprogluaspphevalglnalatyrlys115120125gluthrserpheilevalglyleuprogluglutyrglyglyasngly130135140aspserservalthrthralapheglyvalmetglnglyleuargala145150155160valserglnpheleutrpglythraspvalleuthrgluargvalphe165170175alavalglnglyleuglylysvalglyphelysvalalagluglyleu180185190leulysgluglyalaasnvaltyrvalthraspvalaspprogluthr195200205ilealalysleugluglulysalatyrglntyrproglyhisvalgln210215220alavalthralaaspaspiletyrglyvalglyalaaspvalpheval225230235240procysalaileglyglyileileasnaspgluthrilegluargleu245250255lysvallysalavalcysglyalaalaleuasnglnleuleugluasp260265270arghisglylysvalleuglnalalysasnileleutyralaproasp275280285tyrilevalasnalaglyglyleuileglnvalseraspgluleutyr290295300glyproasnlysalaargvalleulyslysthrargalaleutyrasp305310315320thrleuphegluilepheglnseralaglulyslysalavalserthr325330335valglualaalaasnglnphevalglugluargleuglnlysargala340345350argleuasnserphepheserproaspasnproprolystrpargval355360365argarg370<210>8<211>366<212>prt<213>artificialsequence<400>8metlysilepheglutyrmetglylystyrasptyrgluglnleuval151015leucyshisaspgluglnserglyleulysalaileilecysilehis202530aspthrthrleuglyproalaleuglyglythrargmettrpthrtyr354045aspsergluaspalaalailegluaspalaleuargleualaarggly505560metthrtyrserasnalaalaalaglyleuasnleuglyglyglylys65707580thrvalvalileglyaspprolyslysasplysserglualaleuphe859095argalaleuglyargtyrileglnglyleuasnglyargtyrilethr100105110alagluaspvalglythrthrvalgluaspmetaspileileargglu115120125gluthrlystyrvalthrglyvalserproalapheglysersergly130135140asnproserprovalthralatyrglyvaltyrlysglymetlysala145150155160alaserlysvalalapheglygluaspserleulysglylysvalval165170175alavalglnglyvalglyhisvalalatyrasnleucyslyshisleu180185190hisalagluglyalalysleuilevalthraspileasnglnalaasn195200205glyaspargalavalglnglupheglyalaglualavalserproasp210215220lysiletyraspvalaspcysaspilepheserprocysalaleugly225230235240alaileileasnaspgluthrilegluargleuthrcyslysvalval245250255alaglyalaalaleuasnglnleulysgluglulyshisglyglumet260265270leugluglnlysglyileiletyralaproasptyrvalileasnala275280285glyglyvalileasnvalalaaspgluleutyrglytyrasnargasp290295300argalametlysargvalgluthriletyraspasnmetleulysval305310315320phegluilealalysargaspglyileprosertyrlysalaalaasp325330335argmetalaglugluargilealaalametarglysthrargserthr340345350pheleuvalasnglyglnserileleuserhisargleuglu355360365<210>9<211>380<212>prt<213>artificialsequence<400>9metserleuvalglulysthrserileilelysaspphethrleuphe151015glulysmetsergluhisgluglnvalvalphecysasnaspproala202530thrglyleuargalaileilealailehisaspthrthrleuglypro354045alaleuglyglycysargmetglnprotyrasnservalglugluala505560leugluaspalaleuargleuserlysglymetthrtyrsercysala65707580alaseraspvalasppheglyglyglylysalavalileileglyasp859095proglnlysasplysserprogluleupheargalapheglyglnphe100105110valaspserleuglyglyargphetyrthrglythraspmetglythr115120125asnmetgluasppheilehisalametlysgluthrasncysileval130135140glyvalproglualatyrglyglyglyglyaspserserileprothr145150155160alametglyvalleutyrglyilelysalathrasnlysmetleuphe165170175glylysaspaspleuglyglyvalthrtyralaileglnglyleugly180185190lysvalglytyrlysvalalagluglyleuleuglugluglyalahis195200205leuphevalthraspileasngluglnthrleuglualaileglnglu210215220lysalalysthrthrserglyservalthrvalvalalaseraspglu225230235240iletyrserglnglualaaspvalphevalprocysalapheglygly245250255valvalasnaspgluthrmetlysglnphelysvallysalaileala260265270glyseralaleuasnglnleuleuthrgluasphisglyarghisleu275280285alaasplysglyileleutyralaproasptyrilevalasnsergly290295300glyleuileglnvalalaaspgluleutyrgluvalasnlysgluarg305310315320valleualalysthrlyshisiletyraspalaileleugluvaltyr325330335glnglnalagluleuaspglnilethrthrmetglualaalaasnarg340345350metcysgluglnargmetalaalaargglyargargasnserphephe355360365thrserservallysprolystrpaspileargasn370375380当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1