一种ERK和JNK信号通路抑制剂Mce2E的制作方法
本发明涉及一种erk和jnk信号通路抑制剂mce2e,是一种能同时抑制erk和jnk信号通路的结核分枝杆菌分泌蛋白,属于细胞生物学领域。
背景技术:
结核分枝杆菌是一种兼性胞内致病菌,以巨噬细胞为主要的宿主细胞,并在其中存活和生长。哺乳动物细胞入侵蛋白(mammaliancellentry,mce)mce是由mce基因编码的一组分泌或者膜表面粘附蛋白,已被证明具有促进分枝杆菌入侵宿主细胞的功能。其中mce2操纵子就与分枝杆菌的毒力和肉芽肿形成有关,但是,mce2操纵子编码的mce2e蛋白在炎症和免疫相关信号通路的相关功能尚不清楚。
c-jun氨基末端激酶(c-junn-terminalkinase,jnk)和细胞外信号调节蛋白激酶(erk)p42/p44通路属于丝裂原活化蛋白激酶(mitogen-activatedproteinkinases,mapks)信号转导通路。因此,erk和jnk信号通路是正常与疾病状态时细胞的一个重要调节靶点。
jnk信号通路在细胞应激中发挥重要作用。目前,jnk的研究与肥胖和胰岛素抵抗有密切关系,具体的分子机制研究也比较清楚。越来越多的证据表明jnk1和jnk2的活化能促进肥胖和胰岛素抵抗的发展,在特定的疾病发展阶段,jnk引起的细胞应激与肥胖和二型糖尿病有关。
erk1/2是一种丝氨酸/苏氨酸蛋白激酶,通常与细胞生长和增殖有关。分子量分别为44kd和42kd的蛋白,二者之间有90%的同源性。在静止的细胞中,erk通过与各种锚定蛋白(包括mek)的相互作用被固定在细胞质中,当细胞受到刺激,erk1/2通路被激活后,p-erk1/2由于构象的变化脱离锚定蛋白,相关的研究表明,erk1/2信号通路的活性与炎症反应、肝脏疾病、肥胖症和糖尿病等代谢性疾病,尤其是erk信号的激活能够促进肿瘤的发展。
技术实现要素:
本发明旨在提供一种erk和/或jnk信号通路抑制剂,可以抑制erk1/2和jnk蛋白的磷酸化,并且通过与erk1/2和jnk相互作用使其定位于内质网无法入核,从而抑制了erk和jnk信号通路的活化。
所述抑制剂含有氨基酸序列包括如以下(a)或(b)或(c)所示氨基酸序列的多肽或蛋白质作为活性成分:
(a)seqidno:1所示的氨基酸序列;
(b)在(a)中的氨基酸序列经过取代、缺失或添加一个氨基酸或几个氨基酸且具有结合erk1/2活性的由(a)衍生的多肽或其类似物;
(c)与(a)、(b)中氨基酸序列整体相似度在85%以上的由(a)衍生的多肽或其类似物;
或者,含有氨基酸序列包括如seqidno:2所示氨基酸序列的多肽作为活性成分。
本发明要解决的另一个技术问题是提供所述抑制剂的制备方法,包括以下步骤:
(1)构建含有编码seqidno.1所示蛋白的基因的重组质粒pgex-6p-1-mce2e,并将质粒转化到大肠杆菌中进行表达;
(2)收集菌体,破碎后离心收取上清液,将上清液流过填充好的glutathione-sepharosebeads中,然后加入柱洗涤缓冲液充分洗涤柱子,最后加入含有谷胱甘肽的洗脱液洗脱蛋白,将收集的洗脱液加入到10kd的蛋白浓缩管中,离心浓缩;
(3)在蛋白浓缩管中加入pbs混匀后离心、分装蛋白,-80℃保存待用。
本发明发现seqidno.1所示蛋白(mce2e)中的关键模序rtlasvalil可与erk和jnk相互作用。进一步研究发现mce2e定位在内质网,并通过与erk和jnk的直接互作将其隔离到内质网,从而阻止erk1/2和jnk蛋白的磷酸化及入核。本发明找到了可以抑制erk和jnk信号通路的seqidno.1所示蛋白及其关键模序。由于erk和jnk信号通路与多种疾病的发生,发展有密切相关,因此抑制这两种信号通路的活化这对于很多疾病都有控制作用。本发明成果将来可为临床多种疾病的治疗提供新的工具和思路,也可以直接应用于科研领域或指导开发erk1/2和jnk通路的抑制剂。
附图说明
图1:(a)mce2e能抑制erk信号通路;(b)mce2e抑制jnk信号通路;
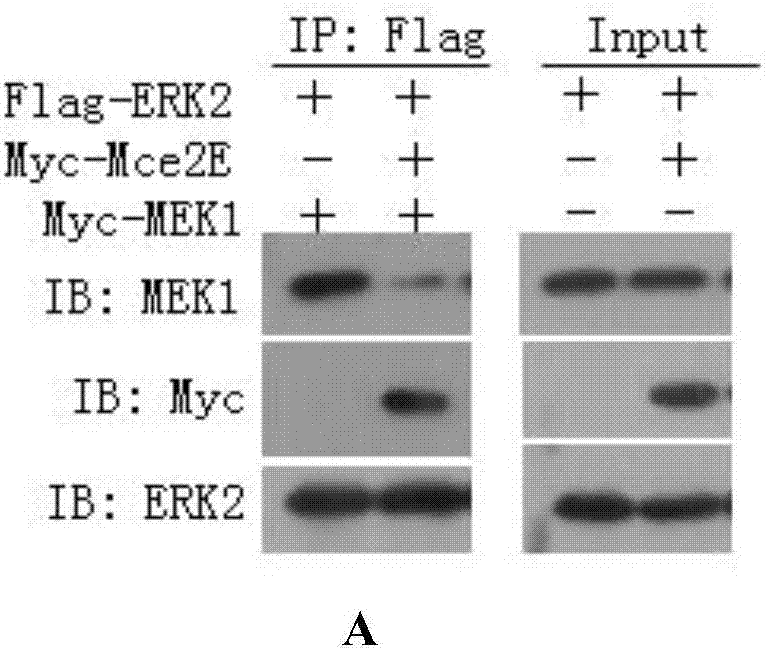
图2:(a)mce2e与erk2相互作用,阻断erk与上游信号mek1的作用,抑制erk信号通路;(b)mce2e与jnk1相互作用,阻断mce2e与上游信号通路mmk4的作用,抑制jnk信号通路;
图3:mce2e主要通过模序rtlasvalil抑制erk和jnk信号通路;
图4:(a)mce2e将erk隔离到内质网,阻止erk入核;(b)mce2e将jnk隔离到内质网,从而阻止jnk入核;
具体实施方式
实施例1结核分枝杆菌分泌蛋白mce2e
结核分枝杆菌分泌蛋白mce2e,其氨基酸序列如seqidno:1所示。mce2e及其结构类似物可用于erk1/2和jnk活性抑制剂的开发。erk2基因序列如geneid:5594所示,jnk1基因序列如geneid:5602所示。
实施例2mce2e能够抑制erk和jnk通路的活化
(a)采用双荧光报告基因技术检测mce2e对erk1/2通路活化的影响
1)构建含有编码mce2e基因的重组质粒pcdna6a-mce2e;
2)构建含有编码mek1-ed基因的重组质粒pcs2ha-mek1-ed,采用重叠延伸pcr法将mek1蛋白中的218位丝氨酸以及222位丝氨酸分别突变为谷氨酸和天冬氨酸后获得mek1-ed基因片段;
3)采用磷酸钙转染法将重组质粒pcdna6a、pcdna6a-mce2e和检测erk通路活化情况的质粒pgal4-elk(0.6μg)、pgal4-luc(0.6μg)、prl-tk(50ng)(购买自promega公司)以及表达erk1/2通路组成型激活剂的pcs2ha-mek1-ed转染进293t细胞;
4)转染6h后换新鲜培养基,24h后用promega公司的试剂盒以及荧光测量仪检测mce2e对erk通路活化的影响。
(b)采用双荧光报告基因技术检测mce2e对jnk通路活化的影响
1)采用磷酸钙转染法将重组质粒pcdna6a、pcdna6a-mce2e和检测jnk通路活化情况的质粒pfa-cjun(0.3μg),pgal4-luc(0.9μg),prl-tk(50ng)(购买自promega公司)以及表达jnk通路组成型激活剂racl61的质粒转染进293t细胞;
2)转染6h后换新鲜培养基,24h后用promega公司的试剂盒以及荧光测量仪检测mce2e对jnk通路活化的影响。
(c)实验结果分析
双荧光报告基因实验结果表明elk的活性明显降低,从而揭示erk活化收到抑制,mce2e可以显著抑制由mek1-ed激活的erk通路(见图1a);ap-1的活性明显降低,从而揭示jnk活化收到抑制,mce2e可以显著抑制racl61激活的jnk通路(见图1b);因此mce2e是同时抑制erk1/2和jnk信号通路的活化。
实施例3mce2e蛋白能够与erk2和jnk蛋白互作,并抑制erk和jnk的磷酸化
(a)mce2e和erk2相互作用,阻断erk2与mek1相互作用。
1)构建含有erk2(geneid:5594)基因的重组质粒p3xflag-cmv14-erk2和含有mek1(geneid:5604)基因的重组质粒pcdna6a-mek1。
2)用pei转染试剂在两个10cm细胞盘293t细胞中共转p3xflag-cmv14-erk2,分别转染空载pcdna6a,pcdna6a-mce2e转染4-6h后换液;
3)转染24h后,收集细胞,分别将1/20的细胞置冰上用western及ip细胞裂解液(p0013;碧云天)裂解,高速离心收集上清后,用westernblot检测两组细胞中各个质粒的表达情况;
4)当两组细胞中质粒表达正常时,分别将剩余的细胞用相同的方法裂解并收集上清;
5)取20μl小鼠抗flagm2亲和凝胶颗粒(f3165;sigma-aldrich),用100μl细胞裂解液洗涤后加入上步细胞裂解上清中,在4℃旋转孵育6h使flag标签蛋白能够与亲和凝胶颗粒充分结合;
6)4℃离心收集凝胶颗粒,用细胞裂解液洗涤凝胶颗粒5次(1ml/次)用以除去非特异结合的杂蛋白;
7)用50μl1×sds-page上样缓冲液重悬凝胶颗粒,90℃煮样10min后,用westernblot检测结合在凝胶颗粒上的蛋白。
(b)mce2e和jnk1相互作用,阻断jnk1和map2k4的相互作用。
1)构建含有jnk1(geneid:5602)基因的重组质粒p3xflag-cmv14-jnk1和含有map2k4(geneid:6416)基因的重组质粒pegfp-n1-map2k4;
2)用pei转染试剂在三个10cm细胞盘293t细胞中共转p3xflag-cmv14-jnk1和gfp-map2k4质粒,分别转染空载pcdna6a,pcdna6a-mce2e转染4-6h后换液;
3)转染24h后,收集细胞,分别将1/20的细胞置冰上用western及ip细胞裂解液(p0013;碧云天)裂解,高速离心收集上清后,用westernblot检测两组细胞中各个质粒的表达情况;
4)当两组细胞中质粒表达正常时,分别将剩余的细胞用相同的方法裂解并收集上清;
5)取20μl小鼠抗flagm2亲和凝胶颗粒(f3165;sigma-aldrich),用100μl细胞裂解液洗涤后加入上步细胞裂解上清中,在4℃旋转孵育6h使flag标签蛋白能够与亲和凝胶颗粒充分结合;
6)4℃离心收集凝胶颗粒,用细胞裂解液洗涤凝胶颗粒5次(1ml/次)用以除去非特异结合的杂蛋白;
7)用50μl1×sds-page上样缓冲液重悬凝胶颗粒,90℃煮样10min后,用westernblot检测结合在凝胶颗粒上的蛋白。
(c)实验结果分析
从图2中可以看出,mce2e和erk2、jnk1存在相互作用,并且抑制erk和jnk的磷酸化。
实施例4mce2e主要通过“rtlasvalil”模序抑制erk及jnk信号通路
(a)采用双荧光报告基因技术检测mce2eaaa对erk1/2通路活化的影响
1)将“rtlasvalil”模序中的关键氨基酸v、l、l分别突变为a、a、a(pcdna6a-mce2eaaa),将非关键氨基酸位点r突变为e作为阴性对照(pcdna6a-mce2er168e);
2)采用磷酸钙转染法将重组质粒pcdna6a、pcdna6a-mce2e、pcdna6a-mce2eaaa、pcdna6a-mce2er168e和检测erk通路活化情况的质粒pgal4-elk(0.6μg)、pgal4-luc(0.6μg)、prl-tk(50ng)(购买自promega公司)以及表达erk1/2通路组成型激活剂的pcs2ha-mek1-ed转染进293t细胞;
3)转染6h后换新鲜培养基,24h后用promega公司的试剂盒以及荧光测量仪检测mce2eaaa对erk通路活化的影响。
(b)采用双荧光报告基因技术检测mce2e对jnk通路活化的影响
1)采用磷酸钙转染法将重组质粒pcdna6a、pcdna6a-mce2e、pcdna6a-mce2eaaa、pcdna6a-mce2er168e和检测jnk通路活化情况的质粒pfa-cjun(0.3μg),pgal4-luc(0.9μg),prl-tk(50ng)(购买自promega公司)以及表达jnk通路组成型激活剂racl61的质粒转染进293t细胞;
2)转染6h后换新鲜培养基,24h后用promega公司的试剂盒以及荧光测量仪检测mce2e对jnk通路活化的影响。
(c)实验结果分析
双荧光报告基因实验结果表明mce2eaaa突变体不能抑制erk信号通路的激活,因此mce2e抑制erk通路依赖“rtlasvalil”模序(见图1a);同样的,mce2eaaa突变体也丧失了对jnk信号通路的抑制作用。因此mce2e主要通过“rtlasvalil”模序抑制erk,jnk信号通路激活。
抑制erk信号通路并不是mce蛋白家族特有的,有文献报道,mce家族的蛋白mce3a就无法抑制erk信号通路。mce2e蛋白,和已经报道的抑制erk信号通路的mce3e蛋白不属于一个操纵子,并且不是同源蛋白,两者序列的相似度只有47.34%。mce3e只抑制erk信号通路,与erk相互作用的是fxfp模序;mce2e除了抑制erk信号通路,对jnk信号通路也有抑制作用,且作用位点是“rtlasvalil”模序。但该模序并不与p38蛋白结合。
实施例5mce2e蛋白与erk1/2和jnk蛋白共定位
(a)mce2e和erk2在细胞中的共定位
1)构建含有erk2基因的重组质粒p3xflag-cmv14-erk2;
2)通过lipofectamine2000(invitrogen,carlsbad,ca)转染法将重组质粒p3xflag-cmv14-erk2和pegfp-n1-mce2e共转染到铺在盖玻片的hela细胞中;
3)转染6h后,更换新鲜去血清的dmem培养基继续培养;
4)24h后用于flag染色的细胞直接固定、透化和封闭;
5)封闭完以后的细胞用flag抗体(cellsignalingtechnology)室温孵育1h;
6)pbs洗涤细胞三遍后用alexa594红色荧光二抗室温孵育1h,之后pbs洗涤细胞三遍并用含有dapi的封片剂封片并用于激光共聚焦显微镜镜检;
(b)mce2e和jnk1在细胞中的共定位
1)构建含有jnk1基因的重组质粒p3xflag-cmv14-jnk1;
2)通过lipofectamine2000(invitrogen,carlsbad,ca)转染法将重组质粒p3xflag-cmv14-jnk1和pegfp-n1-mce2e共转染到铺在盖玻片的hela细胞中;
3)转染6h后,更换新鲜去血清的dmem培养基继续培养;
4)24h后用于jnk1染色的细胞直接固定、透化和封闭;
5)封闭完以后的细胞用flag抗体(cellsignalingtechnology)抗体室温孵育1h;
6)pbs洗涤细胞三遍后用alexa594红色荧光二抗室温孵育1h,之后pbs洗涤细胞三遍并用含有dapi的封片剂封片并用于激光共聚焦显微镜镜检;
(c)实验结果分析:如图4,在非活化的hela细胞中,mce2e和erk2有明显的共定位现象(见图4a),mce2e和jnk1也有显著的共定位现象(见图4b)。未转染mce2e时,erk2和jnk1蛋白处于弥散状态,过表达mce2e后,mce2e将erk和jnk蛋白共定位在内质网中。以上结果说明mce2e通过空间调控阻止erk和jnk蛋白入核发挥作用。
虽然本发明已以较佳实施例公开如上,但其并非用以限定本发明,任何熟悉此技术的人,在不脱离本发明的精神和范围内,都可做各种的改动与修饰,因此本发明的保护范围应该以权利要求书所界定的为准。
sequencelisting
<110>中国科学院微生物研究所
<120>一种erk和jnk信号通路抑制剂mce2e
<160>2
<170>patentinversion3.3
<210>1
<211>402
<212>prt
<213>结核分枝杆菌
<400>1
metargcysglyvalseralaglyseralaasnglylysproasnarg
151015
trpthrleuargcysglyvalseralaglyhisargglyservalphe
202530
leuleualavalleuleualaprovalvalleuthrsercysthrtrp
354045
argglyilealaasnvalproleuprovalglyargglymetglypro
505560
aspargmetthriletyrvalglnmetproaspthrleualaleuasn
65707580
thrasnserargvalargvalalaaspvaltrpvalglythrvalarg
859095
aspileserleuargasntrpilealathrleuthrleugluleuglu
100105110
prothrvalargleuproalaasnalathralalysileglyglnthr
115120125
serleuleuglythrglnhisvalgluleualaalaproproilepro
130135140
serproglnproleulysserglyaspthrileglyleulysasnser
145150155160
seralatyrprothrvalgluargthrleualaservalalaleuile
165170175
leuthrglyglyglyilevalasnleuaspvalileglnthrgluile
180185190
leuasnileleuaspglyhisalaglyglnileargglupheleuglu
195200205
argleualathrphethralagluleuasnasnglnargglyaspleu
210215220
thrargalaileaspserthrasnglnleuleuthrileilealaasn
225230235240
argasnaspthrleuaspargvalleuthraspvalproproleuile
245250255
gluhisphealaaspthrglyglnleuphealaaspalathrgluser
260265270
leuglyargphesergluvalalaasnargalaleualaalathrarg
275280285
proasnleuhisglnthrleuglnserleuglnargproleuarggln
290295300
leugluargalaserprotyrvalvalglyalaleulysleuglyleu
305310315320
thralapropheasnileaspgluvalproasnvalileargglyasp
325330335
tyrvalasnvalseralathrpheaspvalthrleuseralaleuasp
340345350
asnalaleuleuserglythrglyileserglymetleuargalaleu
355360365
gluglnalatrpglyargaspproaspthrmetileproaspvalarg
370375380
tyrthrproasnproasnaspalaproglyglyproleuvalgluarg
385390395400
alaglu
<210>2
<211>10
<212>prt
<213>结核分枝杆菌
<400>2
argthrleualaservalalaleuileleu
1510
- 还没有人留言评论。精彩留言会获得点赞!