一种与棉纤维发育相关的WRKY转录因子基因及其应用的制作方法
本发明属于基因工程领域,具体涉及一种与棉纤维发育相关的wrky转录因子基因及其应用。
背景技术:
棉花是世界上最主要的农作物之一,也是我国最大的经济作物。棉花是关系着国计民生的特殊商品,在我国国民经济中占有重要地位。棉纤维作为重要的天然纤维,是纺织工业的主要原材料,其产量和纤维品质直接决定了棉花的商业价值。随着世界纺织工业的发展,对棉纤维强度、细度和成熟度等内在品质的要求更高,然而目前大多国产原棉的棉纤维长度和强度都达不到纺织工业的要求。因此,提高棉纤维的产量,改良棉纤维的品质,一直是棉花产业发展的研究方向和育种目标。
棉花纤维细胞的形态发生包含了一个高度调控的细胞分化过程。每个纤维细胞都是由胚珠表面的外表皮细胞分化而来的单细胞毛状突起。纤维发育经历几个连续又重叠的阶段,即纤维细胞的起始、伸长、次生壁增厚和脱水成熟。棉纤维细胞壁持续约50天的形态发生过程决定了棉纤维的品质参数。胚珠表皮细胞纤维的起始数目决定成熟纤维的产量,纤维的伸长和次生壁的增厚程度决定成熟纤维的长度和强度。因此,深入研究棉纤维发育过程及调控机理,可望为提高棉纤维产量和品质开辟新的途径。
棉花纤维发育过程受转录水平的调控。目前已发现的主要转录因子家族hd-zip蛋白、myb蛋白、nac蛋白等都在纤维细胞起始或发育过程中起到至关重要的作用。wrky转录因子是近年来在植物中发现的特有新型转录调控因子,因在其n端含有由wrkygqk组成的高度保守的氨基酸序列而得名。已有研究表明,wrky转录因子在植物的生长发育、衰老、生物和非生物胁迫应答中发挥非常重要的作用。但是对于wrky转录因子是否参与调控棉花纤维发育还尚无报道。因此,阐明wrky转录因子在棉纤维发育中的作用以及调控机理,分离鉴定相关的重要调控基因,对于棉花纤维品质改良,具有重要意义。
wrky蛋白家族属于锌指型转录调控因子,最主要的结构特点是其dna结合域中至少含有一个wrky结构域,该结构域是一段由大约60个高度保守的氨基酸残基所组成的多肽序列,在其n端有一个具有高度保守的7肽区域(wrkygqk),为其核心区域,其c末端为一个锌指结构(zincfingermotif),一般组成为cx4~5cx22~23hx1h。依据wrky结构域中wrky基序数目和锌指结构特征的差异,将wrky家族分成3类:第i类成员含有2个wrky基序和c2h2型锌指结构,第ii类成员含有1个wrky基序和c2h2型锌指结构,第iii类成员则含有1个wrky基序和c2hc型锌指结构。研究发现,大多数wrky转录因子都只含有1个wrky结构域,属于ⅱ类,而ⅲ类wrky转录因子只存在于高等植物中,在一些低等植物如苔藓植物中却不存在。同时,高等植物中几乎所有的ⅲ类wrky转录因子都与植物的生物胁迫应答反应有关,这说明ⅲ类wrky转录因子可能是由于植物受环境压力通过适应性进化所产生。
wrky蛋白能特异的与dna结合区的(t)(t)tgac(c/t)序列结合,这一结合区称为w-box,其中包含一个tgac保守核心序列,它对于wrky转录因子的结合必不可少。此外,zn2+等金属离子的参与和磷酸化作用等也利于wrky转录因子与w-box的结合。大多数wrky转录因子的目标基因启动子中都含有数量不定的w-box,它们之间或同向排列或形成回文结构。wrky转录因子与功能基因或其他调控因子基因启动子中存在的w-box结合,调节这些下游基因的表达。
wrky转录因子参与调控植物的一些生长发育过程。如大豆gmwrky13和拟南芥atwrky75与侧根的生长有关;水稻oswrky31也与侧根的形成和延伸有关。拟南芥atwrky6和水稻oswrky23可以引发衰老的进程,而atwrky70可以负调控衰老反应。拟南芥atttg2是一个编码wrky转录因子的基因,它在拟南芥表皮毛状体以及种皮的形成中起了重要的作用。另外,wrky转录因子还参与了植物的一些新陈代谢过程,如大麦susiba2参与了体内的糖代谢,棉花gawrky1调控倍半烯的生物合成。水稻oswrky71以及大麦hvwrky38可以抑制糊粉细胞中ga诱导的α-淀粉酶的活性,从而抑制种子的萌发。但是,wrky转录因子在棉花纤维发育过程中的调控功能至今未见研究报道。因此,研究阐明wrky转录因子在棉纤维发育中的调控作用,具有重要的理论和实践意义。
技术实现要素:
本发明针对现有技术的不足,目的在于提供一种与棉纤维发育相关的wrky转录因子基因及其应用。
本发明提供与棉纤维发育相关的wrky转录因子基因,其具有:
1)核苷酸序列如seqidno.1所示;或
2)如seqidno.1所示核苷酸序列经取代、缺失和/或增加一个或几个核苷酸;或
3)在严格条件下与1)限定的dna序列杂交的核苷酸序列。
本发明还提供与棉纤维发育相关的wrky转录因子基因的cdna序列,核苷酸序列如seqidno.2所示。
本发明还提供与棉纤维发育相关的wrky转录因子基因编码的蛋白质,氨基酸序列如seqidno.3所示。
上述与棉纤维发育相关的wrky转录因子基因在改良棉纤维产量和品质方面的应用。
本发明的有益效果:本发明提供了一个新的与棉纤维发育相关的ghwrky16基因序列及cdna序列,该基因在棉纤维中高水平表达,表明该基因可能在调控棉纤维起始和伸长发育过程中起重要作用;在ghwrky16rnai转基因棉花中,在棉纤维发育过程中发挥重要作用的ghhox3和ghmyb109等重要调控基因的表达下调,纤维素合酶基因表达量也下降;与野生型相比,ghwrky16rnai转基因棉花成熟纤维长度显著变短,而且短绒较少或缺失;ghwrky16蛋白定位于细胞核中,能够直接与ghhox3和ghmyb109基因启动子上的w-box顺式元件结合,这表明ghwrky16基因可能通过调控棉纤维相关的下游靶基因来调节棉纤维细胞起始和伸长发育过程,影响棉纤维产量和品质。
附图说明
图1为荧光定量rt-pcr分析ghwrky16在棉花组织中的表达,其中r:根;l:叶片;c:子叶;h:下胚轴;p:花瓣;a:花药;f:纤维;0d:开花当天胚珠;3d:开花后3天纤维;6d:开花后6天纤维;9d:开花后9天纤维;12d:开花后12天纤维;15d:开花后15天纤维;18d:开花后18天纤维,误差线代表标准误差。
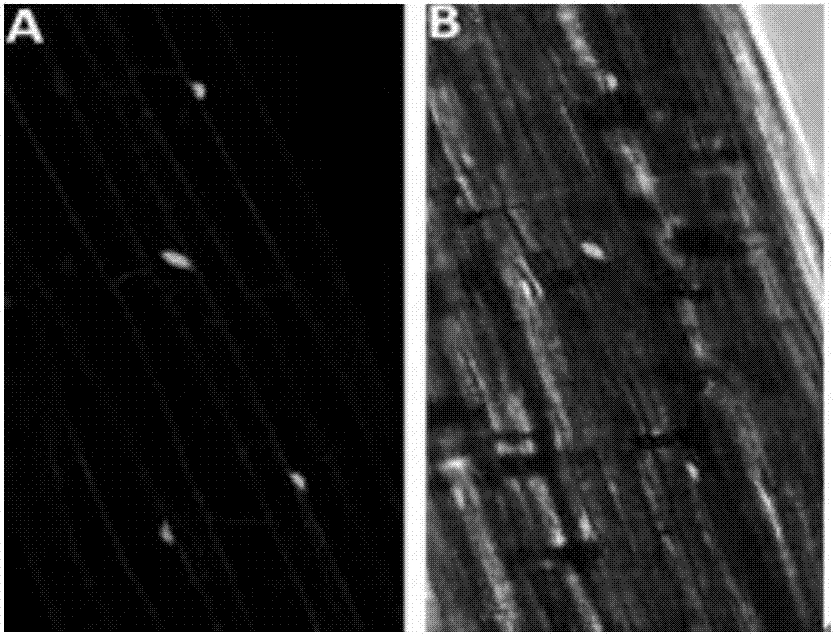
图2为ghwrky16蛋白在拟南芥下胚轴细胞中的亚细胞定位,其中a:拟南芥植株下胚轴细胞中ghwrky16-gfp荧光信号;b:gfp荧光图像与同一细胞的明视野图像的重合。
图3为ghwrky16转基因棉花表型分析,其中a:荧光定量rt-pcr分析ghwrky16基因在野生型(wt)和转基因棉花株系(l1~l8)开花9天后的纤维中的表达;b:野生型和ghwrky16rnai转基因棉花株系(l1、l3、l5)开花当天的胚珠体外离体培养;c:野生型和ghwrky16rnai离体胚珠培养12天、15天和18天后纤维长度统计分析(n>30);d:野生型和ghwrky16rnai转基因棉花成熟纤维长绒和短绒形态比较;e:野生型和ghwrky16rnai转基因棉花成熟纤维长度测量及统计分析(n>50);f:野生型和ghwrky16rnai转基因棉花开花当天(0d)及开花后1天(1d)、2天(2d)胚珠横切片,显示开花0天、1天、2天后的纤维细胞突起分化情况;误差线代表标准误差,*表示转基因与野生型在t-检验中p值<0.05,存在显著差异;**表示转基因与野生型在t-检验中p值<0.01,存在极显著差异。wt,野生型;l1~l8,ghwrky16-rnai转基因棉花株系。
图4为纤维素合酶ghcesa3、ghcesa5、ghcesa10在野生型(wt)及ghwrky16rnai转基因棉花(l1、l3、l5)开花后9天的纤维中的表达情况。wt,野生型;l1,l3和l5,ghwrky16-rnai转基因棉花株系。
图5为与棉纤维发育相关基因ghhox3、ghrdl1、ghexp1以及ghmyb109在野生型(wt)和ghwrky16rnai转基因棉花(l1、l3、l5)开花后9天的纤维中的表达情况。wt,野生型;l1,l3和l5,ghwrky16-rnai转基因棉花株系。
图6为ghwrky16蛋白与ghhox3和ghmyb109启动子结合的emsa分析。
具体实施方式
为了更好地理解本发明,下面结合实施例进一步阐明本发明的内容,但本发明的内容不仅仅局限于下面的实施例。
实施例1棉花wrky家族基因ghwrky16全长序列的克隆鉴定和功能分析
1.ghwrky16基因及cdna序列的分离鉴定:
在棉花基因组及est数据库中筛查相关的ghwrky基因及cdna序列,鉴定了22个棉花wrky基因,其中有一个基因(命名为ghwrky16)在棉纤维中优势表达,通过dna和蛋白质序列分析,利用棉花纤维细胞的转录本(总rna),分离获得ghwrky16全长cdna序列。按照ghwrky16cdna序列设计引物,以棉花基因组dna为模板,采用pcr技术扩增获得该基因的dna全长序列。序列分析表明,ghwrky16基因全长序列(如seqidno.1所示)包含5’-上游启动子区(1~2000bp)、3个外显子、2个内含子(下划线)和3’-utr(3527–3649bp);ghwrky16基因的cdna序列如seqidno.2所示,其中含有957bp开放阅读框(从起始密码子atg到终止密码子taa),编码318个氨基酸(氨基酸序列如seqidno.3所示),分子量为34kda。ghwrky16蛋白dna结合域中含有一个wrky结构域,该结构域的n端有一个具有高度保守的wrkygqk7肽区域,为其核心区域,其c末端锌指基序为c2h2,属于二类wrky转录因子。
2.定量rt-pcr分析ghwrky16基因的表达:
利用rna提取试剂盒(qiagin)提取纯化棉花不同组织和纤维的总rna,实时荧光定量rt-pcr研究基因的表达(按照lixb,fanxp,wangxl,cail,yangwc,2005.thecottonactin1geneisfunctionallyexpressedinfibersandparticipatesinfiberelongation.plantcell17:859–875进行)。首先,将棉花不同组织(根、叶片、子叶、下胚轴、花瓣、花药、开花后0天胚珠、开花后3、6、9、12、15和18天纤维等)的总rna(2μg/样)用m-mlvrnaseh-reversetranscriptase(promega)反转录成cdna;然后,以cdna为模板,用基因特异的引物(ghwrky16rtp1:5’-gaagtgtaattccatggatg-3’和ghwrky16rtp2:5’-tgactaatctgaacaaccct-3’)和real-timepcrmastermix(toyobo,japan)进行定量pcr反应。棉花ghubi1基因作为rt-pcr反应的内标,目标基因每一个循环的扩增都被sybr-green荧光检测,计算目标基因的表达水平相对值。每一实验重复3次,统计分析实验结果。荧光定量rt-pcr结果显示ghwrky16在棉花各组织和棉纤维不同发育阶段均有不同程度的表达,在开花后3~9天的棉纤维中优势表达(图1),推测该基因可能在伸长期的棉纤维细胞中发挥调控作用。
3.ghwrky16蛋白的细胞亚定位分析
构建ghwrky16:egfp植物表达载体,将该载体通过电转化法转入gv3101农杆菌中,通过浸花法转化拟南芥,收获成熟拟南芥的t0代种子,筛选阳性植株。将转基因拟南芥t2代种子在含卡那霉素的ms培养基上萌发生长5天后,置于共聚焦显微镜下观察转基因拟南芥小苗的下胚轴细胞的绿色荧光蛋白(gfp)的荧光信号。亚细胞定位分析结果显示,gfp荧光信号主要分布在下胚轴细胞的细胞核中,ghwrky16蛋白位于细胞核中(图2)。
4.ghwrky16rnai转基因棉花表型分析:
构建35s启动子驱动的pmd-ghwrky16rnai载体,转化农杆菌lba4404。利用农杆菌浸染棉花下胚轴外植体,共培养2天后,将下胚轴转移到选择培养基上培养,诱导并筛选到转化愈伤组织。将该愈伤组织经过8~10个月的继代培养后,诱导分化出胚性愈伤组织和体细胞胚,体细胞胚进而萌发再生为转基因棉花幼苗。将棉花幼苗移栽至土中,生长发育至开花结实。提取棉花基因组dna,利用pcr技术检测鉴定转基因棉花植株。
(1)实时荧光定量rt-pcr研究基因的表达:按照lixb,fanxp,wangxl,cail,yangwc,2005.thecottonactin1geneisfunctionallyexpressedinfibersandparticipatesinfiberelongation.plantcell17:859–875进行,提取开花后9天的纤维rna,逆转录成cdna后,以cdna为模板,用基因特异的引物(ghwrky16rtp1和ghwrky16rtp2)和real-timepcrmastermix(toyobo,japan)进行定量pcr反应。以ghubi1做为内参分析ghwrky16在野生型(wt)及转基因植株各株系中的表达情况。重复3次,统计分析实验结果。
(2)离体胚珠培养实验:选取开花当天的野生型(wt)及3个ghwrky16rnai转基因棉花株系(l1、l3和l5)的胚珠置于bt培养基中分别无菌培养(暗处理,30℃)12天、15天和18天后,用固定液固定后测量胚珠表面纤维长度进行统计分析,误差线代表标准误差,*表示转基因与野生型在t-test中p值<0.05存在显著差异,**表示转基因与野生型在t-test中p值<0.01存在极显著差异。
(3)成熟棉纤维统计测量:纤维成熟后,对野生型(wt)及ghwrky16rnai转基因植株各株系成熟纤维长度进行测量与统计(n>50)。ck为两个分离出来的非转基因植株,剩下都为不同的ghwrky16rnai转基因株系。误差线代表标准误差,*表示转基因与野生型在t-test中p值<0.05存在显著差异,**表示转基因与野生型在t-test中p值<0.01存在极显著差异。
(4)胚珠徒手切片:取野生型(wt)及ghwrky16rnai转基因植株开花当天(0天)、1天和2天的胚珠进行徒手切片,切成约1mm的薄片在显微镜下观察并拍照。
本发明构建了ghwrky16rna干扰(rnai)载体,转化棉花,获得了转基因棉花植株。首先对ghwrky16转基因植株进行rna水平上表达量的检测,结果显示在大多数ghwrky16rnai转基因植株中,ghwrky16基因的表达量均大幅度下调(图3a)。ghwrky16rnai转基因植株的棉桃相较与野生型要小一些。对开花当天的ghwrky16rnai转基因棉花及野生型胚珠进行体外培养,12~18天后观察发现,转基因棉花胚珠发育迟缓,纤维长度明显短于野生型(图3b、c)。对rnai转基因植株成熟棉纤维长度进行测量分析,结果表明,大多数ghwrky16rnai转基因棉花植株的成熟纤维长度均显著短于野生型(图3e)。而且,剥除长纤维后,发现rnai转基因棉花种子上的短绒也变少,一些株系的种子表面几乎没有短绒(图3d)。上述结果表明,ghwrky16表达下调可能影响棉花纤维发育过程,导致成熟棉纤维长度变短,甚至短绒缺失现象。为进一步研究ghwrky16是否影响棉纤维细胞的起始分化,对开花当天、开花后1天和2天的胚珠进行徒手切片,在显微镜下观察胚珠表面纤维细胞突起的数目及排列方式是否发生变化,发现rnai转基因棉花胚珠表面的纤维细胞突起明显减少,纤维细胞伸长发育迟缓(图3f)。这说明ghwrky16在棉纤维发育的起始和伸长阶段都发挥重要作用。
5.ghwrky16rnai转基因棉花中纤维素合酶及其他相关基因表达分析
按照lixb,fanxp,wangxl,cail,yangwc,2005.thecottonactin1geneisfunctionallyexpressedinfibersandparticipatesinfiberelongation.plantcell17:859–875进行。提取野生型(wt)及ghwrky16rnai转基因棉花植株开花后9天的纤维rna,以ghubi1为内参进行实时定量pcr分析纤维素合酶以及棉纤维发育相关基因的表达。误差线代表标准误差,*表示转基因与野生型在t-test中p值<0.05存在显著差异,**表示转基因与野生型在t-test中p值<0.01存在极显著差异。
本发明分析了ghwrky16rnai转基因棉花中ghcesa3、ghcesa5和ghcesa10等纤维素合酶基因的表达情况(图4)。结果显示,与野生型相比,在ghwrky16下调比较显著的两个转基因株系l1、l3、l5中,ghcesa3、ghcesa5和ghcesa10的表达量明显下调。同时对这三个基因的启动子进行分析时发现,ghcesa3和ghcesa5的启动子中均有wrky家族特异性结合元件“w-box”。我们推断ghwrky16可能通过与ghcesa3和ghcesa5的启动子中的“w-box”元件结合而调控它们的表达,从而影响棉花纤维的早期发育。
6.emsa实验分析ghwrky16蛋白与ghhox3、ghmyb109启动子上w-box元件的结合
在大肠杆菌中表达纯化带有his标签的ghwrky16蛋白,从ghhox3和ghmyb109启动子上选取含有w-box的dna序列片段,合成带有生物素标记探针。以带有生物素标记的上述dna序列为探针,以未标记的同一dna序列作为冷探针竞争,进行emsa实验。结果显示,ghwrky16蛋白能够与ghhox3和ghmyb109启动子上的w-box顺式元件结合,加入未标记的探针能够减弱所标记探针的荧光强度,使emsa实验中的结合条带减弱或消失。这表明ghwrky16可能通过调控下游ghhox3和ghmyb109基因的表达来调节棉纤维发育。
ghhox3在棉纤维发育初期表达量较高,且通过调控其下游的靶基因ghexp1以及ghrdl1的表达来影响棉纤维伸长。ghmyb109在起始和伸长的棉花纤维中表达量较高且调控棉纤维发育,在ghmyb109表达下调的转基因棉花中成熟纤维长度变短。因此分析了这些基因在ghwrky16rnai转基因棉花纤维中的表达情况(图5)。结果显示,与野生型相比,ghhox3、ghrdl1、ghexp1及ghmyb109在ghwrky16rnai转基因植株中的表达量均明显下降。分析它们的启动子序列发现,只有ghhox3及ghmyb109的启动子序列中含有wrky家族特异性结合元件“w-box”。因此,我们推测ghwrky16可能通过与ghhox3、ghmyb109启动子区域的“w-box”结合来调控其表达,从而影响棉纤维发育。进一步实验证实,ghwrky16转录因子能够与ghhox3、ghmyb25和ghmyb109基因的启动子结合。如图6所示,esma实验结果显示ghwrky16蛋白可以与ghhox3和ghmyb109的dna探针结合形成复合物,且随着竞争性探针的增加,检测到的结合复合物减少或消失。上述实验结果表明,ghwrky16可能通过调控下游ghhox3和ghmyb109等来调节棉纤维发育过程,影响棉纤维产量和品质。
显然,上述实施例仅仅是为清楚地说明所作的实例,而并非对实施方式的限制。对于所属领域的普通技术人员来说,在上述说明的基础上还可以做出其它不同形式的变化或变动。这里无需也无法对所有的实施方式予以穷举。而因此所引申的显而易见的变化或变动仍处于本发明创造的保护范围之内。
序列表
<110>华中师范大学
<120>一种与棉纤维发育相关的wrky转录因子基因及其应用
<160>3
<210>1
<211>3649bp
<212>dna
<213>棉花ghwrky16基因序列
<400>1
gagtttaatttttattttgagtataaaataattttaaaattttttattaacatttacctc60
ttaataaattaataaaatacgaaaaattaattattaaatttaccccaagaaataatttaa120
aatataaaaaaattcgaactgtttatattaaatcaaaatggctaagggttccactttttt180
gaggaaaaaaaaagagaatcccatgtgacttgtgggatgagactttgtcacattgaccgc240
cgtttccagtttccggtgaaaaagtagatcggggccccactttttattgtgggtcccact300
aataactttaaaatttattattgtttattaattttgtttttaaggtcaactctaatcact360
gtctccaactccaacctaaaataaataaataaaaggtttaatttttttaaaattataata420
atagagtaaagttcgaataacaaacacgattagctcgatttaaatataggctatacttga480
ggcggtgaacacccttagcaatcacatgggattcaaataaaaggtttaattttataatcc540
tatttttaaattttaaaatattataattgagttattaaagtatgaattttttttgtcaat600
ttaatcataaattcgagtattaaatgttattttgaatgtgacgtggaatataactaaaac660
acgtaacataaattattaaatgaagtttgatacgattggtaaagaagaagattcaagacc720
taccacatgtaaaagttaaattcgagtactaaatgttattttgaaagtgacgtaaaatat780
aattaaaatacgtaaattatttaatgaagttttggcatgattggtaaagaggaagactta840
ctgcatgcaaaagttagatttgagtattaaatgttattttgattgtgacgtgaaatataa900
ttaaaacacgtaaattattaaatgaataatttgcacgaatttttgagtctcatggtaaag960
ttttagtccagttgtcagttttagcccgaatcgtattagctcttaatctacttgtgttgt1020
aataatttgatcctattttgtgattagttcataattgtattgtatttgtaatacttaaaa1080
aaacacgtaatttaaattgtaggcaagtgtaaaatcaacccatacttagatctaacctgt1140
taaaaaaatttaacatcaataaattttaagaaaattatttatttatttttgaacacgata1200
tattcaaattttgtatgtaactgttttttcgtttttaaaatgttttcaaatatctttcgt1260
gttaaatttaaattggcatttaatatctacattaattattaagttaccaaaatgatggga1320
caacaaattaaaattatgactaaataagaaaaattaagaaatcaaaaatcgatacaaatt1380
aggtaaaattactacaaaactccttatatcatacacacactagatgagttagcataacaa1440
tttatctgttaaaatttggtgtttatatataaagtataataatatatttagctatcaact1500
tttacatatttataaaatttagcctatactcttttattgggtcgatattattattttaac1560
agaagattgttttactcacgcatccaatgtaaatagaatactttatttattttttaaata1620
aagaaaataaaatgcaattcttggtacagtttttagaatacaattaaaaagaaatcagaa1680
tttatttatgagaaattaatattaaatatttttttagaaaaaaaatcccaaatttacaga1740
gtcatcaatttcaaacggctaatccccaccttttgaccttatctcttgtcagtgacccaa1800
tgagaaaggcatccctttgacgaaaactcgaatctctttcatcataaatcataatcaatc1860
tttcactttttcttcgtaattaaatctcaattcatttcagttcccccctctttgttctgc1920
ttcatctatataataaactctgcttttatttgggagtttctcaaaagggctgggtgttct1980
ttgttttcgtatgagcagagatgacggtggaattgatgatgggtcacggcggcgttggcg2040
gtggaggcaacagttttacagtcaaaatggaggaaactgctttgaaagaagctgctaatg2100
ctgggattcaaggttttgaagaactgataagattgatgtctaaaagtcaacagcttcgct2160
cgcaagatgtttgttttaatgccccttcaagttcagaaccagctatggaaatccaagctg2220
ttacagataagactgtaagttcctttaaaaaagttatttcgttattgggtcgacctagaa2280
ctggtcatgctcggttcagacgagctcctcttaatcatctcccacaacaacaacagattc2340
aagtgagcaaagatgaagaagtatcgacttttaagcctttatgttcaaccccaagttata2400
agttacctcctttgccaaccaaaagtagccatgttttaaagactggtgttttagagcctg2460
ggaattctgttatgtcttcattgactggggatactgatagcatacaacaccaaccgtgtt2520
tctctattggttttcaattcactaacccttcttctcatggcaaacctcctttgtcttcaa2580
aaaggaagtgtaattccatggatgatgttgctaatctcaagtgtggatcatcatcctctg2640
ctcgttgccattgttccaagaagaggttggtccttggtgcttggttcttcactttttgat2700
tgtgatctttgatggtttcatttgattttgattatgtatgtgttcatgtaggaaatcaag2760
agtgaaaagagtcattagagtcccagctgttagcaacaaaatggctgatattcctcacga2820
tgattattcttggagaaaatatggccaaaaaccaatcaaaggttctcctcatccaaggtg2880
tgttttttcgattaaatttcgaatggattctaatagagtgaattctacaccaaagtttca2940
atttttttgaagttgactaaagggttccaaagttttcaaacaataattcatgtatgtatt3000
gaatgcctttttgtttggttttcaactacaaacattaatcatattctatttggcaaaaaa3060
ttaattgttaatttgactttttgcggggaagtgcatggagacttttgaattagaaattct3120
ttttactagaaaaatagataaattagtatttagattaaagagtaaatttgtatgggatga3180
aatttttaataaaaataattaatttactcttttatttaggactaatttattaaattttca3240
taatagagatttaaaatacaatagtatttgtattataacattgaattattgccttactac3300
cgagttggttcattgagttcgcttttttaactcagtcccatatgttttaacaaatttcag3360
gggctattacaaatgtagcagtgtcagaggttgtcctgcaaggaagcatgtagaaagagc3420
tgtcgaggatccaaggatgttgattgtaacatacgaaggtgatcacaatcatagccataa3480
tatcaccacagatgtcccatcggcgctagttcttgaatcatcataaagatcatcccatcc3540
aacggctaaaatatctatagggttgttcagattagtcaatttagatccataggttctgtc3600
tccaacaagacaaaagaagattgattagtgggtaagtttagctgtttct3649
<210>2
<211>1109bp
<212>dna
<213>棉花ghwrky16基因的cdna序列
<400>2
atgacggtggaattgatgatgggtcacggcggcgttggcggtggaggcaacagttttaca60
gtcaaaatggaggaaactgctttgaaagaagctgctaatgctgggattcaaggttttgaa120
gaactgataagattgatgtctaaaagtcaacagcttcgctcgcaagatgtttgttttaat180
gccccttcaagttcagaaccagctatggaaatccaagctgttacagataagactgtaagt240
tcctttaaaaaagttatttcgttattgggtcgacctagaactggtcatgctcggttcaga300
cgagctcctcttaatcatctcccacaacaacaacagattcaagtgagcaaagatgaagaa360
gtatcgacttttaagcctttatgttcaaccccaagttataagttacctcctttgccaacc420
aaaagtagccatgttttaaagactggtgttttagagcctgggaattctgttatgtcttca480
ttgactggggatactgatagcatacaacaccaaccgtgtttctctattggttttcaattc540
actaacccttcttctcatggcaaacctcctttgtcttcaaaaaggaagtgtaattccatg600
gatgatgttgctaatctcaagtgtggatcatcatcctctgctcgttgccattgttccaag660
aagaggaaatcaagagtgaaaagagtcattagagtcccagctgttagcaacaaaatggct720
gatattcctcacgatgattattcttggagaaaatatggccaaaaaccaatcaaaggttct780
cctcatccaaggggctattacaaatgtagcagtgtcagaggttgtcctgcaaggaagcat840
gtagaaagagctgtcgaggatccaaggatgttgattgtaacatacgaaggtgatcacaat900
catagccataatatcaccacagatgtcccatcggcgctagttcttgaatcatcataaaga960
tcatcccatccaacggctaaaatatctatagggttgttcagattagtcaatttagatcca1020
taggttctgtctccaacaagacaaaagaagattgattagtgggtaagtttagctgtttct1080
aaaaagtaaaaaaaaaaaaaaaaaaaaaa1109
<210>3
<211>318
<212>prt
<213>棉花ghwrky16基因编码的蛋白质
<400>3
metthrvalgluleumetmetglyhisglyglyvalglyglyglyglyasnserphethr
15101520
vallysmetglugluthralaleulysglualaalaasnalaglyileglnglypheglu
25303540
gluleuiieargleumetserlysserglnglnleuargserglnaspvalcyspheasn
45505560
alaprosersersergluproalametgluiieglnalavalthrasplysthrvalser
65707580
serphelyslysvaliieserleuleuglyargproargiieglyhisalaargphearg
859095100
argalaproleuasnhisleuproglnglnglnglniieglnvalserlysaspgluglu
105110115120
valserthrphelysproleucysserthrprosertyrlysleuproproleuprothr
125130135140
lysserserhisvalleulysthrglyvalleugluargglyasnservalmetserser
145150155160
leuthrglyaspthraspseriieglnhisglnprocyspheserleuglypheglnphe
165170175180
thrasnproserserhisglylysproproleuserserlysarglyscysasnsermet
185190195200
aspaspvalalaasnleulyscysglyserserserseralaargcyshiscysserlys
205210215220
lysarglysserargvallysargvaliieargvalproalavalserasnlysmetala
225230235240
aspiiephohisaspasptyrsertrparglystyrglyglnlysproiielysglyser
245250255260
prohisproargglytyrtyrlyscysserservalargglycysproalaarglyshis
265270275280
valgluargalavalgluaspproargmetleuiievalthrtyrgluglyasphisasn
285290295300
hisserhisasniiethrthraspvalproseralaleuvalleugluserser
305310315318
- 还没有人留言评论。精彩留言会获得点赞!
- 大豆威廉姆斯82中NAC膜结合转录因子基因GmNTL1及其应用的制作方法与工艺
- 一种锌指蛋白转录因子基因RkMSN4及其应用的制作方法与工艺
- 三七MYB转录因子基因PnMYB2及其应用的制作方法与工艺
- WRKY转录因子在制备抗逆转基因甜橙中的应用的制造方法与工艺
- 能够提供热耐受性的转录调节因子的表达的制造方法与工艺
- 具有用于舒适性、空间调节和个性化舒适性基因的辅助空气源的空气悬浮座椅的制造方法与工艺
- 一种大豆转录因子GmDISS1及其编码基因与应用的制造方法与工艺
- 一种与棉纤维发育相关的WRKY转录因子基因及其应用的制造方法与工艺
- 一个棉花产量性状关联的乙烯响应转录因子基因的制造方法与工艺
- 一种NF‑Y类核转录因子基因ZmNF‑YA13、其编码的蛋白及其应用的制造方法与工艺