茶树开花基因CsFT及其编码蛋白的制作方法
本发明属于分子生物学技术领域,具体涉及茶树开花基因csft及其编码蛋白。
背景技术:
在上个世纪,经过对植物开化机制的长时间研究,人们发现植物花芽形成的是受叶片中产生的一种激素样物质诱导的。该类物质在成熟叶片中产生,然后被传输到顶芽执行花芽诱导的功能。近年来分子生物学及生物化学领域对这一促进开花的物质进了深入研究,结果发现,这种促进开花的物质为一种floweringlocust(ft)基因编码的蛋白。ft蛋白在植物生长发育中具有多重功能,不仅作为一个具有可转移性的开花诱导信号,同时还参与季节性生长停止及接下来的休眠调控。茶树是常绿木本植物,冬季会形成休眠芽以应对低温胁迫。而且绝大多数的茶树品种为开花品种,从花芽形成到种子成熟要经历大约一年半的时间。可见与一般的木本植物不同,茶树在一年中都在进行着生殖发育。因此茶树的生长-休眠循环及生殖发育在茶树生命循环中至关重要,同时还直接影响着茶叶生产。同时部分茶树资源的果实可以用来榨油,是良好的食用油来源,具有很好的保健功能。但是目前还没有关于茶树csft基因序列、蛋白序列及其功能的相关专利报道、发表文献。
技术实现要素:
针对现有技术存在的问题,本发明的目的在于设计提供茶树开花基因csft及其编码蛋白的技术方案。
所述的茶树开花基因csft,其特征在于该基因具有两个转录本csfta和csftb,转录本csfta具有如seqidno.1所示的核苷酸序列或该序列经替换、缺失或添加一个或几个核苷酸形成的具有同等功能的核苷酸序列,所述的转录本csftb具有如seqidno.2所示的核苷酸序列或该序列经替换、缺失或添加一个或几个核苷酸形成的具有同等功能的核苷酸序列。
所述的茶树开花基因csft在诱导植物早花性状的用途。
所述的含有茶树开花基因csft转录本的载体。
所述的含有茶树开花基因csft转录本的载体的宿主。
所述的茶树开花基因csft的转录本csfta,其特征在于该转录本csfta的核苷酸序列如seqidno:1所示。
所述的茶树开花基因csft的转录本csfta的编码蛋白,其特征在于该蛋白的氨基酸序列如seqidno:3所示。
所述的茶树开花基因csft的转录本csfta及其编码蛋白在诱导植物早花性状的用途。
所述的茶树开花基因csft的转录本csftb,其特征在于该转录本csftb的核苷酸序列如seqidno:2所示。
所述的茶树开花基因csft的转录本csftb的编码蛋白,其特征在于该蛋白的氨基酸序列如seqidno:4所示。
所述的茶树开花基因csft的转录本csftb及其编码蛋白在抑制短日照引起的植物生长停滞和休眠芽形成的用途。
本发明提供一种茶树开花基因csft转录本csfta、转录本csftb及其编码蛋白,并利用该基因构建植物表达载体,经农杆菌转化不同遗传背景的杨树,获得的转基因杨树植株均表现显著的早花现象,表明该基因具有诱导杨树早花的功能,将csft基因的植物表达载体应用于茶树,可以加快茶树开花,缩短茶树生育期,促进茶树遗传改良和品种选育。
附图说明
图1为csfta及csftb基因的编码区序列及所编码氨基酸,图1中虚线框显示csfta及csftb基因编码的第100位氨基酸。
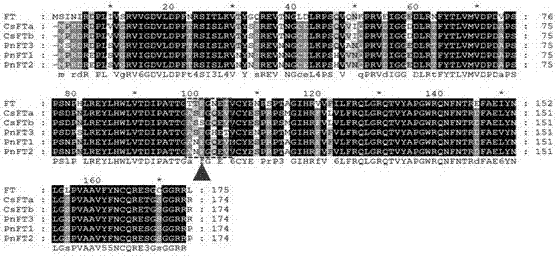
图2为茶树(csfta,csftb)、拟南芥(ft)及杨树(pnft1,pnft2)中的ft蛋白序列比对图,图2中方框显示茶树csfta/csftb蛋白的第100位氨基酸所在区域,三角形指示第100位氨基酸。
图3为以来自茶树成熟叶片的总cdna为模板测序csft基因的结果图,图3中三角形代表ft基因开放阅读框中差异碱基的位置。
图4为茶树csft基因的组织特异性表达图。
图5为利用基因特异性引物检测csfta和csftb在茶树休眠转换过程中在腋芽中的表达图。
图6为利用基因特异性引物检测csfta和csftb在休眠诱导下在叶片和嫩芽中的表达变化图。
图7为茶树csfta基因过表达杨树克隆717和克隆6表现早花现象图,图7中a到d是克隆717,e到f是克隆6,d和h是3个月大的野生型对照,从a到c,从e到g经转化的克隆717和克隆6在被移入生根培养基2-3周后即表现出强烈的早花现象,箭头指示花状结构。
图8为5个月大的csftb过表达杨树植株在温暖的长日照条件下的表型。
图9为csftb过表达杨树植株在短日照条件下的表型鉴定,图9中a到d是克隆717,e到h是克隆6。经过3个星期的短日照处理,野生型对照(a和e)停止生长并形成不活动的顶端,但是csftb过表达的植株顶端仍在继续生长(b和f),经过9个星期的短日照处理后,野生型对照已经已经形成了完整的休眠芽(c和g),但是过表达植株的生长点仍在继续生长(d和h)。
具体实施方式
以下结合实施例来进一步说明本发明。
实施例1:茶树csft基因的克隆及序列分析
以龙井43作为供试材料,获得茶树csft基因的不同转录本以转录组测序获得的csft基因片段为基础,利用race技术获得了1050bp的csft全长cdna序列信息,csft全长cdna核苷酸序列如seqidno:5所示。对其开放阅读框(orf)预测(http://www.ncbi.nlm.nih.gov/projects/gorf/)显示该基因可能编码一个含有174个氨基酸的小分子量蛋白。随后克隆了csft的全长orf,orf核苷酸序列如seqidno:6所示,并挑取多个克隆进行测序验证,结果获得了两种csft基因的转录本,两个转录本之间仅有1个碱基的差异。但是这1个碱基的差异导致ft基因编码的第100位氨基酸发生变化。一个为苯丙氨酸(phe),另一个为丝氨酸(ser)。分别命名这两个转录本为csfta和csftb(图1)。csfta的核苷酸序列如seqidno:1所示,csfta编码的蛋白质氨基酸序列如seqidno:3所示,csftb的核苷酸序列如seqidno:2所示,csftb编码的蛋白质氨基酸序列如csftaseqidno:4所示。
然后对茶树csft基因编码氨基酸序列与其他植物中ft蛋白序列进行了比对,结果发现克隆到的茶树csft基因编码蛋白序列与拟南芥和杨树中的ft蛋白序列有很高的相似性(图2)。因此克隆所得的ft基因应为拟南芥中ft基因的同源基因。同时结果还表明,csft第100位氨基酸所在位置的氨基酸序列保守性并不高,但是在拟南芥和杨树中第100位氨基酸均为苯丙氨酸。因此为了进一步确认csfta和csftb序列的准确性,利用成熟叶片中获得的mrna反转录成的cdna作为模板,对csft基因进行了重测序,结果显示在差异碱基处有两个明显的信号峰(图3)。这说明茶树中存在csfta和csftb两种转录本。
实施例2:茶树csft基因的表达分析
首先以csptb1为内参基因,设计通用引物检测csft基因的组织特异性表达和在不同生长状态的叶片和腋芽中的表达。结果显示,csft基因主要在叶片、花和芽内高表达,特别是在芽内,其表达量远高于在叶片中的表达量。但是在嫩芽中的表达非常低,而茎和根内则无表达(图4)。这说明csft基因与花的形成以及芽的发育密切相关。
为了进一步了解茶树中两个转录本csfta和csftb的表达差异,我们设计了特异性引物(如表1),以区分二者在不同休眠阶段和休眠诱导下的表达模式变化。结果显示,在处于类休眠(paradormancy)、生理休眠(endodormancy)、生态休眠(ecodormancy)和萌芽阶段(sprouting)的腋芽中,csfta和csftb在生态休眠阶段都有较高的表达水平,但是在类休眠和生态休眠阶段,二者之间的表达水平则没有明显差异(图5)。而在生理休眠和萌芽阶段,csfta和csftb的表达水平存在明显的差异。特别是在生理休眠阶段,二者之间的表达差异更加明显。在生理休眠和萌芽阶段csftb的表达水平都显著高于csfta的表达。
表1引物序列信息
在短日照和低温的休眠诱导条件下,csfta和csftb的表达在叶片和顶端嫩芽中的表达明显受到抑制(图6)。在叶片和顶端嫩芽中,csfta和csftb的表达在休眠诱导后的第1天都有明显的升高,随后二者的表达被持续显著抑制。在休眠诱导后的第4周,二者的表达水平最低。在叶片中,与csftb相比,csfta的表达受休眠诱导抑制更明显。csfta的表达在休眠诱导的第3天已达最低,而csftb的表达则逐渐下降,在休眠诱导后的第4周其表达水平仍高于csfta。在顶端嫩芽中,csfta和csftb有着相同的表达变化。
实施例3:功能验证及表型观察
目前茶树的转化体系还不成熟,杨树作为木本植物中的模式植物,有着非常全面的遗传信息,其休眠机理的研究也最为透彻。同时杨树和茶树均为木本植物,有着非常接近的遗传背景。因此利用杨树转化体系来验证茶树中基因的功能是最可靠的。本研究所选取的杨树克隆717和克隆6为目前实验室通用的转化材料,有着表型容易分辨、遗传背景清晰、转化率高的优点。另外,研究发现,杨树的休眠(生长停滞和休眠芽形成)仅受光周期影响,与温度无关(resmanetal.2010)。因此可以很好的控制休眠诱导的条件,从而了解茶树基因在杨树中发挥功能时受何种环境因素影响。
csfta和csftb的过表达载体被分别转入杨树克隆717和克隆6,并分别以转入空载体的对应植株为野生型对照。经转化体系成功筛选到的阳性植株又经过潮霉素抗性基因筛选以及表达筛选。经过验证已经成功被转化且高效表达目的基因的多个独立转化植株用于接下来的表型观察和试验处理。
在杨树中过表达csfta和csftb。结果显示csfta过表达的杨树植株表现出强烈的促进早花现象。一般情况下杨树需要十几年的时间才能开花,而csfta过表达的植株在被移入生根诱导培养基后2-3周就开始形成明显的花状结构。在克隆717,一些外植体形成的芽直接在顶端形成放射状的花絮结构(图7a),还有一些则在顶端形成花蕾或者蝶状花托结构(图7b和c)。在克隆6,同样出现花絮状结构(图7e,f,g)。同时csfta过表达的植株在被移入生根诱导培养基后,并不生根,而是直接开花。而且开花后的枝条或者植株很快就衰老死亡(图7e,f)。可见csft基因的过表达持续促进开花形成,已经影响了植株的正常发育。同时转化植株形成的花状结构并非完整的花器官,这也说明,花的正常形成除ft基因外还需要其他基因的共同参与。图7中的d和h分别为克隆717和克隆6的野生型对照。对照植株已经3个月大,生长正常,未表现出早花现象。
但是在同样条件下,csftb过表达的杨树植株并未表现出早花现象。转化植株与野生型对照有着相同的长势和表现。图8显示5个月大的csftb过表达杨树植株和野生型对照的表型。为了鉴定csftb过表达植株是否影响植物的休眠形成,5个月大的转化植株被移入短日照的休眠诱导培养箱以及长日照的低温培养箱进行处理,结果显示在短日照处理条件下,处理3周后野生型对照即停止生长,顶端生长点开始形成驻芽(图9)。而csftb过表达的植株则受短日照影响,表现出继续生长的状态。短日照诱导9周后,对照组植株已完全停止生长,顶端叶片已经开始衰老,而且已经形成饱满且有鳞片覆盖的休眠芽。而过表达植株则仍在继续生长,特别是克隆717并未表现出生长停滞或者休眠芽形成的现象。克隆6生长放缓,但是生长点仍处于活动状态。移入长日照低温培养条件的过表达植株与对照相比未表现出表型差异。但是低温直接影响杨树的生长,在试验植株被移入低温条件下,植株生长很快受到抑制。由于杨树休眠只受光周期影响,所以在低温条件下的植株没有休眠芽形成。
因此,过表达茶树csfta能显著促进开花,而茶树csftb则没有促进早开花的现象。但是将csftb过表达植株进行短日照处理后,与对照相比过表达植株能显著抑制短日照引起的生长停滞和休眠芽的形成。
sequencelisting
<110>中国农业科学院茶叶研究所
<120>茶树开花基因csft及其编码蛋白
<130>1
<160>6
<170>patentinversion3.3
<210>1
<211>525
<212>dna
<213>茶树
<400>1
atgcctcgggacagggatcctcttgttgttgggcgtgttataggggatgttttggacccc60
tttacaaggtctatctctctcagggtgacctacagcagtagggaagttaccaatggttgt120
gagctcaggccttctcaagttgtcatccaacctagggttgacattggtggtgatgacctt180
aggaccttctacactcttgttatggtggaccctgatgctcccagcccaagtgacccaaac240
ctaagggaatacttgcattggttggtgactgatatcccagcaaccactggagcaagcttt300
gggcaagaggtggtttgttatgagagtccgaggccatcaatgggaatccatcgattcgtt360
ttggtgttgtttagacagttgggaaggcaaacagtgtatgctccagggtggcgtcagaat420
ttcaacactcgggactttgcggagctttacaatctcggctctcctgtcgccgctgtttat480
tttaactgccaaagggagaccggctcaggaggccgaagacgatga525
<210>2
<211>525
<212>dna
<213>茶树
<400>2
atgcctcgggacagggatcctcttgttgttgggcgtgttataggggatgttttggacccc60
tttacaaggtctatctctctcagggtgacctacagcagtagggaagttaccaatggttgt120
gagctcaggccttctcaagttgtcatccaacctagggttgacattggtggtgatgacctt180
aggaccttctacactcttgttatggtggaccctgatgctcccagcccaagtgacccaaac240
ctaagggaatacttgcattggttggtgactgatatcccagcaaccactggagcaagctct300
gggcaagaggtggtttgttatgagagtccgaggccatcaatgggaatccatcgattcgtt360
ttggtgttgtttagacagttgggaaggcaaacagtgtatgctccagggtggcgtcagaat420
ttcaacactcgggactttgcggagctttacaatctcggctctcctgtcgccgctgtttat480
tttaactgccaaagggagaccggctcaggaggccgaagacgatga525
<210>3
<211>174
<212>prt
<213>茶树
<400>3
metproargaspargaspproleuvalvalglyargvalileglyasp
151015
valleuaspprophethrargserileserleuargvalthrtyrser
202530
serarggluvalthrasnglycysgluleuargproserglnvalval
354045
ileglnproargvalaspileglyglyaspaspleuargthrphetyr
505560
thrleuvalmetvalaspproaspalaproserproseraspproasn
65707580
leuargglutyrleuhistrpleuvalthraspileproalathrthr
859095
glyalaserpheglyglngluvalvalcystyrgluserproargpro
100105110
sermetglyilehisargphevalleuvalleupheargglnleugly
115120125
argglnthrvaltyralaproglytrpargglnasnpheasnthrarg
130135140
aspphealagluleutyrasnleuglyserprovalalaalavaltyr
145150155160
pheasncysglnarggluthrglyserglyglyargargarg
165170
<210>4
<211>174
<212>prt
<213>茶树
<400>4
metproargaspargaspproleuvalvalglyargvalileglyasp
151015
valleuaspprophethrargserileserleuargvalthrtyrser
202530
serarggluvalthrasnglycysgluleuargproserglnvalval
354045
ileglnproargvalaspileglyglyaspaspleuargthrphetyr
505560
thrleuvalmetvalaspproaspalaproserproseraspproasn
65707580
leuargglutyrleuhistrpleuvalthraspileproalathrthr
859095
glyalaserserglyglngluvalvalcystyrgluserproargpro
100105110
sermetglyilehisargphevalleuvalleupheargglnleugly
115120125
argglnthrvaltyralaproglytrpargglnasnpheasnthrarg
130135140
aspphealagluleutyrasnleuglyserprovalalaalavaltyr
145150155160
pheasncysglnarggluthrglyserglyglyargargarg
165170
<210>5
<211>1050
<212>dna
<213>茶树
<400>5
tacatggggattgcaataaacaaacaagtagtcagtcagcctcctccaatatatatattt60
tttccttttgatcattagtaggattagattgggttgtgttgcatattaatctctctccct120
ctctctcaatctctcagtttcctcccacttgtgatgcctcgggacagggatcctcttgtt180
gttgggcgtgttataggggatgttttggacccctttacaaggtctatctctctcagggtg240
acctacagcagtagggaagttaccaatggttgtgagctcaggccttctcaagttgtcatc300
caacctagggttgacattggtggtgatgaccttaggaccttctacactcttgttatggtg360
gaccctgatgctcccagcccaagtgacccaaacctaagggaatacttgcattggttggtg420
actgatatcccagcaaccactggagcaagctttgggcaagaggtggtttgttatgagagt480
ccgaggccatcaatgggaatccatcgattcgttttggtgttgtttagacagttgggaagg540
caaacagtgtatgctccagggtggcgtcagaatttcaacactcgggactttgcggagctt600
tacaatctcggctctcctgtcgccgctgtttattttaactgccaaagggagaccggctca660
ggaggccgaagacgatgaatcacatcatcatcatcatatattatactatactatcaccgt720
ttagtttaatctatctatatatgtctctctttactgtttgtgtctaaatctatctttaca780
cattgaagaggttgcctgaaagttgaaaccgctcactaggtagatcatatgtacttggac840
ttgtatcaaattaaaagatgcatgttgcaatgctattaattatatagatagatagataga900
tagggagagagagaaaggaggagggggctagctatcaagcgtgtggaaccaaatatgttg960
gaaatgtatacgcacgtctaacaaatacatatattataataaaatgaatgataatttagy1020
attgtaaaaaaaaaaaaaaaaaaaaaaaaa1050
<210>6
<211>525
<212>dna
<213>茶树
<400>6
atgcctcgggacagggatcctcttgttgttgggcgtgttataggggatgttttggacccc60
tttacaaggtctatctctctcagggtgacctacagcagtagggaagttaccaatggttgt120
gagctcaggccttctcaagttgtcatccaacctagggttgacattggtggtgatgacctt180
aggaccttctacactcttgttatggtggaccctgatgctcccagcccaagtgacccaaac240
ctaagggaatacttgcattggttggtgactgatatcccagcaaccactggagcaagcttt300
gggcaagaggtggtttgttatgagagtccgaggccatcaatgggaatccatcgattcgtt360
ttggtgttgtttagacagttgggaaggcaaacagtgtatgctccagggtggcgtcagaat420
ttcaacactcgggactttgcggagctttacaatctcggctctcctgtcgccgctgtttat480
tttaactgccaaagggagaccggctcaggaggccgaagacgatga525
- 还没有人留言评论。精彩留言会获得点赞!
- 蝎神经毒素AaIT及其编码基因与应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种花狭口蛙的基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种泽陆蛙基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 降低种子重量的白桦AP2基因及其编码蛋白的制造方法与工艺
- 可编码ArtRD蛋白基因及其蛋白和应用的制造方法与工艺
- 蝎神经毒素AaIT及其编码基因与应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种花狭口蛙的基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种泽陆蛙基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 蛋白质及其编码基因在调控植物抗黄萎病性中的应用的制造方法与工艺
- 甘薯耐逆相关蛋白IbFBA2、编码基因、重组载体与应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 核盘菌凝集素SSL‑6蛋白及其编码基因与应用的制造方法与工艺
- 植物抗病毒相关蛋白GmSN1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 蛋白质及其编码基因在调控植物抗黄萎病性中的应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 核盘菌凝集素SSL‑6蛋白及其编码基因与应用的制造方法与工艺