分离的T细胞受体、其修饰的细胞、编码核酸、表达载体、制备方法、药物组合物和应用与流程
本发明属于生物技术领域,具体而言,涉及分离的t细胞受体、其修饰的细胞、编码核酸、表达载体、制备方法、药物组合物和应用。
背景技术:
her2/neu(erbb2)是属于egfr家族的一个跨膜蛋白,和家族其它蛋白形成二聚体并通过一系列细胞内信号传导途径来调控细胞增值,分化以及癌变等过程(参见文献“growthfactors,2008;26:263”、“oncolbiol.phys,2004;58:903”)。her2/neu蛋白在多种上皮来源的癌细胞中过度表达,如乳腺癌,胃癌,大肠癌,卵巢癌,胰腺癌,肺癌,食管癌,膀胱癌,肾癌等(参见文献“trendsinmolecularmed,2013;19:677”),并且在原发灶和转移灶癌细胞中的表达相对均一(参见文献“jclinoncol,1998;8:103”),因此,her2/neu成为靶向治疗的适当靶点。
靶向her2/neu的人源化单抗药物herceptin可以显著延长her2/neu阳性的乳腺癌患者生存期(参见文献“nengljmed,2001,344:783”),然而单独使用herceptin治疗her2阳性的转移乳腺癌的临床反应率只有11%到26%(参见文献“jclinoncol,2002;20:7169”),表明单用heceptin对大部分her2高表达的转移乳腺癌的疗效并不理想。尽管heceptin联合化疗可提高临床应答率,但大多数her2/neu过度表达的乳腺癌病人在一年后会对heceptin产生抗性(参见文献“jclinoncol,2001;19:2587”)。
her2/neu阳性肿瘤患者体内会产生针对her2/neu抗原的内源性抗体和t细胞反应(参见文献“cancerres,2005;65:650”),因而,靶向her2/neu抗原的特异性免疫治疗成为一种颇有前景的治疗手段。特异性识别her2/neu抗原表位多肽(epitopepeptide)369-377的t细胞可以从her2/neu高表达的卵巢癌腹水中成功分离(参见文献“j.exp.med.1995;181:2109–2117”)。靶向her2/neu369-377多肽抗原的肿瘤疫苗进入临床试验,尽管临床1/2期显示此疫苗可以诱导出针对her2/neu369-377多肽抗原的特异性t杀伤细胞(参见文献“breastcare,2016;11:116”),但临床三期却没达到延长病人生存期的预定目标(http://www.onclive.com/web-exclusives/phase-iii-nelipepimuts-study-in-breast-cancer-halted-after-futility-review)。过继传输经体外培养的,基于嵌合抗原受体(chimericantigenreceptors,cars)的肿瘤特异性t细胞疗法被开发后,作为第一个针对实体瘤的,靶向her2/neu抗原的car-t细胞疗法进入临床试验,但由于产生强烈的细胞因子风暴(cytokinereleasesyndrome,crs)导致病人死亡而被终止(参见文献“naturemed,2016;22:26”)。严重的细胞因子风暴以及神经毒性是car-t治疗中常见的毒性反应(参见文献“blood,2016;127:3321”),部分原因可能和car这种非天然的t细胞受体不受限制的细胞活化有关(参见文献“natned,2015;21:581”),或者与不需抗原刺激的细胞因子的自分泌有关(参见文献“cancerimmunolres,2015;3:356”)。
过继转输经过特异性t细胞受体(即tcr)基因修饰的t细胞的tcr-t疗法被认为是针对实体瘤最具前景的免疫细胞基因疗法(参见文献“advimmunol.2016;130:279-94”)。其中,靶向ny-eso-1抗原的tcr-t疗法的临床研究显示出令人鼓舞的临床治疗效果(参见文献“natmed.2015aug;21(8):914-921”)。然而,目前已知的靶向肿瘤抗原、并有效识别肿瘤细胞的特异性tcr的数量十分有限,因此限制了tcr-t疗法的广泛应用。另外,尽管tcr-t疗法没有出现car-t疗法中所表现出的严重的细胞因子风暴毒性,但如果靶抗原来源于自身蛋白,针对正常组织细胞中低表达的靶抗原有可能会导致严重的自身免疫反应,即转接脱靶(或称中靶脱瘤)(ontargetofftumor)毒性反应(blood2009;114:535-546)。另外,为了获得有效识别肿瘤细胞的高亲和性tcr,一般的策略是体外通过对tcr上的互补性决定区(complementaritydetermingregions,cdrs)进行基因点突变,或者通过从未经过中枢耐受机制筛选的人源化小鼠t细胞库中进行诱导(参见文献“frontimmunol.2013;4:363”)。然而,这种高亲和性tcr可能会对正常自身蛋白产生交叉反应而导致对正常组织细胞的杀伤作用,即严重甚至致命的脱靶(off-target)毒性(参见文献“curropinimmunol2015;33:16–22”、参见文献“scitranslmed.2013;5(197):197ra103”)。因此,获得特异性靶向肿瘤抗原并有效识别肿瘤细胞,同时避免可能出现的脱靶毒副反应的新型tcr基因,是成功开发tcr-t免疫细胞基因疗法面临的主要挑战。
技术实现要素:
为解决上述现有技术中所存在的问题,本发明提供了分离的t细胞受体、其修饰的细胞、编码核酸、表达载体、制备方法、药物组合物和应用。
具体而言,本发明提供了:
(1)一种分离的t细胞受体,包括α链和β链中的至少一者,所述α链和β链均包含可变区和恒定区,其特征在于,所述t细胞受体能够特异性识别肿瘤细胞所表达的抗原her2/neu,并且所述α链的所述可变区的氨基酸序列具有与seqidno:19所示的氨基酸序列至少98%的一致性,所述β链的所述可变区的氨基酸序列具有与seqidno:20所示的氨基酸序列至少98%的一致性。
(2)根据(1)所述的t细胞受体,其中所述的t细胞受体能够特异性识别被hla-a2分子所提呈的所述抗原her2/neu的抗原表位多肽;优选的是,所述抗原表位多肽包括如seqidno:18所示的her2/neu369-377。
(3)根据(1)所述的t细胞受体,其中所述α链的所述恒定区和/或所述β链的所述恒定区来源于人;优选地,所述α链的所述恒定区全部或部分地被来源于其它物种的同源序列所替换,并且/或者所述β链的所述恒定区全部或部分地被来源于其它物种的同源序列所替换;更优选地,所述其它物种为小鼠。
(4)根据(1)所述的t细胞受体,其中所述α链的所述恒定区修饰有一个或多个二硫键,并且/或者所述β链的所述恒定区修饰有一个或多个二硫键。
(5)根据(1)所述的t细胞受体,其中所述α链的氨基酸序列如seqidnos:2、6或10所示,所述β链的氨基酸序列如seqidnos:4、8或12所示。
(6)一种分离的、编码t细胞受体的核酸,包含所述t细胞受体的α链和β链中的至少一者的编码序列,所述α链编码序列和β链编码序列均包含可变区编码序列和恒定区编码序列,其特征在于,所述t细胞受体能够特异性识别肿瘤细胞表达的抗原her2/neu,并且所述α链可变区编码序列编码的氨基酸序列具有与seqidno:19所示的氨基酸序列至少98%的一致性,所述β链可变区编码序列编码的氨基酸序列具有与seqidno:20所示的氨基酸序列至少98%的一致性。
(7)根据(6)所述的核酸,其中所述核酸为dna或rna。
(8)根据(6)所述的核酸,其中所述α链可变区编码序列如seqidno:21所示,所述β链可变区编码序列如seqidno:22所示。
(9)根据(6)所述的核酸,其中被所述核酸编码的所述t细胞受体能够特异性识别被hla-a2分子所提呈的所述抗原her2/neu的抗原表位多肽;优选的是,所述抗原表位多肽包括如seqidno:18所示的her2/neu369-377。
(10)根据(6)所述的核酸,其中所述α链恒定区编码序列和/或所述β链恒定区编码序列来源于人;优选地,所述α链恒定区编码序列全部或部分地被来源于其它物种的同源序列所替换,并且/或者所述β链恒定区编码序列全部或部分地被来源于其它物种的同源序列所替换;更优选地,所述其它物种为小鼠。
(11)根据(6)所述的核酸,其中所述α链恒定区编码序列包含一个或多个二硫键编码序列,并且/或者所述β链恒定区编码序列包含一个或多个二硫键编码序列。
(12)根据(6)所述的核酸,其中所述α链编码序列如seqidnos:1、5或9所示,所述β链编码序列如seqidnos:3、7或11所示。
(13)根据(6)-(11)中任一项所述的核酸,其中所述α链编码序列和所述β链编码序列之间由可切割性连接多肽的编码序列连接。
(14)根据(13)所述的核酸,其序列如seqidnos:13、15、或23所示。
(15)一种重组表达载体,其含有与启动子有效连接的、根据(6)-(14)中任一项所述的核酸,和/或其互补序列。
(16)一种t细胞受体修饰的细胞,该细胞的表面被(1)-(5)中任一项所述的t细胞受体修饰,其中所述细胞包括原始t细胞或其前体细胞,nkt细胞,或t细胞株。
(17)一种制备根据(16)所述的t细胞受体修饰的细胞的方法,包括以下步骤:
1)提供细胞;
2)提供编码根据(1)-(5)中任一项所述的t细胞受体的核酸;
3)将所述核酸转染入所述细胞中。
(18)根据(17)所述的方法,其中步骤1)所述的细胞来自自体或异体。
(19)根据(17)所述的方法,其中所述转染的方式包括:采用病毒载体转染的方式,优选的是,所述病毒载体包括γ逆转录病毒载体或慢病毒载体;化学方式,优选的是,所述化学方式包括采用脂质体转染的方式;物理方式,优选的是,所述物理方式包括电转染方式。
(20)根据(17)所述的方法,其中步骤2)所述的核酸为根据(6)-(14)中任一项所述的核酸。
(21)根据(16)所述的t细胞受体修饰的细胞在制备用于治疗或预防肿瘤和/或癌症的药物中的用途。
(22)根据(21)所述的用途,其中所述肿瘤和/或癌症是抗原her2/neu阳性的,并且是hla-a2阳性的。
(23)根据(16)所述的t细胞受体修饰的细胞在制备用于检测宿主的肿瘤和/或癌症的药物中的用途。
(24)一种药物组合物,其中该药物组合物包括作为活性成分的根据(16)所述的t细胞受体修饰的细胞,及可药用辅料。
(25)根据(24)所述的药物组合物,其中所述药物组合物包含每个患者每个疗程总剂量范围为1×103-1×109个细胞/kg体重的所述t细胞受体修饰的细胞。
(26)根据(24)所述的药物组合物,其中所述药物组合物适于经动脉、静脉、皮下、皮内、瘤内、淋巴管内、淋巴结内、蛛网膜下腔内、骨髓内、肌肉内和腹膜内给药。
(27)一种治疗肿瘤和/或癌症的方法,包括对肿瘤和/或癌症患者施用根据(16)所述的t细胞受体修饰的细胞。
(28)根据(27)所述的方法,其中所述t细胞受体修饰的细胞的施用剂量为每个患者每个疗程总剂量范围为1×103-1×109个细胞/kg体重。
(29)根据(27)所述的方法,其中所述t细胞受体修饰的细胞通过动脉、静脉、皮下、皮内、瘤内、淋巴管内、淋巴结内、蛛网膜下腔内、骨髓内、肌肉内和腹膜内给药。
(30)根据(27)所述的方法,其中所述肿瘤和/或癌症是抗原her2/neu阳性的,并且是hla-a2阳性的。
本发明与现有技术相比具有以下优点和积极效果:
本发明从hla-a2阳性的健康供体外周血中成功诱导出对hla-a2提呈的her2/neu抗原表位多肽(如her2/neu369-377多肽)有特异性的t细胞克隆,并从中筛选出携带有特异性识别her2/neu抗原表位多肽(如her2/neu369-377多肽)的天然tcr的t细胞克隆,进而获得了该tcr全序列。此tcr属于cd8分子依赖性,对her2/neu抗原表位多肽(如her2/neu369-377多肽)具有中等亲和性,可特异性识别hla-a2阳性并表达her2/neu抗原的肿瘤细胞。另外,携带此tcr的t细胞克隆经过中枢免疫耐受机制筛选后进入外周t细胞库。携带此tcr的杀伤t细胞曾存在于正常人外周血,并未对微量表达her2/neu蛋白的正常组织细胞产生交叉反应。因此,本发明获得了能够特异性识别肿瘤抗原,同时能够避免可能出现的脱靶毒副反应的新型tcr。
在进一步的发明中还对tcr的恒定区进行了改造(例如进行二硫键修饰或鼠源化改造),以使得此tcr在免疫细胞上表达时能够进一步减少或避免与内源tcr错配的发生。
用此tcr修饰的免疫细胞(例如原始t细胞、其前体细胞、nkt细胞、t细胞株)可特异性识别多种hla-a2+且her2/neu+的肿瘤细胞株,具有显著的抗肿瘤效果。另一方面,用于修饰免疫细胞的tcr不会对微量表达her2/neu的正常细胞产生交叉反应。因此,基于此tcr的tcr-t疗法可望治疗多种实体瘤。
用本发明的tcr修饰的免疫细胞治疗肿瘤时,可有效避免采用car-t治疗时所引起的细胞因子风暴和免疫排斥反应。
本发明的tcr修饰的免疫细胞为治疗hla-a2+且her2/neu+肿瘤患者提供了一种新的选择,具有良好的产业应用前景。
附图说明
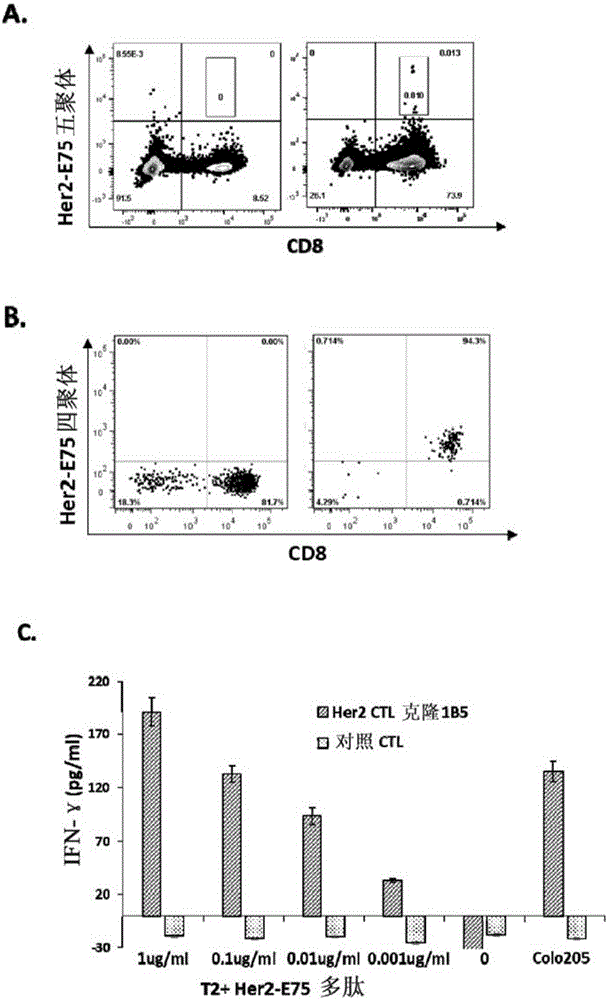
图1示出本发明实施例1中从hla-a2+正常供体pbmc(具体为#2pbmc)中诱导出的her2/neu369-377多肽(her2-e75)特异性杀伤性t细胞的表型和功能检测结果。图1a为经过两轮her2-e75抗原多肽体外刺激后,pbmc细胞经cd8-apc抗体和her2-e75五聚体-pe染色后进行流式细胞分析结果,右图是经多肽刺激的细胞,对cd8+五聚体+杀伤t细胞群进行facs分选,以获得t细胞克隆。左图为没有多肽刺激的对照组细胞。横坐标表示cd8分子表达的荧光强度,纵坐标表示结合的her2-e75五聚体的荧光强度。图1b为cd8+e75-tetramer+杀伤t细胞克隆经cd8-apc和her2-e75四聚体-pe染色后流式细胞的表型分析,右图显示cd8+her2四聚体+t细胞克隆her2ctl1b5为纯化的her2-e75多肽特异性ctl细胞克隆。左图为没有多肽刺激的对照组ctl细胞。横坐标表示cd8分子表达的荧光强度,纵坐标表示结合的her2-e75四聚体的荧光强度。图1c为t细胞克隆功能检测结果,t细胞克隆her2ctl1b5(斜线柱图)或者没有多肽刺激的对照ctl细胞(点柱图)与被t2细胞提呈的不同浓度her-e75多肽共培养,或者与hla-a2+her2/neu+的大肠癌细胞株colo205共培养后,取细胞上清进行ifn-γ的elisa分析,各试验组和对照组均为复孔,结果显示为平均值±sd。横坐标表示不同实验组别,纵坐标表示t细胞分泌ifn-γ的浓度。
图2示出所构建的两种携带her2tcr-1b5tcr基因的慢病毒载体(图中分别示为“pcdh-ef1α-her2tcr-(pgk-gfp)载体”、“pcdh-ef1α-her2tcr载体”)的主要功能片段。上方示出的片段同时表达ef-1α启动子所驱动的tcr基因和pkg启动子所驱动的gfp基因,下方示出的片段只表达tcr基因。各tcr的β链和α链均被可切割性连接多肽的编码序列(furin-f2a)所链接。
图3示出经her2tcr-1b5tcr基因转染的t细胞株的表型和功能检测结果。用编码her2tcr-1b5tcr和gfp的慢病毒载体转染j.rt3-t3.5t细胞株(j.rt3),经her2-e75四聚体-pe染色后进行流式细胞分析。图3a示出的gfp+her2-e75四聚体+细胞群为表达her2tcr-1b5tcr的细胞,所示百分率为各阳性细胞群占总细胞数的比率。左图涉及的tcr为α链和β链的恒定区增加了一个二硫键结构(her2tcr-1b5-dis),右图涉及的tcr为人α链和β链的恒定区被小鼠恒定区的同源序列所置换(her2tcr-1b5-mc)。横坐标表示gfp分子表达的荧光强度,纵坐标表示结合的her2-e75四聚体的荧光强度。图3b示出tcrα链和β链的恒定区经不同方式修饰后在两种t细胞株的表达情况。图中“her2tcr-1b5-dis”指α链和β链的恒定区各增加了一个二硫键结构的tcr;“her2tcr-1b5-mc”指人α链和β链的恒定区被小鼠恒定区的同源序列所置换的tcr。gfp+her2-e75四聚体+细胞为表达her2tcr-1b5tcr的阳性细胞,纵坐标为该tcr阳性细胞占总gfp+细胞的百分比。横坐标表示不同t细胞株组别,其中“jurkat(tcra+b+)”是指jurkat细胞,其α链和β链均表达,而“j.rt3(tcra+b-)”是指j.rt3-t3.5细胞,其来源于jurkat细胞,并且β链基因缺失,α链仍有表达。图3c示出经编码her2tcr-1b5tcr基因的慢病毒载体转染的t细胞株可以识别被t2细胞所提呈的her2-e75多肽。tcrα链和β链的恒定区经不同方式修饰,表达tcr的t细胞株与提呈不同浓度her2-e75多肽的t2细胞混合培养16小时,用抗cd69-pe抗体染色后进行流式细胞分析。图中“j.rt3-her2-1b5-dis”表示表达her2tcr-1b5-dis的j.rt3-t3.5细胞,“j.rt3-her2-1b5-mc”表示表达her2tcr-1b5-mc的j.rt3-t3.5细胞,“jurkat-her2-1b5-dis”表示表达her2tcr-1b5-dis的jurkat细胞,“jurkat-her2-1b5-mc”表示表达her2tcr-1b5-mc的jurkat细胞。横坐标表示t2细胞所提呈的her2-e75多肽的浓度。纵坐标为cd69+细胞占总gfp+细胞的百分比。
图4示出经her2tcr-1b5-mctcr基因转染的外周血单个核细胞(pbmc)的表型和功能检测结果。图4a为编码her2tcr-1b5-mc的慢病毒载体转染来自两个不同供体的pbmc,经her2-e75四聚体-pe和抗cd8-apc抗体染色后进行流式细胞分析的结果。首先根据细胞形态和大小分出淋巴细胞群,her2-e75四聚体+细胞群为表达her2tcr-1b5tcr的细胞。横坐标表示cd8分子表达的荧光强度,纵坐标表示结合的her2-e75四聚体的荧光强度。所示百分率为各阳性细胞群占分出的淋巴细胞数的比率。左图涉及一个供体所提供的外周血单个核细胞(#1pbmc),右图涉及另一个不同供体提供的pbmc(#2pbmc)。cd8+her2-e75四聚体+细胞为表达her2tcr-1b5-mc的杀伤性t细胞。cd8-her2-e75四聚体+细胞可能为表达her2tcr-1b5-mc的cd4+辅助t细胞。图4b示出表达her2tcr-1b5-mc的t细胞可以识别被t2细胞所提呈的her2-e75多肽。经编码her2tcr-1b5-mc和gfp的慢病毒载体转染的两个不同供体pbmc分别与提呈不同浓度梯度her2-e75多肽的t2细胞混合培养16小时,取细胞上清进行ifn-γ的elisa分析。对照组中靶细胞为提呈可以结合hla-a2分子的ebv病毒抗原多肽lmp2426-434的t2细胞。图中“t2+her2-e750.1μg/ml”表示提呈0.1μg/ml的her2-e75多肽的t2细胞组,“t2+her2-e750.01μg/ml”表示提呈0.01μg/ml的her2-e75多肽的t2细胞组,“t2+her2-e750.001μg/ml”表示提呈0.001μg/ml的her2-e75多肽的t2细胞组,“t2+ebv-lmp1μg/ml”表示提呈1μg/ml的ebv病毒抗原多肽lmp2426-434的t2细胞组。横坐标表示不同供体的pbmc组别,纵坐标表示t细胞分泌的ifn-γ的浓度。图4c示出t细胞功能的cd8抗体阻断试验结果。其中,经编码her2tcr-1b5-mc和gfp基因的慢病毒载体转染的#2pbmc与t2细胞提呈的抗原多肽共培养时加入抗人cd8抗体,检测t细胞分泌ifn-γ的功能是否被抑制。图中“t2+her2-e750.1μg/ml”表示未加入抗人cd8抗体的、提呈0.1μg/ml的her2-e75多肽的t2细胞组,“t2+her2-e750.1μg/ml+anti-cd8”表示加入抗人cd8抗体的、提呈0.1μg/ml的her2-e75多肽的t2细胞组。横坐标表示不同实验组别,纵坐标表示t细胞分泌的ifn-γ的浓度。图4b和4c中各试验组和对照组均为复孔,结果显示为平均值±sd。
图5示出经her2tcr-1b5-mctcr基因转染的外周血单个核细(pbmc)识别肿瘤细胞株的功能检测结果。图5a示出编码her2tcr-1b5-mctcr基因的慢病毒载体转染#2pbmc,与不同肿瘤细胞株细胞混合培养16小时后,取细胞上清进行ifn-γ的elisa分析结果。各试验组和对照组均为复孔,结果显示为平均值±sd。横坐标表示不同实验组别,纵坐标表示t细胞分泌的ifn-γ的浓度。图5b示出上述转染后的#2pbmc靶向colo205细胞的培养孔中分别加入抗cd8抗体(图中示为“colo205+anti-cd8”)或者抗hla-abc抗体后(图中示为“colo205+anti-hla-abc”),进行的功能阻断试验结果。横坐标表示不同肿瘤细胞株组别,纵坐标表示t细胞分泌的ifn-γ的浓度。colo205和coca-2为hla-a2阳性her2/neu阳性结肠癌细胞,mad-mb-231为hla-a2阳性her2/neu阳性乳腺癌细胞,h647为hla-a2阴性her2/neu阳性肺癌细胞,h1355为hla-a2阳性her2/neu阳性肺癌细胞,sk-ov-3为hla-a2阴性her2/neu阳性卵巢癌细胞,bjab为hla-a2阳性her2/neu阴性淋巴瘤细胞。
具体实施方式
以下通过具体实施方式的描述并参照附图对本发明作进一步说明,但这并非是对本发明的限制,本领域技术人员根据本发明的基本思想,可以做出各种修改或改进,但是只要不脱离本发明的基本思想,均在本发明的范围之内。
在本发明中,词语“肿瘤”、“癌症”、“肿瘤细胞”、“癌细胞”、“t细胞”、“t细胞受体”、“t细胞受体修饰”、“tcr可变区”、“tcr恒定区”、“抗原”、“抗原表位多肽”、“同源序列”、“编码”、“抗原提呈”、“重组dna表达载体”、“启动子”、“互补序列”、“转染”、“自体”、“异体”、“特异性识别”、“tcr-t疗法”涵盖本领域通常认为的含义。
her2/neu抗原属于肿瘤相关抗原,识别her2/neu抗原的高亲和性t细胞多数被中枢耐受机制所清除,以避免导致可能的自身免疫反应(参见文献“immunolrev.2016;271(1):127-40”)。因此,从外周血t细胞库中诱导出具有特异性识别肿瘤细胞所表达的her2/neu抗原的t细胞克隆变得十分困难。利用树突状细胞(dendriticcell)提呈her2/neu369-377多肽抗原,进而从her2/neu369-377多肽疫苗免疫过的患者外周血中诱导出的高亲和性tcr尽管可以识别极低量外源所负载的(exogenouslyloaded)her2/neu369-377多肽,但不能识别肿瘤细胞内源提呈的(endogenouslypresented)抗原多肽(参见文献“cancerres.1998;58:4902–4908”)。这可能是由于外源负载的多肽/hla复合物在构型(conformation)上与细胞内部自然提呈的hla/多肽复合物有所不同而导致,或者由于her2/neu369-377多肽位于her2蛋白高度糖化区,细胞内部自然提成的her2/neu369-377多肽可能被糖基化而导致tcr识别构型的差异(参见文献“proc.natl.acad.sci.usa2003;100:15029–15034”)。体外通过her2/neu369-377抗原多肽诱导t细胞的过程中,仅能识别外源负载抗原多肽的高亲和性t细胞克隆往往获得优势生长(dominantexpansion),而能特异性识别被细胞提成的内源性her2/neu抗原多肽的t细胞克隆生长受到抑制(参见文献“jexpmed.2016nov14;213(12):2811-2829”),因而增加了获得可识别肿瘤细胞的功能性tcr的难度。经过中枢耐受机制筛选后,这些识别肿瘤细胞的tcr多属于中等亲和性,其功能往往依赖于cd8分子的辅助作用。有研究小组从hla-a2阴性的外周血中诱导出异体t细胞(allo–tcells),可以特异性识别hla-a2限制的her2/neu369-377抗原多肽,用获得的tcr基因转染t细胞后,不仅可以识别肿瘤细胞提成的her2/neu369-377抗原多肽,也可交叉识别同家族的her3以及her4抗原表位(参见文献“journalofimmunology,2008,180:8135–8145”)。然而,基于异体allo-tcr的tcr-t疗法存在产生针对其它正常自身蛋白抗原表位的异体反应(allo-reaction)的风险(参见文献“int.j.cancer2009;125,649–655。natimmunol2007;8:388–97”)。另外一个研究小组从her2/neu369-377多肽疫苗免疫过的肿瘤患者外周血中诱导出her2/neu369-377多肽特异性t细胞,并把来源于不同t细胞的alpha和beta链进行配对并筛选出一个高亲和性tcr,转染此高亲和性tcr的t细胞可识别hla-a2+her2/neu+的多种肿瘤细胞(参见文献“humangenetherapy2014;25:730–739”)。这个tcr不是从单克隆t细胞获得,因此不能确定此tcr是否是存在于经过中枢耐受筛选过的t细胞库的天然tcr。her2/neu蛋白也在心肌、肺、食道、肾、膀胱这些重要脏器有微量表达(参见文献“oncogene.1990jul;5(7):953-62”),因此基于高亲和性her2/neu抗原特异性tcr的tcr-t疗法有对正常组织产生脱靶毒性反应的风险。
肿瘤细胞高表达her2/neu蛋白,因此细胞表面被hla提呈的抗原多肽的数量也会相应增加,在肿瘤细胞和正常细胞上hla/抗原多肽复合物数量的差异可成为特异性t细胞区分正常和肿瘤组织的窗口。本发明提出从自体t细胞库(auto-tcellrepertoire)中获得天然tcr的序列,进而在体外使tcr表达在t细胞上,以使所得到的表达tcr的t细胞可识别肿瘤细胞增加表达的her2/neu抗原,是成功开发有效低毒的tcr-t疗法的关键。
为了获得能够特异性识别肿瘤抗原,同时能够避免可能出现的脱靶毒副反应的tcr,本发明从hla-a2阳性的健康供体外周血中诱导对hla-a2提呈的her2/neu369-377多肽有特异性的t细胞克隆,并从中筛选出携带有对her2/neu369-377多肽具有中等亲和性的天然tcr的t细胞克隆。这不同于其他研究小组从经过her2/neu369-377多肽疫苗免疫过的肿瘤患者外周血诱导her2/neu369-377多肽特异性t细胞的策略(参见文献“humangenetherapy2014,25:730–739”),本发明认为经过her2/neu369-377抗原多肽免疫后,针对her2/neu369-377多肽的特定t细胞克隆会优势增殖,因而不能代表体内t细胞库(repertoire)中自然存在的可识别靶细胞所提呈的her2/neu369-377多肽抗原的特异性t细胞群。本发明也没有采取其他研究小组从hla-a2阴性外周血中诱导多肽特异性t细胞的方式(参见文献“thejournalofimmunology,2010,184:1617–1629”),尽管从异体pbmc中更容易获得高亲和性的识别her2/neu369-377多肽抗原的allo-t细胞,但这也同时增加了t细胞交叉识别被hla-a2分子提呈的其它多肽而导致的异体反应。
基于上述构思,本发明提供了一种分离的t细胞受体,包括α链和β链中的至少一者,所述α链和β链均包含可变区和恒定区,其特征在于,所述t细胞受体能够特异性识别肿瘤细胞所表达的抗原her2/neu,并且所述α链的所述可变区的氨基酸序列具有与seqidno:19所示的氨基酸序列至少98%、优选至少98.5%、更优选至少99%的一致性,所述β链的所述可变区的氨基酸序列具有与seqidno:20所示的氨基酸序列至少98%、优选至少98.5%、更优选至少99%的一致性,只要不显著影响本发明的效果即可。还优选的是,所述α链的所述可变区的氨基酸序列如seqidno:19所示,所述β链的所述可变区的氨基酸序列如seqidno:20所示。
tcrα链和β链的可变区用于结合抗原多肽/主要组织相容性复合体(mhci),分别包括三个超变区或称为互补决定区(complementaritydeterminingregions,cdrs),即,cdr1、cdr2、cdr3。其中cdr3区域对特异性识别被mhc分子提呈的抗原多肽至关重要。tcrα链是不同的v和j基因片段重组而成,β链则是不同的v、d和j基因片段重组而成。特定基因片段重组结合所形成的相应cdr3区域,以及结合区域回文以及随机插入的核苷酸(palindromicandrandomnucleotideadditions)形成了tcr对抗原多肽识别的特异性(参见文献“immunobiology:theimmunesysteminhealthanddisese.5theditin,chapter4,thegenerationoflymphocyteantigenreceptors”)。所述mhci类分子包括人hla。所述hla包括:hla-a、b、c。
进一步具体地,所述的t细胞受体能够特异性识别被hla-a2分子所提呈的所述抗原her2/neu的抗原表位多肽。抗原her2/neu的氨基酸序列如seqidno:17所示。优选的是,所述抗原表位多肽包括如seqidno:18所示的her2/neu369-377。hla-a2阳性细胞表达的hla-a2等位基因包括hla-a*0201、0202、0203、0204、0205、0206和0207。优选的是,所述hla-a2分子优选为hla-a*0201。
在一个实施方案中,所述抗原her2/neu的抗原表位多肽为her2/neu369-377多肽(seqidno:18)。在其它实施方案中,所述抗原her2/neu的抗原表位多肽为与her2/neu369-377多肽具有4-9个连续的相同氨基酸(例如,4、5、6、7、8或9个连续的相同氨基酸)的抗原表位多肽,并且这些多肽的长度为8-11个氨基酸。例如,在一个实施方案中,所述抗原her2/neu的抗原表位多肽为her2/neu373-382多肽(seqidno:25)。
优选地,所述t细胞受体识别her2/neu369-377多肽的最大半反应多肽浓度在1.0-10ng/ml之间(例如,在3.0-8.0ng/ml、5.0-7.0ng/ml之间)。在本发明的一个实施方案中,所述最大半反应多肽浓度约为6.9ng/ml。术语“最大半反应多肽浓度”是指诱导t细胞反应达到最大值的50%所需多肽的浓度。据报道,针对巨细胞病毒(cmv)抗原cmvpp65(495-503)多肽的特异性t细胞的最大半反应多肽浓度在0.1-1ng/ml之间,而此tcr对cmv抗原多肽被认为具有高亲和性(参见文献“journalofimmunogicalmethds2007;320:119-131”)。在本发明中,所述t细胞受体对her2/neu抗原具有中等亲和性,从而可避免高亲和性可能带来的脱靶毒性。
t细胞表达的外源tcrα链和β链有可能和t细胞本身tcr的α链和β链发生错配,不仅会稀释正确配对的外源tcr的表达量,错配tcr的抗原特异性也不明确,因而有识别自身抗原的潜在危险,因此优选将tcrα链和β链的恒定区修饰以减少或避免错配。
在一个实施方案中,所述α链的所述恒定区和/或所述β链的所述恒定区来源于人;优选地,本发明发现所述α链的所述恒定区可以全部或部分地被来源于其它物种的同源序列所替换,并且/或者所述β链的所述恒定区可以全部或部分地被来源于其它物种的同源序列所替换。更优选地,所述其它物种为小鼠。所述替换可以增加细胞中tcr的表达量,并且可以进一步提高被该tcr修饰的细胞对her2/neu抗原的特异性。
所述α链的所述恒定区可以修饰有一个或多个二硫键,并且/或者所述β链的所述恒定区可以修饰有一个或多个二硫键,例如1个或2个。
在具体的实施方式中,制备了两种不同方式修饰的tcr,her2tcr-1b5-dis是通过点突变在tcr恒定区增加一个二硫键,方法在文献“cancerres.2007apr15;67(8):3898-903.”中描述,其全文通过引用方式并入本文。her2tcr-1b5-mc是用小鼠tcr恒定区序列置换相应的人tcr恒定区序列,方法在文献“eur.j.immunol.200636:3052–3059”中描述,其全文通过引用方式并入本文。
在具体的实施方案中,所述α链的氨基酸序列如seqidnos:2、6或10所示,所述β链的氨基酸序列如seqidnos:4、8或12所示。
其中,对于氨基酸序列如seqidno:2所示的α链,其序列为原始的人源序列;对于氨基酸序列如seqidno:6所示的α链,其在恒定区修饰有1个二硫键;对于氨基酸序列如seqidno:10所示的α链,其恒定区替换为鼠源恒定区。
其中,对于氨基酸序列如seqidno:4所示的β链,其序列为原始的人源序列;对于氨基酸序列如seqidno:8所示的β链,其在恒定区修饰有1个二硫键;对于氨基酸序列如seqidno:12所示的β链,其恒定区替换为鼠源恒定区。
在一个具体实施方案中,所述tcr的α链的氨基酸序列如seqidno:2所示,β链的氨基酸序列如seqidno:4所示。在另一个具体实施方案中,所述tcr的α链的氨基酸序列如seqidno:6所示,β链的氨基酸序列如seqidno:8所示。在又一个具体实施方案中,所述tcr的α链的氨基酸序列如seqidno:10所示,β链的氨基酸序列如seqidno:12所示。
在本发明其它具体的实施方案中,所述tcr的α链具有在seqidnos:2、6或10所示氨基酸序列中替换、删除、和/或添加一个或多个氨基酸而得到的氨基酸序列;例如,所述α链具有与seqidnos:2、6或10所示氨基酸序列至少90%、优选至少95%、更优选至少99%的一致性。
在本发明其它具体的实施方案中,所述tcr的β链具有在seqidnos:4、8或12所示氨基酸序列中替换、删除、和/或添加一个或多个氨基酸而得到的氨基酸序列;例如,所述β链具有与seqidnos:4、8或12所示氨基酸序列至少90%、优选至少95%、更优选至少99%的一致性。
本发明的tcr的α链和/或β链还可以在末端(例如c末端)结合其它功能性序列,例如共刺激信号cd28、4-1bb和/或cd3zeta的功能区序列。
本发明还提供了一种分离的、编码t细胞受体的核酸,包含所述t细胞受体的α链和β链中的至少一者的编码序列,所述α链编码序列和β链编码序列均包含可变区编码序列和恒定区编码序列,其特征在于,所述t细胞受体能够特异性识别肿瘤细胞表达的抗原her2/neu,并且所述α链可变区编码序列编码的氨基酸序列具有与seqidno:19所示的氨基酸序列至少98%、优选至少98.5%、更优选至少99%的一致性,所述β链可变区编码序列编码的氨基酸序列具有与seqidno:20所示的氨基酸序列至少98%、优选至少98.5%、更优选至少99%的一致性,只要不显著影响本发明的效果即可。还优选的是,所述α链可变区编码序列编码如seqidno:19所示的氨基酸序列,所述β链可变区编码序列编码如seqidno:20所示的氨基酸序列。
所述核酸可以为dna或rna。
优选地,所述α链可变区编码序列如seqidno:21所示,所述β链可变区编码序列如seqidno:22所示。
进一步具体地,被所述核酸编码的所述t细胞受体能够特异性识别被hla-a2分子所提呈的所述抗原her2/neu的抗原表位多肽。
在一个实施方案中,所述抗原her2/neu的抗原表位多肽为her2/neu369-377多肽(seqidno:18)。在其它实施方案中,所述抗原her2/neu的抗原表位多肽为与her2/neu369-377多肽具有4-9个连续的相同氨基酸(例如,4、5、6、7、8或9个连续的相同氨基酸)的抗原表位多肽,并且这些多肽的长度为8-10个氨基酸。例如,在一个实施方案中,所述抗原her2/neu的抗原表位多肽为her2/neu373-382多肽(seqidno:25)。
优选地,被所述核酸编码的所述t细胞受体识别her2/neu369-377多肽的最大半反应多肽浓度在1.0-10ng/ml之间(例如,在3.0-8.0ng/ml、5.0-7.0ng/ml之间)。在本发明的一个实施方案中,所述最大半反应多肽浓度约为6.9ng/ml。在此情况下,所述t细胞受体对her2/neu抗原具有中等亲和性,可避免高亲和性可能带来的脱靶毒性。
在一个实施方案中,所述α链的所述恒定区和/或所述β链的所述恒定区来源于人;优选地,所述α链恒定区编码序列全部或部分地被来源于其它物种的同源序列所替换,并且/或者所述β链恒定区编码序列全部或部分地被来源于其它物种的同源序列所替换。更优选地,所述其它物种为小鼠。所述替换可以增加细胞中tcr的表达量,并且可以进一步提高被该tcr修饰的细胞对her2/neu抗原的特异性。
所述α链恒定区编码序列可以包含一个或多个二硫键的编码序列,并且/或者所述β链恒定区编码序列可以包含一个或多个二硫键的编码序列。
在具体的实施方案中,所述α链编码序列如seqidnos:1、5或9所示,所述β链编码序列如seqidnos:3、7或11所示。
其中,对于编码序列如seqidno:1所示的α链,其序列为原始的人源序列;对于编码序列如seqidno:5所示的α链,其在恒定区修饰有1个二硫键;对于编码序列如seqidno:9所示的α链,其恒定区替换为鼠源恒定区。
其中,对于编码序列如seqidno:3所示的β链,其序列为原始的人源序列;对于编码序列如seqidno:7所示的β链,其在恒定区修饰有1个二硫键;对于编码序列如seqidno:11所示的β链,其恒定区替换为鼠源恒定区。
在一个具体实施方案中,所述tcr的α链的编码序列如seqidno:1所示,β链的编码序列如seqidno:3所示。在另一个具体实施方案中,所述tcr的α链的编码序列如seqidno:5所示,β链的编码序列如seqidno:7所示。在又一个具体实施方案中,所述tcr的α链的编码序列如seqidno:9所示,β链的编码序列如seqidno:11所示。
在另外的实施方案中,所述α链编码序列和所述β链编码序列之间由可切割性连接多肽的编码序列连接,这样可以增加tcr在细胞内的表达。术语“可切割性连接多肽”是指该多肽起到连接作用,并且可以被特定的酶切割,或者编码此多肽的核酸序列通过核糖体跳跃方式(ribosomeskipping)进行翻译,从而使被其连接的多肽彼此分离。可切割性连接多肽的例子是本领域已知的,例如f2a多肽,f2a多肽序列包括但不限于来自微小核糖核酸病毒的f2a多肽、以及来自其它病毒相似的2a类序列。另外,可切割性连接多肽也包括可被furin酶切割的标准的四氨基酸基序(canonicalfouraminoacidmotif),即r-x-[kr]-r氨基酸序列。该实施方案所编码的tcr为单链嵌合t细胞受体,该单链嵌合t细胞受体表达完成后,连接α链和β链的可切割性连接多肽会被细胞中的特定酶切割,从而形成等量游离的α链和β链。
组成单链嵌合tcr的α链和β链也可如上文所述,恒定区(及其相应的编码序列)全部或部分地被来源于其它物种的同源序列所替换,并且/或者修饰有(编码)一个或多个二硫键。
在具体的实施方案中,所述核酸的序列如seqidnos:13、15、或23所示。
优选地,对所述核酸的核苷酸序列进行编码子优化以增加基因表达、蛋白翻译效率以及蛋白表达,从而增强tcr识别抗原的能力。编码子优化包括但不限于翻译启动区域的修饰、改变mrna结构片段、以及使用编码同一氨基酸的不同密码子。
在其它的实施方案中,可以对上述tcr编码核酸的序列进行突变,包括去除、插入和/或置换一个或多个氨基酸密码子,使得所表达的tcr识别her2/neu抗原的功能不变或者增强。例如,在一个实施方案中,进行保守氨基酸置换,包括对上述tcrα链和/或β链的可变区中的一个氨基酸用结构和/或化学属性相似的另一个氨基酸进行置换。术语“相似的氨基酸”是指具有相似的极性、电负荷、可溶性、疏水性、亲水性等属性的氨基酸残基。突变后的tcr仍具有识别上述被靶细胞提呈的her2/neu抗原多肽的生物活性。在另一个实施方案中,进行tcr成熟性(tcrmaturation)修饰,即,包括对上述tcrα链和/或β链的可变区中的互补决定区2(cdr2)和/或cdr3区域的氨基酸进行去除、插入和/或置换,从而改变tcr结合her2/neu抗原的亲和性。
本发明还提供了一种分离的、由根据本发明所述的dna转录的mrna。
本发明还提供了一种重组表达载体,其含有与启动子有效连接的根据本发明所述的核酸(例如dna),和/或其互补序列。
优选地,在所述重组表达载体中,本发明所述的dna合适地与启动子、增强子、终止子和/或polya信号序列有效连接。
本发明的重组表达载体的上述作用元件的组合能够促进dna的转录和翻译,并增强mrna的稳定性。
重组表达载体的基本骨架可以是任何已知的表达载体,包括质粒或病毒,病毒载体包括但不限于(例如)逆转录病毒载体(病毒原型为莫洛尼鼠白血病病毒(mmlv))和慢病毒载体(病毒原型为人类免疫缺陷i型病毒(hiv))。表达本发明所述tcr的重组载体可以通过本领域常规的重组dna技术来获得。
在一个实施方案中,重组表达载体上的α链和β链基因的表达可以由两个不同的启动子所驱动,启动子包括各种已知的类型,例如强表达的、弱表达的、可诱导的、组织特异性的、和分化特异性的启动子。启动子可以是病毒来源的或者非病毒来源的,例如cmv启动子、mscv的ltr上的启动子、ef1-α启动子、和pgk-1启动子。两个启动子的驱动方向可以是同向也可以是反向的。
在另一个实施方案中,重组表达载体上的α链和β链基因的表达可以由同一个启动子所驱动,例如编码单链嵌合t细胞受体的情况,α链的核苷酸序列和β链的核苷酸序列由furin-f2a多肽编码序列相连接。
在另一些实施方案中,重组表达载体除了包含α链和β链基因外,还可以包含其它功能分子的编码序列。一个实施方案包括表达自发荧光蛋白(如gfp或其它荧光蛋白)以用于体内追踪成像。另一个实施方案包括表达可诱导的自杀基因系统,例如诱导表达单纯疱疹病毒-胸腺嘧啶核苷激酶(hsv-tk)蛋白,或者诱导表达caspase9(icasp9)蛋白。表达这些“安全转换分子”(safety-switch)可以增加经本发明所述tcr基因修饰的细胞在体内使用的安全性。另一个实施方案包括表达人cd8基因,包括单独或联合表达cd8α链和β链,经本发明所述tcr基因修饰的细胞表达cd8分子后可以增强其特异性识别her2/neu抗原的能力,或使cd8阴性t细胞(如cd4+t辅助细胞)获得特异性识别her2/neu抗原的能力。另一个实施方案包括表达人趋化因子受体基因,例如ccr2,这些趋化因子受体可结合肿瘤组织中高表达的相应趋化因子配体,从而可以增加经本发明所述tcr基因修饰的细胞在肿瘤组织中的归巢。
本发明还提供了一种t细胞受体修饰的细胞,该细胞的表面被本发明所述的t细胞受体修饰,其中所述细胞包括原始t细胞或其前体细胞,nkt细胞,或t细胞株。
所述“t细胞受体修饰”中的“修饰”是指,通过基因转染使细胞表达有本发明所述的t细胞受体,即,所述t细胞受体通过跨膜区锚定在所修饰的细胞的细胞膜上,并具有识别抗原多肽/mhc复合物的功能。
本发明还提供了一种制备根据本发明所述的t细胞受体修饰的细胞的方法,包括以下步骤:
1)提供细胞;
2)提供编码本发明t细胞受体的核酸;
3)将所述核酸转染入所述细胞中。
步骤1)所述的细胞可以来源于哺乳动物,包括人、犬、小鼠、大鼠及其转基因动物。所述细胞可以来自自体或异体。异体细胞包括来自同卵双胞胎的细胞、异体干细胞、经基因改造的异体t细胞。
步骤1)所述的细胞包括原始t细胞或其前体细胞、nkt细胞、或t细胞株。术语“原始t细胞(naivetcell)”是指外周血中尚未被相应抗原活化的成熟t细胞。这些细胞可以通过本领域已知的方法分离得到。例如,t细胞可以从不同组织器官获得,包括外周血、骨髓、淋巴组织、脾脏、脐带血、肿瘤组织。一个实施方案中,t细胞可以来自造血干细胞(hscs),包括来自骨髓、外周血或者脐带血,通过干细胞标记分子例如cd34而分离获得。一个实施方案中,t细胞可以来自诱导性多功能干细胞(ipscells),包括把特定基因或特定基因产物导入体细胞,使该体细胞转化为干细胞后,体外诱导分化成t细胞或其前体细胞。t细胞可以通过常用方法如密度梯度离心法而获得,密度梯度离心法的例子包括ficoll或者percoll密度离心。一个实施方案是利用血浆分离置换法(apheresis)或白细胞去除法(leukapheresis)从外周血获得富集的t细胞的产物。一个实施方案是用抗体标记特定细胞群后,通过磁珠分离的方式(如
优选地,所述t细胞前体细胞为造血干细胞。可以将本发明所述tcr的编码基因直接引入造血干细胞,然后转输到体内,进一步分化成为成熟t细胞;也可以将编码基因引入在体外特定条件下由造血干细胞分化成熟的t细胞中。
所述细胞可以被重悬于冻存溶液里置于液氮中保存。常用冻存溶液包括但不限包含10%(v/v)dmso和90%(v/v)人血清或胎牛血清。细胞以每分钟降低温度1℃的条件冻存于-80℃,然后保存于液氮罐的气相部分。其它冻存方法是把置于冻存液的细胞直接放入-80℃或液氮中进行冻存。
步骤2)所述的核酸为根据本发明所述的核酸,包括所述dna和rna。
所述转染包括物理方式、生物方式和化学方式。物理方式是通过磷酸钙沉淀、脂质体、微注射、电穿孔、基因枪等途径把tcr基因以dna或rna的形式导入细胞内。目前已有商业化的仪器,包括电转移仪(例如amaxanucleofector-ii(德国amaxabiosystems公司)、ecm830(btx)(美国harvardinstruments)、genepulserii(美国biorad公司)、multiporator(德国eppendort公司)。生物方式是通过dna或rna载体把tcr基因引入细胞内,逆转录病毒载体(例如γ逆转录病毒载体)是转染并插入外源基因片段到动物细胞(包括人细胞)的常用工具,其它病毒载体来源于慢病毒、痘病毒、单纯疱疹病毒、腺病毒以及腺病毒相关病毒等。化学方式是把多核苷酸引入细胞内,包括胶态分散系统,比如大分子复合物、纳米胶囊、微球体、微珠、微团和脂质体。无论以什么方式把tcr基因引入细胞,要用各种检测方法分析目的基因是否引入靶细胞内,所述检测方法包括常见的分子生物学方法(例如southern印迹和northern印迹、rt-pcr和pcr等),或者常见的生物化学方法(例如elisa和western印迹),以及本发明所提及的方法。
优选地,所述转染通过逆转录病毒载体或慢病毒载体进行。
转染后所述细胞的培养可以根据实际应用通过其各自的常规方法和条件进行。例如,t细胞通过表面的tcr/cd3复合体,以及辅助刺激分子(如cd28)共同激活后,可获得体外扩增。激活tcr、cd3和cd28的刺激物(如抗tcr、cd3或cd28的抗体)可以吸附在培养容器表面,或者共培养物(比如磁珠)表面,也可以直接加入细胞培养液中共同培养。另一个实施方案是将t细胞与滋养细胞共同培养,所述滋养细胞表达辅助刺激分子或者相应的配体,包括但不限于hla-a2、β2-微球蛋白、cd40、cd83、cd86、cd127、4-1bb。
依照通常的哺乳动物细胞体外培养的方法,t细胞培在适当培养条件下进行培养和扩增。例如,细胞达到70%以上融合状态(confluence)时可进行传代,一般2到3天换新鲜培养液。当细胞达到一定数目时直接使用,或按上述描述进行冻存。体外培养的时间可以是24小时之内,也可以长达14天或更长。冻存细胞解冻后可进行下一步应用。
在一个实施方案中,细胞可以在体外培养数小时到14天,或者之间任何小时数。t细胞培养条件包括使用基础培养液,包括但不限于rpmi1640、aim-v、dmem、mem、a-mem、f-12、x-vivo15和x-vivo。其它细胞生存和增殖所需要的条件包括但不限于使用血清(人或胎牛血清)、白介素-2(il-2)、胰岛素、ifn-γ、il-4、il-7、gm-csf、il-10、il-12、il-15、il-21、tgf-β和tnf-a,其它培养添加物(包括氨基酸、丙酮酸钠、维生素c、2-巯基乙醇、生长激素、生长因子)。细胞可置于适当的培养条件,例如,温度可处于37℃、32℃、30℃或者室温,并且空气条件可为(例如)含5%co2的空气。
本发明还提供了根据本发明所述的t细胞受体修饰的细胞在制备用于治疗或预防肿瘤和/或癌症的药物中的用途。
所述肿瘤和/或癌症是抗原her2/neu阳性的,并且是hla-a2阳性的,包括但不限于乳腺癌、卵巢癌、胃癌、食管癌、肠癌、胰腺癌、膀胱癌、肾癌、前列腺癌、子宫颈癌、子宫内膜癌、唾液腺癌、皮肤癌、肺癌、骨癌以及脑癌。
本发明还提供了根据本发明所述的t细胞受体修饰的细胞在制备用于检测宿主的肿瘤和/或癌症的药物中的用途。
在本发明的一个实施方案中,可将从宿主取出的肿瘤和/或癌症细胞的样本与本发明所述的t细胞受体修饰的细胞以一定浓度进行接触,根据二者的反应程度可以判断所述肿瘤和/或癌症是hla-a2阳性的还是hla-a2阴性的,以及是否表达抗原her2/neu。
本发明还提供了一种药物组合物,其中该药物组合物包括作为活性成分的根据本发明所述的t细胞受体修饰的细胞,及可药用辅料。
所述药物组合物优选包含每个患者每个疗程总剂量范围为1×103-1×109个细胞/kg体重的所述t细胞受体修饰的细胞,包括两个端点之间的任何数量的细胞。优选的是,每个疗程1-3天,每天施用1-3次。可以根据实际情况和需要对患者进行一个或多个疗程的治疗。
所述可药用辅料包括药用或生理载体、赋形剂、稀释剂(包括生理盐水、pbs溶液)、以及各种添加剂,包括糖类、脂类、多肽、氨基酸、抗氧化剂、佐剂、保鲜剂等。
所述药物组合物可通过合适的给药途径给药,其适于经动脉、静脉、皮下、皮内、瘤内、淋巴管内、淋巴结内、蛛网膜下腔内、骨髓内、肌肉内和腹膜内给药。
本发明还提供了一种治疗肿瘤和/或癌症的方法,包括对肿瘤和/或癌症患者施用根据本发明所述的t细胞受体修饰的细胞。
所述肿瘤和/或癌症是抗原her2/neu阳性的,并且是hla-a2阳性的,包括但不限于乳腺癌、卵巢癌、胃癌、食管癌、肠癌、胰腺癌、膀胱癌、肾癌、前列腺癌、子宫颈癌、子宫内膜癌、唾液腺癌、皮肤癌、肺癌、骨癌以及脑癌。
所述t细胞受体修饰的细胞的施用剂量优选为每个患者每个疗程总剂量范围为1×103-1×109个细胞/kg体重。优选的是,每个疗程1-3天,每天施用1-3次。可以根据实际情况和需要对患者进行一个或多个疗程的治疗。
所述t细胞受体修饰的细胞可通过合适的给药途径给药,其适于经动脉、静脉、皮下、皮内、瘤内、淋巴管内、淋巴结内、蛛网膜下腔内、骨髓内、肌肉内和腹膜内给药。
所述t细胞受体修饰的细胞进入治疗对象体内后可以消除表达her2/neu抗原的肿瘤细胞,和/或改变肿瘤组织的微环境而诱发其它抗肿瘤免疫反应。
本发明还提供了所述分离的t细胞受体用于检测接受该tcr修饰的t细胞(即tcr-t细胞)治疗的患者体内的该tcr-t细胞的增殖或生存情况的应用,从而进行药物代谢研究,和了解该tcr-t细胞的疗效和毒性。具体而言,tcr序列可作为引物,通过pcr方法检测体内携带此tcr的tcr-t细胞的数量。与荧光标记的hla/多肽复合物多聚体染色后用流式细胞法进行分析的方法相比,所述应用所需要的细胞量少,也更敏感。
以下通过例子的方式进一步解释或说明本发明的内容,但这些例子不应被理解为对本发明的保护范围的限制。
例子
以下除非特别说明,否则以下例子中所用实验方法均使用生物工程领域的常规实验流程、操作、材料和条件进行。
以下除非特别说明,否则各试剂的百分浓度(%)均指该试剂的体积百分浓度(%(v/v))。
材料和方法
细胞株:用于制备慢病毒颗粒的细胞株为293t细胞(atcccrl-3216)。用于检测tcr表型和功能的t细胞株为jurkat细胞(clonee6-1,atcctib-152)和j.rt3-t3.5细胞(atcctib-153)。用于提呈抗原多肽的提呈细胞株为t2细胞(174xcem.t2,atcccrl-1992)。用于检测tcr功能的肿瘤细胞株为人结肠癌colo205细胞(atccccl-222)和caco-2细胞(atcchtb-37)、人乳腺癌mda-mb-231细胞(atcchtb-26)、人卵巢癌sk-ov-3细胞(atcchtb-77)、人淋巴瘤细胞bjab(acc-757-dsmz)、人肺癌h647细胞(atcccrl-5824)和h1355细胞(atcccrl-5865)。细胞株用rpmi-1640完全培养基(lonza,cat#12-115f)维持培养,rpmi-1640完全培养基中加入10%小牛血清fbs(atcc30-2020),2mmol/ll-谷氨酸,100μg/ml青霉素和100μg/ml链霉素。
外周血:试验所用健康供者的人外周血制品均来自位于旧金山的pacific血液中心(#1pbmc和#2pbmc分别为来自apheresis法收集试剂盒的trima残留细胞组分#r32334和#r33941)。
体外诱导her2/neu369-377特异性杀伤t细胞(ctl):外周血经ficoll-paquepremium(sigma-alorich公司,cat#ge-17-5442-02)密度梯度离心(×400g)30分钟后获得单个核细胞(pbmc)。首先用荧光素fitc标记的抗hla-a2抗体(biolegend公司,cat#343303)染色检测细胞的hla-a2表型,流式细胞分析后提取阳性细胞的rna,逆转录为cdna并克隆到载体上,之后进行hla基因测序分析,确定细胞配型为hla-a*0201。hla-a2阳性的pbmc细胞培养在24-孔培养板的培养孔,培养液为上述rpmi-1640完全培养基。每孔2×10e6/mlpbmc,加入her2/neu369-377多肽(her2-e75,用peptide2.0合成,10μg/ml溶于dmso),浓度为1μg/ml。置于5%co2、37℃条件下的培养箱培养16-24小时后加入以下细胞因子,人il-2(peprtech公司,cat#200-02)100iu/ml,人il-7(peprotech公司,cat#200-07)5ng/ml,人il-15(peprotech公司,cat#200-15)5ng/ml。培养10到14天,对培养的t细胞进行抗原再刺激:在24-孔板中每孔加入10e6个培养细胞,同时加入2×10e6个经25μg/ml丝裂霉素c(santacruzbiotechnology公司,cat#sc-3514)处理2小时的hla-a2阳性的自体pbmc细胞作为滋养细胞,每孔加入终浓度为1μg/ml的her2/neu369-377多肽,培养过夜后加入il-2100iu/ml,il-75ng/ml,il-155ng/ml。经两轮上述抗原刺激和再刺激后,收集扩增的t细胞进行表型分析以及t细胞克隆。
流式细胞分析及单细胞分离:表达her2/neu369-377特异性tcr的t细胞表型是通过流式细胞来分析的。收集被检测的细胞置于1.5ml管(细胞数目约为10e5个),用1mldpbs溶液(2.7mmkcl,1.5mmkh2po4,136.9mmnacl,8.9mmna2hpo4·7h2o,ph7.4)洗一遍,并重置于100μl含有1%小牛血清的dpbs中,加入5μl荧光素apc标记的抗人cd8抗体(biolegend公司,cat#300912),以及10μl荧光素pe标记的her2-e75/hla-a2四聚体(her2-e75四聚体,mblinternationalco公司,cat#t01014)或者her2-e75/hla-a2五聚体(her2-e75五聚体,proimmune公司,cat#f214-2a-d),冰上孵育30分钟后用dpbs溶液洗两遍,重悬于100μlpbs溶液进行流式细胞分析。流式细胞仪为macsquantanalyzer10(miltenyibiotec公司),用flowjo软件(flowjo公司)进行结果分析。t细胞克隆是利用流式细胞分离仪(facssorter)进行单细胞分离后培养获得。对her2/neu369-377多肽抗原刺激过的pbmc用apc标记的抗人cd8抗体和pe标记的her2-e75/hla-a2五聚体染色,然后进行流式细胞分离(型号:sonycellsortersh800)。单个cd8+her2-e75/hla-a2五聚体+细胞被分选到96-孔培养板的单个培养孔后,加入经25μg/ml丝裂霉素c处理2小时的hla-a2阳性的自体pbmc细胞,每孔10e5个细胞,加入1μg/mlher2/neu369-377多肽培养过夜后,加入含有il-2100iu/ml、il-75ng/ml、il-155ng/ml的rpmi-1640完全培养液。每3-4天换新鲜含有所述细胞因子的培养液,显微镜下观察是否有t细胞克隆生长。收集增殖的t细胞,按上述方法进行抗原再刺激以获得足够数量的细胞,进行表型或功能检测,以及提取rna进行tcr基因的克隆。
t细胞功能检测:经her2/neu369-377多肽特异性tcr修饰的t细胞株的功能是通过t细胞表面cd69的表达来检测的。在96-孔板的每孔中加入10e5个转染tcr基因的t细胞以及10e5个t2细胞,在100μl/每孔rpmi-1640完全培养基中进行混合培养,各试验组为复孔。再加入不同浓度(分别为1μg/ml、0.5μg/ml、0.1μg/ml、0.05μg/ml、0.01μg/ml、0.005μg/ml、0.001μg/ml和0μg/ml)her2/neu369-377多肽后置于5%co2、37℃条件下的孵育箱过夜培养。收集细胞并悬浮于dpbs+1%fbs溶液中,用apc标记的抗人cd69抗体进行染色后进行流式细胞分析。经her2/neu369-377抗原刺激后表达cd69的t细胞被认为其携带并表达her2/neu369-377特异性的tcr。her2/neu369-377特异性ctl克隆以及pbmc中的转染tcr基因的原代t细胞的功能,是通过检测抗原刺激后细胞上清中分泌的γ干扰素来确定的。her2/neu369-377特异性ctl克隆或者转染tcr基因的pbmc细胞分别与靶细胞在96孔板中,在100μl/每孔rpmi-1640完全培养基中进行混合培养,靶细胞是加入10倍稀释浓度的her2/neu369-377多肽的t2细胞,或者各种肿瘤细胞株细胞,各试验组为复孔。抗体功能阻断试验中,细胞培养孔中同时加入10μg/ml抗人cd8抗体(biolegend公司,cat#300912),或者抗hla-abc抗体(w6/32克隆,biolegend公司,cat#311402),细胞置于5%co2、37℃条件下的孵育箱过夜培养。18-24小时收集细胞上清,并用人ifn-γelisaread-set-go试剂盒(ebioscience公司,cat#88-7316)对上清中的ifn-γ进行检测。
获得单克隆tcr基因:利用zymoquick-rnamicroprep试剂盒(zymoresearch公司,cat#r1050)从t细胞克隆提纯总rna,以此为模板利用smarterrace5’/3’试剂盒获得cdna(美国takarabio公司,cat#634858)。用5’-cds引物和tcrβ链3’引物5’-gcctctggaatcctttctcttg-3’(seqidno:26)以及α链3’引物5’-tcagctggaccacagccgcag-3’(seqidno:27)进行pcr,扩增出tcrα和β全序列基因片段,并分别克隆到prace载体(美国takarabio,cat#634858)上。转化感受态细菌stellar(美国takarabio公司,cat#636763)并获得质粒后进行测序。
重组tcr慢病毒表达载体的制备:用于表达tcr的病毒载体为复制缺陷型慢病毒载体,包括:表达gfp的慢病毒载体pcdh-ef1α-mcs-(pgk-gfp),可购自systembiosciences公司(cat#cd811a-1);以及不表达gfp的载体pcdh-ef1α-mcs,通过采用本领域常规技术去除pcdh-ef1α-mcs-(pgk-gfp)载体上的pgk启动子及gfp基因而得到。根据所获得的tcr基因序列,合成tcrβ链和α链以及之间可切割的f2a序列和furin酶切片段的全基因序列,并链接到所述载体的ef-1α启动子下游的多克隆位点,插入tcr的转录顺序依次为tcrβ链(无终止密码子),furin酶切片段,f2a片段,tcrα链(方法参见文献“genether.2008nov;15(21):1411–1423”)。表达gfp的载体是被反向的pgk启动子驱动的。不表达gfp的载体则是去除了pgk启动子以及gfp片段。
重组tcr慢病毒颗粒的制备:tcr慢病毒颗粒是通过lipofectaine3000转染试剂(thermofisher公司,cat#l3000001)转染293t细胞而获得的。依照厂家说明书准备293t细胞以及转染流程。转染在96孔培养板进行,首先用opti-mem1培养液(thermofisher公司,cat#51985091)制备转染质粒的脂质体溶液,依照厂家说明在250μl培养液中加入p3000试剂(p3000reagent)、以及tcr慢病毒载体质粒和pcdh系统的病毒包装质粒(sbi公司,cat#lv500a-1),另外250μl培养液中加入lipofectaine3000试剂,混合孵育15分钟后加入293t细胞培养孔。5%co2、37℃条件下培养16小时,换含10%fbs的dmem培养液(thermofisher公司,cat#11965092),继续培养24小时和48小时后分别收集细胞上清,2000g离心后,用0.4μm过滤膜过滤后得到的病毒颗粒用于感染细胞。
重组tcr慢病毒转染人t细胞:冻存的原代pbmc细胞解冻后在rpmi-1640完全培养液中培养24小时,经ficoll-paquepremium密度梯度离心(×400g)30分钟去除死细胞,置于用2μg/ml抗人cd3抗体(biolegend公司,okt3克隆cat#317303)和2μg/ml抗人cd28抗体(biolegend公司,cat#302914)处理(其中每孔加入100μl含有上述cd3抗体和cd28抗体的dpbs溶液)24小时的24孔板培养孔中,细胞浓度为2×10e6/ml,培养48小时后收集细胞,重悬于新鲜的重组tcr慢病毒颗粒上清液中置于24孔板的孔中,加入4ng/ml聚凝胺(santacruzbiotechnology公司,cat#sc-134220)后,于32℃,1000g离心2小时,用含有il-2100iu/ml、il-75ng/ml、il-155ng/ml的rpmi-1640完全培养液置换一半病毒上清后继续培养,每3天换新鲜含有上述细胞因子的培养液。一般72小时后可进行表型和功能检测。转染t细胞株也依照上述步骤进行,如果病毒载体上带有gfp标记,一般转染后48小时即可在荧光显微镜下观察到gfp阳性细胞。
实施例1:从hla-a2阳性的正常供体外周血诱导her2/neu369-377多肽特异性杀伤t细胞
本实施例用1μg/ml的低浓度her2/neu369-377多肽经过两轮体外刺激从正常pbmc(#2pbmc)中诱导出多肽特异性杀伤t细胞,并进行流式细胞分析及单细胞分离。具体方法如上文所述。结果如下:
图1a右图显示0.013%的淋巴细胞为可结合her2/neu369-377/hla-a2五聚体(即her2-e75五聚体)的cd8阳性杀伤性t细胞,左图中没有经her2多肽刺激的对照细胞没有出现cd8阳性五聚体阳性细胞。结果说明在自然t细胞库中,识别her2/neu369-377抗原多肽的特异性t细胞数量很少。尽管数量少,这群可识别her2/neu369-377多肽的t细胞仍可被清晰地区分出来。另外根据结合her2-e75五聚体的荧光强度,阳性细胞中又包含高亲和性t细胞和低亲和性t细胞。通过流式细胞分离出453个cd8阳性五聚体阳性细胞后进行单克隆培养,经过两轮抗原多肽再刺激以及细胞因子扩增,从这453个分离出的单个t细胞中仅获得一个增殖的t细胞克隆her2ctl克隆1b5(称为her2ctl1b5)。图1b右图显示此纯化的cd8+ctl克隆可结合her2/neu369-377/hla-a2四聚体(即her2-e75四聚体)。每孔5x103个her2ctl1b5细胞与被5x103个t2细胞提呈的不同浓度her2/neu369-377抗原多肽(her2/neu369-377抗原多肽从1μg/ml开始进行10倍稀释,从而得到终浓度为1μg/ml、0.1μg/ml、0.01μg/ml、0.001μg/ml的不同组)混合培养后,检测上清中t细胞分泌的ifn-γ,用以确定此t细胞克隆特异性识别her2/neu369-377多肽的功能。肿瘤细胞培养孔中加入5x103个colo205细胞进行混合培养后,检测上清中t细胞分泌的ifn-γ。图1c结果显示,t细胞克隆1b5可以被最低浓度为1ng/ml的抗原多肽所激活,而且t细胞的识别功能与抗原多肽的浓度成正相关(具有剂量依赖性),说明此t细胞克隆特异性识别被hla-a2所提呈的her2/neu369-377多肽。更重要的是,此t细胞克隆还识别一株hla-a2+her2/neu+的结肠癌细胞株colo205。因此,此t细胞克隆her2tcr-1b5不仅可识别her-e75多肽,还可以被colo205细胞激活而分泌ifn-γ。
实施例2:her2/neu369-377多肽特异性tcr全序列的获得和鉴定
本实施例直接从由实施例1得到的her2/neu369-377多肽特异性ctl克隆中获得包含相匹配(即,两条链可共同组成识别抗原多肽的功能性tcr)的α和β链的完整tcr基因序列,其编码的tcr称为“her2tcr-1b5”。该tcr的α链的氨基酸序列如seqidno:2所示,编码序列如seqidno:1所示,并且该tcr的β链的氨基酸序列如seqidno:4所示,编码序列如seqidno:3所示。此tcr存在于hla-a2阳性正常人的外周t细胞库中,不会对微量表达her2/neu蛋白的正常细胞产生交叉反应而导致自身免疫反应。具体检测方法如上文所述。结果如下:
为了检测所获tcr的抗原特异性及其功能,tcrα链和β链序列被克隆到复制缺陷型慢病毒表达载体中。图2显示所构建的tcr慢病毒载体结构片段示意图。α链和β链由可切除的furin识别片段和f2a多肽片段相联结。为了追踪被tcr慢病毒载体转染的t细胞,该慢病毒载体(图中示为“pcdh-ef1α-her2tcr-(pgk-gfp)载体”)可同时表达gfp,由反向的pgk启动子所驱动(图2上方)。另一种表达载体(图中示为“pcdh-ef1α-her2tcr载体”)(图2下方)去除了gfp及其启动子序列。通过减小载体长度以增加慢病毒颗粒的产出,又避免了两个启动子之间的相互影响,从而增加tcr的表达量。
将以下序列分别连接至上述表达gfp的载体和不表达gfp的载体:i)由可切割性连接多肽链接的、在恒定区增加1个二硫键的tcrβ链和α链的核苷酸序列(seqidno:13)(对应的tcr为her2tcr-1b5-dis,氨基酸序列如seqidno:14所示);ii)由可切割性连接多肽链接的、恒定区由人源序列替换为鼠源序列的tcrβ链和α链的核苷酸序列(seqidno:15)(对应的tcr为her2tcr-1b5-mc,氨基酸序列如seqidno:16所示);iii)由可切割性连接多肽链接的原始tcrβ链和α链的核苷酸序列(seqidno:23)(对应的tcr为her2tcr-1b5-wt,编码的氨基酸序列如seqidno:24所示);从而得到6种重组慢病毒载体:
1)her2tcr-1b5-dis重组慢病毒载体(携带gfp);
2)her2tcr-1b5-disw/ogfp重组慢病毒载体(不携带gfp);
3)her2tcr-1b5-mc重组慢病毒载体(携带gfp);
4)her2tcr-1b5-mcw/ogfp重组慢病毒载体(不携带gfp)。
5)her2tcr-1b5-wt重组慢病毒载体(携带gfp);
6)her2tcr-1b5-wtw/ogfp重组慢病毒载体(不携带gfp)。
各her2tcr基因片段通过pcr扩增后,分别克隆到上述两种慢病毒载体(即pcdh-ef1α-mcs-(pgk-gfp)和pcdh-ef1α-mcs)的ef1-α启动子下游:her2tcr-1b5-dis和tcr-1b5-wt的tcr片段均是用5’引物5’-agagctagcgaattcaacatggatacctggctcgtatg-3’(seqidno:28)和3’引物5’-gttgattgtcgacgccctcagctggaccacagccgcag-3’(seqidno:29)扩增而得。her2tcr-1b5-mc的tcr片段是用5’引物5’-agagctagcgaattcaacatggatacctggctcgtatg-3’(seqidno:30)和3’引物5’-gttgattgtcgacgccctcaactggaccacagcct-3’(seqidno:31)扩增而得。pcr使用q5高保真pcr试剂盒(neb,cat#m0543s),反应条件为98℃30秒后,进行25个循环98℃10秒,65℃10秒,72℃3分钟。获得的tcr片段克隆到pcdh-ef1α-mcs-(pgk-gfp)载体或pcdh-ef1α-mcs载体的ef1α启动子下游的mcs区域。
将构建得到的重组tcr慢病毒表达载体按前述方法制备得到各自的重组tcr慢病毒颗粒。
按照上述方式能够得到两种不同方式修饰的tcr,her2tcr-1b5-dis是通过点突变在tcr恒定区增加一个二硫键,方法在文献“cancerres.2007apr15;67(8):3898-903.”中描述,其全文通过引用方式并入本文。her2tcr-1b5-mc是用小鼠tcr恒定区序列置换相应的人tcr恒定区序列,方法在文献“eur.j.immunol.200636:3052–3059”中描述,其全文通过引用方式并入本文。图3a显示,用携带her2tcr-1b5tcr和gfp基因的慢病毒(即,上述her2tcr-1b5-dis重组慢病毒载体和her2tcr-1b5-mc重组慢病毒载体)转染t细胞株后,一部分gfp+细胞可以结合her2-e75四聚体,说明这些被慢病毒转染的t细胞可以识别被hla-a2分子提呈的her2/neu369-377多肽。由所得数据计算,如果tcr恒定区增加一个二硫键,37.5%(13.2/(13.2+22)×100%)被转染的t细胞(gfp+)所表达的tcr可以识别her2/neu多肽抗原(图3a左图)。而tcr恒定区被小鼠tcr相应的恒定区所置换的,则有60%(34.6/(34.6+23)×100%)被转染的t细胞(gfp+)所表达的tcr可以识别her2/neu多肽抗原(图3a右图)。与外源tcrα链和β链经二硫键修饰后得到的tcr相比,外源tcr恒定区被相应小鼠tcr恒定区所置换而形成的杂交tcr可以进一步减少与内源tcr的错配几率。图3b显示,不论转染只表达beta链的j.rt-t3.5细胞还是同时表达α链和β链的jurkat细胞,恒定区被小鼠恒定区序列置换后得到的外源tcr(her2tcr-1b5-mc)的表达量明显高于只在恒定区增加一个二硫键的tcr(her2tcr-1b5-dis)。图3c显示,表达外源her2tcr-1b5的t细胞株可以被t2细胞提呈的her2/neu369-377多肽所激活而表达cd69,说明此tcr具有识别her2/neu369-377多肽抗原的功能。恒定区被小鼠恒定区序列置换后得到的外源tcr(her2tcr-1b5-mc)的表达量不仅增加,对多肽抗原的特异性识别能力也明显高于经额外二硫键修饰的tcr(her2tcr-1b5-dis),可以被更低浓度的her2/neu369-377多肽所激活。然而,激活表达her2tcr-1b5-mc的jurkat细胞的最低多肽浓度约为0.05μg/ml,比图1c显示的激活her2ctl1b5克隆的多肽浓度高约50倍。这说明jurkat细胞表达的her2tcr-1b5tcr识别多肽抗原的能力(avidity)要明显低于cd8+ctl上表达的her2tcr-1b5tcr。可能的一个原因是jurkat细胞不表达cd8分子,而cd8对her2tcr-1b5的识别功能起着重要的辅助作用。
实施例3:正常外周血t细胞经her2tcr-1b5-mc重组慢病毒转染后表达可识别her2/neu369-377多肽的特异性tcr
为了进一步验证本发明所获得的tcr能否在原代t细胞表达并具有识别her2/neu抗原多肽的功能,用携带her2tcr-1b5-mc基因的重组慢病毒颗粒(her2tcr-1b5-mc重组慢病毒载体)转染经cd3/cd28抗体活化的、来自两个不同正常供体的外周血t细胞,7天后收集细胞进行her2-e75四聚体染色。具体方法如上文所述。结果如下:
图4a显示,两个供体外周血单个核细胞(分别为#1pbmc和#2pbmc)中均有淋巴细胞可以结合her2-e75四聚体,说明这些细胞表达的her2tcr-1b5-mc可以特异性识别被hla-a2提呈的her2/neu抗原多肽。结果还显示,her2-e75四聚体阳性细胞(即表达her2tcr-1b5-mc)中,大多数阳性细胞为cd8+t杀伤细胞,小部分阳性细胞为cd8-的淋巴细胞,这很可能是cd4+的t辅助细胞。cd8+ctl结合her2-e75四聚体的荧光强度(#2pbmc样本中的geom.mean为1469,#1pbmc样本中的geom.mean为1404),也明显大于cd4+t细胞(#2pbmc样本中的geommean为560,#1pbmc样本中的geom.mean为504)。如果慢病毒感染cd8+和cd4+t细胞的转染效率一样,说明一部分表达在cd4+细胞上的外源her2/neu369-377特异性tcr并不能结合her2-e75四聚体,即使结合,亲和力也低于表达在cd8+t细胞上的外源转染tcr。这也进一步说明转染的tcr需要cd8分子的辅助功能才能有效结合her2/hla-a2复合物。
在96-孔板的每孔中加入10e5个转染tcr的pbmc细胞,与不同浓度被t2细胞(每孔10e5个)提呈的her2/neu369-377抗原多肽(her2/neu369-377抗原多肽从0.1μg/ml开始进行10倍稀释,从而得到终浓度为0.1μg/ml、0.01μg/ml、0.001μg/ml的不同组)混合培养后,检测上清中t细胞分泌的ifn-γ,用以确定此表达tcr的pbmc细胞特异性识别her2/neu369-377多肽的功能。对照组靶细胞为提呈1μg/ml的可以结合hla-a2分子的ebv病毒抗原多肽lmp2426-434的t2细胞。图4b显示,表达her2tcr-1b5-mc的pbmc可以被t2细胞提呈的her2/neu369-377抗原多肽所激活而分泌ifn-γ,说明表达外源her2tcr-1b5-mc的原代t细胞可以特异性识别被hla-a2分子提呈的her2/neu369-377多肽。识别抗原多肽的能力与外源tcr在t细胞上的表达量相关。其识别抗原多肽的最大半反应(half-maximumreaction,ec50)多肽浓度经曲线拟合推算为约6.9ng/ml(ic50toolkit,http://www.ic50.tk/),尽管此反应敏感度低于识别病毒抗原等外源抗原的高亲和性tcr的ec50(ec50约10e-10m)(参见文献“cancerresearch1998,58.4902-4908”和“humangenetherapy2014,25:730–739”),但仍处于可识别常见肿瘤相关抗原的tcr亲和力范围之内(如文献“eurjimmunol(2012)42:3174–9”所述)。
图4c显示t细胞与t2细胞提呈的抗原多肽(t2+her2-e75,即her2/neu369-377多肽)共培养时加入抗人cd8抗体后,t细胞分泌ifn-γ的功能被显著抑制。这说明cd8分子的辅助作用对于外源tcr识别her2/neu369-377抗原多肽至关重要,也显示本发明所述的her2tcr-1b5是cd8功能依赖型tcr。
实施例4:正常外周血t细胞经her2tcr-1b5-mc重组慢病毒转染后表达的her2/neu369-377多肽特异性tcr可识别hla-a2+her2/neu+肿瘤细胞
在96-孔板的每孔中加入10e5个经her2tcr-1b5-mcw/ogfp重组慢病毒转染后的pbmc(图中所示为转染her2tcr-1b5的#2pbmc),或者10e5个不转染tcr的对照#2pbmc,与10e5个不同肿瘤细胞株混合培养,之后检测上清液中分泌的ifn-γ。具体方法如上文所述。结果如下:
图5a显示,表达her2tcr-1b5-mc的t细胞均可被hla-a2+her2/neu+的肿瘤细胞株所激活并分泌ifn-γ,肿瘤细胞株包括结肠癌colo205细胞、乳腺癌mda-mb-231细胞、结肠癌caco-2细胞、以及肺癌h1355细胞。而hla-a2-her2/neu+的卵巢癌sk-ov3细胞、肺癌h647细胞、以及hla-a2+her2/neu-的淋巴瘤bjab细胞却不能激活转染her2tcr-1b5-mctcr的t细胞。说明her2tcr-1b5-mctcr可以特异性识别肿瘤细胞表面被hla-a2提呈的her2/neu抗原。来源同一供体pbmc,平行培养但没有转染her2tcr-1b5-mc的对照组t细胞不能被所列肿瘤细胞株所激活,说明对肿瘤细胞的反应不是非特异性的。图5b显示,表达her2tcr-1b5-mctcr的t细胞(转染her2tcr-1b5的#2pbmc)对colo205细胞表现出明显的反应,而且识别功能可以被抗cd8抗体以及抗hla一类分子的抗体几乎完全阻断,这进一步说明识别肿瘤细胞表面her2/neu抗原的效应细胞为cd8+杀伤性t细胞,其特异性抗原识别功能依赖于cd8的辅助功能,这也和识别t2细胞提呈的her2/neu369-377多肽抗原时her2tcr-1b5-mctcr的表现是一致的。
讨论
不同肿瘤细胞株对特异性t细胞的反应敏感性差异可能与肿瘤细胞表达不同水平her2/neu抗原多肽/hla-a2复合体有关,也可能与肿瘤细胞本身对t细胞功能不同的抑制作用有关。尽管特异性识别her2/neu369-377多肽的高亲和性tcr可以通过her2/neu369-377多肽体外诱导而获得,但这些高亲和性tcr往往不能识别肿瘤细胞所提呈的her2/neu抗原(cancerres.1998;58:4902–4908.cancerimmunol.immunother.2008;57:271–280)。一个原因可能是外源her2/neu369-377多肽结合hla-a2分子的构型与细胞内所提呈的多肽/hla复合物的构型有所不同(参见文献“journalofimmunology,2008,180:8135–8145”)。另一个可能原因为,her2/neu369-377多肽作为模拟表位(mimotope)抗原,所诱导的特异性tcr既可识别her2/neu369-377多肽,也可识别被肿瘤细胞提呈的相似多肽,例如her2/neu373-382多肽(参见文献“jimmunol.2013jan1;190(1):479–488”),然而高亲和性tcr虽然对hla-a2提呈的her2/neu369-377多肽具有高亲和力,却不能有效识别相应的被肿瘤细胞提呈的模拟表位多肽。本发明所述的特异性识别her2/neu369-377多肽的tcr,不仅靶向肿瘤细胞所提呈的her2/neu369-377多肽,也可能同时识别被肿瘤细胞所提呈的来源于her2/neu的其它模拟表位多肽。
由于识别自身抗原的高亲和性t细胞大多被中枢耐受机制所清除,外周t细胞库中自然存在的可以识别her2/neu抗原的tcr大多为中低亲和性。另外一个可以识别肿瘤细胞的cd8功能非依赖型的高亲和性tcr是来自经her2/neu373-382多肽特异性t细胞群的多个α链和β链进行配对后,通过功能检测筛选而出(参见文献“humangenetherapy2024,25:730–739”;wo/2016/133779)。由于不是从特异性的单克隆t细胞直接获得,不能确定此tcr是否存在于外周自然t细胞库。一般认为,高亲和性t细胞的过继转输治疗的疗效要优于靶向同一抗原的低亲和性t细胞(参见文献“clinexpimmunol(2015)180:255–70”)。然而,高亲和性tcr本身容易产生识别自身抗原的自身免疫性反应(参见文献“blood(2009)114:535–46”),没有经过中枢耐受机制筛选的tcr也会识别抗原低表达的正常组织,或者针对其它类似的自身抗原表位产生交叉反应的脱靶毒性(参见文献“scitranslmed(2013)5:197ra103.blood(2013)122:863–71”)。选择高亲和性tcr的另一个原因是这些tcr的功能不依赖cd8的辅助功能,因而可以通过转染cd4+t细胞而获得对cd8+杀伤t细胞功能的辅助作用。对于cd8功能依赖型tcr,通过表达载体同时表达tcr和外源cd8分子也可以达到同样的目的。
总之,本发明提供了一种从hla-a2+的自体外周t细胞库中诱导而来的her2/neu373-382多肽特异性tcrα链和β链全序列,经转染后表达此tcr及恒定区被修饰的tcr的原代杀伤性t细胞可以识别多种hla-a2+her2/neu+的肿瘤细胞。为开发和临床应用过继转输经特异性tcr修饰的t细胞来治疗肿瘤提供了新的方法和途径。
sequencelisting
<110>杭州康万达医药科技有限公司
合成免疫股份有限公司(synimmune,inc.)
<120>分离的t细胞受体、其修饰的细胞、编码核酸、表达载体、制备方法、药物组合物和应用
<130>fi-173891-59:52/c
<160>31
<170>patentinversion3.5
<210>1
<211>831
<212>dna
<213>人(homosapiens)
<400>1
atggcatgccctggcttcctgtgggcacttgtgatctccacctgtcttgaatttagcatg60
gctcagacagtcactcagtctcaaccagagatgtctgtgcaggaggcagagaccgtgacc120
ctgagctgcacatatgacaccagtgagagtgattattatttattctggtacaagcagcct180
cccagcaggcagatgattctcgttattcgccaagaagcttataagcaacagaatgcaaca240
gagaatcgtttctctgtgaacttccagaaagcagccaaatccttcagtctcaagatctca300
gactcacagctgggggatgccgcgatgtatttctgtgcttataggctctttaataatgca360
ggcaacatgctcacctttggagggggaacaaggttaatggtcaaaccccatatccagaac420
cctgaccctgccgtgtaccagctgagagactctaaatccagtgacaagtctgtctgccta480
ttcaccgattttgattctcaaacaaatgtgtcacaaagtaaggattctgatgtgtatatc540
acagacaaaaccgtgctagacatgaggtctatggacttcaagagcaacagtgctgtggcc600
tggagcaacaaatctgactttgcatgtgcaaacgccttcaacaacagcattattccagaa660
gacaccttcttccccagcccagaaagttcctgtgatgtcaagctggtcgagaaaagcttt720
gaaacagatacgaacctaaactttcaaaacctgtcagtgattgggttccgaatcctcctc780
ctgaaagtggccgggtttaatctgctcatgacgctgcggctgtggtccagc831
<210>2
<211>277
<212>prt
<213>人(homosapiens)
<400>2
metalacysproglypheleutrpalaleuvalileserthrcysleu
151015
gluphesermetalaglnthrvalthrglnserglnproglumetser
202530
valglnglualagluthrvalthrleusercysthrtyraspthrser
354045
gluserasptyrtyrleuphetrptyrlysglnproproserarggln
505560
metileleuvalileargglnglualatyrlysglnglnasnalathr
65707580
gluasnargpheservalasnpheglnlysalaalalysserpheser
859095
leulysileseraspserglnleuglyaspalaalamettyrphecys
100105110
alatyrargleupheasnasnalaglyasnmetleuthrpheglygly
115120125
glythrargleumetvallysprohisileglnasnproaspproala
130135140
valtyrglnleuargaspserlysserserasplysservalcysleu
145150155160
phethrasppheaspserglnthrasnvalserglnserlysaspser
165170175
aspvaltyrilethrasplysthrvalleuaspmetargsermetasp
180185190
phelysserasnseralavalalatrpserasnlysserasppheala
195200205
cysalaasnalapheasnasnserileileprogluaspthrphephe
210215220
proserproglusersercysaspvallysleuvalglulysserphe
225230235240
gluthraspthrasnleuasnpheglnasnleuservalileglyphe
245250255
argileleuleuleulysvalalaglypheasnleuleumetthrleu
260265270
argleutrpserser
275
<210>3
<211>939
<212>dna
<213>人(homosapiens)
<400>3
atggatacctggctcgtatgctgggcaatttttagtctcttgaaagcaggactcacagaa60
cctgaagtcacccagactcccagccatcaggtcacacagatgggacaggaagtgatcttg120
cgctgtgtccccatctctaatcacttatacttctattggtacagacaaatcttggggcag180
aaagtcgagtttctggtttccttttataataatgaaatctcagagaagtctgaaatattc240
gatgatcaattctcagttgaaaggcctgatggatcaaatttcactctgaagatccggtcc300
acaaagctggaggactcagccatgtacttctgtgccagcagtaccgatttggtaggggtc360
gaagaagctttctttggacaaggcaccagactcacagttgtagaggacctgaacaaggtg420
ttcccacccgaggtcgctgtgtttgagccatcagaagcagagatctcccacacccaaaag480
gccacactggtatgcctggccacaggcttctaccccgaccacgtggagctgagctggtgg540
gtgaatgggaaggaggtgcacagtggggtcagcacagacccgcagcccctcaaggagcag600
cccgccctcaatgactccagatactgcctgagcagccgcctgagggtctcggccaccttc660
tggcagaacccccgcaaccacttccgctgtcaagtccagttctacgggctctcggagaat720
gacgagtggacccaggatagggccaaacccgtcacccagatcgtcagcgccgaggcctgg780
ggtagagcagactgtggcttcacctccgagtcttaccagcaaggggtcctgtctgccacc840
atcctctatgagatcttgctagggaaggccaccttgtatgccgtgctggtcagtgccctc900
gtgctgatggccatggtcaagagaaaggattccagaggc939
<210>4
<211>313
<212>prt
<213>人(homosapiens)
<400>4
metaspthrtrpleuvalcystrpalailepheserleuleulysala
151015
glyleuthrgluprogluvalthrglnthrproserhisglnvalthr
202530
glnmetglyglngluvalileleuargcysvalproileserasnhis
354045
leutyrphetyrtrptyrargglnileleuglyglnlysvalgluphe
505560
leuvalserphetyrasnasngluileserglulyssergluilephe
65707580
aspaspglnpheservalgluargproaspglyserasnphethrleu
859095
lysileargserthrlysleugluaspseralamettyrphecysala
100105110
serserthraspleuvalglyvalgluglualaphepheglyglngly
115120125
thrargleuthrvalvalgluaspleuasnlysvalpheproproglu
130135140
valalavalphegluproserglualagluileserhisthrglnlys
145150155160
alathrleuvalcysleualathrglyphetyrproasphisvalglu
165170175
leusertrptrpvalasnglylysgluvalhisserglyvalserthr
180185190
aspproglnproleulysgluglnproalaleuasnaspserargtyr
195200205
cysleuserserargleuargvalseralathrphetrpglnasnpro
210215220
argasnhispheargcysglnvalglnphetyrglyleusergluasn
225230235240
aspglutrpthrglnaspargalalysprovalthrglnilevalser
245250255
alaglualatrpglyargalaaspcysglyphethrserglusertyr
260265270
glnglnglyvalleuseralathrileleutyrgluileleuleugly
275280285
lysalathrleutyralavalleuvalseralaleuvalleumetala
290295300
metvallysarglysaspserarggly
305310
<210>5
<211>831
<212>dna
<213>人工序列(artificialsequence)
<400>5
atggcatgccctggcttcctgtgggcacttgtgatctccacctgtcttgaatttagcatg60
gctcagacagtcactcagtctcaaccagagatgtctgtgcaggaggcagagaccgtgacc120
ctgagctgcacatatgacaccagtgagagtgattattatttattctggtacaagcagcct180
cccagcaggcagatgattctcgttattcgccaagaagcttataagcaacagaatgcaaca240
gagaatcgtttctctgtgaacttccagaaagcagccaaatccttcagtctcaagatctca300
gactcacagctgggggatgccgcgatgtatttctgtgcttataggctctttaataatgca360
ggcaacatgctcacctttggagggggaacaaggttaatggtcaaaccccatatccagaac420
cctgaccctgccgtgtaccagctgagagactctaaatccagtgacaagtctgtctgccta480
ttcaccgattttgattctcaaacaaatgtgtcacaaagtaaggattctgatgtgtatatc540
acagacaaatgcgtgctagacatgaggtctatggacttcaagagcaacagtgctgtggcc600
tggagcaacaaatctgactttgcatgtgcaaacgccttcaacaacagcattattccagaa660
gacaccttcttccccagcccagaaagttcctgtgatgtcaagctggtcgagaaaagcttt720
gaaacagatacgaacctaaactttcaaaacctgtcagtgattgggttccgaatcctcctc780
ctgaaagtggccgggtttaatctgctcatgacgctgcggctgtggtccagc831
<210>6
<211>277
<212>prt
<213>人工序列(artificialsequence)
<400>6
metalacysproglypheleutrpalaleuvalileserthrcysleu
151015
gluphesermetalaglnthrvalthrglnserglnproglumetser
202530
valglnglualagluthrvalthrleusercysthrtyraspthrser
354045
gluserasptyrtyrleuphetrptyrlysglnproproserarggln
505560
metileleuvalileargglnglualatyrlysglnglnasnalathr
65707580
gluasnargpheservalasnpheglnlysalaalalysserpheser
859095
leulysileseraspserglnleuglyaspalaalamettyrphecys
100105110
alatyrargleupheasnasnalaglyasnmetleuthrpheglygly
115120125
glythrargleumetvallysprohisileglnasnproaspproala
130135140
valtyrglnleuargaspserlysserserasplysservalcysleu
145150155160
phethrasppheaspserglnthrasnvalserglnserlysaspser
165170175
aspvaltyrilethrasplyscysvalleuaspmetargsermetasp
180185190
phelysserasnseralavalalatrpserasnlysserasppheala
195200205
cysalaasnalapheasnasnserileileprogluaspthrphephe
210215220
proserproglusersercysaspvallysleuvalglulysserphe
225230235240
gluthraspthrasnleuasnpheglnasnleuservalileglyphe
245250255
argileleuleuleulysvalalaglypheasnleuleumetthrleu
260265270
argleutrpserser
275
<210>7
<211>939
<212>dna
<213>人工序列(artificialsequence)
<400>7
atggatacctggctcgtatgctgggcaatttttagtctcttgaaagcaggactcacagaa60
cctgaagtcacccagactcccagccatcaggtcacacagatgggacaggaagtgatcttg120
cgctgtgtccccatctctaatcacttatacttctattggtacagacaaatcttggggcag180
aaagtcgagtttctggtttccttttataataatgaaatctcagagaagtctgaaatattc240
gatgatcaattctcagttgaaaggcctgatggatcaaatttcactctgaagatccggtcc300
acaaagctggaggactcagccatgtacttctgtgccagcagtaccgatttggtaggggtc360
gaagaagctttctttggacaaggcaccagactcacagttgtagaggacctgaacaaggtg420
ttcccacccgaggtcgctgtgtttgagccatcagaagcagagatctcccacacccaaaag480
gccacactggtatgcctggccacaggcttctaccccgaccacgtggagctgagctggtgg540
gtgaatgggaaggaggtgcacagtggggtctgcacagacccgcagcccctcaaggagcag600
cccgccctcaatgactccagatactgcctgagcagccgcctgagggtctcggccaccttc660
tggcagaacccccgcaaccacttccgctgtcaagtccagttctacgggctctcggagaat720
gacgagtggacccaggatagggccaaacccgtcacccagatcgtcagcgccgaggcctgg780
ggtagagcagactgtggcttcacctccgagtcttaccagcaaggggtcctgtctgccacc840
atcctctatgagatcttgctagggaaggccaccttgtatgccgtgctggtcagtgccctc900
gtgctgatggccatggtcaagagaaaggattccagaggc939
<210>8
<211>313
<212>prt
<213>人工序列(artificialsequence)
<400>8
metaspthrtrpleuvalcystrpalailepheserleuleulysala
151015
glyleuthrgluprogluvalthrglnthrproserhisglnvalthr
202530
glnmetglyglngluvalileleuargcysvalproileserasnhis
354045
leutyrphetyrtrptyrargglnileleuglyglnlysvalgluphe
505560
leuvalserphetyrasnasngluileserglulyssergluilephe
65707580
aspaspglnpheservalgluargproaspglyserasnphethrleu
859095
lysileargserthrlysleugluaspseralamettyrphecysala
100105110
serserthraspleuvalglyvalgluglualaphepheglyglngly
115120125
thrargleuthrvalvalgluaspleuasnlysvalpheproproglu
130135140
valalavalphegluproserglualagluileserhisthrglnlys
145150155160
alathrleuvalcysleualathrglyphetyrproasphisvalglu
165170175
leusertrptrpvalasnglylysgluvalhisserglyvalcysthr
180185190
aspproglnproleulysgluglnproalaleuasnaspserargtyr
195200205
cysleuserserargleuargvalseralathrphetrpglnasnpro
210215220
argasnhispheargcysglnvalglnphetyrglyleusergluasn
225230235240
aspglutrpthrglnaspargalalysprovalthrglnilevalser
245250255
alaglualatrpglyargalaaspcysglyphethrserglusertyr
260265270
glnglnglyvalleuseralathrileleutyrgluileleuleugly
275280285
lysalathrleutyralavalleuvalseralaleuvalleumetala
290295300
metvallysarglysaspserarggly
305310
<210>9
<211>822
<212>dna
<213>人工序列(artificialsequence)
<400>9
atggcatgccctggcttcctgtgggcacttgtgatctccacctgtcttgaatttagcatg60
gctcagacagtcactcagtctcaaccagagatgtctgtgcaggaggcagagaccgtgacc120
ctgagctgcacatatgacaccagtgagagtgattattatttattctggtacaagcagcct180
cccagcaggcagatgattctcgttattcgccaagaagcttataagcaacagaatgcaaca240
gagaatcgtttctctgtgaacttccagaaagcagccaaatccttcagtctcaagatctca300
gactcacagctgggggatgccgcgatgtatttctgtgcttataggctctttaataatgca360
ggcaacatgctcacctttggagggggaacaaggttaatggtcaaaccccatatccagaac420
ccagaacctgctgtgtaccagttaaaagatcctcggtctcaggacagcaccctctgcctg480
ttcaccgactttgactcccaaatcaatgtgccgaaaaccatggaatctggaacgttcatc540
actgacaaaactgtgctggacatgaaagctatggattccaagagcaatggggccattgcc600
tggagcaaccagacaagcttcacctgccaagatatcttcaaagagaccaacgccacctac660
cccagttcagacgttccctgtgatgccacgttgaccgagaaaagctttgaaacagatatg720
aacctaaactttcaaaacctgtcagttatgggactccgaatcctcctgctgaaagtagcg780
ggatttaacctgctcatgacgctgaggctgtggtccagttga822
<210>10
<211>273
<212>prt
<213>人工序列(artificialsequence)
<400>10
metalacysproglypheleutrpalaleuvalileserthrcysleu
151015
gluphesermetalaglnthrvalthrglnserglnproglumetser
202530
valglnglualagluthrvalthrleusercysthrtyraspthrser
354045
gluserasptyrtyrleuphetrptyrlysglnproproserarggln
505560
metileleuvalileargglnglualatyrlysglnglnasnalathr
65707580
gluasnargpheservalasnpheglnlysalaalalysserpheser
859095
leulysileseraspserglnleuglyaspalaalamettyrphecys
100105110
alatyrargleupheasnasnalaglyasnmetleuthrpheglygly
115120125
glythrargleumetvallysprohisileglnasnprogluproala
130135140
valtyrglnleulysaspproargserglnaspserthrleucysleu
145150155160
phethrasppheaspserglnileasnvalprolysthrmetgluser
165170175
glythrpheilethrasplysthrvalleuaspmetlysalametasp
180185190
serlysserasnglyalailealatrpserasnglnthrserphethr
195200205
cysglnaspilephelysgluthrasnalathrtyrproserserasp
210215220
valprocysaspalathrleuthrglulysserphegluthraspmet
225230235240
asnleuasnpheglnasnleuservalmetglyleuargileleuleu
245250255
leulysvalalaglypheasnleuleumetthrleuargleutrpser
260265270
ser
<210>11
<211>921
<212>dna
<213>人工序列(artificialsequence)
<400>11
atggatacctggctcgtatgctgggcaatttttagtctcttgaaagcaggactcacagaa60
cctgaagtcacccagactcccagccatcaggtcacacagatgggacaggaagtgatcttg120
cgctgtgtccccatctctaatcacttatacttctattggtacagacaaatcttggggcag180
aaagtcgagtttctggtttccttttataataatgaaatctcagagaagtctgaaatattc240
gatgatcaattctcagttgaaaggcctgatggatcaaatttcactctgaagatccggtcc300
acaaagctggaggactcagccatgtacttctgtgccagcagtaccgatttggtaggggtc360
gaagaagctttctttggacaaggcaccagactcacagttgtagaggatctgagaaatgtg420
actccacccaaggtctccttgtttgagccatcaaaagcagagattgcaaacaaacaaaag480
gctaccctcgtgtgcttggccaggggcttcttccctgaccacgtggagctgagctggtgg540
gtgaatggcaaggaggtccacagtggggtcagcacggaccctcaggcctacaaggagagc600
aattatagctactgcctgagcagccgcctgagggtctctgctaccttctggcacaatcct660
cgcaaccacttccgctgccaagtgcagttccatgggctttcagaggaggacaagtggcca720
gagggctcacccaaacctgtcacacagaacatcagtgcagaggcctggggccgagcagac780
tgtgggattacctcagcatcctatcaacaaggggtcttgtctgccaccatcctctatgag840
atcctgctagggaaagccaccctgtatgctgtgcttgtcagtacactggtggtgatggct900
atggtcaaaagaaagaattca921
<210>12
<211>307
<212>prt
<213>人工序列(artificialsequence)
<400>12
metaspthrtrpleuvalcystrpalailepheserleuleulysala
151015
glyleuthrgluprogluvalthrglnthrproserhisglnvalthr
202530
glnmetglyglngluvalileleuargcysvalproileserasnhis
354045
leutyrphetyrtrptyrargglnileleuglyglnlysvalgluphe
505560
leuvalserphetyrasnasngluileserglulyssergluilephe
65707580
aspaspglnpheservalgluargproaspglyserasnphethrleu
859095
lysileargserthrlysleugluaspseralamettyrphecysala
100105110
serserthraspleuvalglyvalgluglualaphepheglyglngly
115120125
thrargleuthrvalvalgluaspleuargasnvalthrproprolys
130135140
valserleuphegluproserlysalagluilealaasnlysglnlys
145150155160
alathrleuvalcysleualaargglyphepheproasphisvalglu
165170175
leusertrptrpvalasnglylysgluvalhisserglyvalserthr
180185190
aspproglnalatyrlysgluserasntyrsertyrcysleuserser
195200205
argleuargvalseralathrphetrphisasnproargasnhisphe
210215220
argcysglnvalglnphehisglyleuserglugluasplystrppro
225230235240
gluglyserprolysprovalthrglnasnileseralaglualatrp
245250255
glyargalaaspcysglyilethrseralasertyrglnglnglyval
260265270
leuseralathrileleutyrgluileleuleuglylysalathrleu
275280285
tyralavalleuvalserthrleuvalvalmetalametvallysarg
290295300
lysasnser
305
<210>13
<211>1866
<212>dna
<213>人工序列(artificialsequence)
<400>13
atggatacctggctcgtatgctgggcaatttttagtctcttgaaagcaggactcacagaa60
cctgaagtcacccagactcccagccatcaggtcacacagatgggacaggaagtgatcttg120
cgctgtgtccccatctctaatcacttatacttctattggtacagacaaatcttggggcag180
aaagtcgagtttctggtttccttttataataatgaaatctcagagaagtctgaaatattc240
gatgatcaattctcagttgaaaggcctgatggatcaaatttcactctgaagatccggtcc300
acaaagctggaggactcagccatgtacttctgtgccagcagtaccgatttggtaggggtc360
gaagaagctttctttggacaaggcaccagactcacagttgtagaggacctgaacaaggtg420
ttcccacccgaggtcgctgtgtttgagccatcagaagcagagatctcccacacccaaaag480
gccacactggtatgcctggccacaggcttctaccccgaccacgtggagctgagctggtgg540
gtgaatgggaaggaggtgcacagtggggtctgcacagacccgcagcccctcaaggagcag600
cccgccctcaatgactccagatactgcctgagcagccgcctgagggtctcggccaccttc660
tggcagaacccccgcaaccacttccgctgtcaagtccagttctacgggctctcggagaat720
gacgagtggacccaggatagggccaaacccgtcacccagatcgtcagcgccgaggcctgg780
ggtagagcagactgtggcttcacctccgagtcttaccagcaaggggtcctgtctgccacc840
atcctctatgagatcttgctagggaaggccaccttgtatgccgtgctggtcagtgccctc900
gtgctgatggccatggtcaagagaaaggattccagaggccgtgccaagcgatccggaagc960
ggagcccctgtaaagcagactttgaattttgaccttctcaagttggcgggagacgtcgag1020
tccaaccctgggcccatggcatgccctggcttcctgtgggcacttgtgatctccacctgt1080
cttgaatttagcatggctcagacagtcactcagtctcaaccagagatgtctgtgcaggag1140
gcagagaccgtgaccctgagctgcacatatgacaccagtgagagtgattattatttattc1200
tggtacaagcagcctcccagcaggcagatgattctcgttattcgccaagaagcttataag1260
caacagaatgcaacagagaatcgtttctctgtgaacttccagaaagcagccaaatccttc1320
agtctcaagatctcagactcacagctgggggatgccgcgatgtatttctgtgcttatagg1380
ctctttaataatgcaggcaacatgctcacctttggagggggaacaaggttaatggtcaaa1440
ccccatatccagaaccctgaccctgccgtgtaccagctgagagactctaaatccagtgac1500
aagtctgtctgcctattcaccgattttgattctcaaacaaatgtgtcacaaagtaaggat1560
tctgatgtgtatatcacagacaaatgcgtgctagacatgaggtctatggacttcaagagc1620
aacagtgctgtggcctggagcaacaaatctgactttgcatgtgcaaacgccttcaacaac1680
agcattattccagaagacaccttcttccccagcccagaaagttcctgtgatgtcaagctg1740
gtcgagaaaagctttgaaacagatacgaacctaaactttcaaaacctgtcagtgattggg1800
ttccgaatcctcctcctgaaagtggccgggtttaatctgctcatgacgctgcggctgtgg1860
tccagc1866
<210>14
<211>622
<212>prt
<213>人工序列(artificialsequence)
<400>14
metaspthrtrpleuvalcystrpalailepheserleuleulysala
151015
glyleuthrgluprogluvalthrglnthrproserhisglnvalthr
202530
glnmetglyglngluvalileleuargcysvalproileserasnhis
354045
leutyrphetyrtrptyrargglnileleuglyglnlysvalgluphe
505560
leuvalserphetyrasnasngluileserglulyssergluilephe
65707580
aspaspglnpheservalgluargproaspglyserasnphethrleu
859095
lysileargserthrlysleugluaspseralamettyrphecysala
100105110
serserthraspleuvalglyvalgluglualaphepheglyglngly
115120125
thrargleuthrvalvalgluaspleuasnlysvalpheproproglu
130135140
valalavalphegluproserglualagluileserhisthrglnlys
145150155160
alathrleuvalcysleualathrglyphetyrproasphisvalglu
165170175
leusertrptrpvalasnglylysgluvalhisserglyvalcysthr
180185190
aspproglnproleulysgluglnproalaleuasnaspserargtyr
195200205
cysleuserserargleuargvalseralathrphetrpglnasnpro
210215220
argasnhispheargcysglnvalglnphetyrglyleusergluasn
225230235240
aspglutrpthrglnaspargalalysprovalthrglnilevalser
245250255
alaglualatrpglyargalaaspcysglyphethrserglusertyr
260265270
glnglnglyvalleuseralathrileleutyrgluileleuleugly
275280285
lysalathrleutyralavalleuvalseralaleuvalleumetala
290295300
metvallysarglysaspserargglyargalalysargserglyser
305310315320
glyalaprovallysglnthrleuasnpheaspleuleulysleuala
325330335
glyaspvalgluserasnproglyprometalacysproglypheleu
340345350
trpalaleuvalileserthrcysleugluphesermetalaglnthr
355360365
valthrglnserglnproglumetservalglnglualagluthrval
370375380
thrleusercysthrtyraspthrsergluserasptyrtyrleuphe
385390395400
trptyrlysglnproproserargglnmetileleuvalilearggln
405410415
glualatyrlysglnglnasnalathrgluasnargpheservalasn
420425430
pheglnlysalaalalysserpheserleulysileseraspsergln
435440445
leuglyaspalaalamettyrphecysalatyrargleupheasnasn
450455460
alaglyasnmetleuthrpheglyglyglythrargleumetvallys
465470475480
prohisileglnasnproaspproalavaltyrglnleuargaspser
485490495
lysserserasplysservalcysleuphethrasppheaspsergln
500505510
thrasnvalserglnserlysaspseraspvaltyrilethrasplys
515520525
cysvalleuaspmetargsermetaspphelysserasnseralaval
530535540
alatrpserasnlysseraspphealacysalaasnalapheasnasn
545550555560
serileileprogluaspthrphepheproserproglusersercys
565570575
aspvallysleuvalglulysserphegluthraspthrasnleuasn
580585590
pheglnasnleuservalileglypheargileleuleuleulysval
595600605
alaglypheasnleuleumetthrleuargleutrpserser
610615620
<210>15
<211>1836
<212>dna
<213>人工序列(artificialsequence)
<400>15
atggatacctggctcgtatgctgggcaatttttagtctcttgaaagcaggactcacagaa60
cctgaagtcacccagactcccagccatcaggtcacacagatgggacaggaagtgatcttg120
cgctgtgtccccatctctaatcacttatacttctattggtacagacaaatcttggggcag180
aaagtcgagtttctggtttccttttataataatgaaatctcagagaagtctgaaatattc240
gatgatcaattctcagttgaaaggcctgatggatcaaatttcactctgaagatccggtcc300
acaaagctggaggactcagccatgtacttctgtgccagcagtaccgatttggtaggggtc360
gaagaagctttctttggacaaggcaccagactcacagttgtagaggatctgagaaatgtg420
actccacccaaggtctccttgtttgagccatcaaaagcagagattgcaaacaaacaaaag480
gctaccctcgtgtgcttggccaggggcttcttccctgaccacgtggagctgagctggtgg540
gtgaatggcaaggaggtccacagtggggtcagcacggaccctcaggcctacaaggagagc600
aattatagctactgcctgagcagccgcctgagggtctctgctaccttctggcacaatcct660
cgcaaccacttccgctgccaagtgcagttccatgggctttcagaggaggacaagtggcca720
gagggctcacccaaacctgtcacacagaacatcagtgcagaggcctggggccgagcagac780
tgtgggattacctcagcatcctatcaacaaggggtcttgtctgccaccatcctctatgag840
atcctgctagggaaagccaccctgtatgctgtgcttgtcagtacactggtggtgatggct900
atggtcaaaagaaagaattcacgtgccaagcgatccggaagcggagcccctgtaaagcag960
actttgaattttgaccttctcaagttggcgggagacgtcgagtccaaccctgggcccatg1020
gcatgccctggcttcctgtgggcacttgtgatctccacctgtcttgaatttagcatggct1080
cagacagtcactcagtctcaaccagagatgtctgtgcaggaggcagagaccgtgaccctg1140
agctgcacatatgacaccagtgagagtgattattatttattctggtacaagcagcctccc1200
agcaggcagatgattctcgttattcgccaagaagcttataagcaacagaatgcaacagag1260
aatcgtttctctgtgaacttccagaaagcagccaaatccttcagtctcaagatctcagac1320
tcacagctgggggatgccgcgatgtatttctgtgcttataggctctttaataatgcaggc1380
aacatgctcacctttggagggggaacaaggttaatggtcaaaccccatatccagaaccca1440
gaacctgctgtgtaccagttaaaagatcctcggtctcaggacagcaccctctgcctgttc1500
accgactttgactcccaaatcaatgtgccgaaaaccatggaatctggaacgttcatcact1560
gacaaaactgtgctggacatgaaagctatggattccaagagcaatggggccattgcctgg1620
agcaaccagacaagcttcacctgccaagatatcttcaaagagaccaacgccacctacccc1680
agttcagacgttccctgtgatgccacgttgaccgagaaaagctttgaaacagatatgaac1740
ctaaactttcaaaacctgtcagttatgggactccgaatcctcctgctgaaagtagcggga1800
tttaacctgctcatgacgctgaggctgtggtccagt1836
<210>16
<211>612
<212>prt
<213>人工序列(artificialsequence)
<400>16
metaspthrtrpleuvalcystrpalailepheserleuleulysala
151015
glyleuthrgluprogluvalthrglnthrproserhisglnvalthr
202530
glnmetglyglngluvalileleuargcysvalproileserasnhis
354045
leutyrphetyrtrptyrargglnileleuglyglnlysvalgluphe
505560
leuvalserphetyrasnasngluileserglulyssergluilephe
65707580
aspaspglnpheservalgluargproaspglyserasnphethrleu
859095
lysileargserthrlysleugluaspseralamettyrphecysala
100105110
serserthraspleuvalglyvalgluglualaphepheglyglngly
115120125
thrargleuthrvalvalgluaspleuargasnvalthrproprolys
130135140
valserleuphegluproserlysalagluilealaasnlysglnlys
145150155160
alathrleuvalcysleualaargglyphepheproasphisvalglu
165170175
leusertrptrpvalasnglylysgluvalhisserglyvalserthr
180185190
aspproglnalatyrlysgluserasntyrsertyrcysleuserser
195200205
argleuargvalseralathrphetrphisasnproargasnhisphe
210215220
argcysglnvalglnphehisglyleuserglugluasplystrppro
225230235240
gluglyserprolysprovalthrglnasnileseralaglualatrp
245250255
glyargalaaspcysglyilethrseralasertyrglnglnglyval
260265270
leuseralathrileleutyrgluileleuleuglylysalathrleu
275280285
tyralavalleuvalserthrleuvalvalmetalametvallysarg
290295300
lysasnserargalalysargserglyserglyalaprovallysgln
305310315320
thrleuasnpheaspleuleulysleualaglyaspvalgluserasn
325330335
proglyprometalacysproglypheleutrpalaleuvalileser
340345350
thrcysleugluphesermetalaglnthrvalthrglnserglnpro
355360365
glumetservalglnglualagluthrvalthrleusercysthrtyr
370375380
aspthrsergluserasptyrtyrleuphetrptyrlysglnpropro
385390395400
serargglnmetileleuvalileargglnglualatyrlysglngln
405410415
asnalathrgluasnargpheservalasnpheglnlysalaalalys
420425430
serpheserleulysileseraspserglnleuglyaspalaalamet
435440445
tyrphecysalatyrargleupheasnasnalaglyasnmetleuthr
450455460
pheglyglyglythrargleumetvallysprohisileglnasnpro
465470475480
gluproalavaltyrglnleulysaspproargserglnaspserthr
485490495
leucysleuphethrasppheaspserglnileasnvalprolysthr
500505510
metgluserglythrpheilethrasplysthrvalleuaspmetlys
515520525
alametaspserlysserasnglyalailealatrpserasnglnthr
530535540
serphethrcysglnaspilephelysgluthrasnalathrtyrpro
545550555560
serseraspvalprocysaspalathrleuthrglulysserpheglu
565570575
thraspmetasnleuasnpheglnasnleuservalmetglyleuarg
580585590
ileleuleuleulysvalalaglypheasnleuleumetthrleuarg
595600605
leutrpserser
610
<210>17
<211>1254
<212>prt
<213>人(homosapiens)
<400>17
metgluleualaalaleucysargtrpglyleuleuleualaleuleu
151015
proproglyalaalaserthrglnvalcysthrglythraspmetlys
202530
leuargleuproalaserprogluthrhisleuaspmetleuarghis
354045
leutyrglnglycysglnvalvalglnglyasnleugluleuthrtyr
505560
leuprothrasnalaserleuserpheleuglnaspileglngluval
65707580
glnglytyrvalleuilealahisasnglnvalargglnvalproleu
859095
glnargleuargilevalargglythrglnleuphegluaspasntyr
100105110
alaleualavalleuaspasnglyaspproleuasnasnthrthrpro
115120125
valthrglyalaserproglyglyleuarggluleuglnleuargser
130135140
leuthrgluileleulysglyglyvalleuileglnargasnprogln
145150155160
leucystyrglnaspthrileleutrplysaspilephehislysasn
165170175
asnglnleualaleuthrleuileaspthrasnargserargalacys
180185190
hisprocysserprometcyslysglyserargcystrpglygluser
195200205
sergluaspcysglnserleuthrargthrvalcysalaglyglycys
210215220
alaargcyslysglyproleuprothraspcyscyshisgluglncys
225230235240
alaalaglycysthrglyprolyshisseraspcysleualacysleu
245250255
hispheasnhisserglyilecysgluleuhiscysproalaleuval
260265270
thrtyrasnthraspthrpheglusermetproasnprogluglyarg
275280285
tyrthrpheglyalasercysvalthralacysprotyrasntyrleu
290295300
serthraspvalglysercysthrleuvalcysproleuhisasngln
305310315320
gluvalthralagluaspglythrglnargcysglulyscysserlys
325330335
procysalaargvalcystyrglyleuglymetgluhisleuargglu
340345350
valargalavalthrseralaasnileglngluphealaglycyslys
355360365
lysilepheglyserleualapheleuprogluserpheaspglyasp
370375380
proalaserasnthralaproleuglnprogluglnleuglnvalphe
385390395400
gluthrleuglugluilethrglytyrleutyrileseralatrppro
405410415
aspserleuproaspleuservalpheglnasnleuglnvalilearg
420425430
glyargileleuhisasnglyalatyrserleuthrleuglnglyleu
435440445
glyilesertrpleuglyleuargserleuarggluleuglysergly
450455460
leualaleuilehishisasnthrhisleucysphevalthrvalpro
465470475480
trpaspglnleupheargasnprohisglnalaleuleuhisthrala
485490495
asnargprogluaspglucysvalglygluglyleualacyshisgln
500505510
leucysalaargglyhiscystrpglyproglyprothrglncysval
515520525
asncysserglnpheleuargglyglnglucysvalgluglucysarg
530535540
valleuglnglyleuproargglutyrvalasnalaarghiscysleu
545550555560
procyshisproglucysglnproglnasnglyservalthrcysphe
565570575
glyproglualaaspglncysvalalacysalahistyrlysasppro
580585590
prophecysvalalaargcysproserglyvallysproaspleuser
595600605
tyrmetproiletrplyspheproaspglugluglyalacysglnpro
610615620
cysproileasncysthrhissercysvalaspleuaspasplysgly
625630635640
cysproalagluglnargalaserproleuthrserileileserala
645650655
valvalglyileleuleuvalvalvalleuglyvalvalpheglyile
660665670
leuilelysargargglnglnlysilearglystyrthrmetargarg
675680685
leuleuglngluthrgluleuvalgluproleuthrproserglyala
690695700
metproasnglnalaglnmetargileleulysgluthrgluleuarg
705710715720
lysvallysvalleuglyserglyalapheglythrvaltyrlysgly
725730735
iletrpileproaspglygluasnvallysileprovalalailelys
740745750
valleuarggluasnthrserprolysalaasnlysgluileleuasp
755760765
glualatyrvalmetalaglyvalglyserprotyrvalserargleu
770775780
leuglyilecysleuthrserthrvalglnleuvalthrglnleumet
785790795800
protyrglycysleuleuasphisvalarggluasnargglyargleu
805810815
glyserglnaspleuleuasntrpcysmetglnilealalysglymet
820825830
sertyrleugluaspvalargleuvalhisargaspleualaalaarg
835840845
asnvalleuvallysserproasnhisvallysilethraspphegly
850855860
leualaargleuleuaspileaspgluthrglutyrhisalaaspgly
865870875880
glylysvalproilelystrpmetalaleugluserileleuargarg
885890895
argphethrhisglnseraspvaltrpsertyrglyvalthrvaltrp
900905910
gluleumetthrpheglyalalysprotyraspglyileproalaarg
915920925
gluileproaspleuleuglulysglygluargleuproglnpropro
930935940
ilecysthrileaspvaltyrmetilemetvallyscystrpmetile
945950955960
aspserglucysargproargphearggluleuvalserglupheser
965970975
argmetalaargaspproglnargphevalvalileglnasngluasp
980985990
leuglyproalaserproleuaspserthrphetyrargserleuleu
99510001005
gluaspaspaspmetglyaspleuvalaspalagluglutyrleu
101010151020
valproglnglnglyphephecysproaspproalaproglyala
102510301035
glyglymetvalhishisarghisargserserserthrargser
104010451050
glyglyglyaspleuthrleuglyleugluproserglugluglu
105510601065
alaproargserproleualaprosergluglyalaglyserasp
107010751080
valpheaspglyaspleuglymetglyalaalalysglyleugln
108510901095
serleuprothrhisaspproserproleuglnargtyrserglu
110011051110
aspprothrvalproleuprosergluthraspglytyrvalala
111511201125
proleuthrcysserproglnproglutyrvalasnglnproasp
113011351140
valargproglnproproserproarggluglyproleuproala
114511501155
alaargproalaglyalathrleugluargprolysthrleuser
116011651170
proglylysasnglyvalvallysaspvalphealapheglygly
117511801185
alavalgluasnproglutyrleuthrproglnglyglyalaala
119011951200
proglnprohisproproproalapheserproalapheaspasn
120512101215
leutyrtyrtrpaspglnaspproprogluargglyalapropro
122012251230
serthrphelysglythrprothralagluasnproglutyrleu
123512401245
glyleuaspvalproval
1250
<210>18
<211>9
<212>prt
<213>人(homosapiens)
<400>18
lysilepheglyserleualapheleu
15
<210>19
<211>137
<212>prt
<213>人(homosapiens)
<400>19
metalacysproglypheleutrpalaleuvalileserthrcysleu
151015
gluphesermetalaglnthrvalthrglnserglnproglumetser
202530
valglnglualagluthrvalthrleusercysthrtyraspthrser
354045
gluserasptyrtyrleuphetrptyrlysglnproproserarggln
505560
metileleuvalileargglnglualatyrlysglnglnasnalathr
65707580
gluasnargpheservalasnpheglnlysalaalalysserpheser
859095
leulysileseraspserglnleuglyaspalaalamettyrphecys
100105110
alatyrargleupheasnasnalaglyasnmetleuthrpheglygly
115120125
glythrargleumetvallysprohis
130135
<210>20
<211>134
<212>prt
<213>人(homosapiens)
<400>20
metaspthrtrpleuvalcystrpalailepheserleuleulysala
151015
glyleuthrgluprogluvalthrglnthrproserhisglnvalthr
202530
glnmetglyglngluvalileleuargcysvalproileserasnhis
354045
leutyrphetyrtrptyrargglnileleuglyglnlysvalgluphe
505560
leuvalserphetyrasnasngluileserglulyssergluilephe
65707580
aspaspglnpheservalgluargproaspglyserasnphethrleu
859095
lysileargserthrlysleugluaspseralamettyrphecysala
100105110
serserthraspleuvalglyvalgluglualaphepheglyglngly
115120125
thrargleuthrvalval
130
<210>21
<211>411
<212>dna
<213>人(homosapiens)
<400>21
atggcatgccctggcttcctgtgggcacttgtgatctccacctgtcttgaatttagcatg60
gctcagacagtcactcagtctcaaccagagatgtctgtgcaggaggcagagaccgtgacc120
ctgagctgcacatatgacaccagtgagagtgattattatttattctggtacaagcagcct180
cccagcaggcagatgattctcgttattcgccaagaagcttataagcaacagaatgcaaca240
gagaatcgtttctctgtgaacttccagaaagcagccaaatccttcagtctcaagatctca300
gactcacagctgggggatgccgcgatgtatttctgtgcttataggctctttaataatgca360
ggcaacatgctcacctttggagggggaacaaggttaatggtcaaaccccat411
<210>22
<211>402
<212>dna
<213>人(homosapiens)
<400>22
atggatacctggctcgtatgctgggcaatttttagtctcttgaaagcaggactcacagaa60
cctgaagtcacccagactcccagccatcaggtcacacagatgggacaggaagtgatcttg120
cgctgtgtccccatctctaatcacttatacttctattggtacagacaaatcttggggcag180
aaagtcgagtttctggtttccttttataataatgaaatctcagagaagtctgaaatattc240
gatgatcaattctcagttgaaaggcctgatggatcaaatttcactctgaagatccggtcc300
acaaagctggaggactcagccatgtacttctgtgccagcagtaccgatttggtaggggtc360
gaagaagctttctttggacaaggcaccagactcacagttgta402
<210>23
<211>1866
<212>dna
<213>人工序列(artificialsequence)
<400>23
atggatacctggctcgtatgctgggcaatttttagtctcttgaaagcaggactcacagaa60
cctgaagtcacccagactcccagccatcaggtcacacagatgggacaggaagtgatcttg120
cgctgtgtccccatctctaatcacttatacttctattggtacagacaaatcttggggcag180
aaagtcgagtttctggtttccttttataataatgaaatctcagagaagtctgaaatattc240
gatgatcaattctcagttgaaaggcctgatggatcaaatttcactctgaagatccggtcc300
acaaagctggaggactcagccatgtacttctgtgccagcagtaccgatttggtaggggtc360
gaagaagctttctttggacaaggcaccagactcacagttgtagaggacctgaacaaggtg420
ttcccacccgaggtcgctgtgtttgagccatcagaagcagagatctcccacacccaaaag480
gccacactggtatgcctggccacaggcttctaccccgaccacgtggagctgagctggtgg540
gtgaatgggaaggaggtgcacagtggggtcagcacagacccgcagcccctcaaggagcag600
cccgccctcaatgactccagatactgcctgagcagccgcctgagggtctcggccaccttc660
tggcagaacccccgcaaccacttccgctgtcaagtccagttctacgggctctcggagaat720
gacgagtggacccaggatagggccaaacccgtcacccagatcgtcagcgccgaggcctgg780
ggtagagcagactgtggcttcacctccgagtcttaccagcaaggggtcctgtctgccacc840
atcctctatgagatcttgctagggaaggccaccttgtatgccgtgctggtcagtgccctc900
gtgctgatggccatggtcaagagaaaggattccagaggccgtgccaagcgatccggaagc960
ggagcccctgtaaagcagactttgaattttgaccttctcaagttggcgggagacgtcgag1020
tccaaccctgggcccatggcatgccctggcttcctgtgggcacttgtgatctccacctgt1080
cttgaatttagcatggctcagacagtcactcagtctcaaccagagatgtctgtgcaggag1140
gcagagaccgtgaccctgagctgcacatatgacaccagtgagagtgattattatttattc1200
tggtacaagcagcctcccagcaggcagatgattctcgttattcgccaagaagcttataag1260
caacagaatgcaacagagaatcgtttctctgtgaacttccagaaagcagccaaatccttc1320
agtctcaagatctcagactcacagctgggggatgccgcgatgtatttctgtgcttatagg1380
ctctttaataatgcaggcaacatgctcacctttggagggggaacaaggttaatggtcaaa1440
ccccatatccagaaccctgaccctgccgtgtaccagctgagagactctaaatccagtgac1500
aagtctgtctgcctattcaccgattttgattctcaaacaaatgtgtcacaaagtaaggat1560
tctgatgtgtatatcacagacaaaaccgtgctagacatgaggtctatggacttcaagagc1620
aacagtgctgtggcctggagcaacaaatctgactttgcatgtgcaaacgccttcaacaac1680
agcattattccagaagacaccttcttccccagcccagaaagttcctgtgatgtcaagctg1740
gtcgagaaaagctttgaaacagatacgaacctaaactttcaaaacctgtcagtgattggg1800
ttccgaatcctcctcctgaaagtggccgggtttaatctgctcatgacgctgcggctgtgg1860
tccagc1866
<210>24
<211>622
<212>prt
<213>人工序列(artificialsequence)
<400>24
metaspthrtrpleuvalcystrpalailepheserleuleulysala
151015
glyleuthrgluprogluvalthrglnthrproserhisglnvalthr
202530
glnmetglyglngluvalileleuargcysvalproileserasnhis
354045
leutyrphetyrtrptyrargglnileleuglyglnlysvalgluphe
505560
leuvalserphetyrasnasngluileserglulyssergluilephe
65707580
aspaspglnpheservalgluargproaspglyserasnphethrleu
859095
lysileargserthrlysleugluaspseralamettyrphecysala
100105110
serserthraspleuvalglyvalgluglualaphepheglyglngly
115120125
thrargleuthrvalvalgluaspleuasnlysvalpheproproglu
130135140
valalavalphegluproserglualagluileserhisthrglnlys
145150155160
alathrleuvalcysleualathrglyphetyrproasphisvalglu
165170175
leusertrptrpvalasnglylysgluvalhisserglyvalserthr
180185190
aspproglnproleulysgluglnproalaleuasnaspserargtyr
195200205
cysleuserserargleuargvalseralathrphetrpglnasnpro
210215220
argasnhispheargcysglnvalglnphetyrglyleusergluasn
225230235240
aspglutrpthrglnaspargalalysprovalthrglnilevalser
245250255
alaglualatrpglyargalaaspcysglyphethrserglusertyr
260265270
glnglnglyvalleuseralathrileleutyrgluileleuleugly
275280285
lysalathrleutyralavalleuvalseralaleuvalleumetala
290295300
metvallysarglysaspserargglyargalalysargserglyser
305310315320
glyalaprovallysglnthrleuasnpheaspleuleulysleuala
325330335
glyaspvalgluserasnproglyprometalacysproglypheleu
340345350
trpalaleuvalileserthrcysleugluphesermetalaglnthr
355360365
valthrglnserglnproglumetservalglnglualagluthrval
370375380
thrleusercysthrtyraspthrsergluserasptyrtyrleuphe
385390395400
trptyrlysglnproproserargglnmetileleuvalilearggln
405410415
glualatyrlysglnglnasnalathrgluasnargpheservalasn
420425430
pheglnlysalaalalysserpheserleulysileseraspsergln
435440445
leuglyaspalaalamettyrphecysalatyrargleupheasnasn
450455460
alaglyasnmetleuthrpheglyglyglythrargleumetvallys
465470475480
prohisileglnasnproaspproalavaltyrglnleuargaspser
485490495
lysserserasplysservalcysleuphethrasppheaspsergln
500505510
thrasnvalserglnserlysaspseraspvaltyrilethrasplys
515520525
thrvalleuaspmetargsermetaspphelysserasnseralaval
530535540
alatrpserasnlysseraspphealacysalaasnalapheasnasn
545550555560
serileileprogluaspthrphepheproserproglusersercys
565570575
aspvallysleuvalglulysserphegluthraspthrasnleuasn
580585590
pheglnasnleuservalileglypheargileleuleuleulysval
595600605
alaglypheasnleuleumetthrleuargleutrpserser
610615620
<210>25
<211>10
<212>prt
<213>人(homosapiens)
<400>25
serleualapheleuprogluserpheasp
1510
<210>26
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>26
gcctctggaatcctttctcttg22
<210>27
<211>21
<212>dna
<213>人工序列(artificialsequence)
<400>27
tcagctggaccacagccgcag21
<210>28
<211>38
<212>dna
<213>人工序列(artificialsequence)
<400>28
agagctagcgaattcaacatggatacctggctcgtatg38
<210>29
<211>38
<212>dna
<213>人工序列(artificialsequence)
<400>29
gttgattgtcgacgccctcagctggaccacagccgcag38
<210>30
<211>38
<212>dna
<213>人工序列(artificialsequence)
<400>30
agagctagcgaattcaacatggatacctggctcgtatg38
<210>31
<211>35
<212>dna
<213>人工序列(artificialsequence)
<400>31
gttgattgtcgacgccctcaactggaccacagcct35
- 还没有人留言评论。精彩留言会获得点赞!