单分子肽测序的手段和方法与流程
发明领域
本发明涉及生物化学领域,更特别地涉及蛋白质组学,更特别地涉及蛋白质测序,甚至更特别地涉及单分子肽测序。本发明公开了使用裂解诱导剂进行单分子蛋白质测序和/或氨基酸鉴定的手段和方法。所述不是对一种特定氨基酸特异性的裂解诱导剂,从n末端开始逐步裂解多肽,并基于所述裂解诱导剂与多肽之间的啮合(engagement)动力学或所述多肽裂解反应的动力学来提供关于裂解的氨基酸的身份的信息。
背景
生物分子(dna、rna、蛋白质)的高通量测序技术正在迅速发展,并且对于现代科学和医学至关重要。尤其是dna测序技术已经取得了巨大飞跃,目前已从二代技术发展到作用于单分子水平的第三代方法(例如pacificbiosciences,oxfordnanopores;ambardar等,2016indianjmicrobiol56:394-404)。dna的均匀性质和广泛的可用分子工具组确实推动了该领域的发展。相比之下,蛋白质/肽的测序相对滞后,并且很大程度上依赖于基于液相色谱-质谱(lc-ms)的技术。尽管lc-ms仪器在速度、灵敏度和分辨率方面已取得了很大进步,但操作和维护机器却非常复杂且昂贵。此外,基于ms的蛋白质组学仍然需要大约106个拷贝的蛋白质/肽才能被检测到。目前无法进行单蛋白质/肽测序,因此无法实现如单细胞蛋白质组学这样的应用。此外,由于蛋白质无法像dna一样进行扩增,因此单分子测序从概念上讲是更好的选择,而且可以进一步对蛋白质进行数字量化。
下一代蛋白质测序的概念正在萌芽,但面临重大挑战。首先,要区分二十种氨基酸(相比之下,只有四种核苷酸),并且获得单个可测量参数以严格识别每种氨基酸似乎不太可能。然而,甚至通过概率方式能够将某个氨基酸类别归因于每个序列位置,对于通过基于约束的肽识别从数据库进行剖析(如针对lc-ms数据所做的那样),可能就已经足够了(swaminathan等。2015ploscomputbiol.11:e1004080)。其次,就理化性质而言,蛋白质极不均一。通过应用自下而上的蛋白质组学方法,即通过蛋白酶介导的蛋白质组消化成肽,可以在一定程度上减少这种情况。动态范围为几个数量级的这种复杂的肽混合物的并行测序将不可避免地面临巨大的技术障碍。通过从肽库中纯化(蛋白型)肽的子集,可以进一步降低复杂性。通过(固态)纳米孔的肽测序是目前研究中最受欢迎的平台。对纳米孔的工程化进行了广泛的研究,这些工程化能够转运肽并沿序列区分氨基酸或氨基酸类别(kennedy等2016natnanotechnol11:968-976;wilson等2016advfunctmater26:4830-4838)。
mitra等(wo2010/065531)开发了另一种有希望的以灵敏和定量的方式分析蛋白质的技术。这项技术被称为通过末端测序的蛋白质数字分析或dapes,其特征是一种用于单分子蛋白质分析的方法。为了进行dapes,将大量蛋白质变性并裂解成肽。将这些肽固定在应用于显微镜载玻片表面的纳米凝胶表面上,并使用涉及edman降解的方法并行测定其氨基酸序列。将异硫氰酸苯酯(pitc)添加到载玻片中,并与每种肽的n端氨基酸反应形成稳定的苯硫脲衍生物。接着,通过例如用每个pitc衍生的n末端氨基酸特异性的抗体进行20轮抗体结合、检测和剥离来确定n末端氨基酸衍生物的身份。通过升高温度或降低ph值来去除n末端氨基酸,并重复该循环以从载玻片上的每个肽中测序12-20个氨基酸。然后可以根据观察到的不同肽序列的数量来计算原始样品中每种蛋白质的绝对浓度。dapes中使用的pitc化学性质与edman降解中使用的相同,并且高效且耐用(效率>99%)。然而,单个氨基酸的裂解需要强的无水酸,或替换地,高温水性缓冲液。对于用于单分子蛋白质检测(smd)的敏感底物上的多轮分析,不希望在这些严酷条件中的任一个之间进行循环。
替代的肽测序方法使用n末端氨基酸结合蛋白(naab)代替结合pitc衍生的n末端氨基酸的抗体(wo20140273004)。对于每种氨基酸,研发了可以由氨肽酶或trna合成酶修饰的naab。naab被不同地标记,然后通过在孵育和洗涤此类naab时检测与n末端氨基酸结合的特定naab的荧光标记来鉴定多肽的n末端氨基酸。此外,代替化学/物理去除n末端氨基酸,可以使用称为edmanase的酶(以edman降解命名)。尽管edmanase解决了mitra等的局限性,但这种方法依赖于naab的库来推导氨基酸身份信息。所有不同氨基酸的naab应一起存在或顺序添加,从而增加了该系统的复杂性。此外,研发具有足够亲和力的以用于单分子感测中的naab的能力仍未被证实。因此,与仅基于试剂的结合亲和力相比,基于不同的理化原理研发更简单且更精细的蛋白质测序技术将是有利的。
概述
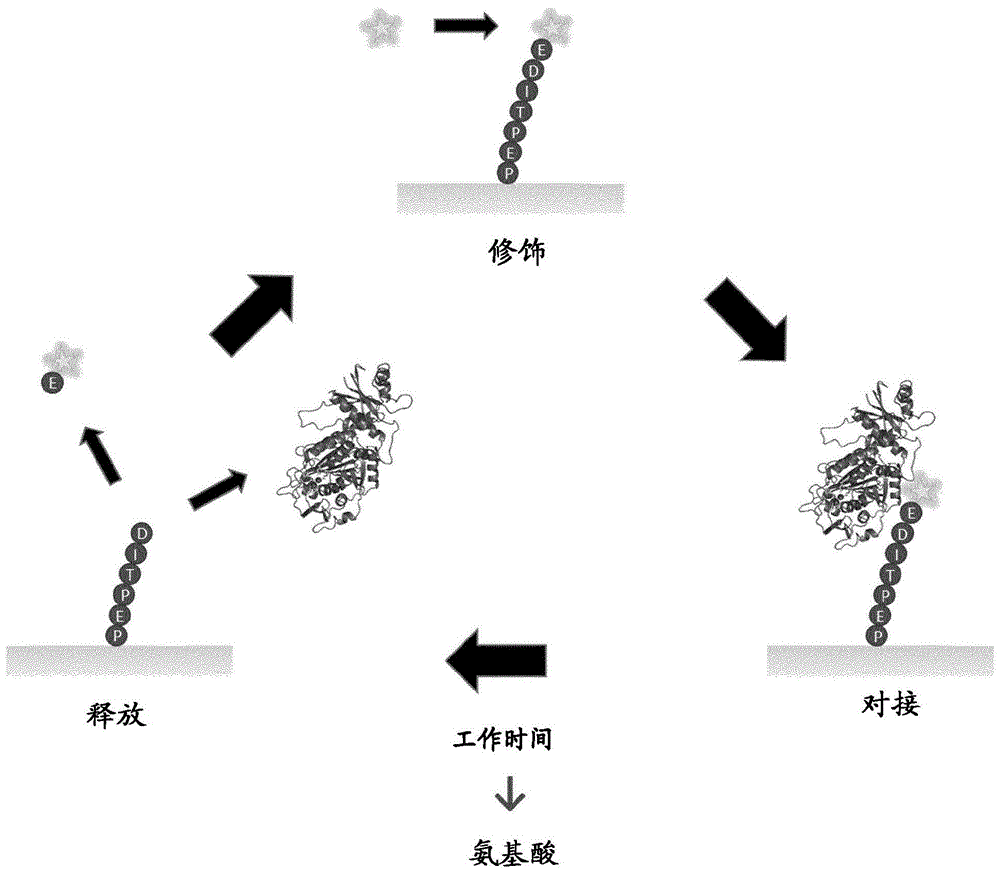
在此,描述了替代的单分子肽测序方法和其中涉及的工程化分子。使用氨肽酶的催化特性和酶促反应的动力学来鉴定(或分类)肽单分子的n末端氨基酸。本申请中描述的方法基于工程化的氨肽酶的周转数(kcat)与其裂解的n端氨基酸之间的相关性。因此,通过测量n-末端氨基酸被切除之前,工程化氨肽酶在添加到肽底物上后停留的时间,可以鉴定出n-末端氨基酸。所述氨肽酶也可以被化学裂解诱导剂代替。与使用氨肽酶所观察到的相似,化学裂解诱导剂的停留时间是对其结合的n端氨基酸身份的读出。更准确地说,本申请提供了一种对蛋白质进行测序的方法,该方法包括以下步骤循环:通过肽c-末端部分与易裂键固定的肽的n-末端衍生,测量裂解诱导剂切除n末端氨基酸导致n末端氨基酸从固定化表面释放所花费的时间,使系统为下一个循环做好准备(图1)。裂解诱导剂可以是催化活性的氨肽酶或异硫氰酸酯样化学物质。
在第一个方面中,提供了工程化的、作用于多肽的催化活性氨肽酶,其中所述多肽通过其c末端或通过所述多肽的第一肽键的c末端肽部分固定在表面上,其中所述氨肽酶裂解所述多肽的n-末端氨基酸,并且其中所述氨肽酶在裂解所述n-末端氨基酸之前的停留时间鉴定或分类所述n-末端氨基酸。所述n-末端氨基酸可以是衍生的n-末端氨基酸,并且如果这样,则所述氨肽酶结合并裂解所述衍生的n-末端氨基酸。所述n末端氨基酸可以是用异硫氰酸酯或异硫氰酸酯类似物衍生的n末端氨基酸。更特别地,上述氨肽酶是与seqidno.1或seqidno.2具有至少80%序列同一性并且具有25位的甘氨酸残基,65位的丝氨酸残基,138位的半胱氨酸残基和208位的组氨酸残基的氨肽酶,所述氨肽酶能够结合用citc或spitc衍生的n端氨基酸。更具体地讲,氨肽酶包含如seqidno.3或seqidno.4中所示的氨基酸序列。
本申请所述的氨肽酶还可以与seqidno.7具有至少80%序列同一性,其中半胱氨酸残基插入第1位的甲硫氨酸残基和第2位的丙氨酸残基之间。更特别地,所述氨肽酶包含seqidno.8或由seqidno.8组成。在特别的实施方案中,以上氨肽酶进一步包含光学、电学或等离子体标记,因此可以通过光学、电学或等离子体检测所述氨肽酶。在其他特别的实施方案中,所述氨肽酶是嗜热的和/或耐溶剂的。
在第二个方面中,提供了裂解诱导剂获得多肽的序列信息的用途,所述多肽通过其c-末端或通过所述多肽的第一肽键的c-末端肽部分固定在表面上,其中所述裂解诱导剂在所述多肽的n-末端氨基酸上的停留时间鉴定或分类所述n-末端氨基酸。所述裂解诱导剂可以是催化活性氨肽酶、异硫氰酸酯或异硫氰酸酯类似物。更特别地,所述催化活性氨肽酶可以是本申请中所述的任一种氨肽酶。在特别的实施方案中,所述n-末端氨基酸选自leu、met、tyr、arg、pro、gly、lys、ala和val。还提供了以上的用途,所述n-末端氨基酸是衍生的n-末端氨基酸并且用于获得单分子水平的序列信息。
在第三个方面中,提供了鉴定或分类多肽的n-末端氨基酸的方法,所述多肽通过其c-末端或通过所述多肽的第一肽键的c-末端肽部分固定在表面上,所述方法包括将所述表面固定的多肽接触裂解诱导剂,其中所述裂解诱导剂结合所述多肽的n-末端氨基酸并从所述多肽裂解n-末端氨基酸;测量所述裂解诱导剂在所述n-末端氨基酸上的停留时间;并将所述测量的停留时间与所述裂解诱导剂和一组n-末端氨基酸特征性的一组参照停留时间进行比较,以鉴定或分类所述n-末端氨基酸。
接着,提供了获得通过c-末端固定在表面上的多肽的序列信息的方法,所述方法包括:
a)将所述表面固定的多肽接触裂解诱导剂,其中所述裂解诱导剂结合所述多肽的n-末端氨基酸并从所述多肽裂解n-末端氨基酸;
b)测量所述裂解诱导剂在所述表面固定多肽的n-末端氨基酸上的停留时间;
c)使所述裂解诱导剂切除所述n-末端氨基酸;
d)通过将所述测量的停留时间与所述裂解诱导剂和一组n-末端氨基酸特征性的一组参照停留时间进行比较,以鉴定或分类所述n-末端氨基酸;
e)重复步骤a)至d)一次或多次或重复步骤b)至d)一次或多次。
还提供了以上方法,其中所述裂解诱导剂是异硫氰酸酯或异硫氰酸酯类似物,其中所述停留时间是直到除去所述n-末端氨基酸的时长,其中通过将所述时长与不同氨基酸的一组参照值进行比较来鉴定所述n-末端氨基酸。
还提供了以上方法,其中所述裂解诱导剂是氨肽酶,并且针对所述氨肽酶与所述n-末端氨基酸的每个结合事件来测量所述氨肽酶的停留时间。
本申请的方法可以另外包括通过测量表面固定多肽的光学、电学或等离子体信号来测定所述n-末端氨基酸裂解的步骤,其中光学、电学或等离子体信号的差异表示所述n-末端氨基酸的裂解。还提供了以上方法,其中将所述表面固定的肽另外接触一种或多种n-末端氨基酸结合蛋白,其中所述一种或多种结合蛋白与所述n-末端氨基酸的结合事件的动力学鉴定所述n-末端氨基酸或提供了所述n-末端氨基酸的更多信息。以上方法还包括第一步的多肽变性,或提供了以上方法,其中在所述方法的一个或多个步骤过程中存在变性条件,其中所述催化活性的氨肽酶是嗜热和/或耐溶剂氨肽酶和/或其中所述裂解诱导剂是异硫氰酸酯或异硫氰酸酯类似物。还提供了以上方法,其中n-末端氨基酸是衍生的。以上方法的氨肽酶可以是本文所述的任一种氨肽酶。
在特别的实施方案中,设想本申请的方法使用单分子水平。尤其是对于这些应用,可以预期通过光学、电学或等离子体测量裂解诱导剂的停留时间。将所述多肽固定在活性传感表面时,这可以高通量地进行。所述活性传感表面可以是金表面或酰胺-、羧基-、硫醇-或叠氮化物-官能化表面,所述多肽化学偶联在所述表面上。
附图简述
图1.酶降解的动力学检测,更准确地用于单分子肽测序的n-末端氨基酸裂解。
图2.酶停留时间的光和电位测量读出的非限制性实例的示意图。
图3.玻璃表面上cy5标记的测试肽(pepcy5)的固定。左,对照;中,1nm浓度的pepcy5;右:显示出成功的空间分布的放大图。
图4.表面固定肽(1nmpepcy5)的胰蛋白酶消化(1nm)。a,不存在钝化剂下的固定pepcy5上的胰蛋白酶反应。b,钝化剂dbco-peg8-酰胺(1μm)存在下的固定pepcy5上的胰蛋白酶处理。在639nm下检测信号(上图)并在561nm的下λem通道中评估背景(下图)。
图5.硫代异硫氰酸苯酯-ala-phe(a)和3-香豆基-异硫氰酸酯-ala-phe(b)在虚拟再造的edmanase上的成功计算对接。
图6.工程化的克氏锥虫(t.cruzi)cruzipain(a)和水生栖热菌(t.aquaticus)氨肽酶t(b)在大肠杆菌中的成功转化。
图7.纯化的水生栖热菌氨肽酶t的sds-page分析。a,原始可溶性级分;b,ni-nta纯化;c,热处理;d,ni-nta纯化+热处理。
图8.使用l-亮氨酸-对硝基苯胺的肽酶测试。e,酶;s,底物。
图9.pet24b(+)质粒的分子图谱。
图10.氨肽酶的酰化和脱酰步骤的示意图。对于肽,情况认为xh=rnh2(因此在方案中产生了肽键的存在)。肽的n-末端部分由红色部分来表示,而肽的c-末端部分由x来表示。
图11.使用不同的氨基酸对硝基苯胺底物的氨肽酶测试。将酶和底物(1.5mm)在pbs中在40℃或80℃下孵育2h,此后通过测量405nm处的吸光度来定量释放的对硝基苯胺。
图12.来自水生栖热菌的氨肽酶t的有机溶剂耐受性。a.使用l-亮氨酸-对硝基苯胺底物测量了氨肽酶t的活性。将酶和底物在含有不同浓度的有机溶剂乙腈(acn)、甲醇(meoh)和乙醇(etoh)的50mmtrishcl(ph8)中在40℃下孵育3h。b.在0%vs.50%meoh(在10mm磷酸氢二钾缓冲液(k2hpo4)中)中的氨肽酶t二级结构的圆二色性分析。c.缓冲液(50mmtrishcl,ph8)vs.去离子水(milliq)中不同浓度的meoh中的酶活性。
图13.氨肽酶t的位点特异性荧光标记。a.用等摩尔和1×、10×、100×和1000×摩尔超量的马来酰亚胺-dylight650标记n-末端半胱氨酸后氨肽酶t的sds-page分析。用荧光(dylight650标记)和考马斯(总蛋白)将氨肽酶t可视化。b.用马来酰亚胺-dylight650标记后用l-亮氨酸-对硝基苯胺检查氨肽酶t活性。
图14.监测酶-底物结合事件和底物裂解事件的组合。通过使用荧光标记的固定肽底物和游离氨肽酶,用tipf显微镜监测单分子酶停留时间。
图15.edman降解机理的示意图。埃德曼降解需要将异硫氰酸苯酯(pitc)偶联到蛋白质/肽的游离n末端上(碱性条件),然后作为乙内酰苯硫脲(pth)衍生物释放n末端氨基酸(酸性条件)。然后用色谱鉴定释放的pth-氨基酸。然后连续重复该程序,从而产生蛋白质/肽序列信息(来源:https://en.wikipedia.org/wiki/edman_degradation)
图16.不同的氨基酸对硝基苯胺底物的自发edman降解。a.将硫代异硫氰酸苯酯(spitc,15mm)和底物(1.5mm)在50%acn中的300mm三乙醇胺中在40℃下孵育30min,此后通过测量405nm处的吸光度来定量释放的对硝基苯胺。b.spitc-诱导的氨基酸对硝基苯胺底物裂解的时间-动力学测量。
详述
定义
将关于特定实施方案并参考某些附图来描述本发明,但是本发明不限于此,而是仅由权利要求来限定。权利要求中的任何附图标记不应被解释为限制所述范围。所描述的附图仅是示意性的而非限制性的。在附图中,出于说明性目的,一些元件的尺寸可能被放大并且未按比例绘制。在本说明书和权利要求书中使用术语“包括”时,其不排除其他元件或步骤。当提到单数名词时使用不定冠词或定冠词的情况中,例如除非特别说明,否则“一个(a)”或“一个(an)”,“该(the)”包括该名词的复数形式。此外,说明书和权利要求书中的术语第一、第二、第三等用于区分相似的元件,而不必定是用于描述顺序或时间顺序。应当理解,如此使用的术语在适当的情况下是可互换的,并且本文描述的本发明的实施方案能够以不同于本文描述或示出的其他顺序来操作。提供以下术语或定义仅是用于帮助理解本发明。除非本文具体定义,否则本文所用的所有术语具有与本发明领域的技术人员所知的相同含义。对于本领域的定义和术语,从业者特别指向michaelr.green和josephsambrook,molecularcloning:alaboratorymanual,第4版,coldspringharborlaboratorypress,plainsview,newyork(2012);和ausubel等,currentprotocolsinmolecularbiology(增补47),johnwiley&sons,newyork(1999)。本文提供的定义不应被解释为具有小于本领域普通技术人员所理解的范围。
在本申请中,申请人描述了一种使用多步法进行肽测序的方法,其中n-末端氨基酸被一个一个地鉴定。更精确地,提供了一种对多肽进行测序的方法,其中所述方法包括以下步骤:a)将所述多肽接触裂解诱导剂,更具体地为催化活性氨肽酶、异硫氰酸酯或异硫氰酸酯类似物;b)测量所述试剂在所述多肽的n-末端氨基酸上的停留时间,或替换地,测量所述酶促反应的kcat值;c)通过所述停留时间或所述kcat值鉴定或分类所述n-末端氨基酸;d)重复步骤a)至c)一次或多次。在一个实施方案中,将所述多肽固定在表面上。不用说,当所述试剂从所述多肽裂解n-末端氨基酸时,所述多肽通过其c-末端固定在表面上。反之亦然,当所述试剂从所述多肽裂解c-末端氨基酸时,所述多肽通过其n末端固定在表面上。在另一个实施方案中,所述方法是在单分子水平上对表面固定的多肽进行测序的方法。
鉴于所述的肽测序方法依赖于n末端氨基酸的顺序鉴定,因此本申请同样公开了一种通过确定裂解诱导剂(更特别地,催化活性氨肽酶、异硫氰酸酯或异硫氰酸酯类似物)在所述n-末端氨基酸上的停留时间来鉴定或分类多肽的n末端氨基酸的方法。所述方法包括将所述多肽接触所述试剂,并测量所述试剂的停留时间,或替换地,所述酶促反应的kcat值。此外,在这种情况下,所述方法可以在单分子水平上使用,使用或不使用表面固定的肽。因此,在一个实施方案中,所述方法以单分子水平鉴定或分类表面固定多肽的n-末端氨基酸。
如本文所用的,术语“肽”和“多肽”可互换使用,并且是指任何长度的氨基酸的聚合形式,其可以包括编码和非编码的氨基酸,天然和非天然的氨基酸,化学或生物化学修饰或衍生的氨基酸,以及具有修饰的肽主链的多肽。如本文所用的,“肽”或“多肽”比衍生它们的全长蛋白质短,并例如但没有限制的目的,通过胰蛋白酶或蛋白酶k蛋白消化来形成。在特定的实施方案中,所述肽或多肽具有20至500个,或25至200个或30至100个氨基酸的长度,或具有小于500个,小于250个,小于200个,小于150个,少于100个或少于50个氨基酸的长度。在任何情况下,“肽”或“多肽”包括至少2个,至少3个,至少4个,至少5个,至少6个,至少7个,至少8个,至少9个,至少10个或至少20个氨基酸。
以单分子方式或以单分子水平或在单分子实验中使用的“单分子”是指对单个分子的性质的研究。单分子研究可能与对分子的整体或大量集合的测量(在这种情况下,无法区分分子的单独行为,只能测量平均特征)形成对比。
蛋白质是氨基酸聚合物。蛋白质由核糖体产生,核糖体在称为翻译的过程中“读取”基因中密码子编码的rna,并根据遗传指令组装必需的氨基酸组合。新形成的蛋白质链随后进行翻译后修饰,其中添加了其他原子或分子,例如铜、锌或铁。一旦这个翻译后修饰过程完成,蛋白质开始折叠(有时自发,而有时在酶促帮助下),自身卷曲,使蛋白质的疏水性元件深埋在结构内部,而亲水性元件最终在蛋白质外。蛋白质的最终形状或结构决定了蛋白质与环境的相互作用方式。因此,蛋白质具有一级结构(即通过共价肽键连接在一起的氨基酸序列),二级结构(即规则的重复模式,如α-螺旋和β折叠),三级结构(即氨基酸侧链之间的共价相互作用,如半胱氨酸基团之间的二硫化物桥)和四级结构(即彼此相互作用的蛋白质亚基)。然而,对于本申请中公开的方法,蛋白质及其n-末端氨基酸应当是本申请的氨肽酶可接近的,并且优选该蛋白质以线性构型固定。因此,在各种实施方案中,将待测序的蛋白质变性。变性是其中蛋白质失去其天然状态中存在的四级结构,三级结构和二级结构,但是氨基酸之间的一级结构的肽键保持完整的过程。蛋白质变性可以通过施加外部应力或化合物来实现,如强酸或强碱,浓无机盐,有机溶剂(例如,醇或氯仿),辐射或热。因此,在本申请的各种实施方案中,待使用的氨肽酶是嗜热的和/或耐溶剂的(见下文)。如本文所用的,“嗜热的”是指“提高的温度耐受性”,更确切地是指在40至122℃之间相对高的温度下其活性旺盛或维持其活性的生物体或酶。嗜热生物体的非限制性实例是水生栖热菌,并且因此其酶,如其氨肽酶t在高温下起作用,因此是嗜热的。在特定的实施方案中,用于本申请的用途和方法的氨肽酶在40℃至100℃或40℃至80℃或50℃至70℃或60℃至80℃的温度范围内具有最佳的肽酶活性。在其他特定的实施方案中,本申请的氨肽酶在溶剂(如乙酸、三氯乙酸、磺基水杨酸、碳酸氢钠、乙醇、醇、交联剂(如甲醛和戊二醛)、离液剂,如脲、氯化胍或高氯酸锂,破坏二硫键的试剂,如2-巯基乙醇、二硫苏糖醇或三(2-羧乙基)膦)的存在下保持其酶活性。
与不同的n-末端氨基酸结合时,使用嗜热和/或耐溶剂的氨肽酶特别有助于微调所述氨肽酶的“工作时间”值。通过改变实验过程(例如蛋白质测序)中的反应条件,可以调节温度、ph值、溶剂……,以进一步区分与氨基酸y的“工作时间(on-time)”值相比的氨基酸x的“工作时间”值。
在本申请的方法中,从多肽底物裂解n端氨基酸。这可以例如通过化学方法或通过肽酶,更特别是通过氨肽酶,通过酶法来实现。所述裂解诱导剂可以共价或非共价地结合所述n端氨基酸。
化学裂解诱导剂
edman降解是一种化学技术,其可以对蛋白质进行n端测序。最早由pehredman在1950年描述,并在1967年将降解反应完全自动化。该方法需要将异硫氰酸苯酯(pitc)偶联到蛋白质/肽的游离n端上(碱性条件),然后作为乙内酰苯硫脲(pth)衍生物释放n末端氨基酸(酸性条件)(图15)。然后用例如色谱鉴定释放的pth-氨基酸。然后连续重复该过程,得到蛋白质/肽序列信息。
在本申请中,令人惊讶地发现,即使在没有酸性条件或热的情况下,pth也被释放,并且因此从多肽底物裂解n端氨基酸。甚至更令人惊讶地,发明人发现结合异硫氰酸酯(或类似物)与n末端氨基酸裂解之间的时间取决于氨基酸的特性。因此,通过测量所述时长,其基本上是异硫氰酸酯(或类似物)在n-末端氨基酸上的停留时间,可以鉴定出n-末端氨基酸。
在本申请的一个优选实施方案中,在本申请的用途和方法中提及的裂解诱导剂是化学试剂,更特别地是选自异硫氰酸酯(itc)、异硫氰酸苯基酯(pitc)、叠氮基-pitc,香豆素-异硫氰酸酯(citc)和磺基异硫氰酸苯酯(spitc)的化学试剂。在本申请中,pitc、叠氮基-pitc、citc和spitc将被称为异硫氰酸酯类似物。因此,提供了本申请的方法,其中裂解诱导剂是异硫氰酸酯或异硫氰酸酯类似物。
本文还提供了裂解诱导剂用于获得经由其c末端固定在表面上的多肽的序列信息的用途,其中所述裂解诱导剂在所述多肽的n-末端氨基酸上的停留时间鉴定了所述n端氨基酸。在特定的实施方案中,所述裂解诱导剂是异硫氰酸酯或异硫氰酸酯类似物,并且所述停留时间是itc或itc类似物在n末端氨基酸上结合与去除所述n末端氨基酸之间的时长。随后可以通过将所述时长与针对不同氨基酸的itc或itc类似物获得的一组参照值进行比较来鉴定所述n末端氨基酸。
氨肽酶
如本文所用的,“氨肽酶”是指催化氨基酸从蛋白质或肽底物的氨基末端(n末端)裂解的酶。它们广泛分布于整个动物和植物界,并存在于许多亚细胞器、胞质和膜成分中。氨肽酶通过以下来分类:1)从底物的氨基末端裂解的氨基酸数量(例如,氨基二肽酶去除完整的氨基末端二肽,氨基三肽酶催化氨基末端三肽的水解),2)氨肽酶在细胞中的位置,3)对抑氨肽酶抑制的易感性;4)金属离子含量和/或将金属结合到酶上的残基;5)观察到最大活性的ph和6)与本申请最相关的,通过使用其除去残基的相对效率(taylor1993fasebj7:290-298)。氨肽酶可以具有广泛或小的底物特异性。在本申请中,重点是广泛底物特异性的氨肽酶的开发或使用,但是在本申请中还设想了使用底物特异性重叠或互补的多种氨肽酶。
通常,可以通过特异性常数kcat/km来量化酶在特定环境条件下对特定底物的特异性。kcat是周转数,每个酶位点每单位时间转化成产物的底物分子数,或每个催化中心和每单位时间生产性底物转化成产物反应物的数量。km定义为在有效稳态酶动力学测量所需的条件下,酶达到其最大速度的一半所需的底物浓度,这是本领域公知的。在区分两种酶底物a和b时,根据这些底物到产物的转化率,成立这种关系:
v为速度,和[a]为a的浓度。
因此,可以从酶对这些底物的转化速度测量中获得关于酶的不同底物的身份的信息。在底物浓度相等的条件下,相对速度由kcat和km来确定。当观察单个底物分子时,一旦添加酶,形成产物分子所需的时间由kcat控制。因此,在单分子观察中,可以从酶在底物上的“工作时间”或停留时间获得底物身份的信息。通过将底物和/或酶工程化,可以进一步补充该信息,从而可以将酶和底物的催化生产性啮合(engagement)与非生产性啮合区分开来。因此,如本文所用的,“工作时间”是指酶在底物上的停留时间,酶溶液与底物的接触时间,或更具体地是kcat的倒数,这是本领域公知的。从这里开始,“工作时间”和停留时间将互换使用,并且可以指一种酶分子作用于一个肽分子直至发生裂解的时间,或指多个酶分子顺序作用于该肽分子直至发生裂解所需要的时间。
底物上酶的“工作时间”的观察可用于鉴定所述底物对于氨肽酶尤其成立。肽酶通常通过两步机制运行(图10)。首先,在酰化反应过程中,肽的n-末端部分(对于氨肽酶)或肽的c-末端部分(对于羧肽酶)被切除并与肽酶共价连接。第二,在脱酰反应中,酶释放裂解的氨基酸。
氨肽酶通过立体电子与酰化反应过渡态的配合而获得其对特定(组的)氨基酸的特异性,这尤其受易裂键n-末端的底物侧链的性质的影响。通常,氨肽酶与易裂键c-末端的肽部分之间的结合相互作用要少得多,因此在确定反应速率的酰化或水解步骤之后,会迅速从肽(或从肽结合的表面)解离。如果将肽在被肽酶裂解的易裂肽键的c-末端固定,则在酰化反应时,在丝氨酸或半胱氨酸肽酶的情况中,肽的n-末端氨基酸或氨基酸衍生物与酶共价连接,或在直接水解肽酶的情况中,与酶非共价结合,而c-末端部分将保持与固定肽的表面结合(图10)。因此,对于选定的氨肽酶,在表面固定的肽底物上的停留时间或“工作时间”与酰化或水解步骤的速率相关,因此与易裂键的n-末端部分的性质相关。在这种情况下,可以通过分子标记所述氨肽酶来容易地确定氨肽酶的“工作时间”。这样,分子标记物充当氨肽酶的“工作时间”的指标,并因此充当被所述氨肽酶切除的n-末端氨基酸的身份的指标。在本申请的特定实施方案中,所述氨肽酶可以通过光、荧光学、电学或等离子体标记(见下文)。
在可替换的实施方案中,将氨肽酶分子的溶解接触肽底物,并且测量直至n-末端氨基酸(或其衍生物)被切除的停留时间/工作时间。在这样的实施方案中,测量直至这样的裂解事件时的酶接触底物的整体停留时间,并且将这个值与所用条件下酶对肽底物上的特定n-末端氨基酸(衍生物)的kcat的倒数相关。
对于来自半胱氨酸和丝氨酸蛋白酶组的羧肽酶,情况则不同。更精确地,在所述羧肽酶的情况中,酶在切除c-末端氨基酸后保持与固定的肽部分共价结合。羧肽酶在酰化步骤之后将不会从肽解离,并且将通过脱酰(水解)步骤的速率来确定其对固定表面上的肽的“工作时间”值。后面的水解步骤对于c-末端氨基酸(其已经在酰化步骤中释放到溶剂中)的性质的信息很少或没有提供信息。然而,在将羧肽酶分子的溶液接触肽底物并测量直至c-末端氨基酸(或其衍生物)被切除的停留时间/工作时间的实施方案中,该值与所用条件下酶对肽底物上的特定c-末端氨基酸(衍生物)的kcat的倒数相关,并且这种羧肽酶可用于本发明的范围中。
令人感兴趣的,来自金属蛋白酶组的羧肽酶不进行这种共价结合,并且不会通过水解切除c-末端氨基酸。因此,所述羧基-金属肽酶的“工作时间”对于其结合并裂解的c-末端氨基酸提供与氨肽酶对于n-末端氨基酸那样同等地提供信息。因此,在本申请中描述的方法中也设想使用羧基-金属肽酶,但重要的区别在于,随后将多肽通过其n端或通过肽的侧链固定在表面上。总之,除了对于本申请特别有用的异硫氰酸酯和/或itc类似物的用途之外,且本文公开的方法是氨肽酶或羧基-金属肽酶。
因此,在本申请的各种具体实施方案中,本申请的方法和用途中所指的裂解诱导剂是肽酶,特别是氨肽酶或羧基-金属肽酶,更特别是氨肽酶。
在本申请的特定实施方案中,提供了一种活性肽酶的用途,所述肽酶的裂解速率或肽酶活性的动力学是特征性的,并因此鉴定了氨基酸底物,尤其是多肽的末端氨基酸。一种合乎需要的策略是利用氨肽酶,特别是一种独特的氨肽酶,更特别地,其识别20种可能的n-末端氨基酸中的每一种的催化活性氨肽酶。然而,还设想使用2、3、4或更多种氨肽酶,其随后可以区分不同组的氨基酸,例如但没有限制目的:芳香族氨基酸与非芳香族氨基酸,或疏水性末端氨基酸,正电荷氨基酸,负电荷氨基酸和小氨基酸。
还可以设想,通过改变实验过程(例如蛋白质测序)中的反应条件,可以改变特定n-末端氨基酸的氨肽酶的“工作时间”值。当所用的氨肽酶对于某些n端氨基酸具有非常相似的“工作时间”值时,这尤其理想。因此,在某些实施方案中,调节反应条件,尤其包括温度、ph、溶剂等,以增加与氨基酸y的“工作时间”值相比时与氨基酸x的“工作时间”值之间的区别。在其他实施方案中,氨肽酶本身被工程化以区分天然氨肽酶对其具有相似停留时间的不同氨基酸。如本文所用的,“工程化的”是“合成的”、“重组的”、“人造的”或“非天然的”的同义词。
作为非限制性实例,来自水生栖热菌的氨肽酶可以用于本申请的方法中(参见下文)。工程化所述氨肽酶的可能的残基是距催化位点中的二价金属离子8埃半径内的残基,更确切地是e250、f252、g315、e316、v317、a318、t336、e340、h345、i346、a347、f348、q350、y352、n355、h376、v377、d378和/或w379。所述残基的位置是指如seqidno.7中所示的野生型氨肽酶中的位置。
另一种理想的策略是本申请中所述的氨肽酶结合时,酶“工作时间”值可以用于鉴定固定多肽的n-末端氨基酸。在本申请中还设想了氨肽酶,更特别地催化活性氨肽酶,其用途和其中使用所述氨肽酶的方法,所述酶的“工作时间”值可提供一组或亚组氨基酸的说明或信息。在某些实施方案中,因此酶“工作时间”值以一定概率将一组或亚组氨基酸中的n-末端氨基酸分类或归类。
在某些实施方案中,本申请中公开的方法中使用的氨肽酶是氨基二肽酶,更特别地是催化活性氨基二肽酶。氨基二肽酶是二氨基肽酶的同义词,并且是指切除多肽的两个最末端的n-末端氨基酸的酶。
在优选实施方案中,所述氨肽酶或本申请中公开的方法中使用的氨肽酶是催化活性的。“催化活性的”表示氨肽酶是完全功能性的催化酶。这与已经工程化来结合n-末端氨基酸但不裂解所述n-末端氨基酸的催化死亡氨肽酶相反,例如,wo20140273004中的。
对于本申请中所述的用途和方法,尤其设想了嗜热氨肽酶。可以用于本申请中所述的方法中的这样的氨肽酶的非限制性实例是来自水生栖热菌的氨肽酶t(ampt_theaq)、来自嗜热栖热菌(thermusthermophilus)的氨肽酶t(ampt_thet8)、来自嗜热链球菌(streptococcusthermophiles)的pepc(pepc_strtr)、来自灰色链霉菌(streptomycesgriseus)的氨肽酶s(apx_strgg)、来自横膈链霉菌(streptomycesseptatus)th-2的氨肽酶(q75v72_9actn)和来自嗜热脂肪芽孢杆菌(bacillusstearothermophilus)的氨肽酶2(amp2_geose)。
本申请方法中设想的和本文公开的催化活性氨肽酶的非限制性实例是工程化克氏锥虫(trypanosomacruzi)cruzipain或cruzain和水生栖热菌氨肽酶t。
在本申请中,提供了工程化的、催化活性氨肽酶,其包含针对多肽的任何n-末端氨基酸或针对多肽的一系列不同的n-末端氨基酸的结合结构域,其中将所述多肽固定在表面上,其中所述氨肽酶在结合时裂解n-末端氨基酸,并且其中所述氨肽酶的酶“工作时间”鉴定或分类所述n-末端氨基酸。本文还提供了结合表面固定多肽的工程化的、催化活性氨肽酶,其中所述氨肽酶裂解所述多肽的n-末端氨基酸,和其中所述氨肽酶在所述n-末端氨基酸上的停留时间鉴定或分类所述n-末端氨基酸。
在特定的实施方案中,将所述多肽通过易裂键的肽c-末端部分固定于所述表面上。在另一个特定的实施方案中,所述n-末端氨基酸是衍生的n-末端氨基酸和所述氨肽酶结合并裂解所述衍生的n-末端氨基酸。在更特别的实施方案中,所述衍生的n-末端氨基酸是用itc、citc、spitc、pitc、叠氮基-pitc或叠氮基-pitc的点击化学修饰的产物(此后总称为“叠氮基-pitc”)衍生的n-末端氨基酸,并且所述氨肽酶分别结合并裂解itc、citc、spitc、pitc或叠氮基-pitc衍生的n-末端氨基酸。
“衍生的”源自“衍生”,衍生是指在化学中使用的技术或生物化学机理,其将化合物转化成相似化学结构的产物(反应的衍生物),称为衍生物。通常,化合物的特定官能团参与衍生反应并将产物转化为偏离、反应性、溶解度、沸点、熔点、聚集态、化学组成、相互作用或光学、电学或等离子体特征性的衍生物。在可替换的实施方案中,衍生的表示标记的。
在另一个实施方案中,所述n-末端氨基酸选自leu、met、tyr、arg、pro、gly、lys、ala和val,并且所述针对一系列不同n-末端氨基酸的结合结构域或所述针对任何n-末端氨基酸的结合结构域是针对选自leu、met、tyr、arg、pro、gly、lys、ala和val的n-末端氨基酸的结合结构域。在另一个实施方案中,所述n-末端氨基酸是leu、met、tyr、arg、pro、gly、lys、ala和val,并且所述针对任何n-末端氨基酸的结合结构域是针对leu、met、tyr、arg、pro、gly、lys、ala和val的结合结构域。
在另一个特定的实施方案,提供了工程化的、合成的或重组的氨肽酶,其包含针对多肽的一个或多个不同衍生的或标记的n-末端氨基酸的结合结构域,其中所述氨肽酶在结合所述衍生的或标记的n-末端氨基酸时裂解所述衍生的或标记的n-末端氨基酸,和其中所述氨肽酶的分裂速率或氨肽酶活性的动力学鉴定所述n-末端氨基酸。在一个实施方案中,所述衍生的或标记的n-末端氨基酸是用itc、citc、spitc、pipc或叠氮基-pitc衍生或标记的n-末端氨基酸。在另一个实施方案中,所述衍生的或标记的n-末端氨基酸是衍生的或标记的leu、met、tyr、arg、pro、gly、lys、ala和val。所述氨肽酶因此是催化活性的,并且不是催化死亡氨肽酶。
在一个方面中,本申请提供了工程化的、合成的或重组的氨肽酶,其包含具有野生型克氏锥虫cruzipain或cruzain的25位的甘氨酸残基,65位的丝氨酸残基,138位的半胱氨酸残基和208位的组氨酸残基的氨基酸序列,其中所述氨肽酶的剩余氨基酸序列包含与所述野生型克氏锥虫的seqidno.1中所示的cruzipain或seqidno.2中所示的cruzain具有至少75%,至少80%,至少81%,至少82%,至少83%,至少84%,至少85%,至少86%,至少87%,至少88%,至少89%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%,至少99%或100%序列同一性的序列。即提供了与seqidno.1或seqidno.2具有至少75%,至少80%,至少81%,至少82%,至少83%,至少84%,至少85%,至少86%,至少87%,至少88%,至少89%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%,至少99%或100%序列同一性并且具有25位的甘氨酸残基,65位的丝氨酸残基,138位的半胱氨酸残基和208位的组氨酸残基的工程化氨肽酶。在一个实施方案中,在所述氨肽酶的第一个甲硫氨酸残基后插入半胱氨酸残基。所述半胱氨酸用于标记所述工程化的cruzipain或cruzain氨肽酶。在特定的实施方案中,所述氨肽酶包含seqidno.3或seqidno.4中所示的氨基酸序列。在更特别的实施方案中,所述氨肽酶由seqidno.5或seqidno.6中所示的氨基酸序列组成。在特别的实施方案中,以上方面及其实施方案的所述氨肽酶是催化活性的氨肽酶。
克氏锥虫cruzipain和cruzain中的上述特定突变允许活性(结合并裂解)朝向衍生的n-末端氨基酸,更特别的citc-或spitc-衍生的n-末端氨基酸。因此,方法或本申请用于对包含衍生的氨基酸的肽测序或鉴定衍生的n-末端氨基酸。在特别的实施方案中,所述衍生的氨基酸是用itc、pitc、叠氮基-pitc、citc和/或spitc衍生的氨基酸,甚至更特别的是用citc和/或spitc衍生的。
除了上述裂解citc-或spitc-衍生的n-末端氨基酸的工程化克氏锥虫cruzipain和cruzain在本申请所述的方法中的使用以外,所述工程化的克氏锥虫cruzipain和cruzain还可以用于wo20140273004所述的方法中。在后一文献中,通过一系列n末端氨基酸结合蛋白来鉴定衍生的n末端氨基酸,因此通过edmanase去除所鉴定的n末端氨基酸。在n-末端氨基酸是用citc或spitc衍生的情况中,本申请中所述的工程化克氏锥虫cruzipain和cruzain可以用作edmanase。
在接来下的方面中,提供了与seqidno.7中所示的野生型水生栖热菌氨肽酶t的氨基酸序列具有至少75%,至少80%,至少81%,至少82%,至少83%,至少84%,至少85%,至少86%,至少87%,至少88%,至少89%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%,至少99%或100%序列同一性的工程化的、合成的或重组的氨肽酶,其中在所述野生型氨肽酶的第一个甲硫氨酸残基后插入半胱氨酸残基。即提供了与seqidno.7具有至少75%,至少80%,至少81%,至少82%,至少83%,至少84%,至少85%,至少86%,至少87%,至少88%,至少89%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%,至少99%或100%序列同一性的工程化氨肽酶,其中在第1位的甲硫氨酸残基和第2位的丙氨酸残基之间插入半胱氨酸残基。在特别的实施方案中,所述氨肽酶t包含seqidno.8中所示的氨基酸序列。在更特别的实施方案中,所述氨肽酶由seqidno.8中所示的氨基酸序列组成。在特别的实施方案中,以上方面及其实施方案的所述氨肽酶是催化活性的氨肽酶。
如本文使用的,在两个或更多个多肽序列语境中的术语“相同的”、“相似性”或“同一性”百分比或“相似性”百分比或“同源性”百分比是指使用序列比较算法或通过人工比对和视觉观察测量的在比较窗口或指定区域针对最大相应性进行比较和比对时,相同或具有特定百分比的相同氨基酸残基的两个或更多个序列或子序列。优选,同一性存在于长度为至少约25个氨基酸的区域上,或更优选存在于50-100个氨基酸的区域中,甚至更优选存在于长度为100-500个氨基酸或甚至更长的区域中。
如本文所用的,术语“序列同一性”或“序列同源性”是指在比较窗口中,序列在氨基酸基础上氨基酸相同的程度。因此,通过在比较窗口中比较两个最佳比对的序列,确定两个序列中出现相同氨基酸的位置数以产生匹配的位置数,将匹配的位置数除以比较窗口中的位置总数(即,窗口大小),然后将结果乘以100得出序列同一性百分比,从而计算出“序列同源性百分比”。认为缺口(即,比对中在一个序列中存在残基但在另一个序列中不存在残基的位置)是具有不同残基的位置。确定序列同源性的百分比可以手动完成,或通过使用本领域可用的计算机程序来完成。有用的算法的实例是pileup(higgins&sharp,cabios5:151(1989),blast和blast2.0(altschul等,j.mol.biol.215:403(1990))。用于执行blast分析的软件可通过国家生物技术中心公开获得(http://www.ncbi.nlm.nih.gov/)。在特定的实施方案中,用于确定两个或更多个多肽(如氨肽酶)的序列同一性的比较窗口是全长蛋白质序列。
固定和标记
如本文所用的“固定在表面上”是指将一个或多个多肽连接惰性、不溶性材料(例如玻璃表面),导致所述多肽的活动性丧失。对于本申请公开的方法,固定允许多肽在多肽的测序或在鉴定或分类所述多肽的n-末端氨基酸的整个过程中保持在适当的位置。由此,n端应当可自由接近,因此多肽应当通过其c-末端固定。而且,固定在高密度表面上的蛋白质允许使用少量样品溶液。过去几年中已经开发了许多固定技术,这些固定技术主要基于以下三种机制:物理、共价和生物亲和力固定(rusmini等2007biomacromolecules8:1775-1789;美国专利号6,475,809;wo2001040310;us7358096;us20100015635;wo1996030409)。在本申请中,根据本领域可用的方案,例如,eeftens等(2015bmcbiophys8:9),使用叠氮化物-二苯并环辛基(dbco)点击反应将多肽固定在玻璃表面上(参见实施例1和2)。
在本申请的各种实施方案中,可以在接触氨肽酶之前,将多肽固定在表面上。可以将肽固定在任何合适的表面上(参见下文)。对于本申请中公开的方法至关重要的是,待测序的多肽或待鉴定或分类其n端氨基酸的多肽是通过多肽的最c-末端的部分或通过易裂键的c-末端部分固定的。因此,多肽以其c端或沿着肽结构的一部分、易裂键的c-末端(例如,通过例如马来酰亚胺化学或金-硫醇键的半胱氨酸的硫醇官能,这是本领域公知的)连接到应用表面。
如本文所用的,“易裂键”是指待通过本申请的氨肽酶之一裂解的共价化学键。
如本文所用的,“表面”是载体或层的同义词。本申请的表面或层适用于检测分子标记、电化学信号、电磁信号、等离子体相关事件。所述分子标记可以是光(包括但不限于发光和荧光标记)或电(包括但不限于电位、伏安、库仑标记)标记。
所述层也可以是多层,即,包含几层的层。在多层的情况中,至少一层应当允许合适地检测所述分子标记或所述电化学、电磁或等离子体相关的事件。因此,根据特定的实施方案,表面是活性传感表面。因此,所述在单分子水平将表面固定的多肽测序的方法的表面固定肽是固定在活性传感表面上的多肽。在更特别的实施方案中,所述活性传感表面是金表面或酰胺-、羧基-、硫醇-或叠氮基-官能化的表面,所述方法的多肽通过化学偶联在其上。在其他特别的实施方案中,所述载体是纳米颗粒、纳米盘、纳米结构、芯片。在最特别的实施方案中,所述表面是自组装单层(sam)。
为了检测“工作时间”值或停留时间,可以选择两种标记选项。首先,例如,可以通过其n-末端氨基酸来标记待测序的多肽。替换地或另外地,例如可以如图14中所示的标记内部氨基酸。可以使用荧光探针,如但不限于荧光胺、邻-苯二醛、丹酰氯和香豆素异硫氰酸酯(citc),来进行多肽的标记。在特别的实施方案中,本申请的固定多肽的n-末端氨基酸是citc衍生的或替换地说是用citc标记的。多肽还可以通过电来标记。电分析方法是一类通过测量分析物、肽、酶……上的电学标记的电位(伏特)和/或电流(安培)来确定分析物、肽、酶……存在的技术。这些方法可以根据标记分为几类。三个主要类别是电位法(测量电极电位的差异)、库仑法(随时间测量电流)和伏安法(在主动改变电位时测量电流)。库仑技术有两个基本类别。恒电位库仑法涉及使用恒电位仪在反应过程中保持电位恒定。另一种称为库仑滴定法或安培库仑法,使用安培计使电流(以安培测量)保持恒定。电标签的非限制性实例是磺异硫氰酸苯酯(spitc)。spitc是用于ms从头肽测序中的异硫氰酸苯酯(pitc)探针的带负电荷的变体,用于中和n末端片段离子(samyn等,2004jamsocmassspectrom15:1838-1852)。在特别的实施方案中,可以通过电位、安培或伏安来进行电标记。
为了检测和测量本申请的氨肽酶在固定多肽的n末端氨基酸上的或直至固定多肽的n末端氨基酸被切除的“工作时间”值或停留时间,(见上文),需要检测所述氨肽酶。氨肽酶可以在测得的停留时间内与底物进行有效裂解(cleavage-productive)或非有效裂解(cleavage-unproductive)的反应,直至n-末端氨基酸被切除。在这两种相互作用的类型中,它们的长度、长度总和和平均长度可以是与本发明有关的所测量的停留时间的一部分,因为这些参数都是测量的一部分,其提供了直至氨肽酶切除n-末端氨基酸需要多长时间的信息。检测的性质对于本发明不是至关重要的,只要可以检测到酶“工作时间”或氨肽酶的停留时间即可。在本申请的某些实施方案中,通过光学、电学或等离子体检测氨肽酶的“工作时间”。检测本申请的氨肽酶的一种方法是将其与分子标记融合,然后检测该分子标记。与以上相似,可以对氨肽酶进行光学、电学或等离子体标记。
光学检测需要光学标记,并且包括但不限于发光和荧光检测。标记因此可以是荧光团。在商业上,有大量可用的光学标记类别,包括但不限于cy3、cy5、香豆素、alexafluor标记、gfp、yfp、rfp,……。在某些实施方案中,氨肽酶的标记与固定多肽的或所述多肽的n-末端氨基酸的或固定表面的标记相互作用。所述标记可以是例如荧光团。在另一个实施方案中,第一组和第二组标记分子之间至少有一个共同的分子。在一个实施方案中,检测步骤产生图像,例如荧光图像(例如,使用荧光共振能量转移(fret)、全内反射荧光(tirf)或零模式波导(zmw)获得的图像)。在另一个实施方案中,图像的汇编产生数字概况,例如,鉴定固定多肽或其n-末端氨基酸的数字概况。在特定的实施方案中,光学标记的是荧光标记的。在甚至更特别的实施方案中,通过tirf显微术测量或检测荧光标记。
无需分子标记,也可以检测氨肽酶在固定蛋白质的n-末端氨基酸上的结合,从而可以测量氨肽酶的停留时间或“工作时间”。无标记电学检测的非限制性实例是使用基于场效应晶体管的生物传感器或biofet。biofet是一种场效应晶体管,其受分子结合(即固定化多肽的n-末端氨基酸上的氨肽酶结合)诱导的表面电势变化所门控(gate)。当带电荷的分子(如spitc标记的氨肽酶)结合fet栅极(其通常是介电材料)时,它们可以改变下面的半导体材料的电荷分布,从而导致fet通道的电导率发生变化。biofet由两个主要部分组成:一个是生物识别元件,另一个是场效应晶体管(fet)。
通过修饰栅极或将其与不同的生物识别元件(受体)耦合,可以简单地由离子敏感场效应晶体管(isfet)、硅纳米线(sinw)、电容性eis传感器和光寻址电位传感器(laps)构造biofet(poghossian和
同样,等离子体读数可以用于检测本申请的氨肽酶或itc或itc类似物的“工作时间”。在物理学中,等离子体可以定义为通常在(贵金属)金属与电介质之间的界面上自由电子集体振荡的量子(quantum)。术语等离子体是指在电磁辐射的影响下,金属中自由电子的等离子体样行为。表面等离子体是相干的离域电子振荡,其存在于任意两种材料之间的界面,其中介电功能的实部在整个界面(例如金属-电介质界面,如空气中的金属薄板)上改变信号。表面等离子体的激发通常可以用一种称为表面等离子体共振(spr)的实验技术来完成,这种激发可以利用电磁波谱可见范围内的光非常有效地完成。在spr中,表面等离子体的最大激发是通过监测来自棱镜耦合器的反射功率(作为入射角或波长的函数)来检测的。该技术可用于观察厚度、密度波动或分子吸收的纳米变化,并用于筛选和定量蛋白质结合事件。使用根据这些原理操作的商业化工具是可获得的。因此,在特定的实施方案中,通过表面等离子体共振来确定本申请的氨肽酶的或itc的或itc类似物的“工作时间”。
可以测量氨肽酶的或itc的或itc类似物的“工作时间”的另一个实例是等离子体增强的回音通道微腔传感器,如针对聚合酶dna相互作用所证明的那样(kim等2017sciadv3:e1603044)。
然而,尽管有上述内容,但必须清楚的是,标记和随后检测的性质对于本发明而言并不重要,只要可以检测到裂解诱导剂的“工作时间”或停留时间即可。
裂解诱导剂的用途
在本申请的另一个方面中,提供了裂解诱导剂获得表面固定多肽的序列信息的用途,其中所述裂解诱导剂在所述多肽的末端氨基酸上的停留时间鉴定了所述末端氨基酸。在一个实施方案中,所述末端氨基酸是衍生的。在另一个实施方案中,所述裂解诱导剂是异硫氰酸酯和异硫氰酸酯类似物或是肽酶。更特别地,所述异硫氰酸酯类似物选自itc、citc、pitc、citc和叠氮基-pitc。所述肽酶是催化活性的肽酶,更特别地,是催化活性的氨肽酶。
在再另一个实施方案中,提供了本申请中所述的任一种氨肽酶用于裂解多肽的n-末端氨基酸的用途。在一个实施方案中,所述多肽是表面固定的。在另一个实施方案中,在单分子水平进行所述多肽的n-末端氨基酸的裂解。在特别的实施方案中,提供了工程化的、合成的或重组的氨肽酶用于裂解多肽的n-末端氨基酸的用途,所述氨肽酶包含具有野生型克氏锥虫cruzipain或cruzain的25位的甘氨酸残基,65位的丝氨酸残基,138位的半胱氨酸残基和208位的组氨酸残基的氨基酸序列,其中所述氨肽酶的剩余氨基酸序列包含与seqidno.1中所示的所述野生型克氏锥虫cruzipain或seqidno.2中所示的cruzain具有至少75%,至少80%,至少81%,至少82%,至少83%,至少84%,至少85%,至少86%,至少87%,至少88%,至少89%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%,至少99%或100%序列同一性的序列。甚至更特别地,为裂解多肽的n-末端氨基酸提供的所述氨肽酶在野生型克氏锥虫cruzipain或cruzain的第一个甲硫氨酸残基后具有插入的半胱氨酸残基。甚至更特别地,为裂解多肽的n-末端氨基酸提供的所述氨肽酶包含seqidno.3或seqidno.4中所示的氨基酸序列或由seqidno.5或seqidno.6中所示的氨基酸序列组成。最特别地,在单分子水平进行所述裂解,并且所述多肽是表面固定的,更特别地,通过所述多肽的c-末端进行表面固定。
在另一个特别的实施方案中,提供了工程化的、合成的或重组的氨肽酶用于裂解多肽的n-末端氨基酸的用途,所述氨肽酶与seqidno.7中所示的野生型水生栖热菌氨肽酶t的氨基酸序列具有至少75%,至少80%,至少81%,至少82%,至少83%,至少84%,至少85%,至少86%,至少87%,至少88%,至少89%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%,至少99%或100%序列同一性,其中在所述野生型氨肽酶的第一个甲硫氨酸残基后插入半胱氨酸残基。在更特别的实施方案中,为裂解多肽的n-末端氨基酸提供的所述氨肽酶t包含seqidno.8中所示的氨基酸序列或由seqidno.8中所示的氨基酸序列组成。最特别地,在单分子水平进行所述裂解,并且所述多肽是表面固定的。在其他实施方案中,所述氨肽酶是催化活性的氨肽酶。还提供了本文公开的水生栖热菌氨肽酶t从表面固定的多肽裂解或鉴定n-末端氨基酸的用途,其中所述n-末端氨基酸选自leu、met、tyr、arg、pro、gly、lys、ala和val。
在再另一个方面中,提供了氨肽酶(更特别的催化活性氨肽酶)用于鉴定或分类多肽的n-末端氨基酸或用于多肽测序的用途。在特别的实施方案中,所述鉴定、分类或测序在单分子水平进行。甚至更特别地,所述多肽通过其c-末端进行表面固定。在一个实施方案中,所述氨肽酶是标记的,特别是用光学、电学或等离子体标记标记的,或通过光学、电学或等离子体来检测所述氨肽酶。在特别的实施方案中,提供了工程化的、合成的和重组的氨肽酶用于鉴定多肽的n-末端氨基酸或用于多肽测序的用途,所述氨肽酶包含具有野生型克氏锥虫cruzipain或cruzain的25位的甘氨酸残基,65位的丝氨酸残基,138位的半胱氨酸残基和208位的组氨酸残基的氨基酸序列,其中所述氨肽酶的剩余氨基酸序列包含与seqidno.1中所示的所述野生型克氏锥虫cruzipain或seqidno.2中所示的cruzain具有至少75%,至少80%,至少81%,至少82%,至少83%,至少84%,至少85%,至少86%,至少87%,至少88%,至少89%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%,至少99%或100%序列同一性的序列。甚至更特别地,用于鉴定多肽的n-末端氨基酸或用于多肽测序的所述氨肽酶在野生型克氏锥虫cruzipain或cruzain的第一个甲硫氨酸残基后具有插入的半胱氨酸残基。甚至更特别地,用于鉴定多肽的n-末端氨基酸或用于多肽测序的所述氨肽酶包含seqidno.3或seqidno.4中所示的氨基酸序列或由seqidno.5或seqidno.6中所示的氨基酸序列组成。在特别的实施方案中,在单分子水平进行所述鉴定、分类或测序。甚至更特别地,通过所述多肽的c-末端进行表面固定。
在另一个特别的实施方案中,提供了工程化的、合成的或重组的氨肽酶用于鉴定多肽的n-末端氨基酸或用于多肽测序的用途,所述氨肽酶与seqidno.7中所示的野生型水生栖热菌氨肽酶t的氨基酸序列具有至少75%,至少80%,至少81%,至少82%,至少83%,至少84%,至少85%,至少86%,至少87%,至少88%,至少89%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%,至少99%或100%序列同一性,其中在所述野生型氨肽酶的第一个甲硫氨酸残基后插入半胱氨酸残基。在更特别的实施方案中,为鉴定或分类多肽的n-末端氨基酸或为多肽测序提供的所述氨肽酶t包含seqidno.8中所示的氨基酸序列或由seqidno.8中所示的氨基酸序列组成。在特别的实施方案中,在单分子水平进行所述鉴定、分类或测序。甚至更特别地,所述氨肽酶通过其c-末端进行表面固定。在其他实施方案中,所述氨肽酶是催化活性的氨肽酶。在最特别的实施方案中,使用所述水生栖热菌氨肽酶t鉴定的所述n-末端氨基酸选自leu、met、tyr、arg、pro、gly、lys、ala和val。
本申请的方法
本申请中所述的发明基于以下公开的几个方法。
在一个方面中,提供了鉴定或分类表面固定多肽的末端氨基酸的方法,所述方法包括:
a)将所述表面固定多肽接触裂解诱导剂,其中所述裂解诱导剂结合所述多肽的末端氨基酸并从所述多肽裂解末端氨基酸;
b)测量所述裂解诱导剂在所述末端氨基酸上的停留时间;
c)将所述测量的停留时间与所述裂解诱导剂和一组末端氨基酸特征性的一组参照停留时间进行比较;
来鉴定或分类所述末端氨基酸。
还提供了获得表面固定多肽的序列信息的方法,所述方法包括:
a)将所述表面固定多肽接触裂解诱导剂,其中所述裂解诱导剂结合所述多肽的末端氨基酸并从所述多肽裂解末端氨基酸;
b)测量所述裂解诱导剂在所述表面固定多肽的末端氨基酸上的停留时间;
c)通过将所述测量的停留时间与所述裂解诱导剂和一组末端氨基酸特征性的一组参照停留时间值进行比较来鉴定或分类所述末端氨基酸;
d)允许所述裂解诱导剂切除所述末端氨基酸;
e)重复步骤a)至d)一次或多次。
在所述方法中,通过光学、电学或等离子体测量所述停留时间。在特别的实施方案中,所述末端氨基酸组包含leu、met、tyr、arg、pro、gly、lys、ala和val或由其组成。
在各种实施方案中,所述方法中的所述裂解诱导剂是异硫氰酸酯或选自pitc、citc、spitc和叠氮基-pitc的itc类似物。在这样的情况中,所述停留时间是直至除去所述末端氨基酸的时长,并且通过将所述时长与针对不同氨基酸的一组参照值进行比较来鉴定所述末端氨基酸。
在其他实施方案中,所述方法中的所述裂解诱导剂时肽酶,更特别是催化肽酶,甚至更特别是催化活性的氨肽酶。在本申请的方法中使用氨肽酶时,所述多肽是通过其c-末端表面固定的。在特定的实施方案中,以上方法中测量所述裂解诱导剂在所述末端氨基酸上的停留时间的所述步骤是测量所述裂解诱导剂在末端氨基酸上直至所述表面固定多肽的末端氨基酸裂解的停留时间。
在整个本申请中,所述裂解诱导剂可以是氨基肽酶、itc或itc类似物。如本文已经讨论的,表面上固定的多肽应当变性,使得n-末端对于化学或酶裂解是可自由接近的(如果多肽是通过其c-末端固定的),而且也要避免位阻或所述裂解的干扰。因此,还提供了包括第一步多肽变性的本申请的方法。在这样的变性条件下,待使用的催化活性的氨肽酶应当能够抵抗变性条件。因此在这些情况中优选所述氨肽酶是嗜热和/或耐溶剂的氨肽酶。
在不同的实施方案中,提供了本文所述的方法,其中所述氨肽酶是本申请中公开的任一种氨肽酶,更确切地本文所述的来自克氏锥虫的cruzain和cruzipain肽酶或来自水生栖热菌的氨肽酶t中的任一种。在特别的实施方案中,提供了所述方法,其中所述n-末端氨基酸选自leu、met、tyr、arg、pro、gly、lys、ala和val。
在其他实施方案中,提供了本文所述的方法,其中所述n-末端氨基酸是衍生的,并且其中所述裂解诱导剂是能够裂解衍生的n-末端氨基酸的氨肽酶。在更特别的实施方案中,用citc或spitc衍生所述n-末端氨基酸,并且所述裂解诱导剂是本文公开的来自克氏锥虫的工程化cruzain或cruzipain中的任一种。
在本申请的各种实施方案中,本文针对从c-末端固定多肽鉴定或分类n-末端氨基酸或针对从所述多肽获得序列信息的方法是在单分子水平进行的方法。
对于单分子测量,设想了将来自本申请方法的多肽固定于活性传感表面上。在特别的实施方案中,所述活性传感表面是金表面或酰胺-、羧基-、硫醇-或叠氮化物官能化表面,所述多肽通过化学偶联于其上。
停留时间的多种测量和结合使用非裂解结合剂
在可替换的实施方案中,所述申请的方法中使用的氨肽酶可以是仅在n-末端氨基酸几轮结合和解开后裂解n-末端氨基酸的氨肽酶。所述氨肽酶的每个停留时间将提供测定直至n-末端氨基酸切除的停留时间的信息,并且可以帮助鉴定n-末端氨基酸。为了通过氨肽酶检测n-末端氨基酸身份变化的时间点以及在单分子设定中更准确地预测n-末端氨基酸,推荐对每个n-末端氨基酸进行多个测量。这可以通过使用发生实际裂解前将停靠(结合)和离开(解离)n-末端氨基酸几次的氨肽酶来实现。因此还设想了本申请的方法中测量催化活性氨肽酶的停留时间的步骤意味着在所述氨肽酶裂解n-末端氨基酸之前测量所述氨肽酶的多个停留时间。换句话说,提供了本申请的方法,其中针对所述氨肽酶与所述n-末端氨基酸的每个结合事件来测量所述催化活性氨肽酶的停留时间。在实施例13和图14中证明了以上内容。
在特别的实施方案中,提供了本申请中公开的方法,其中n-末端氨基酸的酶裂解中使用的氨肽酶在所述氨肽酶裂解n-末端氨基酸所需的时间窗中平均具有至少3个、至少4个、至少5个、至少6个、至少7个、至少8个、至少9个、至少10个、至少20个或至少50个结合/解离循环。这表示在有效裂解事件之间发生至少3个、至少4个、至少5个、至少6个、至少7个、至少8个、至少9个、至少10个、至少20个或至少50个非有效裂解结合/解离循环。
还提供了本申请的方法,其中将所述表面固定的多肽另外接触一种或多种末端氨基酸结合蛋白,其中所述一种或多种结合蛋白与所述末端氨基酸的结合事件的动力学鉴定了所述末端氨基酸。使用n-末端氨基酸结合蛋白的结合特异性来收集底物信息的可能性在理论上得到了rodriques等的证实(2018,biorxiv,doi:http://dx.doi.org/10.1101/352310)。在本申请的方法中另外使用所述不可裂解结合剂(挨着催化活性氨肽酶)可以提供额外的信息,以在单分子实验中较高准确性地预测或鉴定n-末端氨基酸。在特别的实施方案中,所述不可裂解的结合剂在裂解诱导剂裂解所述n-末端氨基酸所需的时间窗口中具有至少3个、至少4个、至少5个、至少6个、至少7个、至少8个、至少9个、至少10个、至少20个或至少50个与n-末端氨基酸的结合/解离循环。所述裂解诱导剂是氨肽酶或itc或itc类似物,更特别地,所述氨肽酶是本申请中所述的氨肽酶中的一种。
裂解的检测
本申请方法的其他部分之一是检测或确认末端氨基酸的裂解。因此,本文还提供了本申请的方法,另外包括通过测量表面固定多肽的光学、电学或等离子体信号来确定所述末端氨基酸的裂解的步骤,其中光学、电学或等离子体信号中的差异表示所述末端氨基酸的裂解。实际上,具有游离n-末端的固定肽具有几种性质,这些性质用于确定本发明的裂解诱导剂何时切除n-末端氨基酸。
在第一个实例中,游离的n-末端胺基团在宽范围的ph内带有正电荷。该正电荷与通过其固定肽的锚定点之间的距离可以例如通过以下方法测量,将肽固定在适当设计的检测器元件(碳纳米管、纳米级晶体管,如场效应晶体管,尤其是鳍片形场效应晶体管、栅极全能场效应晶体管、纳米带场效应晶体管等)上时,通过在电位检测中测量随机电报噪声(sorgenfrei等,2011nanolett11:3739-3743)。用本发明的裂解诱导剂裂解n-末端氨基酸时,带正电荷的n-末端氨基基团更靠近肽的锚定点,从而更接近检测器表面。在完全伸展的肽中,该电荷与锚定点之间的距离缩短的长度约为3.8埃(轮廓长度),这受肽主链中共价键的几何形状约束。因此,在肽二级结构破坏(高温、有机(共)溶剂暴露等)的环境条件下,氨基末端电荷和肽锚定点之间的长度测量值分布的最大值有一个上限,其受到肽主链中共价键几何结构的限制。通过在裂解诱导剂存在期间重复观察肽的氨基末端电荷来测量该最大长度的变化,可以揭示裂解诱导剂确实已经切除n-末端氨基酸的时间点。
在第二个实例中,可以使氨基末端氨基酸与试剂反应,使得形成氨基酸衍生物,其中末端氨基上的正电荷被消除,转化为带有一个或更多个负电荷的氨基酸衍生物,或从单个正电荷增加到多个带正电荷的氨基酸衍生物。例如,这可以通过将固定的肽与适当选择的不带电荷、带一个或多个正电荷或一个或多个负电荷的n-羟基琥珀酰亚胺试剂接触来实现。或者,电荷调节剂可以是裂解诱导剂本身,例如当固定的肽的末端氨基与适当选择的异硫氰酸酯试剂(如pitc、citc、spitc(4-磺基异硫氰酸苯酯)或叠氮基异硫氰酸苯酯)反应时,其中在该试剂与固定的肽接触之前,在该试剂与固定的肽接触期间或之后,可以通过叠氮化物基团上的点击化学进一步修饰后者。以此方式,携带氨基酸衍生物的肽与在切除了n-末端氨基酸衍生物之后该肽之间的电荷差变为二元(中性向正的转化、负向正的转化或多个正电荷向单个正电荷的转化),或增强了,或两者兼有。使用与实例1中类似的检测技术,可以通过检测电荷的这种变化来测量裂解诱导剂有效导致氨基酸裂解的时间。
在第三个实例中,n-末端氨基酸的氨基基团或其侧链可以与赋予光谱学上可区分的特性的试剂反应,从而生成可以用光谱学方法(如荧光法、拉曼光谱法),等离子体共振等检测的n-末端氨基酸衍生物。特别是,使用全内反射荧光(tirf)显微镜进行单分子检测是一种优选方法,因为它旨在检测可以在其上固定肽的并列于反射表面(例如玻璃)上的薄层中的荧光。接触裂解诱导剂(其可以例如是氨肽酶、edmanase或含异硫氰酸酯的分子)时,可以通过固定肽的观察时间序列中的光谱性质变化来检测裂解发生的时间,例如,由于切除了荧光标记的n-末端氨基酸衍生物,导致荧光信号丢失。或者,当固定的肽含有合适的fret供体或受体且n-末端氨基酸衍生物含有匹配的fret受体或供体时,可以观察到
在再另一个实施方案中,通过检测肽结合剂(例如催化活性或无活性氨肽酶或edmanase)与固定肽的结合亲和力或结合动力学的变化,可以检测直至被裂解诱导剂裂解的时间。例如,可以使用上述任一种技术来测量肽结合剂的停留时间(结合和解离之间的时间)。可以测量肽结合剂的几个这样的结合和解离循环,并且可以在n-末端氨基酸裂解时检测平均停留时间的变化。在一个特定的实例中,用于检测的肽结合剂比裂解诱导剂诱导n-末端氨基酸裂解所需的时间具有更快的结合/解离动力学,使得通常在n-末端氨基酸的两个裂解事件之间可以观察到肽结合剂对固定肽的多个结合/解离测量点。在一个特定的实例中,肽结合剂与裂解诱导剂相同。例如,催化活性的氨肽酶具有有效裂解和非有效裂解的缔合/解离循环。通过测量非有效裂解肽结合事件期间氨肽酶随时间的亲和力,尤其是结合/解离动力学,当n-末端氨基酸被切除并且新的n-末端氨基酸因此呈现用于氨肽酶相互作用时,观察到这些性质发生了变化。在特定的实施方案中,在远离酶催化速率的最佳条件的条件下使用氨肽酶,使得氨肽酶与固定肽结合的大多数结合/解离事件是非有效的裂解,并且将这些结合事件的时间序列中的这些事件的动力学变化用于告知氨肽酶切除n-末端氨基酸的时间。
再在其他方面中,提供了一种通过确定氨肽酶在n-末端氨基酸上的“工作时间”值或停留时间在单分子水平鉴定或分类通过其c-末端固定于表面的多肽的n-末端氨基酸的方法,所述方法包括将所述表面固定多肽接触氨肽酶并测量所述氨肽酶的“工作时间”值或停留时间。
在特别的实施方案中,所述氨肽酶是本申请所述的任一种氨肽酶。因此,提供了所述方法,其中所述氨肽酶是工程化的、合成的或重组的氨肽酶,其包含具有野生型克氏锥虫cruzipain或cruzain的25位的甘氨酸残基,65位的丝氨酸残基,138位的半胱氨酸残基和208位的组氨酸残基的氨基酸序列,其中所述氨肽酶的剩余氨基酸序列具有与seqidno.1中所示的所述野生型克氏锥虫cruzipain或seqidno.2中所示的cruzain的氨基酸序列具有至少75%,至少80%,至少81%,至少82%,至少83%,至少84%,至少85%,至少86%,至少87%,至少88%,至少89%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%,至少99%或100%序列同一性的序列。甚至更特别地,为所述方法提供的所述氨肽酶在野生型克氏锥虫cruzipain或cruzain的第一个甲硫氨酸残基后具有插入的半胱氨酸残基。甚至更特别地,所述氨肽酶包含seqidno.3或seqidno.4中所示的氨基酸序列或由seqidno.5或seqidno.6中所示的氨基酸序列组成。在另一个特别的实施方案中,提供了第六个方面的方法,其中所述氨肽酶与seqidno.7中所示的野生型水生栖热菌氨肽酶t的氨基酸序列具有至少75%,至少80%,至少81%,至少82%,至少83%,至少84%,至少85%,至少86%,至少87%,至少88%,至少89%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%,至少99%或100%序列同一性,其中在所述野生型氨肽酶的第一个甲硫氨酸残基后插入了半胱氨酸。在更特别的实施方案中,所述氨肽酶t包含seqidno.8中所示的氨基酸序列或由seqidno.8中所示的氨基酸序列组成。在其他实施方案中,所述氨肽酶是催化活性的氨肽酶。在最特别的实施方案中,所述待鉴定或分类的n-末端氨基酸选自leu、met、tyr、arg、pro、gly、lys、ala和val。
在更特别的实施方案中,n-末端氨基酸是衍生的,并且所述氨肽酶是包含针对所述衍生的n-末端氨基酸的结合结构域并且裂解所述衍生的n-末端氨基酸的氨肽酶。在甚至更特别的实施方案中,所述衍生的n-末端氨基酸是citc-或spitc-衍生的氨基酸并且所述氨肽酶能够结合并裂解所述citc-和spitc-衍生的氨基酸。对于后一个目的,所述氨肽酶是特意工程化的。这样工程化的结合并裂解citc-或spitc衍生的n-末端氨基酸的氨肽酶的非限制性实例是本申请工程化的以具有25位的甘氨酸残基,65位的丝氨酸残基,138位的半胱氨酸残基和208位的组氨酸残基的克氏锥虫cruzipain或cruzain。
再在另一个方面中,提供了在单分子水平将表面固定的多肽测序的方法,所述方法包括a)将所述表面固定的肽接触氨肽酶,更特别地催化活性的氨肽酶;b)测量所述氨肽酶的酶“工作时间”值;c)通过所述“工作时间”值鉴定或分类所述n-末端氨基酸;重复步骤a)至步骤c)一次或多次。在特别的实施方案中,所述氨肽酶是本申请所述的任一种氨肽酶。因此,提供了本申请的方法,其中所述氨肽酶是工程化的、合成的或重组的氨肽酶,其包含具有野生型克氏锥虫cruzipain或cruzain的25位的甘氨酸残基,65位的丝氨酸残基,138位的半胱氨酸残基和208位的组氨酸残基的氨基酸序列,其中所述氨肽酶的剩余氨基酸序列包含与seqidno.1中所示的所述野生型克氏锥虫cruzipain或seqidno.2中所示的cruzain的氨基酸序列具有至少75%,至少80%,至少81%,至少82%,至少83%,至少84%,至少85%,至少86%,至少87%,至少88%,至少89%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%,至少99%或100%序列同一性的序列。甚至更特别地,所述氨肽酶在野生型克氏锥虫cruzipain或cruzain的第一个甲硫氨酸残基后具有插入的半胱氨酸残基。甚至更特别地,所述氨肽酶包含seqidno.3或seqidno.4中所示的氨基酸序列或由seqidno.5或seqidno.6中所示的氨基酸序列组成。在另一个特别的实施方案中,提供了第七个方面的方法,其中所述氨肽酶是工程化的、合成的或重组的氨肽酶,其与seqidno.7中所示的野生型水生栖热菌氨肽酶t的氨基酸序列具有至少75%,至少80%,至少81%,至少82%,至少83%,至少84%,至少85%,至少86%,至少87%,至少88%,至少89%,至少90%,至少91%,至少92%,至少93%,至少94%,至少95%,至少96%,至少97%,至少98%,至少99%或100%序列同一性,其中在所述野生型氨肽酶的第一个甲硫氨酸残基后插入了半胱氨酸。在更特别的实施方案中,所述氨肽酶t包含seqidno.8中所示的氨基酸序列或由seqidno.8中所示的氨基酸序列组成。在最特别的实施方案中,来自所述方法的所述n-末端氨基酸选自leu、met、tyr、arg、pro、gly、lys、ala和val。
在更特别的实施方案中,n-末端氨基酸是衍生的,并且所述氨肽酶包含针对所述衍生的n-末端氨基酸的结合结构域并且裂解所述衍生的n-末端氨基酸。在甚至更特别的实施方案中,所述衍生的n-末端氨基酸是citc-或spitc-衍生的氨基酸,并且所述氨肽酶能够结合并裂解所述citc-和spitc-衍生的氨基酸。在其他实施方案中,所述氨肽酶是催化活性氨肽酶。
在另一个实施方案中,通过光学、电学或等离子体测量所述在单分子水平将表面固定多肽测序的方法中的“工作时间”值。
在本申请最特别的实施方案中,在蛋白质变性条件下进行本文所述的方法。所述蛋白质变性条件通过高温或通过溶解的存在来获得。在特别的实施方案中,所述高温是40℃至120℃或50℃至100℃或60℃至100℃或70℃至90℃的温度。在特别的实施方案中,所述溶剂选自乙酸、三氯乙酸、磺基水杨酸、碳酸氢钠、乙醇、醇、交联剂(如甲醛和戊二醛)、离液剂,如脲、氯化胍或高氯酸锂,以及破坏二硫键的试剂,如2-巯基乙醇、二硫苏糖醇或三(2-羧乙基)膦。最特别地,所述溶剂是乙腈、乙醇或甲醇。
在本申请其他最特别的实施方案中,所述裂解诱导剂是共价裂解诱导剂。在本申请其他最特别的实施方案中,所述不可裂解结合剂是共价不可裂解结合剂。
以下实施例旨在促进进一步理解本发明。尽管已经参照所说明的实施方案描述了本发明,但应当理解本发明不限于此。本领域普通技术人员根据本文的教导将认识到其范围内的其他改变和实施方案。因此,本发明仅受限于本文所附的权利要求。
实施例
havranek和borgo的肽测序依赖于naab的使用,naab是无催化活性的氨肽酶或t-rna合成酶。作为传统的edman降解裂解n-末端氨基酸的替代方案,产生了edmanase酶(wo2014273004)。来自克氏锥虫的这种cruzain半胱氨酸蛋白酶在四个位置进行了修饰,以能够结合(并裂解)pitc衍生的n-末端氨基酸。首次引入的点突变,用甘氨酸取代了25位的催化半胱氨酸(c25g),使该酶失去催化能力,除非它被pitc衍生的肽底物的硫原子拯救。需要另外三个突变,即g65s、a138c和l160y,来提高pitc衍生的底物的适应性。虽然havranek和borgo使用无催化活性的naab进行氨基酸鉴定,并使用edmanase酶在鉴定后裂解n-末端氨基酸,但我们已经开发了一种方法,该方法令人惊讶地不需要无催化活性的naab。该方法完全依赖于工程化的氨肽酶,从其停留时间可告知其结合和裂解的n-末端氨基酸。本文公开的酶对n-末端(衍生的)氨基酸具有亲和力,而与n-末端氨基酸的身份无关。但是,为了获得肽序列信息,这些酶根据n-末端氨基酸的身份显示出催化效率和停留时间的变化(图1-2)。甚至更令人惊讶地,我们证明了一种鉴定n-末端氨基酸的方法,该方法包括仅使itc或itc类似物在所述n-末端氨基酸上的停留时间相关。
实施例1:用于单肽检测的tirf显微镜检查
第一步,开发了一种系统来固定待在表面上测序的肽。将叠氮化物官能化的、用烤箱清洁的玻璃板用作表面,并且将n-末端为dbco-peg8基团且c末端为磺基-cy5荧光探针的肽nnggnnggrgnk用作测试肽。通过叠氮化物-dbco点击反应将测试肽固定。将叠氮化物官能化的玻璃板置于1ml1nm测试肽的顶部,并在黑暗中孵育24h。为了官能化,使用11-叠氮基-十一烷基(三甲氧基)硅烷,其使玻璃表面疏水,允许玻璃漂浮在液体上。24h后,将玻璃板用1mlms级水洗涤3次(每次洗涤30分钟)。对于对照样品,将玻璃板用水孵育。在zeisstirf显微镜上进行显微镜检查,并在λem639nm处拍照。肽成功地固定在玻璃表面上,并以1nm的浓度获得了足够的空间分布(图3)。
实施例2:表面固定的肽的胰蛋白酶消化
接下来,优化酶促裂解表面固定的肽的条件。再次使用测试肽dbco-peg8-nnggnnggrgnk-cy5,但现在与胰蛋白酶一起使用。在精氨酸切割后检测到成功的酶表面反应,其除去了荧光探针。将叠氮化物官能化、烤箱清洁的玻璃板放在1ml1nm测试肽的顶部,并在黑暗中孵育24小时。用ms级水洗涤3次后,将玻璃板与100nm胰蛋白酶(测序级,promega)在室温下孵育1h。对照用水孵育。胰蛋白酶处理后,将板再次用水洗涤3次。在存在1μmdbco-peg8-酰胺钝化剂(在24h叠氮化物-dbco点击反应期间添加到测试肽中)的情况下,也重复了该实验,以评估其对聚类和非特异性胰蛋白酶表面相互作用的影响。
在没有钝化剂的情况下,胰蛋白酶反应不成功,这是由于来自胰蛋白酶在表面上的非特异性结合的高背景信号引起的(图4a)。在561nm的较低λem通道(cy3通道)中评估了背景。存在钝化剂时,背景消失,并且相当大量的固定肽被裂解,如斑点的明显减少所见(图4b)。
实施例3:肽n-末端反应探针
根据所用的氨肽酶(即结合衍生的n-末端氨基酸或结合未标记的n-末端氨基酸,参见实施例4及更多),固定的肽应被标记或不标记。探针的选择将取决于读出策略(图2)。但是,必须仔细选择探针:探针必须针对肽n-末端具有反应性,并且衍生的肽底物必须适合酶的催化位点(有关酶工程化,参见实施例4)。潜在的荧光探针候选物是荧光胺、邻苯二甲醛、丹酰氯和香豆素异硫氰酸酯(citc)衍生物。对于带电荷的探针,令人感兴趣的候选物是磺基异硫氰酸苯酯(spitc),它是pitc探针的带负电荷的变体,用于ms从头肽测序中以中和n-末端片段离子(samyn等2004jamsocmassspectrom15:1838-1852)。尽管可以通过小心地控制ph值来提高对n-末端伯胺的特异性(与赖氨酸伯ε-胺相反),但仍需要考虑一定程度的非特异性,尤其是使用胰蛋白酶肽时。在此,肽用citc或spitc来衍生。
实施例4:edmannase再工程化
为了寻找一种能够结合并裂解任何肽的任何n-末端氨基酸的氨肽酶,探索了两条平行的研究路线:一项是使用来自克氏锥虫的再工程化的cruzipain/cruzain半胱氨酸蛋白酶,而另一条是关于水生栖热菌氨肽酶t的使用。
我们首先再工程化了克氏锥虫的cruzain和cruzipain半胱氨酸蛋白酶,使其能够结合衍生的n-末端氨基酸,更准确地结合citc衍生的和spitc衍生的n-末端氨基酸。荧光或带电异硫氰酸酯探针在cruzain或cruzipain酶上的计算对接(autodock-vina;trott和olsen2010jcomputchem31:455-461)以及虚拟突变体提供了几种潜在的再工程化选择。将tyr160恢复自然(renatived)的leu时,spitc-ala-phe成功地停靠在所述cruzain和cruzipain上(图5a)。附近的glu208可能会与spitc的硫酸根(sulphate)发生电荷冲突,但可以通过将实际残基突变成his来避免。通过这种方式,开发了一种再工程化的克氏锥虫cruzain和cruzipain,其包含四个点突变(即c25g、g65s、a138c、e208h),其上带有荧光3-citc-ala-phe及其5-、6-、7和8香豆素类似物,可在活性位点的凹槽内产生总体正确的对接姿势(图5b)。
实施例5.用克氏锥虫cruzipain_pet24b(+)转化大肠杆菌bl21(de3)
在下一步中,重组生产了工程化的cruzipain(c25g、g65s、a138c、e208h)。将编码再工程化的cruzipain密码子优化的dna序列克隆到pet24b(+)质粒(ndei-bamhi)中。在蛋白质的n-末端添加了cys,并在c-末端添加了6xhis。全长蛋白质的分子量为±37.5kda,估计pi为5.7。其序列描述于seqidno.5中:
获得了再工程化的cruzipain的成功转化(图6a)。接下来,用合成的spitc-和citc-衍生的7-氨基-4-甲基香豆素(amc)氨基酸类似物分析了再工程化的cruzipain的动力学。一旦去除spitc-或citc-衍生的n-末端氨基酸时,释放并检测了荧光amc。然后使用计算机模拟,将动力学数据用于评估使用酶“工作时间”值对肽进行测序的可行性。
实施例6:用水生栖热菌氨肽酶t_pet24b(+)转化大肠杆菌bl21(de3)
合成了编码水生栖热菌的氨肽酶t(从这开始称为taq-apt或taqapt)的密码子优化的dna序列。随后将taq-apt基因克隆到pet24b(+)质粒(ndei-bamhi)中(图9)。在蛋白质的n-末端添加了cys,并在c-末端添加了6xhis。全长蛋白质的分子量为±45.7kda,估计pi为5.6。其序列描述于seqidno.8中:
获得了再工程化的taq-apt的成功转化(图6b)。
实施例7:taq-apt表达和纯化
验证并优化了taq-apt在大肠杆菌bl21(de3)转化子中的重组表达。在此,(嗜热)蛋白的纯化通过ni-nta离心柱或热处理或两者结合进行测试。将bl21(de3)pet-24b(+)-taq-apt转化细胞的等分试样加入5mllb+50μg/ml卡那霉素中,在37℃下生长过夜,并在5ml新鲜lb+50μg/ml卡那霉素中稀释100倍。在37℃下孵育2h后,用1mmiptg进行诱导,然后将培养物在28℃下生长过夜。收集细胞(4,000xg,10分钟),重悬于1mlpbs+10mm咪唑(ph7.5-8)中,超声处理(1s开,1s关,90秒,30%振幅)并全速离心5分钟。收集上清液并分成4×200μl,用于测试4种不同条件:(a)原始可溶性级分,(b)ni-nta纯化,(c)热处理和(d)ni-nta纯化+热处理。处理后,将样品用10kda离心柱浓缩至约50μl的体积。从这将10μl与10μlsds样品缓冲液混合,并用sds-page分析样品(图8)。有关ni-nta纯化方案的详细信息,参见实验程序。通过加热的纯化在80℃下进行30分钟。
ni-nta和加热的结合产生了可接受的纯taq-apt提取物(图7)。用l-亮氨酸-对硝基苯胺(140μlpbs缓冲液,10μlmeoh中的24mml-亮氨酸-对硝基苯胺和10μl纯化的tag-apt)进行的肽酶分析证实了活性(氨基)肽酶的存在,具有嗜热性能(70℃时taq活性最佳)(图8)。
实施例8.水生栖热菌氨肽酶t的动力学参数的测定
用对硝基苯胺测试确定了水生栖热菌氨肽酶t裂解不同氨基酸底物的动力学参数。底物由n末端氨基酸与其c-末端连接的对硝基苯胺组成。氨基酸裂解后,可以通过测量405nm的吸光度来监测游离的硝基苯胺。将seqidno8中所述的水生栖热菌氨肽酶t以2.0625μm的浓度(在pbs中)添加到不同浓度的氨基酸对硝基苯胺底物(pbs中0.0625、0.125、0.25、0.5、1和2mm)中。随后,使用fluostaromega微平板阅读器(mbglabtech)在40℃下连续测量对硝基苯胺释放。由此得出在每种底物浓度下的反应初始速度(v0)。对于每种氨基酸,生成一个lineweaver-burke图,从中可以确定反应vmax和酶-底物km。从vmax和酶浓度计算周转数kcat(kcat=vmax/[e])。然后通过从kcat值取倒数来计算酶的工作时间值。表1中列出了9种氨基酸的动力学参数。表1中所示的“工作时间”值以1/kcat计算,并且因此是酶溶液在肽上直至催化发生所需的总时间。
表1.使用九种不同氨基酸的对硝基苯胺测定的动力学参数,包括水生栖热菌氨肽酶t针对所述九种不同氨基酸的工作时间。
在低于酶的最佳温度的40℃下进行测定。更接近最佳温度70℃的工作加快了反应速度(kcat和酶工作时间)。总而言之,我们令人惊讶地发现了本文所述的水生栖热菌氨肽酶t能够以不同的动力学结合并裂解九种不同的氨基酸。此外,反应的动力学与氨基酸的身份有关,甚至更令人惊讶的是,不同氨基酸的kcat值的分布允许彼此区分并因此鉴定不同的氨基酸。此处显示的结果不仅证实了本文公开的氨肽酶的效用,而且巩固并证实了本申请中公开的方法和用途。
实施例9.水生栖热菌氨肽酶t在40℃和80℃下针对不同氨基酸对硝基苯胺底物的活性
taqapt酶在当前测试组中显示出针对所有氨基酸对硝基苯胺底物的活性。如本文所述并且与当前申请中描述的发明一致,活性在不同氨基酸之间不同。在80℃下,可以将氨基酸底物组大致分为快裂解(l、m、y、r、f)和慢裂解(d、p)底物。然而,在40℃下,组似乎分为快裂解(y、r)和慢裂解(d、p),以及介于其中的底物(l、m、f)。在40℃下,根据n-末端氨基酸,活性大约降低10到3倍。最后,taqapt不仅在对硝基苯胺测定中具有活性。更重要地,发明人证明了taqapt也裂解了肽底物。例如在图11中显示,taqapt裂解了二肽,即使在第二位具有脯氨酸。来自在第二位具有脯氨酸的肽的n-末端氨基酸不容易被裂解,因为与氨基酸脯氨酸相邻的肽键对大多数肽酶都有抵抗力(iyver等,2015febsopenbio.2015年4月2日;5:292-302)。与minagawa等(1988agriculturalandbiologicalchemistry52:1755-1763)中所述相反,我们在此表明,即使脯氨酸在第二位置,水生栖热菌氨肽酶t也可以裂解n-末端氨基酸。这一令人惊讶的发现大大增强了taq-apt在本申请公开的方法和用途中以及在通常的单分子肽测序中的通用用途。
实施例10.来自水生栖热菌的氨肽酶t的有机溶剂耐受性
对于表面固定多肽的单肽检测,至关重要的是将所述多肽完全变性并且不再具有任何二级结构。本领域公知的是这可以通过溶剂(如甲醇)或高温来实现。当需要这些苛刻条件时,所用的氨肽酶应能耐受溶剂和/或高温。除了证明taqapt在80℃的温度下具有活性外,我们还研究了本文所述的来自水生栖热菌的氨肽酶是否耐受有机溶剂。如此处所示,直到50%的甲醇,33%的乙腈和33%的乙醇,来自水生栖热菌的氨肽酶t仍保持完全活性,这表明该酶对有机溶剂具有相当的耐受性(图12a)。在较高的有机溶剂浓度下,尽管较低,但仍可检测到活性。在0%甲醇对50%甲醇中用圆二色性(cd)分析酶时,未观察到结构差异(图12b)。此外,该酶似乎在去离子(ms级)水中具有完全活性,这在酶与超灵敏芯片技术(例如电生物传感器,如场效应晶体管)一起使用时可能是有利的(图12c)。
实施例11.来自水生栖热菌的氨肽酶t的位点特异性n-末端标记
在还原剂tcep(10mm)存在下,将含有额外n-末端半胱氨酸的重组taq氨肽酶t与荧光马来酰亚胺-dylight650探针孵育过夜。以等摩尔浓度以及10倍、100倍和1000倍过量摩尔浓度添加马来酰亚胺-dylight650。用sds-page分离出氨肽酶后,使用考马斯染色以及荧光法对其进行可视化,以评估dylight650标记的蛋白质。图13a显示氨肽酶被dylight650标记。此外,l-亮氨酸-对硝基苯胺测定法证明了标记氨基肽酶不会损害其功能(图13b)。
实施例12.氨肽酶标记
为了测序步骤的读取,并因此检测酶的“工作时间”值,使用了两个传感器选项:光学和电位计(图2)。实施例11中显示了氨肽酶的光学标记。使用荧光探针的可替代光学读出策略可以在氨肽酶以及在肽底物上进行,以测量荧光共振能量转移(fret)的工作时间和检测成功的裂解事件。
考虑到该概念是基于单分子的,因此需要酶的可预测且特异性的标记。为了合理覆盖人类蛋白质组,需要多达109个读数(例如10,000个表达的蛋白,动态范围为104,并且至少需要丰度最低蛋白的10个读数)(geiger等,2012molcellproteomics11:m111.014050)。可以使用零模式波导(zmw)来完成单分子检测,如根据rhoads和au(2015genomicsproteomicsbioinformatics13:278-289)用于单分子dna测序中的那些。另一方面,电位计读数需要带电探针,以影响场效应晶体管(fet)电势。
对于位点特异性标记的氨肽酶,需要在酶n-末端进行一步化学修饰。如macdonald等(2015natchembiol11:326-331)所述,该酶在n-末端伯胺上被吡啶羧醛衍生物特异性修饰。当不存在其他(表面暴露的)半胱氨酸时,用醛类探针或用马来酰亚胺类探针特异性地修饰两种氨肽酶(参见实施例5和6)中添加的半胱氨酸(gunnoo和madder2016chembiochem17:529-553)。对于电位计工作时间测量,是否需要向酶添加电荷取决于其等电点(pi)。如果需要,可以通过(位点特异性)修饰引入带电荷的探针,或通过单侧中和带正电或带负电的残基(例如赖氨酸甲酰化)来改变酶的净电荷。
实施例13.通过测量氨肽酶停留时间的单分子肽n-末端氨基酸鉴定或分类
在独立的实验装置中进一步验证了水生栖热菌氨肽酶t的使用。固定一系列除了n-末端氨基酸以外具有相同一级结构的合成肽底物,然后使用tirf显微镜测量来自水生栖热菌的标记的氨肽酶t的停留时间。肽底物具有以下整体结构:x-dggnnggk(fluo)ggk(dbco/mal/nhs),其中c-末端赖氨酸的侧链连接有dbco、马来酰亚胺或n-羟基琥珀酰亚胺基团,用于将底物固定在表面上。第二赖氨酸在其侧链上具有连接的荧光基团,用于精确定位该表面上的单分子底物。n-末端将具有可变的氨基酸或可变的氨基酸序列(x),其前面为作为反应制动器的天冬氨酸残基,这是考虑到来自水生栖热菌的氨肽酶t对该残基的活性非常低。
固定肽底物并确定视野中的单分子底物位置后,添加荧光标记的氨肽酶,并测量底物位置上的连续酶停留时间。考虑到酶-底物结合动力学(km)和底物裂解动力学(kcat)都取决于n-末端氨基酸的身份,通过测量酶-底物“结合-解离”事件的数量和直至底物裂解的总时间,得到n-末端氨基酸的身份或将n-末端氨基酸的身份分类(图14)。底物裂解的验证源自裂解前后酶-底物“结合-解离”事件频率的可测量变化(图14下)。在远低于最佳温度下使用嗜热氨基肽酶(例如水生栖热菌氨肽酶t)时,裂解动力学会显著降低,从而导致酶底物“结合-解离”事件的数量增加。
如本申请的详细描述中所述,单个氨基肽酶可用于捕获n-末端氨基酸上的“结合-解离”事件,以及用于裂解n-末端氨基酸。替换地,也可以使用两种不同的氨肽酶的组合,或者氨肽酶与化学n-末端氨基酸结合剂/裂解剂的组合(参见描述)。
实施例14.edman降解反应动力学取决于n-末端氨基酸残基
接下来,我们令人惊讶地发现了edman降解化学适用于单分子水平的固定肽的测序。首先,蛋白质通过其c-末端固定在表面上,然后通过edman降解化学连续切除氨基酸。尽管edman试剂的n-末端偶联与n-末端氨基酸的身份无关,但裂解反应的速度取决于它。因此,通过监测每个随后的n-末端氨基酸的裂解反应时间,可以获得序列信息。为了规避n-末端修饰的问题,首先对固定的蛋白进行蛋白水解(例如,用胰蛋白酶),然后在c-末端多肽上留下游离的、可接近的n-末端。通过使用可追踪的itc试剂,可以监测化学反应时间。例如,磺基异硫氰酸苯酯带有负电荷,可用于电学测量中。或者可以使用叠氮基异硫氰酸苯酯,其可以通过点击化学(带电基团,荧光探针)进行标记。最后,反应可以在高有机溶剂中进行,使结构彻底变性,并允许控制反应。
为了检查edman试剂4-磺基异硫氰酸苯酯(spitc)在不同氨基酸对硝基苯胺底物上的自发裂解活性,将5μl24mm氨基酸对硝基苯胺底物(在甲醇中)和240mmspitc(在水中)加入70μl300mm三乙醇胺(在50%乙腈中,ph9)中,并在40℃下孵育30分钟。终点活性测量将测试的氨基酸底物分为快裂解底物(l、m、y、r、f)和慢裂解底物(d、p)(图16a)。时间动力学分析显示了同一组底物的反应动力学差异(图16b)。重要地,在经典edman降解反应的裂解步骤过程中通常不使用的条件下也观察到了这种自发裂解活性。在经典edman降解中,itc偶联在弱碱性条件(吡啶、三甲胺、n-甲基哌啶)下实现,在酸性条件下(三氟乙酸)氨基酸裂解。在此,itc偶联和氨基酸裂解均在弱碱性条件下(三乙醇胺)完成。
实验程序
大肠杆菌的转化
在ice(neb)上解冻化学感受态大肠杆菌bl21(de3)细胞,添加100ng质粒dna并在冰上放置30min。在42℃的温水浴中孵育1.5min,然后放在冰上10min。向小瓶中加入1mllb培养基,并用胶带将其固定摇床上(laf),在37℃下1h。涂在lb-kan琼脂平板上(50μg/ml卡那霉素),并在37℃下生长过夜。
培养物挑选
从平板中挑选菌落(平板保存在4℃下),然后将菌落添加到10ml液体tb培养基+50μg/mlkan中。在37℃下生长过夜,准备500μl等分试样(+500μl甘油)并在-80℃下保存。
克隆
为了克隆,使用了pet-24b(+)质粒(图9)。
ni-nta纯化
将200μl裂解液(在pbs+10mm咪唑中)加到预先平衡的qiagenni-nta离心柱上,并在100×g下离心5分钟。用500μlpbs+20mm咪唑洗涤离心柱3次,并用500μlpbs+250mm咪唑洗脱。
热处理
在80℃下加热样品30min,并在全速下离心10min。
- 还没有人留言评论。精彩留言会获得点赞!