通过具有组成型转录激活因子MAL等位基因的酵母增加乙醇生产的制作方法
本发明的组合物和方法涉及具有组成型转录激活因子mal等位基因的经修饰的酵母。酵母产生的乙醇量增加,该增加似乎不是麦芽糖代谢的结果。此类酵母对于来自淀粉底物的大规模乙醇生产特别有用。
背景技术:
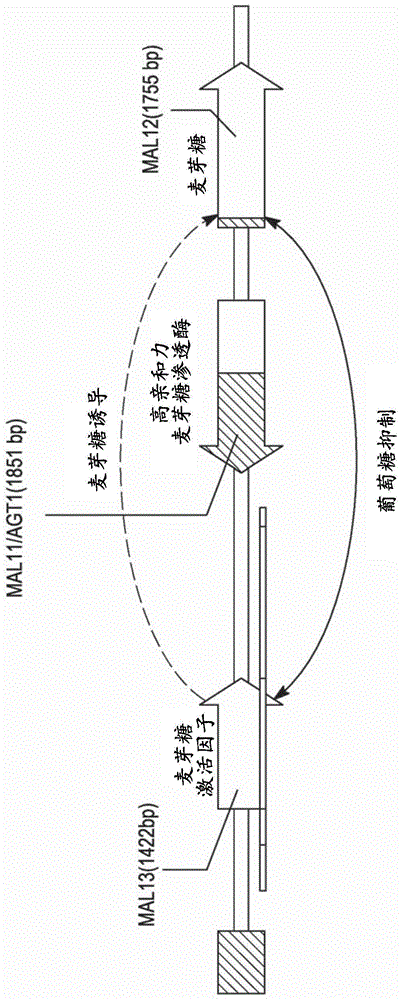
:第一代基于酵母的乙醇生产将糖转化为燃料乙醇。全世界每年通过酵母生产的燃料乙醇约900亿升(gombert,a.k.和vanmaris.a.j.(2015)curr.opin.biotechnol.[生物技术新见]33:81-86)。据估计,乙醇生产成本的约70%是原料。因为生产量如此之大,所以甚至小的产率提升将对于该产业也具有巨大的经济影响。酵母,例如酵母属(saccharomyces)能够代谢许多单糖和双糖,包括麦芽糖。酵母属中的麦芽糖发酵涉及五个似乎冗余的、未连接的mal基因座(称为mal1、mal2、mal3、mal4和mal5)中的至少一个(参见,例如,needleman,r.b.(1991)mol.microbiol.[分子微生物学]9:2079-84)。如图1中所示,每个基因座包括三个基因,编码(i)麦芽糖渗透酶、(ii)麦芽糖酶和(iii)转录激活因子(kim,j.和michels,c.a.(1988)curr.genet.[当代遗传学]14:319-23以及cheng,q.和michels(1989)genetics[遗传学]123:477-84。所述基因常规分别编号为1、2和3,使得例如在mal2基因座的基因分别编号为mal21、mal22和mal23。mal基因的转录受麦芽糖的诱导且受葡萄糖的抑制;然而,已在所有的mal基因座鉴定了组成型突变(winge,o.和roberts,c.(1950)c.r.trav.lab.carlsbergser.physiol.25:35-81,kahn,n.a.和eaton,n.r.(1971)mol.gen.genet.[分子遗传学与普通遗传学]112:317-22;charronm,j.和michels,c.a.(1987)genetics[遗传学]11623-31;zimmerman和eaton,n.r.(1974)mol.gen.genet.[分子遗传学与普通遗传学]134261-271;rodicio,r.(1986)curr.genet.[当代遗传学]11:235-41和tenberge,a.m.a.等人(1973)mol.gen.genet.[分子遗传学与普通遗传学]125:139-46。麦芽糖在商业规模的淀粉水解物中以低水平存在,其典型地为一种不希望的dp2组分,不太可能是额外乙醇的重要来源。此外,mal基因的转录受到高水平葡萄糖的抑制,使得酵母难以在高糖条件下利用麦芽糖(例如在燃料乙醇生产期间存在的麦芽糖)。有意表达麦芽糖代谢酶实际上可能会减缓葡萄糖代谢和废碳,考虑到乙醇生产商的需求,这是不可接受的。技术实现要素:本发明的组合物和方法涉及具有组成型转录激活因子mal等位基因的经修饰的酵母。虽然此类酵母可能能够代谢麦芽糖,即使在葡萄糖存在的情况下,所述酵母似乎产生的乙醇量增加,其不一定是麦芽糖代谢的结果。所述组合物和方法的方面和实施例描述于以下独立编号的段落中。1.在一方面,提供了一种用于增加来自淀粉水解物发酵产生的醇量和/或增加来自淀粉水解物发酵的醇产生速率的方法,所述方法包括用具有组成型转录激活因子mal等位基因的经修饰的酵母发酵所述淀粉水解物,其中与其他方面相同的亲本酵母产生的乙醇量相比,在经修饰的酵母中,在发酵结束时产生的乙醇量增加,或经一个时间段产生的乙醇量增加。2.在如段落1中所述的方法的一些实施例中,基于通过麦芽糖代谢途径的碳通量,至少一部分增加的乙醇量不能归因于麦芽糖发酵。3.在如段落1或2中所述的方法的一些实施例中,发酵结束时所述淀粉水解物中的麦芽糖水平与发酵开始时所述淀粉水解物中的麦芽糖水平大致相同。4.在如前述段落中任一项所述的方法的一些实施例中,发酵开始时所述淀粉水解物中的麦芽糖量不超过10g/l。5.在如前述段落中任一项所述的方法的一些实施例中,所述具有组成型转录激活因子mal等位基因的酵母进一步包含遗传改变,所述遗传改变引入编码磷酸转酮酶途径中的多肽的多核苷酸。6.在如前述段落中任一项所述的方法的一些实施例中,所述具有组成型转录激活因子mal等位基因的酵母进一步包含甘油途径和/或乙酰辅酶a途径中的改变。7.在如前述段落中任一项所述的方法的一些实施例中,所述具有组成型转录激活因子mal等位基因的酵母进一步包含编码dls1的基因的破坏。8.在如前述段落中任一项所述的方法的一些实施例中,所述具有组成型转录激活因子mal等位基因的酵母包含编码碳水化合物加工酶的外源基因。9.在如前述段落中任一项所述的方法的一些实施例中,所述酵母是酵母属(saccharomyces)物种。10.在另一方面,提供了一种经修饰的酵母,其包含组成型转录激活因子mal等位基因以及与麦芽糖代谢无关的至少一种额外的遗传修饰,当在淀粉水解物中生长时,在发酵期间,与其他方面相同的亲本酵母产生的量相比,所述酵母在发酵结束时产生的乙醇量增加。11.在如段落10中所述的经修饰的酵母的一些实施例中,所述酵母进一步包含遗传改变,所述遗传改变引入编码磷酸转酮酶途径中的多肽的多核苷酸。12.在如段落10或11中所述的经修饰的酵母的一些实施例中,所述酵母进一步包含在甘油途径和/或乙酰辅酶a途径中的改变。13.在如段落10-12中任一项所述的经修饰的酵母的一些实施例中,所述酵母进一步包含编码dls1的基因的破坏。14.在如段落10-13中任一项所述的经修饰的酵母的一些实施例中,所述酵母进一步包含编码碳水化合物加工酶的外源基因。15.在如段落10-14中任一项所述的经修饰的酵母的一些实施例中,所述细胞是酵母属物种。根据包括附图/图的说明书,本发明的经修饰的细胞和方法的这些和其他方面以及实施例将是清楚的。附图说明图1是典型的mal基因座以及它通过麦芽糖和葡萄糖调节的图图2描绘了mal23c表达盒。图3是质粒phx19的图谱。图4是用菌株fg和a28发酵期间累积co2压力指数的比较。图5是质粒pzkap1m-(h3c19)的图谱。图6是质粒ptopoii-bluntura3-loxp-kanmx-loxp-ura3的图谱。图7是质粒pgal-cre-316的图谱。图8是用rhy723及其亲本菌株gpy10008发酵期间累积co2压力指数的比较。具体实施方式i.定义在详细地描述本发明的酵母和方法之前,为了清楚起见定义以下术语。未定义的术语应当符合相关领域中所使用的常规含义。如本文所使用的,术语“醇”是指其中羟基官能团(-oh)与饱和碳原子键合的有机化合物。如本文所使用的,术语“酵母细胞”、酵母菌株,或简称“酵母”是指来自子囊菌门和担子菌门的生物体。示例性酵母是来自酵母目(saccharomycetales)的芽殖酵母。酵母的具体实例是酵母属物种,包括但不限于酿酒酵母(s.cerevisiae)。酵母包括用于生产燃料醇的生物体以及用于生产可饮用醇的生物体,包括用于制备独特味道的啤酒、葡萄酒和其他发酵饮料的特种和专有酵母菌株。如本文所使用的,短语“工程化的酵母细胞”、“变体酵母细胞”、“经修饰的酵母细胞”、或相似短语是指包括本文所述的遗传修饰和特征的酵母。变体/经修饰的酵母不包括天然存在的酵母。如本文所使用的,术语“多肽”和“蛋白”(以及它们各自的复数形式)可互换地使用,是指包含通过肽键连接的氨基酸残基的任何长度的聚合物。本文使用氨基酸残基的常规一字母或三字母代码,并且所有序列均从n-末端到c-末端方向进行呈现。聚合物可以包含经修饰的氨基酸,并且它可以被非氨基酸中断。这些术语还包括天然地修饰的或通过干预而修饰的氨基酸聚合物;例如,通过二硫键形成、糖基化、脂化、乙酰化、磷酸化或任何其他操作或修饰,如与标记组分偶联。该定义内还包括例如含有一种或多种氨基酸类似物(包括例如非天然氨基酸等)以及本领域已知的其他修饰的多肽。如本文所使用的,功能上和/或结构上相似的蛋白被认为是“相关的蛋白”或“同源物”。这类蛋白质可以衍生自不同属和/或物种的生物体,或不同纲的生物体(例如,细菌和真菌),或者是人工设计的蛋白质。相关的蛋白质还涵盖通过一级序列分析确定的、通过二级或三级结构分析确定的、或者通过免疫交叉反应性确定的、或者通过它们的功能确定的同源物。如本文所使用的,术语“同源蛋白”是指与参考蛋白具有相似活性和/或结构的蛋白。这并不旨在意味着同源物必定与进化相关。因此,该术语旨在涵盖从不同生物体获得的相同、相似、或相应(即,在结构和功能方面)的一种或多种酶。在一些实施例中,希望鉴定与参考蛋白具有类似的四级、三级和/或一级结构的同源物。在一些实施例中,同源蛋白诱导与参考蛋白相似的一种或多种免疫应答。在一些实施例中,将同源蛋白工程化以产生具有所希望活性的酶。序列之间的同源性程度可以使用本领域已知的任何适合方法确定(参见,例如,smith和waterman(1981)adv.appl.math.[应用数学进展]2:482;needleman和wunsch(1970)j.mol.biol.[分子生物学杂志],48:443;pearson和lipman(1988)proc.natl.acad.sci.usa[美国科学院院报]85:2444;威斯康星遗传学软件包(wisconsingeneticssoftwarepackage)(遗传学计算机组公司(geneticscomputergroup),麦迪逊,威斯康星州)中的程序,如gap、bestfit、fasta和tfasta;以及thompson等人(1984)nucleicacidsres.[核酸研究]12:387-95)。例如,pileup是确定序列同源性水平的有用程序。pileup使用渐进的、两两比对创建了来自一组相关序列的多重序列比对。它还可以标绘显示用于创建该比对的聚类关系的一个树。pileup使用feng和doolittle的渐进比对方法的简化(feng和doolittle(1987)j.mol.evol.[分子进化杂志]35:351-60)。该方法类似于higgins和sharp描述的方法((1989)cabios5:151-53)。有用的pileup参数包括为3.00的默认空位权重,为0.10的默认空位长度权重,以及加权的末端空位。有用算法的另一个实例是blast算法,由以下描述:altschul等人((1990)j.mol.biol.[分子生物学杂志]215:403-10)以及karlin等人((1993)proc.natl.acad.sci.usa[美国科学院院报]90:5873-87)。一个特别有用的blast程序是wu-blast-2程序(参见,例如,altschul等人(1996)meth.enzymol.[酶学方法]266:460-80)。参数“w”、“t”、以及“x”确定了该比对的灵敏度与速度。所述blast程序使用字长(w)为11、blosum62得分矩阵(参见,例如,henikoff和henikoff(1989)proc.natl.acad.sci.usa[美国科学院院报]89:10915)比对(b)为50、期望值(e)为10、m'5、n'-4、以及两条链的比较作为默认值。如本文所使用的,在至少两个核酸或多肽的上下文中,短语“基本上相似”和“基本上相同”典型地意指多核苷酸或多肽包含与参考(即,野生型)序列相比具有至少约70%同一性、至少约75%同一性、至少约80%同一性、至少约85%同一性、至少约90%同一性、至少约91%同一性、至少约92%同一性、至少约93%同一性、至少约94%同一性、至少约95%同一性、至少约96%同一性、至少约97%同一性、至少约98%同一性、或甚至至少约99%同一性、或更高同一性的序列。使用具有默认参数的clustalw算法计算序列同一性百分比。参见thompson等人(1994)nucleicacidsres.[核酸研究]22:4673-4680。clustalw算法的默认参数是:两种多肽基本上相同的另一个指示是第一多肽与第二多肽具有免疫交叉反应性。典型地,差别在于保守氨基酸取代的多肽具有免疫交叉反应性。因此,多肽与第二多肽基本上相同,例如,其中两个肽仅相差保守取代。两个核酸序列基本上相同的另一个指示是两个分子在严格条件下(例如,在中等至高严格的范围内)彼此杂交。如本文所使用的,术语“基因”与术语“等位基因”同义,是指编码和指导蛋白或rna表达的核酸。丝状真菌的营养体形式通常是单倍体,因此指定基因的单拷贝(即单个等位基因)足以赋予特定表型。当生物体含有多于一个相似基因时,术语“等位基因”通常是优选的,在这种情况下,每个不同的相似基因被称为不同的“等位基因”。如本文所使用的,“组成型”表达是指基本上在所有典型生长条件下,由特定基因编码的多肽的产生,而不是“条件”表达,所述“条件”表达需要特定的底物的存在、温度等来诱导或激活表达。如本文所使用的,术语“表达多肽”和相似术语是指使用细胞的翻译机器(例如,核糖体)产生多肽的细胞过程。如本文所使用的,“过表达多肽”、“提高多肽的表达”和类似术语是指与不包括指定的遗传修饰的亲本或“野生型细胞”所观察到的相比,以比正常较高的水平表达多肽。如本文所使用的,“表达盒”是指包括启动子、和氨基酸编码区与终止子(即启动子::氨基酸编码区::终止子)以及允许在细胞中产生编码的多肽需要的其他核酸序列的dna片段。表达盒可以是外源的(即,引入细胞中)或内源的(即,存在于细胞中)。如本文所使用的,关于两个dna片段的术语“融合的”和“融合”(例如启动子和多肽的编码区)是指致使两个dna片段变成单个分子的物理键合。如本文所使用的,术语“野生型”和“天然”可互换地使用,并且是指在自然界中发现的基因、蛋白质或菌株,或者不是为了目前所述酵母的优点而有意修饰的基因、蛋白质或菌株。如本文所使用的,术语“目的蛋白”是指希望在经修饰的酵母中表达的多肽。此类蛋白质可以是酶、底物结合蛋白、表面活性蛋白、结构蛋白、选择性标记等,并且可以表达。目的蛋白由相对于亲本菌株的内源基因或异源基因(即,目的基因)编码。目的蛋白可以在细胞内表达或作为分泌的蛋白表达。如本文所使用的,“基因的破坏”泛指任何实质上阻止细胞在宿主细胞中产生功能性基因产物(例如,蛋白)的遗传的或化学的操作(即,突变)。示例性破坏方法包括基因的任何部分的完全或部分(包括多肽编码序列、启动子、增强子或额外的调节元件)缺失、或其诱变,其中诱变涵盖取代、插入、缺失、倒位、及其组合和变化,任何这些突变基本上阻止功能基因产物的产生。也可以使用rnai、反义、或任何其他消除基因表达的方法破坏基因。可以通过非相邻控制元件的缺失或遗传操作来破坏基因。如本文所使用的,“基因缺失”是指该基因从宿主细胞的基因组中去除。当基因包括与基因编码序列不紧邻的控制元件(例如,增强子元件)时,基因的缺失是指编码序列、以及任选地相邻增强子元件(例如,包括但不限于启动子和/或终止子序列)的缺失,但未要求非相邻控制元件的缺失。如本文所使用的,术语“遗传操作”和“遗传改变”可互换地使用,并且是指核酸序列的改变/变化。改变可包括但不限于核酸序列中至少一种核酸的取代、缺失、插入或化学修饰。如本文所使用的,“功能性多肽/蛋白”是具有活性(诸如酶活性、结合活性、表面活性特性等)的蛋白质,并且其未被诱变、截短、或以其他方式修饰而消除或减少此活性。如所指出的,功能性多肽可以是热稳定的或不耐热的。如本文所使用的,“功能性基因”是能够被细胞组分用于产生活性基因产物(典型地是蛋白)的基因。功能性基因是破坏的基因的对立体,破坏的基因被修饰使得它们不能被细胞组分用于产生活性基因产物,或者具有减少的被细胞组分用于产生活性基因产物的能力。如本文所使用的,如果已对酵母细胞进行遗传或化学改变以防止产生呈现出野生型蛋白质的活性特征的功能性蛋白质/多肽,则对所述酵母细胞已经进行了“修饰以防止产生特定蛋白质”。此类修饰包括但不限于编码蛋白质(如本文所述)的基因的缺失或破坏、使得编码的多肽缺乏前述活性的基因修饰、影响翻译后加工或稳定性的基因修饰、及其组合。如本文所使用的,“途径的减弱”或“通过途径的通量的减弱”(即,生物化学途径)泛指通过代谢途径减少或完全阻止生物化学底物或中间体的通量的任何遗传的或化学的操作。可以通过各种众所周知的方法实现途径的减弱。此类方法包括,但不限于:完全或部分缺失一个或多个基因、用编码具有减少的催化活性或增加的km值的酶的突变体形式替代这些基因的野生型等位基因、修饰启动子或控制一种或多个基因表达的其他调控元件、针对减少的稳定性工程化所述酶或编码这些酶的mrna、将酶错误指导进不太可能与底物和中间体相互作用的细胞区室、使用干扰rna等。如本文所使用的,“需氧发酵”是指在氧存在下的生长。如本文所使用的,“厌氧发酵”是指在不存在氧的情况下的生长。如本文所使用的,表达“发酵结束”是指就固定和可变成本而言,当连续发酵产生少量额外的醇的经济优势被连续发酵的成本超过时的发酵阶段。在更一般的意义上,“发酵结束”是指发酵不再产生大量额外的醇,即不超过约1%的额外的醇的点。如本文所使用的,表达“碳通量”是指碳分子通过代谢途径的周转率。碳通量是由参与代谢途径,例如葡萄糖代谢途径和麦芽糖代谢途径的酶调节的。如本文所使用的,单数冠词“一个/一种(a/an)”以及“所述”涵盖复数个指示物,除非上下文中另外明确指明。本文引用的所有参考文献均通过引用以其全文特此并入。除非另外说明,以下缩写/首字母缩略词具有以下含义:ec酶学委员会pkl磷酸转酮酶pta磷酸转乙酰酶aadh乙醛脱氢酶adh醇脱氢酶etoh乙醇aaα-淀粉酶ga葡糖淀粉酶℃摄氏度bp碱基对dna脱氧核糖核酸ds或ds干固体g或gm克g/l克/升h2o水hplc高效液相色谱hr或h小时kg千克m摩尔的mg毫克ml或ml毫升min分钟mm毫摩尔的n正常nm纳米pcr聚合酶链式反应ppm份/百万份δ与缺失有关μg微克μl和μl微升μm微摩尔的ii.具有组成型mal等位基因的经修饰的酵母本发明的诸位发明人发现,与在其他方面相同的亲本酵母相比,具有组成型转录激活因子mal等位基因的经修饰的酵母在淀粉水解物发酵中产生更多乙醇,和/或在更短时间内产生相同量的乙醇。值得注意的是,产生的额外的乙醇似乎不是麦芽糖代谢的结果,或不是至少仅仅麦芽糖代谢的结果,因为淀粉水解物中存在的少量麦芽糖在发酵后基本保持不变。与在其他方面相同的亲本酵母相比,在淀粉水解物发酵中,具有组成型转录激活因子mal等位基因的酵母也以更高速率产生乙醇。因此,转录激活因子mal等位基因的组成型表达似乎对高糖环境中的葡萄糖代谢产生了出乎意料的益处。组成型转录激活因子mal等位基因的存在,任选地在其他遗传修饰的存在下,导致乙醇生产的增加,或乙醇生产速率增加至少0.5%、至少0.6%、至少0.7%、至少0.8%、至少0.9%、至少1.0%、至少1.1%、至少1.2%或更多。尽管此类增长幅度较小,考虑到当前生产的乙醇量,这相当于在体积上大幅度的增加。本发明的酵母菌株和方法使用代表性的组成型转录激活因子mal23等位基因为例,如以下seqidno:1所示:然而,完全可以预期,其他组成型转录激活因子mal等位基因的引入将产生类似结果。组成型转录激活因子mal等位基因已经被认知超过半个世纪,并以以下所述为例:例如,winge,o.和roberts,c.(1950)c.r.trav.lab.carlsbergser.physiol.25:35-81,kahn,n.a.和eaton,n.r.(1971)mol.gen.genet.[分子遗传学与普通遗传学]112:317-22;charronm,j.和michels,c.a.(1987)genetics[遗传学]11623-31;zimmerman和eaton,n.r.(1974)mol.gen.genet.[分子遗传学与普通遗传学]134261-271;rodicio,r.(1986)curr.genet.[当代遗传学]11:235-41和tenberge,a.m.a.等人(1973)mol.gen.genet.[分子遗传学与普通遗传学]125:139-46,所述文献各自通过引用并入本文。具有组成型转录激活因子mal等位基因的酵母还可以包括增加醇生产的其他遗传操作,特别是与增强的麦芽糖代谢无关的修饰。iii.具有与外源pkl途径基因组合的组成型mal等位基因的经修饰的酵母组成型转录激活因子mal等位基因的存在可以与pkl途径中基因的表达组合,以增加细胞的生长速率,并进一步增加乙醇生产。先前已经描述了具有异源pkl途径的工程化酵母细胞(例如,wo2015148272)。这些细胞表达异源pkl(ec4.1.2.9)和pta(ec2.3.1.8),任选地具有其他酶,以使通道碳通量远离甘油途径导向,并且朝向乙酰辅酶a的合成导向,所述乙酰辅酶a然后转化为乙醇。与在其他方面相同的亲本酵母细胞相比,此类经修饰的细胞能够在发酵过程中增加乙醇生产。iv.具有组成型mal等位基因并且具有降低的dls1表达的经修饰的酵母dls1,由yjl065c编码,是isw2酵母染色质可接近性复合物(ychrac)的167个氨基酸多肽亚基,所述isw2酵母染色质可接近性复合物含有isw2、itc1、dpb3样亚基(dls1)、和dpb4(参见,例如,peterson,c.l.(1996)curr.opin.genet.dev.[遗传学与发育新观点]6:171-75和winston,f.和carlson,m.(1992)trendsgenet.[遗传学趋势]8:387-91)。在没有其他遗传修饰的情况下,具有降低细胞中功能性dls1的量的遗传改变的酵母在醇发酵中表现出增加的稳健性,这允许更高温度和可能更短的发酵(数据未示出)。减少细胞中产生的功能性dls1的量可以通过破坏yjl065c基因来完成。yjl065c基因的破坏可以使用基本上阻止功能性yjl065c基因产物(即,dls1)表达的任何适合方法进行。如本领域技术人员已知的示例性破坏方法包括但不限于:完全或部分缺失yjl065c基因,包括完全或部分缺失例如dls1编码序列、启动子、终止子、增强子或其他调节元件;以及完全或部分缺失包含yjl065c基因任何部分的染色体的一部分。破坏yjl065c基因的具体方法包括在yjl065c基因的任何部分(例如,dls1编码序列、启动子、终止子、增强子或另一个调节元件)中进行核苷酸取代或插入。优选地,缺失、插入和/或取代(统称为突变)通过使用序列特异性分子生物学技术的遗传操作来进行,与通过化学诱变相反,化学诱变通常不靶向特定核酸序列。尽管如此,理论上,化学诱变仍可用于破坏yjl065c基因。v.影响醇生产的额外的突变本发明的经修饰的酵母可进一步包括或可以明确排除导致天然甘油生物合成途径减弱的突变,已知所述突变增加醇生产。用于减弱酵母中甘油生物合成途径的方法是已知的,并包括例如通过破坏基因gpd1、gpd2、gpp1和/或gpp2中的一个或多个来降低或消除内源性nad依赖性甘油3-磷酸脱氢酶(gpd)或磷酸甘油磷酸酶(gpp)活性。参见例如美国专利号9,175,270(elke等人)、8,795,998(pronk等人)和8,956,851(argyros等人)。经修饰的酵母的特征可进一步在于增加的乙酰辅酶a合酶(也称为乙酰辅酶a连接酶)活性(ec6.2.1.1)以清除(即捕获)通过化学或酶水解乙酰-磷酸产生(或出于任何其他原因存在于酵母的培养基中)的乙酸并将其转化为乙酰辅酶a。这避免了乙酸对酵母细胞生长的不良影响,并且可进一步有助于醇产量的提高。增加乙酰辅酶a合酶活性可以通过将异源乙酰辅酶a合酶基因引入细胞、增加内源乙酰辅酶a合酶基因的表达等来实现。用于引入细胞中的特别有用的乙酰辅酶a合酶可以从康斯力鬃毛甲烷菌(methanosaetaconcilii)(uniprot/trembl登录号:wp_013718460)获得。该酶的同源物,包括与上述来自康斯力鬃毛甲烷菌的乙酰辅酶a合酶具有至少85%、至少90%、至少92%、至少95%、至少97%、至少98%、以及甚至至少99%氨基酸序列同一性的酶,也可用于本发明的组合物和方法中。在其他实施例中,本发明的经修饰的酵母不具有增加的乙酰辅酶a合酶。在一些实施例中,本发明的经修饰的酵母可进一步包括编码具有nad+依赖性乙酰化乙醛脱氢酶活性的蛋白质的异源基因和/或编码丙酮酸甲酸裂解酶的异源基因。例如,在美国专利号8,795,998(pronk等人)中描述了与甘油途径减弱进行组合的此类基因的引入。然而,在本发明组合物和方法的大多数实施例中,不需要引入乙酰化乙醛脱氢酶和/或丙酮酸甲酸裂解酶,因为通过减弱导致氧化还原辅因子失衡的用于制备乙酰辅酶a的天然生物合成途径消除了对这些活性的需要。因此,在一些实施例中,本发明的酵母不具有编码nad+依赖性乙酰化乙醛脱氢酶的异源基因和/或编码丙酮酸甲酸裂解酶的异源基因。在一些实施例中,本发明的经修饰的酵母进一步包含丁醇生物合成途径。在一些实施例中,所述丁醇生物合成途径是异丁醇生物合成途径。所述异丁醇生物合成途径可以包含编码多肽的多核苷酸,所述多肽催化选自由以下组成的组的底物至产物的转化:(a)丙酮酸至乙酰乳酸;(b)乙酰乳酸至2,3-二羟基异戊酸盐;(c)2,3-二羟基异戊酸盐至2-酮异戊酸盐;(d)2-酮异戊酸盐至异丁醛;和(e)异丁醛至异丁醇。所述异丁醇生物合成途径可以包含编码具有乙酰乳酸合酶、酮酸还原异构酶、二羟酸脱水酶、酮异戊酸脱羧酶、和醇脱氢酶活性的多肽的多核苷酸。在一些实施例中,包含丁醇生物合成途径的经修饰的酵母进一步包含编码具有丙酮酸脱羧酶活性的多肽的多核苷酸中的修饰。所述酵母在编码具有丙酮酸脱羧酶活性的多肽的内源多核苷酸中可以包含缺失、突变和/或取代。在一些实施例中,具有丙酮酸脱羧酶活性的多肽选自下组,该组由以下组成:pdc1、pdc5、pdc6、及其组合。在一些实施例中,酵母细胞在编码fra2、ald6、adh1、gpd2、bdh1、和ymr226c的一个或多个内源多核苷酸中进一步包含缺失、突变和/或取代。在其他实施例中,本发明的经修饰的酵母细胞没有进一步包含丁醇生物合成途径。本发明的经修饰的酵母可以包括编码目的蛋白的任何数量的额外的目的基因,所述目的蛋白包括选择性标记、碳水化合物加工酶以及其他商业上相关的多肽,包括但不限于选自由以下项组成的组的酶:脱氢酶、转酮醇酶、磷酸转酮酶、转醛醇酶、差向异构酶、植酸酶、木聚糖酶、β-葡聚糖酶、磷酸酶、蛋白酶、α-淀粉酶、β-淀粉酶、葡糖淀粉酶、支链淀粉酶、异淀粉酶、纤维素酶、海藻糖酶、脂肪酶、果胶酶、聚酯酶、角质酶、氧化酶、转移酶、还原酶、半纤维素酶、甘露聚糖酶、酯酶、异构酶、果胶酶、乳糖酶、过氧化物酶和漆酶。目的蛋白可以被分泌、糖基化和以其他方式修饰。vi.经修饰的酵母用于增加醇生产的用途本发明的酵母及其使用方法包括用于增加发酵反应中醇生产的方法。这类方法不限于特定的发酵过程。预期本发明的工程化酵母是任何醇发酵设施中常规酵母的“滴入式(drop-in)”替代品。虽然主要旨在用于燃料乙醇生产,但是本发明的酵母也可以用于生产可饮用酒精,包括葡萄酒和啤酒。vii.适合修饰的酵母酵母是被归类为真菌界成员的单细胞真核微生物,并且包括来自子囊菌门和担子菌门的生物体。可以用于醇生产的酵母包括但不限于酵母属物种,包括酿酒酵母、以及克鲁维酵母属(kluyveromyces)、克鲁维酵母属(lachancea)和裂殖酵母属(schizosaccharomyces)物种。许多酵母菌株是可商购获得的,其中许多已被选择或基因工程化以获得所需的特征,诸如高乙醇生产、快速生长速率等。一些酵母已经被基因工程化以产生异源酶,例如葡糖淀粉酶或α-淀粉酶。viii.底物和产物从许多碳水化合物底物(包括但不限于玉米淀粉、甘蔗、木薯和糖蜜)中生产醇是众所周知的,正如酶条件和化学条件以及机械方法的无数变化和改善也是众所周知的。据信本发明的组合物和方法与此类底物和条件完全相容。高水平的麦芽糖不需要从本发明的组合物和方法中受益。在一些实施例中,所述浓度小于约10g/l。鉴于本说明书,本发明的菌株和方法的这些和其他方面以及实施方案对于技术人员是清楚的。以下实例旨在进一步说明但不限制菌株和方法。实例实例1材料与方法液化物制备:液化物(玉米醪浆料)通过添加600ppm尿素、0.124sapu/gdsfermgentm2.5x(酸性真菌蛋白酶)、0.33gau/gdscs4(木霉属(trichoderma)葡糖淀粉酶变体)以及1.46sscu/gdsakaa(白曲霉(aspergilluskawachii)α-淀粉酶)、调节至ph4.8来制备。血清瓶测定:向在24孔板中的2mlypd接种酵母细胞,并且使培养物生长过夜至25-30之间的od。将2.5ml液化物转移到血清瓶(chemglass公司,目录号:cg-4904-01),并且将酵母添加到每个小瓶,达到约0.4-0.6的最终od。安装小瓶的盖子,并且用针(bd公司,目录号:305111)刺穿以用于通风(以释放co2),然后在振荡(200rpm,持续65小时)下在32℃孵育。ankom测定:将300μl浓缩的酵母过夜培养物添加到填充有50g制备的液化物(参见上文)的多个ankom瓶中的每个中,以达到0.3的最终od。然后将所述瓶在振荡(以150rpm)下在32℃孵育65小时。hplc分析:通过在14,000rpm下离心12分钟在eppendorf管中收集来自血清瓶和ankom测定的培养物的样品。将上清液使用0.2μmptfe过滤器过滤,并且然后在以下条件下用于hplc(agilenttechnologies1200系列)分析:bio-radaminexhpx-87h柱,运行温度为55c。0.6ml/min等度流速,0.01nh2so4,2.5μl注射体积。使用校准标准物用于定量乙酸、乙醇、甘油、葡萄糖和其他分子。所有值以g/l报告。实例2组成型mal23等位基因盒的制备使用标准程序制备mal23c表达盒,所述mal23c表达盒由在mal23c启动子(seqidno:3)和mal23终止子(seqidno:4)控制下的mal23的组成型等位基因(seqidno:2)组成。所述核苷酸序列如下所示,并且所述盒的表示如图2中所示:组成型mal23等位基因的dna编码区(seqidno:2):mal23c启动子的dna序列(seqidno:3):mal23c终止子的dna序列(seqidno:4):通过用适当的引物扩增mal23c表达盒,制备包括侧接ydl227c基因座序列的dna片段。所述片段被插入到一个命名为phx19的质粒中,所述质粒在酵母属染色体ydl227c基因座处包括整合的组成型mal23c表达盒,如图3中所示。质粒phx19的功能和结构组成在表1中描述。表1.质粒phx19的功能和结构元件实例3具有mal23c表达盒的酵母的产生为了研究mal23c在工业酵母中的作用,野生型fermaxtm黄金菌株(martrex,inc.公司,查斯卡市,明尼苏达州,美国),以下简称“fg”,被用作亲本,以在ydl227c整合位点引入mal23c表达盒。用3,362-bp的pcr扩增dna片段转化细胞,所述pcr扩增dna片段使用适当的侧接引物并以phx19质粒为模板。选择转化子,并将一个代表性成员命名为菌株a28。实例4具有或不具有mal23c的fg酵母的比较菌株a28和亲本菌株fg生长在ankom瓶中,且其发酵产物如实施例1中所述进行分析。关于累积co2压力指数(cpi)(指示发酵速率)的性能,在图4中示出。关于乙醇、甘油和乙酸生产的性能在表2中示出。表2.fg相对于a28,在ankom测定中在麦芽三糖、麦芽糖和葡萄糖的残留量方面,fg亲本和菌株a28的性能相似,这表明在ydl227c基因座处整合mal23cdna片段不会影响葡萄糖基玉米醪发酵中的麦芽糖代谢。mal23c表达菌株的乙醇滴度比fg对照的乙醇滴度高1.1%。图4中的cpi数据证明菌株a28的发酵速率也高于亲本fg菌株的发酵速率。重复进行实验,并确认结果。实例5含有完全pkl途径的质粒的产生设计质粒pzkap1m-(h3c19)以整合aap1基因座(yhr047c)上游的四个单独的多肽表达盒。所述四个盒如以下:(i)hxt3启动子::pklc::fba1终止子;(ii)pgk1启动子::lppta::pgk1终止子;(iii)tdh3启动子::eutea19::eno终止子;和(iv)pdc1启动子::acsa1::pdc1终止子,所述四个盒被设计以分别表达编码衍生自加德纳菌(gardnerellavaginalis)的磷酸转酮酶(pkl)、衍生自植物乳杆菌(lactobacillusplantarum)的磷酸转乙酰酶(pta)、衍生自desulfospirajoergensenii的乙酰化乙醛脱氢酶、和衍生自康斯力鬃毛甲烷菌的乙酰辅酶a合酶的密码子优化基因。pzkap1m-(h3c19)的图谱在图5中示出。功能和结构元件在表3中详细列出。表3.构建体pzkap1m-(h3c19)的功能和结构元件实例6fg-ura3酵母的产生fg菌株被用作亲本菌株,以制备ura3营养缺陷型菌株fg-ura3。将质粒ptopoii-bluntura3-loxp-kanmx-loxp-ura3(图6中描绘的)设计成用突变的ura3和ura3-loxp-tefp-kanmx-teft-loxp-ura3片段替代菌株fg中的ura3基因。质粒的功能和结构元件在表4中列出。表4.ptopoii-bluntura3-loxp-kanmx-loxp-ura3的功能/结构元件含有ura3-loxp-kanmx-loxp-ura3盒的2,018-bpdna片段通过ecori消化从质粒topoii-bluntura3-loxp-kanmx-loxp-ura3释放。通过电穿孔使用所述片段转化酿酒酵母fg细胞。能够在含有g418的培养基上生长的转化的菌落在含有20μg/ml尿嘧啶和2mg/ml5-氟乳清酸(5-foa)的合成小型板上划线。通过sd-ura板上表型的生长和通过pcr进一步确认能够在5-foa板上生长的菌落的ura3缺失。ura3缺失转化体不能在sd-ura板上生长。用测试引物获得单个1.98-kbpcr片段。相比之下,使用来自亲本fg菌株的dna,相同的引物对生成1.3-kb片段,从而指示完整ura3基因的存在。ura3缺失菌株命名为fg-kanmx-ura3。为了从菌株fg-kanmx-ura3去除kanmx表达盒,使用质粒pgal-cre-316(如图7中描绘的)通过电穿孔用于转化菌株fg-kanmx-ura3的细胞。使用此质粒的目的是暂时表达cre酶,这样使得loxp夹层kanmx基因将从菌株fg-kanmx-ura3去除以生成菌株fg-ura3。pgal-cre-316是随后从菌株fg-ura3去除的自我复制的环状质粒。来自pgal-cre-316的所有序列元件均不插入到菌株fg-ura3基因座中。质粒pgal-cre-316的功能和结构元件在表5中列出。表5.pgal-cre-316的功能和结构元件。功能/结构元件酵母-细菌穿梭载体prs316序列pbr322复制起点酿酒酵母ura3基因f1起点galp-cre-adht盒,反向取向将转化的细胞铺板在sd-ura板上。将单个菌落转移到ypg板上,并且在30℃孵育2至3天。然后将菌落转移到新的ypd板,再持续几天。最后,将来自ypd板的细胞悬浮液点样在以下板上:ypd,g418(150μg/ml),5-foa(2mg/ml)和sd-ura。选择能够在ypd和5-foa上生长并且不能够在g418和sd-ura板上生长的细胞用于如上所述的pcr确认。预期的pcr产物大小是0.4-kb,并且确认衍生自fg-kanmx-ura3的kanmx(遗传霉素)敏感性ura3缺失菌株的身份。此菌株命名为fg-ura3。实例7具有pkl途径的酵母的产生fg-ura3菌株被作为亲本以引入pkl途径。用14,993-bpkasi片段转化细胞,所述片段含有来自实例5的pzkap1m-(h3c19)的四个表达盒。选择具有整合yhr047c基因座上游的kasi片段的转化子,并命名为菌株gpy10000。菌株fg和gpy10000在小瓶培养物中生长,并且其发酵产物如实施例1中所述进行分析。关于乙醇、甘油和乙酸生产的性能在表6中示出。表6.fg相对于g176,在小瓶测定中菌株表达的一种或多种转基因甘油乙酸etohfg无17.300.70140.9gpy10000pkl途径12.941.86146.3表达pkl途径的gpy10000菌株的乙醇滴度比fg菌株的乙醇滴度高3.8%。不出人意料地,乙酸的水平也升高了,这是具有pkl途径的酵母的一个已知特性。实例8具有pkl途径并且dls1减产的酵母的产生菌株gpy10008是由菌株gpy10000中的yjl065c(其编码dls1)的cas9介导的缺失产生的。具体来说,从yjl065c的起始密码子前4-bp到终止密码子前10-bp造成缺失。fg酵母菌株gpy10008及其亲本菌株g10008在ankom瓶中生长,并且其发酵产物如实例1中所述进行分析。关于乙醇、甘油和乙酸生产的性能在表7中示出。表7.gpy10000相对于gpy10008,在ankom测定中菌株表达的一种或多种转基因甘油乙酸etohgpy10000pkl途径12.851.80143.39gpy10008pkl途径+δyjl065c13.151.78147.23具有yjl065c缺失的菌株的乙醇滴度比未缺失菌株的乙醇滴度高2.7%。乙酸水平大致相同。实例9组合mal23c与pkl途径并且dls1减产的酵母的产生为了研究mal23c在具有pkl途径并且dls1表达降低的工程fg菌株中的作用,菌株gpy10008被用作亲本以引入mal23c表达盒,在这种情况下,在yhl041w基因座的3′区域,其他如实例3中所述。用含有mal23c表达盒的pcr扩增dna片段转化gpy10008细胞,所述pcr扩增dna片段是使用作为模板的phx19质粒和包括侧接yhl041w基因座序列的引物制备的。将命名为rhy723的新的fg酵母菌株及其亲本菌株gpy10008在ankoms瓶中生长,并且其发酵产物如实例1中所述进行分析。关于乙醇、甘油和乙酸生产的性能如表8所示,并且发酵期间以cpi表示的发酵速率如图8中所示。表8.rhy720相对于gpy10008,在ankom测定中菌株rhy723的乙醇滴度比菌株gpy10008的乙醇滴度高0.83%,证明mal23c的表达增加了乙醇生产,即使在也含有pkl途径并且不表达dls1的酵母中。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1