靶向肿瘤抗原的嵌合抗原受体的制作方法
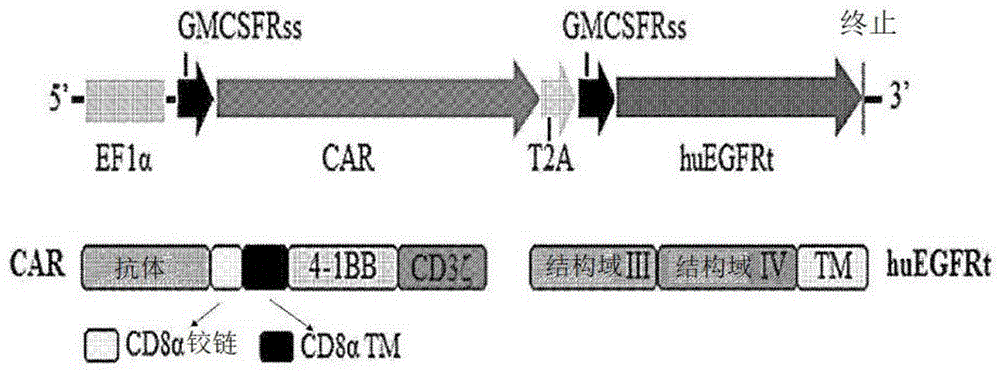
相关申请的交叉引用本申请要求2017年11月10日提交的美国临时申请第62/584,421号的权益,其通过引用结合入本文。本公开涉及对肿瘤抗原特异的嵌合抗原受体,及其用于癌症免疫疗法的用途。政府支持的致谢本发明在美国国家卫生研究院、国家癌症研究所(nationalinstitutesofhealth,nationalcancerinstitute)所授予的项目号z01bc010891的政府支持下做出。美国政府对本发明拥有一定的权利。
背景技术:
:嵌合抗原受体(car)由与跨膜结构域和t细胞信号传导部分融合的对肿瘤抗原特异的抗体片段组成。当所述受体在t细胞表面上表达时,其介导与靶的结合并激活t细胞,最终诱导靶细胞裂解。car正成为最有希望的治疗血液恶性肿瘤的方法之一(kochenderferetal.,blood119:2709-2720,2012;kochenderferandrosenberg,natrevclinoncol10:267-276,2013;porteretal.,newengljmed365:725-733,2011;maudeetal.,newengljmed371:1507-1517,2014;gruppetal.,newengljmed368:1509-1518,2013)。在美国已经批准了两种cd19-靶向的car,即axicabtageneciloleucel(yescartatm)和tisagenlecleucel(kymriahtm),它们分别用于治疗b细胞非霍奇金淋巴瘤和b细胞急性淋巴细胞白血病。目前正在进行临床试验,以测试用于治疗实体瘤的各种cart细胞疗法(yuetal.,jhematoloncol10(1):78,2017)。发明概要本文公开了编码嵌合抗原受体(car)和截短的人表皮生长因子受体(huegfrt)的核酸构建体。编码的car包括与细胞外铰链区、跨膜区、细胞内共刺激结构域和细胞内信号传导结构域融合的肿瘤抗原特异性单克隆抗体片段。huegfrt包括两个egfr胞外结构域(结构域iii和结构域iv)和一个egfr跨膜结构域,但是缺少两个膜远端胞外结构域和所有的细胞内结构域。还公开了共表达所公开的car和huegfrt的分离的细胞,例如t淋巴细胞。用car构建体转导的t细胞可用于癌症免疫疗法。本文提供了编码car和huegfrt的核酸分子。在一些实施方式中,所述核酸分子以5’至3’的方向包括编码第一信号序列的核酸;编码抗原特异性抗体或其抗原结合片段的核酸;编码细胞外铰链区的核酸;编码跨膜结构域的核酸;编码细胞内共刺激结构域的核酸;编码细胞内信号传导结构域的核酸;编码自裂解2a肽的核酸;编码第二信号序列的核酸;和编码huegfrt的核酸。在一些实例中,第一和/或第二信号序列是粒细胞-巨噬细胞集落刺激因子受体信号序列(gmcsfrss),细胞外铰链区是cd8α铰链区,跨膜结构域是cd8α跨膜结构域,细胞内共刺激结构域是4-1bb共刺激结构域,并且细胞内信号传导结构域是cd3ζ信号传导结构域。在一些实例中,所述抗体或抗原结合片段特异性结合肿瘤抗原,例如glypican-3(gpc3)、gpc2或间皮素。还提供了包括本文公开的核酸分子的载体,例如病毒载体。在具体的非限制性实例中,病毒载体是慢病毒载体。进一步提供了包括本文公开的核酸分子的分离的宿主细胞。还提供了共表达car和huegfrt的分离的宿主细胞。在一些实施方式中,car包括抗原特异性抗体或其抗原结合片段、细胞外铰链区、跨膜结构域、细胞内共刺激结构域和细胞内信号传导结构域;和/或huegfrt包括来自人egfr的结构域iii、结构域iv和跨膜结构域,但是缺少表皮生长因子(egf)-结合结构域和细胞质结构域。在一些实例中,细胞外铰链区包括cd8α铰链区,跨膜结构域包括cd8α跨膜结构域,细胞内共刺激结构域包括4-1bb共刺激结构域,并且细胞内信号传导结构域包括cd3ζ信号传导结构域。在一些实例中,所述抗体或抗原结合片段特异性结合肿瘤抗原,例如gpc3、gpc2或间皮素。进一步提供了包括本文公开的分离的宿主细胞和药学上可接受的载体的组合物。在一些实施方式中,分离的宿主细胞是t淋巴细胞。进一步提供了在受试者中治疗gpc3-阳性癌症、gpc2-阳性癌症或间皮素-阳性癌症的方法,通过给所述受试者施用本文公开的分离的宿主细胞。在一些实施方式中,分离的宿主细胞是t淋巴细胞,例如自体t淋巴细胞。从以下参考附图进行的详细描述中,本发明的前述和其它目的、特征和优点将变得更加明显。附图说明图1是用于产生肿瘤-靶向嵌合抗原受体(car)的慢病毒构建体的示意图。该慢病毒构建体包括car编码区和编码截短的人表皮生长因子受体(huegfrt)的区域,各自之前都有一个粒细胞-巨噬细胞集落刺激因子受体信号序列(gmcsfrss)。所述两个区域被自裂解t2a序列分开,使得在该构建体表达后,car从huegfrt上切割下来。该构建体的表达由人延伸因子1α(ef1α)启动子驱动。所述car包括抗原结合区、cd8α铰链区、cd8α跨膜(tm)结构域、4-1bb共刺激区和cd3ζ信号传导结构域。所述huegfrt包括两个胞外结构域(结构域iii和结构域iv)和一个tm结构域。图2a-2c为以下构建体的载体图谱:编码car.hn3的pmh288(图2a)、编码car.hyp7的pmh289(图2b)和编码car.lh7的pmh290(图2c)。图3a-3c是显示gpc3-靶向cart细胞的转导效率的流式细胞图。使用抗-huegfrt抗体西妥昔单抗(cetuximab)测定转导效率。编码car.hn3(图3a)和car.hyp7(图3b)的慢病毒载体分别转导65%和45.4%的t细胞。(图3c)对照人血清igg。图4a-4g是显示gpc3-靶向cart细胞对人细胞系的细胞毒性的图。使用1:2、1.5:1、5:1和16:1的效应子:靶比例,在gpc3+g1细胞(图4a)、gpc3+hep3b细胞(图4b)、gpc3+hepg2细胞(图4c)、gpc3+huh7细胞(图4d)、gpc3-a431细胞(图4e)、gpc3-t3m4细胞(图4f)和gpc3-imr32细胞(图4g)上测试car.hyp7。car.hyp7对gpc3-阳性细胞系有细胞毒性,但对gpc3-阴性细胞系没有细胞毒性。图5是显示car.hyp7t细胞诱导靶gpc-阳性hep3b、huh7和g1肿瘤细胞的干扰素(ifn)-γ分泌的图。图6显示在用gpc3-靶向t细胞治疗的小鼠中hep3b肿瘤抑制的生物发光图像。小鼠在第0天静脉内注射400万个hep3b细胞。在第10天,对小鼠进行空白注射或注射pbs、1000万个car.hn3t细胞(hn3-10m)、1000万个car.hyp7t细胞(hyp7-10m)、2000万个car.hyp7t细胞(hyp7-20m)或4000万个car.hyp7t细胞(hyp7-40m)。通过生物发光成像测量肿瘤尺寸。图7a-7c是显示car.hyp7t细胞对小鼠的hep3b异种移植肿瘤具有持久抗肿瘤活性的图。(图7a)在治疗后多至3周,用pbs治疗、空白治疗、用1000万个car.hn3t细胞、1000万car.hyp7t细胞、2000万car.hyp7t细胞或4000万car.hyp7t细胞治疗的荷hep3b肿瘤的小鼠的肿瘤体积。(图7b)在治疗后多至7周,用pbs、1000万个car.hn3t细胞,1000万个car.hyp7t细胞或4000万个car.hyp7t细胞治疗的荷hep3b肿瘤的小鼠的肿瘤体积。(图7c)荷hep3b肿瘤的小鼠的存活曲线。在hep3b接种后10天,向小鼠注射pbs、1000万个car.hyp7t细胞或4000万个car.hyp7t细胞,并对生存期进行评估70天。用4000万个car.hyp7t细胞的治疗可导致100%存活。图8a-8d是显示经空白治疗(mock-treated)(图8a)或用1000万个car.hyp7t细胞治疗(图8b)或4000万个car.hyp7t细胞治疗(图8c)的hepg2异种移植nsg小鼠的肿瘤体积的图。在图10a-10c中,每行代表单个动物。图8d显示了所有三个治疗组的平均肿瘤体积。图9显示通过定量实时pcr测量的人正常组织中gpc3mrna的水平。将不同正常组织中的相对gpc3水平与胎盘中gpc3的表达进行比较。图10a-10e显示gpc3cart细胞的产生和表达。(图10a)hn3和hyp7抗体分别与成熟gpc3的n-小叶(n-lobe)和c-小叶(c-lobe)结合的示意结构。(图10b)表达靶向gpc3的car与使用t2a核糖体跳跃序列的huegfrt的双顺反子慢病毒构建体的示意图。(图10c)使用流式细胞术通过检测egfr表达来分析用慢病毒颗粒转导的健康供体来源的t细胞上的car表达。(图10d)来自健康供体的空白t细胞和car(hyp7)t细胞以及hcc患者来源的car(hyp7)t细胞的cd3+、cd4+和cd8+t细胞群分析。(图10e)通过台盼蓝(trypanblue)排除测定评估,在8个不同的健康供体和4个不同的hcc患者中car(hyp7)t细胞的增殖。图11a-11g是显示gpc3-靶向cart细胞在体外杀死gpc3阳性hcc细胞的图。(图11a)如通过荧光素酶活性所测量的,gpc3-阳性靶细胞裂解(g1),但gpc3-阴性靶细胞不裂解(a431和t3m4)。空白或gpc3-靶向cart细胞与表达荧光素酶的靶细胞在指定的效应子(e):靶(t)比例下共培养24小时,并且使用基于荧光的细胞溶解测定来测量特异性裂解。(图11b-11c)与hep3b细胞共培养24小时后,来自健康供体(图11b)或hcc患者(图11c)的car(hn3)t细胞和car(hyp7)t细胞的细胞溶解活性。(图11d)用抗-cd3/cd28珠子刺激后,car(hyp7)t细胞强增殖35天。(图11e)与hep3b细胞共培养24小时后,car(hyp7)t细胞在激活后第14天和第28天的细胞溶解活性。(图11f)与hepg2和huh-7细胞共培养24小时后,健康供体来源的gpc3特异性cart细胞的细胞溶解活性。(图11g)使用incucytezoom测定的gpc3-靶向cart细胞-介导的hepg2细胞的杀伤。将hepg2细胞与cart细胞以2:1的e:t比例孵育长达140小时。图12显示了用gpc3-car重定向的t细胞的细胞因子/趋化因子谱和多功能性。将hep3b和hepg2细胞与gpc3-靶向cart细胞以各种e:t比例共培养24小时,并使用luminex测量上清液中指定的细胞因子/趋化因子水平。条形从左到右:空白、hyp7和hn3。显示平均和sd。*p<0.05;**p<0.01;***p<0.001。图13a-13f显示靶向gpc3通过抑制wnt/β-连环蛋白信号传导来诱导hcc细胞凋亡。(图13a)治疗6小时后,car(hyp7)t细胞抑制hep3b细胞中β-连环蛋白的表达并增加凋亡标记物的表达(裂解的parp和裂解的caspase-9)。(图13b)car(hyp7)t细胞以时间依赖的方式抑制hep3b细胞中β-连环蛋白的表达。(图13c)crispr/cas9介导的gpc3敲除后,hep3b细胞中的gpc3蛋白表达。(图13d)靶向gpc3的外显子5的sgrna5-2的抗肿瘤活性。将无胸腺nu/nu裸鼠皮下接种5×106hep3b细胞。当肿瘤的平均体积达到150mm3时,每隔一天通过肿瘤内注射sgrna5-2质粒或空载体来治疗小鼠,注射6次。(图13e)gpc3的敲除降低了小鼠肿瘤中β-连环蛋白的表达。(图13f)sgrna5-2质粒或空载体对照治疗前后的血清afp水平。显示平均和sd。*p<0.05;**p<0.01。图14a-14f显示car(hyp7)t细胞在hep3b腹膜播散(peritonealdissemination)异种移植小鼠模型中根除肿瘤。(图14a)实验示意图。在肿瘤细胞接种后第12天,对荷hep3b肿瘤的nsg小鼠腹膜注射空白t细胞、5×106car(hn3)t细胞、5×106car(hyp7)t细胞、10×106car(hyp7)t细胞或20×106car(hyp7)t细胞来进行治疗。通过生物发光成像监测肿瘤负荷。(图14b)car(hyp7)t细胞在高剂量时(2,000万个细胞)退缩了已建立的hep3b异种移植,并且在低剂量时(500万或1,000万个细胞)抑制了肿瘤生长。(图14c)图14b中治疗的小鼠中肿瘤生物发光,以平均光子计数。(图14d)用500万或2000万个car(hyp7)t细胞治疗后荷瘤小鼠的kaplan-meier生存曲线。(图14e)cart治疗后两周或六周,从图14b所示的组中收集的血清中的甲胎蛋白水平。收集每组三只不同小鼠的血清用于elisa分析。(图14f)如通过微滴式数字pcr(ddpcr)所测量的,治疗3周后异种移植肿瘤组织中的cart细胞持久性。值代表平均值±sd。*p<0.05;**p<0.01;***p<0.001。图15a-15d显示car(hyp7)t细胞在hepg2腹膜播散异种移植小鼠模型中消除了肿瘤细胞。(图15a)实验示意图。在第21天,对荷hep3b肿瘤的nsg小鼠腹膜注射空白t细胞或20×106car(hyp7)t细胞来进行治疗。(图15b)car(hyp7)t细胞证实强效的抗肿瘤活性和hepg2异种移植肿瘤的介导根除。(图15c)图15b中治疗的小鼠中肿瘤生物发光,以平均光子计数。(图15d)如通过ddpcr所测量的,治疗5周后异种移植肿瘤组织和小鼠脾脏中的cart细胞持久性。图16a-16d显示了在hep3b原位异种移植小鼠模型中通过car(hyp7)t细胞的hcc根除。(图16a)实验示意图。在第21天,向荷hep3b原位肿瘤的nsg小鼠腹膜内或静脉内注射20×106car(hyp7)t细胞。每周对小鼠进行成像。(图16b)经尾静脉用car(hyp7)t细胞治疗的小鼠证实肿瘤根除,而腹膜内治疗导致肿瘤生长抑制。(图16c)图16b中治疗的小鼠中肿瘤生物发光,以平均光子计数。(图16d)如通过ddpcr所测量的,治疗5周后肿瘤组织和小鼠脾脏中的cart细胞持久性。值代表平均值±sd。**p<0.01。图17是一系列流式细胞图和scatchard图,其显示了gpc3-靶向(hn3和hyp7)jurkatcart细胞与gpc3-人fc(hfc)融合蛋白的结合。图18是一系列图,其显示了在gpc3-靶向cart细胞与hep3b和hepg2肿瘤细胞孵育24小时后,通过luminex测量的差异化细胞因子和趋化因子分泌。条形从左到右:空白、hyp7和hn3。*p<0.05,**p<0.01,***p<0.001。图19a-19b显示用gpc3-靶向cart细胞治疗后hep3b和hepg2肿瘤异种移植小鼠的体重。(图19a)腹膜内注射pbs、空白t-细胞、car(hyp7)t细胞或car(hn3)t细胞后hep3b肿瘤模型小鼠的体重。(图19b)腹膜内注射2000万个空白t细胞或car(hyp7)t细胞后,hepg2肿瘤模型小鼠的体重。序列表使用37c.f.r.1.822中定义的标准核苷酸碱基字母缩写和氨基酸三字母代码,显示所附序列表中列出的核酸和氨基酸序列。每个核酸序列仅示出一条链,但是应理解互补链通过任意参考示出的链而被包括在内。序列表以ascii文本文件提交,于2018年10月29日生成,59.3kb,其通过引用并入本文。在所附序列表中:seqidno:1是编码gmcsfrss的核苷酸序列。seqidno:2是gmcsfrss的氨基酸序列。seqidno:3是编码cd8α铰链的核苷酸序列。seqidno:4是cd8α铰链的氨基酸序列。seqidno:5是编码cd8α跨膜结构域的核苷酸序列。seqidno:6是cd8α跨膜结构域的氨基酸序列。seqidno:7是编码4-1bb的核苷酸序列。seqidno:8是4-1bb的氨基酸序列。seqidno:9是编码cd3ζ的核苷酸序列。seqidno:10是cd3ζ的氨基酸序列。seqidno:11是编码自裂解t2a肽的核苷酸序列。seqidno:12是自裂解t2a肽的氨基酸序列。seqidno:13是编码huegfrt的核苷酸序列。seqidno:14是huegfrt的氨基酸序列。seqidno:15是编码car.hyp7的核苷酸序列,具有以下特征:核苷酸1-66=gmcsfrss编码序列核苷酸67-72=ndei限制位点核苷酸73-807=人源化yp7scfv编码序列核苷酸808-813=spei限制位点核苷酸814-948=cd8α铰链区编码序列核苷酸949-1011=cd8α跨膜结构域编码序列核苷酸1012-1137=4-1bb共刺激结构域编码序列核苷酸1138-1473=cd3ζ信号传导结构域编码序列核苷酸1474-1527=t2a编码序列核苷酸1528-1593=gmcsfrss编码序列核苷酸1594-2598=huegfrt编码序列。seqidno:16是car.hyp7的氨基酸序列,具有以下特征:残基1-22=gmcsfrss残基23-24=hm(由ndei限制位点编码)残基25-269=人源化yp7scfv残基270-271=ts(由spei限制位点编码)残基272-316=cd8α铰链区残基317-337=cd8α跨膜结构域残基338-379=4-1bb共刺激结构域残基380-491=cd3ζ信号传导结构域残基492-509=自裂解t2a肽残基510-531=gmcsfrss残基532-866=huegfrt编码序列。seqidno:17是编码car.hn3的核苷酸序列,具有以下特征:核苷酸1-66=gmcsfrss编码序列核苷酸67-72=ndei限制位点核苷酸73-420=hn3编码序列核苷酸421-426=spei限制位点核苷酸427-561=cd8α铰链区编码序列核苷酸562-624=cd8α跨膜结构域编码序列核苷酸625-750=4-1bb共刺激结构域编码序列核苷酸751-1086=cd3ζ信号传导结构域编码序列核苷酸1087-1140=t2a编码序列核苷酸1141-1206=gmcsfrss编码序列核苷酸1207-2211=huegfrt编码序列。seqidno:18是car.hn3的氨基酸序列,具有以下特征:残基1-22=gmcsfrss残基23-24=hm(由ndei限制位点编码)残基25-140=hn3单域抗体残基141-142=ts(由spei限制位点编码)残基143-187=cd8α铰链区残基188-208=cd8α跨膜结构域残基209-250=4-1bb共刺激结构域残基251-362=cd3ζ信号传导结构域残基363-380=自裂解t2a肽残基381-402=gmcsfrss残基403-737=huegfrt编码序列。seqidno:19是编码car.lh7的核苷酸序列,具有以下特征:核苷酸1-66=gmcsfrss编码序列核苷酸67-72=ndei限制位点核苷酸73-432=lh7编码序列核苷酸433-438=spei限制位点核苷酸439-573=cd8α铰链区编码序列核苷酸574-636=cd8α跨膜结构域编码序列核苷酸637-762=4-1bb共刺激结构域编码序列核苷酸763-1098=cd3ζ信号传导结构域编码序列核苷酸1099-1152=t2a编码序列核苷酸1153-1218=gmcsfrss编码序列核苷酸1219-2223=huegfrt编码序列.seqidno:20是car.lh7的氨基酸序列,具有以下特征:残基1-22=gmcsfrss残基23-24=hm(由ndei限制位点编码)残基25-144=lh7单域抗体残基145-146=ts(由spei限制位点编码)残基147-191=cd8α铰链区残基192-212=cd8α跨膜结构域残基213-254=4-1bb共刺激结构域残基255-366=cd3ζ信号传导结构域残基367-384=自裂解t2a肽残基385-406=gmcsfrss残基407-741=huegfrt编码序列。seqidno:21是yp7vh结构域的核苷酸序列。seqidno:22是yp7vh结构域的氨基酸序列。seqidno:23是yp7vl结构域的核苷酸序列。seqidno:24是yp7vl结构域的氨基酸序列。seqidno:25是hyp7vh结构域的核苷酸序列。seqidno:26是hyp7vh结构域的氨基酸序列。seqidno:27是hyp7vl结构域的核苷酸序列。seqidno:28是hyp7vl结构域的氨基酸序列。seqidno:29是hn3单域抗体的核苷酸序列。seqidno:30是hn3单域抗体的氨基酸序列。seqidno:31是lh7单域抗体的核苷酸序列。seqidno:32是lh7单域抗体的氨基酸序列。seqidno:33是lh4单域抗体的核苷酸序列。seqidno:34是lh4单域抗体的氨基酸序列。seqidno:35是lh6单域抗体的核苷酸序列。seqidno:36是lh6单域抗体的氨基酸序列。seqidno:37是yp218vh结构域的核苷酸序列。seqidno:38是yp218vh结构域的氨基酸序列。seqidno:39是yp218vl结构域的核苷酸序列。seqidno:40是yp218vl结构域的氨基酸序列。seqidno:41是sd1单域抗体的核苷酸序列。seqidno:42是sd1单域抗体的氨基酸序列。seqidno:43-51是sgrna序列。发明详述i.缩写adcc抗体依赖性细胞介导的细胞毒性(antibody-dependentcell-mediatedcytotoxicity)car嵌合抗原受体(chimericantigenreceptor)cdr互补决定区(complementaritydeterminingregion)ctl细胞毒性t淋巴细胞(cytotoxictlymphocyte)ddpcr微滴式数字pcr(dropletdigitalpcr)dmemdulbecco改良的eagle培养基(dulbecco’smodifiedeaglemedium)ef1α延伸因子1α(elongationfactor1alpha)egf表皮生长因子(epidermalgrowthfactor)egfr表皮生长因子受体(epidermalgrowthfactorreceptor)elisa酶联免疫吸附测定(enzyme-linkedimmunosorbentassay)facs荧光激活细胞分选(fluorescenceactivatedcellssorting)fbs胎牛血清(fetalbovineserum)gpc2glypican-2gpc3glypican-3gmcsfrss粒细胞-巨噬细胞集落刺激因子受体信号序列(granulocyte-macrophagecolonystimulatingfactorreceptorsignalsequence)hcc肝细胞癌(hepatocellularcarcinoma)hla人白细胞抗原(humanleukocyteantigen)huegfrt人截短的表皮生长因子受体(humantruncatedepidermalgrowthfactorreceptor)ifn干扰素(interferon)ig免疫球蛋白(immunoglobulin)il白细胞介素(interleukin)i.p.腹膜内(intraperitoneal)itam免疫受体酪氨酸激活基序(immunoreceptortyrosine-basedactivationmotif)pbmc外周血单个核细胞(peripheralbloodmononuclearcell)pbs磷酸盐缓冲液(phosphate-bufferedsaline)scfv单链可变片段(single-chainvariablefragment)tm跨膜(transmembrane)vh或vh重链可变(variableheavy)vl或vl轻链可变(variablelight)yst卵黄囊肿瘤(yolksactumor)ii.术语和方法除非另外指出,技术术语根据常规用法使用。分子生物学常用术语的定义可见于以下文献:benjaminlewin,genesv,publishedbyoxforduniversitypress,1994(isbn0-19-854287-9);kendrewetal.(eds.),theencyclopediaofmolecularbiology,publishedbyblackwellscienceltd.,1994(isbn0-632-02182-9);和roberta.meyers(ed.),molecularbiologyandbiotechnology:acomprehensivedeskreference,publishedbyvchpublishers,inc.,1995(isbn1-56081-569-8)。为了便于审阅本公开的各种实施方式,提供对特定术语的下述解释:4-1bb:由t细胞受体(tcr)-激活的淋巴细胞和包括自然杀伤细胞在内的其他细胞表达的共刺激分子。4-1bb的连接诱导信号级联(signalingcascade),所述信号级联导致细胞因子产生,抗凋亡分子的表达和增强的免疫应答。急性淋巴细胞白血病(acutelymphoblasticleukemia,all):急性形式的白血病,以淋巴母细胞过量产生为特征。all在儿童中最常见,在2-5岁的儿童中达到峰值。抗体:一种包含至少一个可变区的多肽配体,其识别和结合(例如特异性识别和特异性结合)抗原的表位。哺乳动物免疫球蛋白分子由重(h)链和轻(l)链组成,它们各自具有可变区,分别称为重链可变(vh)区和轻链可变(vl)区。vh区和vl区共同负责结合抗体识别的抗原。有五种主要重链类别(或同种型)的哺乳动物免疫球蛋白,它们决定抗体分子igm、igd、igg、iga和ige的功能活性。在哺乳动物中未发现的抗体同种型包括igx、igy、igw和ignar。igy是鸟类和爬行动物产生的主要抗体,并且在功能上与哺乳动物igg和ige有些相似。igw和ignar抗体由软骨鱼(cartilaginousfish)产生,而igx抗体则在两栖动物中发现。抗体可变区包含“框架”区和高变区,称为“互补决定区”或“cdr”。cdr主要负责与抗原的表位结合。抗体的框架区用于在三维空间中定位和对齐cdr。可以使用多种已知的编号方案中的任何一种容易地确定给定cdr的氨基酸序列边界,包括kabat等(sequencesofproteinsofimmunologicalinterest,u.s.departmentofhealthandhumanservices,1991;the“kabat”numberingscheme),chothia等(参见chothiaandlesk,jmolbiol196:901-917,1987;chothiaetal.,nature342:877,1989;和al-lazikanietal.,(jmb273,927-948,1997;the“chothia”numberingscheme)和immunogenetics(imgt)数据库(参见lefranc,nucleicacidsres29:207-9,2001;the“imgt”numberingscheme)中所描述的那些。kabat和imgt数据库是在线维护的。“单域抗体(single-domainantibody)”是指具有单个结构域(可变结构域)的抗体,其能够在不存在其他抗体结构域的情况下特异性结合抗原或抗原的表位。单域抗体例如包括vh结构域抗体、vnar抗体、骆驼科动物(camelid)vhh抗体和vl结构域抗体。vnar抗体由软骨鱼产生,例如护士鲨(nurseshark),须鲨(wobbegongshark),白斑角鲨(spinydogfish)和竹鲨(bambooshark)。骆驼科动物vhh抗体是由几种物种产生的,包括骆驼、美洲驼(llama)、羊驼(alpaca)、单峰驼(dromedary)和原驼(guanaco),它们产生天然缺少轻链的重链抗体。“单克隆抗体”是由淋巴细胞的单个克隆或由其中已经转染了单个抗体的编码序列的细胞产生的抗体。抗体通过本领域技术人员已知的方法产生。单克隆抗体包括人源化单克隆抗体。“嵌合抗体”具有来自一种物种例如人的框架残基,和来自另一种物种的cdr(其通常赋予抗原结合)。“人源化”抗体是包括人框架区和一个或更多个来自非人(例如小鼠、兔、大鼠、鲨鱼或合成的)免疫球蛋白的cdr。提供cdr的非人免疫球蛋白被称为“供体”,提供框架的人免疫球蛋白被称为“受体”。在一个实施方式中,人源化免疫球蛋白中的所有cdr均来自供体免疫球蛋白。恒定区无需存在,但如果存在的话,则它们必须与人免疫球蛋白恒定区基本相同,即,具有至少约85-90%,例如约95%或更高的同一性。因此,人源化免疫球蛋白的所有部分,可能除了cdr以外,与天然人免疫球蛋白序列的相应部分基本相同。人源化抗体与提供cdr的供体抗体结合相同的抗原。人源化或其他单克隆抗体可以具有其他保守的氨基酸取代,这些保守的氨基酸取代对抗原结合或其他免疫球蛋白功能基本没有影响。结合亲和力:抗体对抗原的亲和力。在一个实施方式中,亲和力通过frankeletal.,mol.immunol.,16:101-106,1979描述的改进的scatchard方法来计算。在另一个实施方式中,结合亲和力通过抗原/抗体解离速率来测量。在另一个实施方式中,高结合亲和力通过竞争放射免疫测定来测量。在另一个实施方式中,结合亲和力通过elisa测量。在另一个实施方式中,抗体亲和力通过流式细胞术测量。“特异性结合”抗原(例如gpc3)的抗体是以高亲和力结合该抗原并且不会明显结合其他不相关抗原的抗体。乳腺癌:一种在乳腺组织中形成的癌症,所述乳腺组织通常是导管(将乳汁输送到乳头的管)和小叶(制造乳汁的腺体)。三阴性乳腺癌是指其中癌细胞不表达雌激素受体、孕酮受体或显著水平的her2/neu蛋白的乳腺癌类型。三阴性乳腺癌也称为er阴性pr阴性her2/neu-阴性乳腺癌。化疗剂:任何在治疗以异常细胞生长为特征的疾病中具有治疗作用的化学试剂。这些疾病包括肿瘤、赘生物(neoplasm)和癌症,以及以增生性生长为特征的疾病,例如牛皮癣。在一个实施方式中,化疗剂是一种放射性化合物。本领域技术人员可以很容易地确定使用的化疗剂(参见,例如slapakandkufe,principlesofcancertherapy,chapter86inharrison'sprinciplesofinternalmedicine,14thedition;perryetal.,chemotherapy,ch.17inabeloff,clinicaloncology2nded.,2000churchilllivingstone,inc;baltzer,l.,berkery,r.(eds.):oncologypocketguidetochemotherapy,2nded.st.louis,mosby-yearbook,1995;fischer,d.s.,knobf,m.f.,durivage,h.j.(eds):thecancerchemotherapyhandbook,4thed.st.louis,mosby-yearbook,1993)。联合化疗是施用不止一种试剂来治疗癌症。一个实例是施用与放射性或化学化合物组合使用的cart细胞。嵌合抗原受体(car):包含抗原结合部分(例如单域抗体或scfv)和信号传导结构域(例如来自t细胞受体的信号传导结构域(例如cd3ζ))的嵌合分子。通常,car由抗原结合部分、跨膜结构域和胞内结构域组成。胞内结构域通常包括具有免疫受体酪氨酸激活基序(itam)的信号传导链,例如cd3ζ或fcεriγ。在一些情况下,胞内域还包括至少一个其他共刺激结构域的胞内部分,例如cd28、4-1bb(cd137)、icos、ox40(cd134)、cd27和/或dap10。胆管癌(cholangiocarcinoma):在肝胆管内壁细胞中发生的癌症类型。互补决定区(cdr):确定抗体的结合亲和力和特异性的高变氨基酸序列区域。哺乳动物免疫球蛋白的轻链和重链各自具有三个cdr,分别称为l-cdr1、l-cdr2、l-cdr3和h-cdr1、h-cdr2、h-cdr3。保守变体(conservativevariant):“保守”氨基酸替代是那些基本上不影响或降低蛋白例如抗体对gpc3的亲和力的替代。例如,特异性结合gpc3的单克隆抗体可包括至多约1、至多约2、至多约5、至多约10或至多约15个保守替代,并特异性结合gpc3多肽。术语“保守变体”还包括使用取代的氨基酸代替未取代的亲本氨基酸,只要该变体保留活性。非保守取代是降低蛋白质活性的那些。提供功能上相似的氨基酸的保守氨基酸替代表是本领域普通技术人员熟知的。以下六组是被认为是彼此的保守替代的氨基酸示例:1)丙氨酸(a)、丝氨酸(s)、苏氨酸(t);2)天冬氨酸(d)、谷氨酸(e);3)天冬酰胺(n)、谷氨酰胺(q);4)精氨酸(r)、赖氨酸(k);5)异亮氨酸(i)、亮氨酸(l)、甲硫氨酸(m)、缬氨酸(v);和6)苯丙氨酸(f)、酪氨酸(y)、色氨酸(w)。在本文的一些实施方式中,提供了氨基酸序列,所述氨基酸序列相对于seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:12、seqidno:14、seqidno:16、seqidno:18、seqidno:20、seqidno:22、seqidno:24、seqidno:26、seqidno:28、seqidno:30或seqidno:32包括不超过10、不超过9、不超过8、不超过7、不超过6、不超过5、不超过4、不超过3、不超过2或不超过1个氨基酸替代。细胞毒性剂:杀死细胞的任何药物或化合物。细胞毒性:分子对预期被靶向的细胞(相对于生物体其余部分的细胞)的毒性。简并变体:编码包含由于遗传密码而简并的序列的多肽的多核苷酸。有20种天然氨基酸,其中大多数由一个以上的密码子指定。因此,只要多肽的氨基酸序列不变,所有简并的核苷酸序列都包括在内。促结缔组织增生性小圆细胞肿瘤(desmoplasticsmallroundcelltumor,drct):主要发生在儿童时期特别是在男孩中的软组织肉瘤。drct是一种侵袭和罕见型癌症,其主要以腹部肿块的形式出现,但也可以在淋巴结、腹部内膜、横膈膜、脾脏、肝脏、胸壁、颅骨、脊髓、肠道、膀胱、脑、肺、睾丸、卵巢和骨盆中发现。表位:抗原决定簇。这些是分子上具有抗原性,即引起特异性免疫应答的特定化学基团或肽序列。抗体特异性结合多肽上的特定抗原表位。尤文氏肉瘤(ewing’ssarcoma):骨或软组织中发现的罕见类型的恶性肿瘤。尤文氏肉瘤是一种小的蓝色圆形细胞瘤。框架区(frameworkregion):介于cdr之间的氨基酸序列。构架区包括轻链可变和重链可变框架区。框架区用于将cdr保持在抗原结合的适合方向上。融合蛋白:包含两种不同(异源)蛋白的至少一部分的蛋白。glypican-2(gpc2):硫酸乙酰肝素(heparansulfate,hs)蛋白聚糖的六成员glypican家族的成员,所述hs蛋白聚糖通过gpi锚点附着在细胞表面(filmusetal.,genomebiol9:224,2008)。gpc2独特表达于神经系统中(stippetal.,jcellbiol124:149-160,1994),参与细胞粘附并被认为可调节轴突的生长和引导。此外,gpc2mrna高度表达于神经母细胞瘤和其他小儿癌症中(orentasetal.,frontoncol2:194,2012)。gpc2也称为脑聚糖(cerebroglycan)蛋白聚糖和glypican蛋白聚糖2。gpc2基因组、mrna和蛋白质序列是可公开获得的(参见,例如ncbigeneid221914)。gpc2-阳性癌症:过表达gpc2的癌症。gpc2-阳性癌症的实例包括,但不限于神经母细胞瘤、急性淋巴细胞白血病、胚胎横纹肌肉瘤、腺泡状横纹肌肉瘤、尤文氏肉瘤、促结缔组织增生性小圆细胞肿瘤或骨肉瘤。glypican-3(gpc3):硫酸乙酰肝素(hs)蛋白聚糖的glypican家族的成员,所述hs蛋白聚糖通过糖基磷脂酰肌醇锚点附着在细胞表面(filmusandselleck,jclininvest108:497-501,2001)。gpc3基因编码大约70kd的核心蛋白,该蛋白可以被弗林蛋白酶切割以产生40kdn端片段和30kdc端片段。两个hs链连接在gpc3的c端部分。gpc3和其他glypican家族蛋白在细胞分裂和细胞生长调节中起作用。gpc3高度表达于hcc和一些其他人类癌症中,包括黑色素瘤、肺鳞状细胞癌和卵巢透明细胞癌(hoandkim,eurjcancer47(3):333-338,2011),但不表达于正常组织中。gpc3也称为sgb、dgsx、mxr7、sdys、sgbs、oci-5、sgbs1和gtr2-2。人gpc3有四种已知的同种型(同种型1-4)。gpc3的四种同种型的核酸和氨基酸序列是已知的,包括genbank登录号:nm_001164617和np_001158089(同种型1);nm_004484和np_004475(同种型2);nm_001164618和np_001158090(同种型3);以及nm_001164619和np_001158091(同种型4)。gpc3-阳性癌症:过表达gpc3的癌症。gpc3-阳性癌症的实例包括,但不限于hcc、黑色素瘤、卵巢透明细胞癌、卵黄囊肿瘤(yst)、神经母细胞瘤、肝母细胞瘤、wilms肿瘤、肺鳞状细胞癌、睾丸非精原细胞性生殖细胞瘤、脂肪肉瘤、宫颈上皮内瘤变、肾上腺腺瘤、神经鞘瘤和胚胎肿瘤(hoandkim,eurjcancer47(3):333-338,2011;baumhoeretal.,amjclinpathol129(6):899-906,2008;saikaliandsinnett,intjcancer89(5):418-422,2000)。hama(人类抗鼠抗体(humananti-murineantibody))应答:人受试者对已经施用于该患者的鼠抗体的可变和恒定区的免疫应答。重复施用抗体可能会导致抗体从患者血清中清除的速率增加,并且也可能引起患者过敏反应。肝细胞癌(hepatocellularcarcinoma,hcc):肝脏的原发性恶性肿瘤通常发生在具有炎症性肝的患者中,所述炎症性肝由病毒性肝炎、肝毒素或肝硬化(通常由酒精中毒引起)引起。hcc也称为恶性肝癌。异源的(heterologous):来源于单独的遗传资源或物种。免疫反应:免疫系统的细胞(例如b细胞、t细胞或单核细胞)对刺激的反应。在一个实施方式中,该反应对特定抗原是特异性的(“抗原-特异性反应”)。在一个实施方式中,免疫反应是t细胞反应,例如cd4+反应或cd8+反应。在另一个实施方式中,该反应是b细胞反应,并且导致特异性抗体的产生。分离的:“分离的”生物组分(例如核酸、蛋白质(包括抗体)或细胞器)已经与该组分自然存在的环境(例如细胞)中的其他生物组分(即,其他染色体和染色体外的dna和rna、蛋白质和细胞器)基本分离或从其中纯化出来。已经“分离”的核酸和蛋白质包括通过标准纯化方法纯化的核酸和蛋白质。该术语还涵盖通过在宿主细胞中重组表达制备的核酸和蛋白质,以及化学合成的核酸。标记物(label):与另一分子(例如抗体或蛋白)直接或间接缀合以促进该分子检测的可检测化合物或组合物。标记物的非限制性具体实例包括荧光标签、酶连接和放射性同位素。在一个实例中,“标记的抗体”是指在抗体中引入另一种分子。例如,标记物是可检测标记物,例如掺入放射性标记的氨基酸或与可通过标记的亲和素(例如,可通过光学或比色法检测的含有荧光标记或酶活性的链霉亲和素)检测的生物素部分多肽连接。标记多肽和糖蛋白的各种方法是本领域已知的并且可以使用。用于多肽的标记物的例子包括但不限于以下:放射性同位素或放射性核苷酸(例如35s、11c、13n、15o、18f、19f、99mtc、131i、3h、14c、15n、90y、99tc、111in和125i),荧光标记物(如异硫氰酸荧光素(fitc)、若丹明、镧系荧光剂),酶标记物(如辣根过氧化物酶、β-半乳糖苷酶、荧光素酶、碱性磷酸酶),化学发光标记,生物素基团,被二级报告分子识别的预先确定的多肽表位(例如亮氨酸拉链对序列、第二抗体的结合位点、金属结合结构域、表位标签),或磁性试剂,例如钆螯合物。在一些实施方式中,标记物通过各种长度的间隔臂连接,以减少潜在的空间位阻。接头(linker):在一些情况下,接头是抗体结合片段(例如fv片段)内的肽,其用于将可变重链间接结合至可变轻链。“接头”也可以指用于将靶向部分例如抗体与效应分子例如细胞毒素或可检测标记物连接的肽。术语“缀合(conjugating)”,“结合(joining)”,“键合(bonding)”或“连接(linking)”是指使两个多肽成为一个连续的多肽分子,或将放射性核素或其他分子共价附接于多肽,例如scfv。在特定上下文中,该术语包括提及将配体例如抗体部分连接至效应分子。连接可以通过化学或重组方式进行。“化学方式”是指抗体部分和效应分子之间的反应,使得在两个分子之间形成共价键以形成一个分子。肺癌:在肺组织中,通常在气道衬里细胞中形成的癌症。两种主要类型是小细胞肺癌和非小细胞肺癌(nsclc)。根据细胞在显微镜下的外观诊断这些类型。哺乳动物:该术语包括人类和非人类哺乳动物。类似地,术语“受试者”包括人类和兽医学受试者。黑色素瘤(melanoma):一种起源于黑色素细胞(产生黑色素这种色素的细胞)的癌症。黑色素细胞主要在皮肤中发现,但也存在于肠和眼中。皮肤中的黑色素瘤包括浅表扩散性黑色素瘤、结节性黑色素瘤、肢端雀斑样黑色素瘤和恶性雀斑痣(黑色素瘤)。以上任何一种类型都可能产生黑色素或可能是无黑色素的。同样,任何亚型都可能显示出结缔织生成(具有向神经性的密集纤维反应),这是侵袭性行为和局部复发趋势的标志。其他黑色素瘤包括透明细胞肉瘤,粘膜黑色素瘤和葡萄膜黑色素瘤。间皮素(mesothelin):40kda细胞表面糖基磷脂酰肌醇(gpi)-连接的糖蛋白。人间皮素蛋白被合成为70kd的前体,然后进行蛋白水解加工。间皮素的30kd氨基末端被分泌出来,并且被称为巨核细胞增强因子(megakaryocytepotentiatingfactor)(yamaguchietal.,j.biol.chem.269:805808,1994)。作为成熟的间皮素,40kd羧基末端保持与膜结合(changetal.,natl.acad.sci.usa93:136140,1996)。间皮素的示例性核酸和氨基酸序列描述于pct公开号wo97/25,068;美国专利第6,083,502号;changandpastan,int.j.cancer57:90,1994;changandpastan,proc.natl.acad.sciusa93:136,1996;brinkmannetal.,int.j.cancer71:638,1997;andchowdhuryetal.,mol.immunol.34:9,1997。间皮素还指保留在细胞内的间皮素蛋白或多肽以及分泌和/或分离的细胞外间皮素蛋白。间皮素-阳性癌症:过表达间皮素的癌症。间皮素-阳性癌症的实例包括,但不限于间皮瘤、前列腺癌、肺癌、胃癌、鳞状细胞癌、胰腺癌、胆管癌、三阴性乳腺癌和卵巢癌。间皮瘤(mesothelioma):一类来源于胸膜和腹膜衬里细胞的赘生物,其生长成覆盖内脏的厚片,由梭状细胞或纤维组织组成,其可能包围内衬有立方体状细胞的腺样空间。间皮瘤通常起源于肺、心脏或腹部的衬里组织。在某些情况下,间皮瘤是由接触石棉引起的。赘生物、恶性肿瘤、癌症或肿瘤(neoplasia,malignancy,cancerortumor):赘生物是由于过度的细胞分裂导致的组织或细胞的异常生长。赘生性生长可产生肿瘤。个体中肿瘤的量就是“肿瘤负荷(tumorburden)”,其可以用肿瘤的数量、体积或重量来衡量。不转移的肿瘤被称为“良性的”。侵入周围组织和/或可以转移的肿瘤被称为“恶性的”。神经母细胞瘤(neuroblastoma):由胚胎神经嵴细胞引起的实体瘤。神经母细胞瘤通常发生在肾上腺及其周围,但也可能发生在发现交感神经组织的任何地方,例如在腹部、胸部、颈部或脊柱附近的神经组织中。神经母细胞瘤通常发生在5岁以下的儿童中。可操作连接(operablylinked):当第一核酸序列与第二核酸序列处于功能关系时,第一核酸序列与第二核酸序列可操作连接。例如,如果启动子影响编码序列的转录或表达,则启动子与编码序列可操作连接。通常,可操作连接的dna序列是连续的,并且在需要连接两个蛋白编码区的情况下,在相同的阅读框中。骨肉瘤(osteosarcoma):在骨中发现的癌性肿瘤类型。骨肉瘤是一种侵袭性癌症,其起源于间充质来源的原生转化细胞。这类癌症在儿童和年轻人中最为普遍。卵巢癌(ovariancancer):在卵巢(在其中形成卵子或卵的一对雌性生殖腺中的一个)组织中形成的癌症。大多数卵巢癌要么是卵巢上皮癌(始于卵巢表面细胞的癌症),要么是恶性生殖细胞瘤(始于卵细胞的癌症)。卵巢透明细胞癌(ovarianclearcellcarcinoma):上皮性卵巢癌的独特组织病理学亚型,其发生率不到所有卵巢恶性肿瘤的5%。在显微镜下观察时,这种类型的肿瘤细胞内部看起来是透明的。胰腺癌(pancreaticcancer):在胰腺组织中发现恶性(癌)细胞的疾病。也称为外分泌癌。小儿癌症(pediatriccancer):在0至14岁的儿童中发生的癌症。小儿癌症的主要类型包括,例如,神经母细胞瘤、急性淋巴细胞白血病(all)、胚胎横纹肌肉瘤(erms)、腺泡状横纹肌肉瘤(arms)、尤文氏肉瘤、促结缔组织增生性小圆细胞肿瘤(drct)、骨肉瘤、脑和其他cns肿瘤、wilm肿瘤、非霍奇金淋巴瘤和视网膜母细胞瘤。药剂:当适当地施用于受试者或细胞时能够诱导所需治疗或预防效果的化学化合物或组合物。药学可接受的载体:使用的药学可接受的载体是常规的。remington’spharmaceuticalsciences,bye.w.martin,mackpublishingco.,easton,pa,15thedition,1975描述了适用于药学递送本文公开的组合物的组合物和制剂。通常,载体的性质将取决于所采用的具体施用方式。例如,肠胃外制剂通常包括可注射流体,其包括药学上和生理学上可接受的流体,例如水、生理盐水、平衡盐溶液、葡萄糖水溶液、甘油等作为媒介物。对于固体组合物(例如粉末、丸剂、片剂或胶囊形式),常规的非毒性固体载体可以包括,例如药用级甘露醇、乳糖、淀粉或硬脂酸镁。除生物中性载体之外,待施用的药物组合物可以含有少量无毒的辅助物质,例如润湿剂或乳化剂、防腐剂和ph缓冲剂等,例如乙酸钠或脱水山梨醇单月桂酸酯。预防、治疗或改善疾病(preventing,treatingoramelioratingadisease):“预防”疾病是指抑制疾病的全面发展。“治疗”是指改善疾病或病理状况开始发展后的体征或症状的治疗干预,例如减轻肿瘤负荷或减少转移灶的数量尺寸。“改善”是指疾病例如癌症的体征或症状在数量或严重性上的降低。前列腺癌(prostatecancer):在前列腺(位于膀胱下方和直肠前方的男性生殖系统的腺体)组织中形成的癌症。前列腺癌通常发生在老年男性中。纯化的:术语“纯化”不要求绝对的纯度,相反,它旨在作为一个相对术语。因此,例如,纯化的肽制品是这样的肽制品,其中肽或蛋白质比细胞内自然环境中的肽或蛋白质富集程度更高。在一个实施方式中,纯化制剂以使蛋白质或肽至少占制剂的总肽或蛋白质含量的50%。基本纯化是指从其他蛋白质或细胞组分中纯化。基本纯化的蛋白质是至少60%、70%、80%、90%、95%或98%纯的。因此,在一个特定的非限制性实例中,基本纯化的蛋白质是90%不含其他蛋白质或细胞组分。重组的:重组核酸是具有非天然存在序列的核酸或者是具有由序列的两个分离的片段人工组合而制成的序列的核酸。通常通过化学合成或通过人工操作分离的核酸片段,例如通过基因工程技术,来实现这种人工组合。横纹肌肉瘤(rhabdomyosarcoma,rms):骨骼肌来源的软组织恶性肿瘤。横纹肌肉瘤最常见的原发部位是头部和颈部(例如,脑膜旁、眼眶、咽等),泌尿生殖道和四肢。其他较不常见的原发部位包括躯干、胸壁、腹部(包括腹膜后腔和胆道)以及会阴/肛门区域。存在至少有两种类型的rms。最常见的形式是腺泡状rms(arms)和胚胎组织学rms(erms)。大约20%的患有横纹肌肉瘤的儿童具有arms亚型。在青少年以及原发部位涉及四肢、躯干和会阴/肛周区域的患者中,发现该亚型的频率增加。arms与编码融合基因的染色体易位相关,所述融合基因涉及13号染色体上的fkhr和pax家族成员。胚胎亚型是儿童中最常见的亚型,约占儿童横纹肌肉瘤的60-70%。具有胚胎组织学的肿瘤通常出现在头部和颈部或泌尿生殖道中,尽管它们可能发生在任何原发部位。erms的特征是诊断时年龄较小,杂合性丧失和基因组印迹改变。样品(或生物样品):从受试者获得的包含基因组dna、rna(包括mrna)、蛋白质或其组合的生物样本。实例包括但不限于外周血、组织、细胞、尿液、唾液、组织活检、细针抽吸物、手术样本和尸检材料。在一实例中,样品包括肿瘤活检,例如肿瘤组织活检。序列同一性:氨基酸或核酸序列之间的相似性以序列之间的相似性(或称为序列同一性)表示。序列同一性经常以同一性(或相似性或同源性)百分比来测量;百分比越高,两条序列越相似。当使用标准方法比对时,多肽或核酸分子的同源物或变体将具有相当高程度的序列同一性。用于进行比较的序列比对方法是本领域公知的。各种程序和比对算法描述于:smithandwaterman,adv.appl.math.2:482,1981;needlemanandwunsch,j.mol.biol.48:443,1970;pearsonandlipman,proc.natl.acad.sci.u.s.a.85:2444,1988;higginsandsharp,gene73:237,1988;higginsandsharp,cabios5:151,1989;corpetetal.,nucleicacidsresearch16:10881,1988;andpearsonandlipman,proc.natl.acad.sci.u.s.a.85:2444,1988。altschuletal.,naturegenet.6:119,1994陈述了序列比对方法和同源性计算的详细考虑。ncbi基本局部比对搜索工具(ncbibasiclocalalignmentsearchtool,blast)(altschuletal.,j.mol.biol.215:403,1990)可以从几个来源获得,这些来源包括国家生物技术信息中心(nationalcenterforbiotechnologyinformation,ncbi,bethesda,md)和互联网,与序列分析程序blastp、blastn、blastx、tblastn和tblastx结合使用。如何使用该程序确定序列同一性的描述可在互联网的ncbi网站上获得。特异性结合gpc3多肽的抗体的vl或vh的同源物和变体的特征通常在于,根据使用ncbiblast2.0,gappedblastp设置为默认参数的抗体氨基酸序列的全长比对计算,具有至少约75%,例如至少约80%、90%、95%、96%、97%、98%或99%的序列同一性。为了比较大于约30个氨基酸的氨基酸序列,使用设置为默认参数的默认blosum62矩阵((缺口存在扣分(gapexistencecost)为11,每残基缺口扣分(gapcost)为1))来使用blast2序列函数。当比对短肽(少于约30个氨基酸)时,比对应使用blast2序列函数进行,使用设定为默认参数的pam30矩阵(开放缺口9,延伸缺口1罚分)。当通过该方法评估时,与参考序列具有甚至更高相似性的蛋白质将会显示增加的同一性百分比,例如至少80%、至少85%、至少90%、至少95%、至少98%或至少99%的序列同一性。当比较少于完整序列的序列同一性时,同源物和变体通常将在10-20个氨基酸的短窗口内具有至少80%的序列同一性,并且根据它们与参考序列的相似性,可以具有至少85%或至少90%或95%的序列同一性。在互联网的ncbi网站上可以获得在这些短窗口上确定序列同一性的方法。本领域技术人员将会认识到,这些序列同一性范围仅作为指导提供;完全可能获得落在所提供范围之外的强显著性同源物。鳞状细胞癌(squamouscellcarcinoma):一种癌症,其起源于形成皮肤、眼睛、各种内部器官的表面,以及某些腺体的中空器官和导管的衬里的鳞状细胞、薄的扁平细胞。鳞状细胞癌也称为表皮样癌。一种鳞状细胞癌是肺鳞状细胞癌。鳞状细胞癌是最常见的皮肤癌类型。胃癌(stomachcancer):在胃壁组织中形成的癌症。也称为胃癌(gastriccancer)。受试者:活的多细胞脊椎动物生物体,包括人类和兽类受试者的类别,包括人类和非人类哺乳动物。合成的:在实验室中通过人工手段生产的,例如可以在实验室中通过化学合成的合成核酸或蛋白质(例如抗体)。治疗有效量:足以在被治疗的受试者中获得所需效果的特定物质的量。例如,这可以是抑制或阻止肿瘤生长所需的量。在一个实施方式中,治疗有效量是消除、减小肿瘤尺寸或预防肿瘤转移所必需的量。当施用于受试者时,通常将使用达到目标组织浓度(例如,在肿瘤中)的剂量,该目标组织浓度已显示实现所需的体外效果。载体:引入宿主细胞的核酸分子,由此产生转化的宿主细胞。载体可以包括允许其在宿主细胞中复制的核酸序列,例如复制起点。载体还可以包括一种或更多种选择标记基因和其他本领域已知的遗传元件。除非另有解释,本文使用的所有技术和科学术语具有与本公开所属领域技术人员通常理解相同的含义。除非上下文另有明确指示,单数形式“a”、“an”和“the”包括复数。“包含a或b”是指包括a或b,或a和b。还应理解,对核酸或多肽给出的所有碱基大小或氨基酸大小,以及所有分子量或分子质量值是近似的,并且出于描述性目的而提供。尽管在本公开的实践或测试中可以使用与本文所述的那些类似或等同的方法和材料,但下文描述了特别合适的方法和材料。在冲突的情况下,以本说明书(包括术语的解释)为准。此外,这些材料、方法和实施例仅是说明性的,而无意进行限定。iii.若干实施方式的概述本文公开了编码嵌合抗原受体(car)和截短的人表皮生长因子受体(huegfrt)的核酸分子。该编码的car包括与细胞外铰链区、跨膜区、细胞内共刺激结构域和细胞内信号传导结构域融合的肿瘤抗原特异性单克隆抗体。所述huegfrt包括两个egfr胞外结构域(结构域iii和结构域iv)和一个跨膜结构域,但是缺少两个膜远端胞外结构域(结构域i和结构域ii)和所有的胞内结构域(近膜结构域、酪氨酸激酶结构域和c-端尾)。还提供了共表达所公开的car和huegfrt的分离的细胞,例如t淋巴细胞。用该car构建体转导的t细胞可用于癌症免疫疗法。本文提供了编码car和huegfrt的核酸分子。在一些实施方式中,所述核酸分子以5'至3'的方向包括编码第一信号序列的核酸;编码抗原特异性抗体或其抗原结合片段的核酸;编码细胞外铰链区的核酸;编码跨膜结构域的核酸;编码细胞内共刺激结构域的核酸;编码细胞内信号传导结构域的核酸;编码自裂解2a肽的核酸;编码第二信号序列的核酸;和编码huegfrt的核酸。第一和第二信号序列可以是本领域已知的任何合适的信号序列。第一和第二信号序列可以是相同的信号序列或者它们可以是不同的信号序列。在一些实施方式中,第一和/或第二信号序列是粒细胞-巨噬细胞集落刺激因子受体信号序列(gmcsfrss)。在一些实施方式中,细胞外铰链区包括cd8α铰链区、cd28铰链区或者来自另一个免疫球蛋白分子例如igg1、igg4或igd的序列(例如,来自免疫球蛋白分子的ch2和/或ch3结构域)。该铰链区在本领域有时也称作“间隔子区”。在一些实施方式中,跨膜结构域包括cd8α、cd28、cd3ε、cd45、cd4、cd5、cd9、cd16、cd22、cd33、cd37、cd64、cd80、cd86、cd134、cd137或cd154跨膜结构域。跨膜结构域还可以是t细胞受体的α、β或ζ链的跨膜区。在一些实施方式中,细胞内共刺激结构域包括4-1bb(cd137,tnfrsf9)、cd28、icos、ox40(cd134)、cd27、cd30、cd40、pd-1、淋巴细胞功能相关抗原1(lfa-1)、cd2、cd7、light、nkg2c、b7-h3或dap10共刺激结构域。在一些实例中,细胞内共刺激结构域包括4-1bb和cd28。在一些实施方式中,细胞内信号传导结构域是具有免疫受体酪氨酸激活基序(itam)的结构域,例如cd3ζ或fcεriγ信号传导结构域。在具体的实施方式中,第一和第二信号序列包括gmcsfrss,细胞外铰链区包括cd8α铰链区,跨膜结构域包括cd8α跨膜结构域,细胞内共刺激结构域包括4-1bb共刺激结构域,并且细胞内信号传导结构域包括cd3ζ信号传导结构域。在一些实例中,编码cd8α铰链的核酸包括与seqidno:3至少90%、至少95%、至少96%、至少97%、至少98%或至少99%同一性的核苷酸序列。在一个非限制性实例中,编码cd8α铰链的核酸包括seqidno:3的序列。在一些实例中,编码cd8α跨膜结构域的核酸包括与seqidno:5至少90%、至少95%、至少96%、至少97%、至少98%或至少99%同一性的核苷酸序列。在一个非限制性实例中,编码cd8α跨膜结构域的核酸包括seqidno:5的序列。在一些实例中,编码4-1bb共刺激结构域的核酸分子包括与seqidno:7至少90%、至少95%、至少96%、至少97%、至少98%或至少99%同一性的核苷酸序列。在一个非限制性实例中,编码4-1bb共刺激结构域的核酸分子包括seqidno:7的序列。在一些实例中,编码cd3ζ信号传导结构域的核酸包括与seqidno:9至少90%、至少95%、至少96%、至少97%、至少98%或至少99%同一性的核苷酸序列。在一个非限制性实例中,编码cd3ζ信号传导结构域的核酸包括seqidno:9的序列。在一些实例中,编码第一gmcsfrss的核酸和编码第二gmcsfrss的核酸各自包括与seqidno:1至少90%、至少95%、至少96%、至少97%、至少98%或至少99%同一性的核苷酸序列。在一个非限制性实例中,编码第一gmcsfrss的核酸和编码第二gmcsfrss的核酸各自包括seqidno:1的序列。在一些实例中,自裂解2a肽是thoseaasigna病毒2a(thoseaasignavirus2a,t2a)肽。在其他实例中,自裂解2a肽是口蹄疫病毒(footandmouthdiseasevirus2a,f2a)肽、马甲型鼻炎病毒2a(equinerhinitisavirus2a,e2a)肽或猪捷申病毒-1(porcineteschovirus-12a,p2a)肽。在具体实例中,编码自裂解t2a肽的核酸包括与seqidno:11至少90%、至少95%、至少96%、至少97%、至少98%或至少99%同一性的核苷酸序列。在一个非限制性实例中,编码自裂解t2a肽的核酸包括seqidno:11的序列。在一些实例中,编码huegfrt的核酸包括与seqidno:13至少90%、至少95%、至少96%、至少97%、至少98%或至少99%同一性的核苷酸序列。在一个非限制性实例中,编码huegfrt的核酸包括seqidno:13的序列。在一些实施方式中,该核酸分子还在编码第一gmcsfrss的核酸5'包括人延伸因子1α(ef1α)启动子序列。但是,本领域技术人员可以选择任何合适的启动子序列。在一些实施方式中,该抗原结合片段是单链可变片段(scfv)或单域抗体。在一些实施方式中,该抗体或抗原结合片段特异性结合肿瘤抗原。在具体实例中,该肿瘤抗原是gpc3、gpc2或间皮素。在一些实例中,其中所述肿瘤抗原是gpc3,编码抗体结合片段的核酸包括seqidno:25(hyp7vh结构域核苷酸序列)的重链可变(vh)结构域互补决定区1(cdr1)、cdr2和cdr3核酸序列和seqidno:27(hyp7vl结构域核苷酸序列)的轻链可变(vl)结构域cdr1、cdr2和cdr3核酸序列。可以使用任何公知的编号方案例如imgt、kabat或chothia来确定cdr序列。在具体实例中,该vh结构域cdr1、cdr2和cdr3核酸序列各自包括seqidno:25的核苷酸91-105、148-204和301-318;和/或该vl结构域cdr1、cdr2和cdr3核酸序列各自包括seqidno:27的核苷酸70-120、166-186和283-309。在其他具体的实例中,该vh结构域cdr1、cdr2和cdr3核酸序列各自包括seqidno:25的核苷酸76-99、151-180和295-318;和/或该vl结构域cdr1、cdr2和cdr3核酸序列各自包括seqidno:27的核苷酸79-114、166-174和283-309。在一个非限制性实例中,编码抗体结合片段的核酸包括seqidno:15的核苷酸73-807的序列。在其他实例中,其中所述肿瘤抗原是gpc3,编码抗体结合片段的核酸包括seqidno:29(hn3单域抗体核苷酸序列)的cdr1、cdr2和cdr3核酸序列。可以使用任何公知的编号方案例如imgt、kabat或chothia来确定cdr序列。在具体实例中,该cdr1、cdr2和cdr3核酸序列各自包括seqidno:29的核苷酸91-105、148-195和286-315。在其他具体的实例中,该cdr1、cdr2和cdr3核酸序列各自包括seqidno:29的核苷酸76-99、151-171和286-315。在一个非限制性实例中,编码抗体结合片段的核酸包括seqidno:17的核苷酸73-420的序列。在其他实例中,其中所述肿瘤抗原是gpc3,编码抗体结合片段的核酸包括在wo2013/181543或wo2012/145469中公开的gpc3-特异性单克隆抗体的cdr核酸序列,这些专利通过引用整体并入本文。在一些实例中,其中所述肿瘤抗原是gpc2,编码抗体结合片段的核酸包括seqidno:31(lh7单域抗体核苷酸序列)的cdr1、cdr2和cdr3核酸序列。可以使用任何公知的编号方案例如imgt、kabat或chothia来确定cdr序列。在具体实例中,该cdr1、cdr2和cdr3核酸序列各自包括seqidno:31的核苷酸91-105、148-195和286-327。在其他具体的实例中,该cdr1、cdr2和cdr3核酸序列各自包括seqidno:31的核苷酸76-99、151-171和286-327。在一个非限制性实例中,编码抗体结合片段的核酸包括seqidno:19的核苷酸73-432的序列。在其他实例中,其中所述肿瘤抗原是gpc2,编码抗体结合片段的核酸包括seqidno:33(lh4单域抗体核苷酸序列)的cdr1、cdr2和cdr3核酸序列。可以使用任何公知的编号方案例如imgt、kabat或chothia来确定cdr序列。在具体实例中,该cdr1、cdr2和cdr3核酸序列各自包括seqidno:33的核苷酸91-105、148-195和286-327。在其他具体的实例中,该cdr1、cdr2和cdr3核酸序列各自包括seqidno:33的核苷酸76-99、151-171和286-327。在其他实例中,其中所述肿瘤抗原是gpc2,编码抗体结合片段的核酸包括seqidno:35(lh6单域抗体核苷酸序列)的cdr1、cdr2和cdr3核酸序列。可以使用任何公知的编号方案例如imgt、kabat或chothia来确定cdr序列。在具体实例中,该cdr1、cdr2和cdr3核酸序列各自包括seqidno:35的核苷酸91-105、148-198和289-330。在其他具体的实例中,该cdr1、cdr2和cdr3核酸序列各自包括seqidno:35的核苷酸76-99、151-174和289-330。在其他实例中,其中所述肿瘤抗原是gpc2,编码抗体结合片段的核酸包括在lietal.,procnatlacadsciusa114(32):e6623-e6631,2017中公开的gpc2-特异性单克隆抗体的cdr核酸序列。在一些实例中,其中所述肿瘤抗原是间皮素,编码抗体结合片段的核酸包括seqidno:37(yp218vh结构域核苷酸序列)的vh结构域cdr1、cdr2和cdr3核酸序列和seqidno:39(yp218vl结构域核苷酸序列)的vl结构域cdr1、cdr2和cdr3核酸序列。可以使用任何公知的编号方案例如imgt、kabat或chothia来确定cdr序列。在具体实例中,该vh结构域cdr1、cdr2和cdr3核酸序列各自包括seqidno:37的核苷酸91-108、101-204和298-36;和/或该vl结构域cdr1、cdr2和cdr3核酸序列各自包括seqidno:39的核苷酸70-102、148-168和265-303。在其他具体的实例中,该vh结构域cdr1、cdr2和cdr3核酸序列各自包括seqidno:37的核苷酸79-102、154-177和292-336;和/或该vl结构域cdr1、cdr2和cdr3核酸序列各自包括seqidno:39的核苷酸79-96、148-156和265-303。在其他实例中,其中所述肿瘤抗原是间皮素,编码抗体结合片段的核酸包括seqidno:41(sd1单域抗体核苷酸序列)的cdr1、cdr2和cdr3核酸序列。可以使用任何公知的编号方案例如imgt、kabat或chothia来确定cdr序列。在具体实例中,该cdr1、cdr2和cdr3核酸序列各自包括seqidno:41的核苷酸91-105、151-198和295-306。在其他具体的实例中,该cdr1、cdr2和cdr3核酸序列各自包括seqidno:41的核苷酸78-105、151-174和289-309。本文进一步提供了包括本文公开的car-编码核酸分子的载体。在一些实施方式中,该载体是病毒载体,例如,但不限于慢病毒载体。在其他实例中,其中所述肿瘤抗原是间皮素,编码抗体结合片段的核酸包括在wo2014/031476、wo2014/052064、美国专利第8,460,660号、美国专利第6,809,184号或美国专利第7,081,518号中公开的间皮素-特异性单克隆抗体的cdr核酸序列,这些专利各自通过引用整体并入本文。还提供了分离的宿主细胞,其包括本文公开的car-编码核酸分子。在一些实施方式中,所述分离的宿主细胞是t淋巴细胞。本公开还提供了分离的宿主细胞,其共表达嵌合抗原受体(car)和截短的人表皮生长因子受体(huegfrt)。在一些实施方式中,该car包括抗原特异性抗体或其抗原结合片段、细胞外铰链区、跨膜结构域、细胞内共刺激结构域和细胞内信号传导结构域;和/或该huegfrt包括来自人egfr的结构域iii、结构域iv和跨膜结构域,但是缺少表皮生长因子(egf)-结合结构域和细胞质结构域。在一些实施方式中,细胞外铰链区包括cd8α铰链区、cd28铰链区或来自另一个免疫球蛋白分子例如igg1、igg4或igd的序列(例如,来自免疫球蛋白分子的ch2和/或ch3结构域)。在一些实施方式中,跨膜结构域包括cd8α、cd28、cd3ε、cd45、cd4、cd5、cd9、cd16、cd22、cd33、cd37、cd64、cd80、cd86、cd134、cd137或cd154跨膜结构域。跨膜结构域还可以是t细胞受体的α、β或ζ链的跨膜区。在一些实施方式中,所述细胞内共刺激结构域包括4-1bb(cd137,tnfrsf9)、cd28、icos、ox40(cd134)、cd27、cd30、cd40、pd-1、淋巴细胞功能相关抗原1(lfa-1)、cd2、cd7、light、nkg2c、b7-h3或dap10共刺激结构域。在一些实施方式中,细胞内信号传导结构域是具有itam的结构域,例如cd3ζ或fcεriγ信号传导结构域。在具体的实施方式中,细胞外铰链区包括cd8α铰链区,跨膜结构域包括cd8α跨膜结构域,细胞内共刺激结构域包括4-1bb共刺激结构域,并且细胞内信号传导结构域包括cd3ζ信号传导结构域。在一些实例中,cd8α铰链区的氨基酸序列与seqidno:4为至少90%、至少95%、至少96%、至少97%、至少98%或至少99%的同一性。在具体实例中,cd8α铰链区的氨基酸序列包括seqidno:4。在一些实例中,cd8α跨膜结构域的氨基酸序列与seqidno:6为至少90%、至少95%、至少96%、至少97%、至少98%或至少99%的同一性。在具体实例中,cd8α跨膜结构域的氨基酸序列包括seqidno:6。在一些实例中,4-1bb共刺激结构域的氨基酸序列与seqidno:8为至少90%、至少95%、至少96%、至少97%、至少98%或至少99%的同一性。在具体实例中,4-1bb共刺激结构域的氨基酸序列包括seqidno:8。在一些实例中,cd3ζ信号传导结构域的氨基酸序列与seqidno:10为至少90%、至少95%、至少96%、至少97%、至少98%或至少99%的同一性。在具体实例中,cd3ζ信号传导结构域的氨基酸序列包括seqidno:10。在一些实例中,huegfrt的氨基酸序列与seqidno:14为至少90%、至少95%、至少96%、至少97%、至少98%或至少99%的同一性。在具体实例中,huegfrt的氨基酸序列包括seqidno:14。在一些实施方式中,该抗原结合片段是scfv或单域抗体。在一些实施方式中,该抗体或抗原结合片段特异性结合肿瘤抗原。在一些实例中,该肿瘤抗原是gpc3、gpc2或间皮素。在一些实例中,其中肿瘤抗原是gpc3,所述抗原结合片段的氨基酸序列包括seqidno:26(hyp7vh结构域)的vh结构域cdr1、cdr2和cdr3序列和seqidno:28(hyp7vl结构域)的vl结构域cdr1、cdr2和cdr3序列。可以使用任何公知的编号方案例如imgt、kabat或chothia来确定cdr序列。在具体实例中,该vh结构域cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:26的残基31-35、50-68和101-106和/或该vl结构域cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:28的残基24-40、56-62和95-103。在其他具体的实例中,该vh结构域cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:26的残基26-33、51-60和99-106和/或该vl结构域cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:28的残基27-38、56-58和95-103。在一个非限制性实例中,所述抗体结合片段的氨基酸序列包括seqidno:16的残基25-269。在其他实例中,其中所述肿瘤抗原是gpc3,所述抗原结合片段的氨基酸序列包括seqidno:30(hn3单域抗体序列)的cdr1、cdr2和cdr3序列。可以使用任何公知的编号方案例如imgt、kabat或chothia来确定cdr序列。在具体实例中,该cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:30的残基31-35、50-65和96-105。在其他具体的实例中,该cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:30的残基26-33、51-57和96-105。在一个非限制性实例中,所述抗体结合片段的氨基酸序列包括seqidno:18的残基25-140。在其他实例中,其中所述肿瘤抗原是gpc3,所述抗体结合片段的氨基酸序列包括在wo2013/181543或wo2012/145469中公开的gpc3-特异性单克隆抗体的cdr序列,这些专利通过引用整体并入本文。在一些实例中,其中所述肿瘤抗原是gpc2,所述抗原结合片段的氨基酸序列包括seqidno:32(lh7单域抗体序列)的cdr1、cdr2和cdr3序列。可以使用任何公知的编号方案例如imgt、kabat或chothia来确定cdr序列。在具体实例中,该cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:32的残基26-33、51-57和96-109。在其他具体的实例中,该cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:32的残基31-35,50-65和96-109。在一个非限制性实例中,所述抗体结合片段的氨基酸序列包括seqidno:20的残基25-144。在其他实例中,其中所述肿瘤抗原是gpc2,所述抗原结合片段的氨基酸序列包括seqidno:34(lh4单域抗体序列)的cdr1、cdr2和cdr3序列。可以使用任何公知的编号方案例如imgt、kabat或chothia来确定cdr序列。在具体实例中,该cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:34的残基31-35、50-65和96-109。在其他具体的实例中,该cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:34的残基26-33、51-57和96-109。在其他实例中,其中所述肿瘤抗原是gpc2,所述抗原结合片段的氨基酸序列包括seqidno:36(lh6单域抗体序列)的cdr1、cdr2和cdr3序列。可以使用任何公知的编号方案例如imgt、kabat或chothia来确定cdr序列。在具体实例中,该cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:36的残基31-35、50-66和97-110。在其他具体的实例中,该cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:36的残基26-33、51-58和97-110。在其他实例中,其中所述肿瘤抗原是gpc2,所述抗体结合片段的氨基酸序列包括在lietal.,procnatlacadsciusa114(32):e6623-e6631,2017中公开的gpc2-特异性单克隆抗体的cdr序列。在一些实例中,其中所述肿瘤抗原是间皮素,所述抗原结合片段的氨基酸序列包括seqidno:38(yp218vh结构域)的vh结构域cdr1、cdr2和cdr3序列和seqidno:40(yp218vl结构域)的vl结构域cdr1、cdr2和cdr3序列。可以使用任何公知的编号方案例如imgt、kabat或chothia来确定cdr序列。在具体实例中,vh结构域cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:38的残基31-36、51-68和100-112和/或vl结构域cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:40的残基24-34、50-56和89-101。在其他具体的实例中,vh结构域cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:38的残基27-34、52-59和98-112和/或vl结构域cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:40的残基27-32、50-52和89-101。在其他实例中,其中所述肿瘤抗原是间皮素,所述抗原结合片段的氨基酸序列包括seqidno:42(sd1单域抗体序列)的cdr1、cdr2和cdr3序列。可以使用任何公知的编号方案例如imgt、kabat或chothia来确定cdr序列。在具体实例中,该cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:42的残基31-35、51-66和99-102。在其他具体的实例中,该cdr1、cdr2和cdr3氨基酸序列各自包括seqidno:42的残基26-35、51-58和97-103。在其他实例中,其中所述肿瘤抗原是间皮素,所述抗体结合片段的氨基酸序列包括在wo2014/031476、wo2014/052064、美国专利第8,460,660号、美国专利第6,809,184号或美国专利第7,081,518号中公开的间皮素-特异性单克隆抗体的cdr序列,这些专利各自通过引用整体并入本文。在一些实施方式中,所述分离的宿主细胞t淋巴细胞。在一些实例中,t淋巴细胞是自体t淋巴细胞。在其他实例中,t淋巴细胞是同种异体t淋巴细胞。本文还提供包括本文公开的分离的(表达car的-)宿主细胞和药学上可接受的载体的组合物。本文进一步提供治疗受试者中gpc3-阳性癌症的方法。在一些实施方式中,该方法包括给受试者施用治疗有效量的包括编码本文公开的gpc3-靶向car的核酸分子的分离的宿主细胞,或施用治疗有效量的共表达gpc3-靶向car和huegfrt的分离的宿主细胞,如本文所公开的。在一些实例中,gpc3-阳性癌症是肝细胞癌、黑色素瘤、卵巢透明细胞癌、卵黄囊肿瘤、神经母细胞瘤、肝母细胞瘤或wilms肿瘤。还提供治疗受试者中gpc2-阳性癌症的方法。在一些实施方式中,该方法包括给受试者施用治疗有效量的包括编码本文公开的gpc2-靶向car的核酸分子的分离的宿主细胞,或使用治疗有效量共表达gpc2-靶向car和huegfrt的分离的宿主细胞,如本文所公开的。在一些实例中,gpc2-阳性癌症是神经母细胞瘤、急性淋巴细胞白血病、胚胎横纹肌肉瘤、腺泡状横纹肌肉瘤、尤文氏肉瘤、促结缔组织增生性小圆细胞肿瘤或骨肉瘤。进一步提供治疗受试者中间皮素-阳性癌症的方法。在一些实施方式中,该方法包括给受试者施用治疗有效量的包括编码本文公开的间皮素-靶向car的核酸分子的分离的宿主细胞,或施用治疗有效量的共表达间皮素-靶向car和huegfrt的分离的宿主细胞,如本文所公开的。在一些实例中,间皮素-阳性癌症是间皮瘤、前列腺癌、肺癌、胃癌、鳞状细胞癌、胰腺癌、胆管癌、三阴性乳腺癌或卵巢癌。在所述治疗方法的一些实施方式中,分离的宿主细胞是t淋巴细胞。在一些实例中,t淋巴细胞是自体t淋巴细胞。在其他实例中,t淋巴细胞是同种异体t淋巴细胞。在本文的一个实施方式中,提供了编码car的核酸分子,其以5’至3’的方向包括编码第一gmcsfrss的核酸;编码抗原特异性抗体或其抗原结合片段的核酸;编码cd8α铰链区的核酸;编码cd8α跨膜结构域的核酸;编码4-1bb共刺激结构域的核酸;编码cd3ζ信号传导结构域的核酸;编码自裂解2a肽的核酸;编码第二gmcsfrss的核酸;和编码huegfrt的核酸。在一些实例中,编码抗体结合片段的核酸包括seqidno:15的核苷酸73-807、seqidno:17的核苷酸73-420或seqidno:19的核苷酸73-432的序列。在具体实例中,该核酸分子与seqidno:15、seqidno:17或seqidno:19有至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%或至少99%的同一性。在具体的非限制性实例中,该核酸分子包括seqidno:15、seqidno:17或seqidno:19的核苷酸序列。在本文的一个实施方式中,提供了共表达car和huegfrt的分离的宿主细胞,其中所述car包括抗原特异性抗体或其抗原结合片段、cd8α铰链区、cd8α跨膜结构域、4-1bb共刺激结构域和cd3ζ信号传导结构域;和所述huegfrt包括来自人egfr的结构域iii、结构域iv和跨膜结构域,但是缺少egf-结合结构域和细胞质结构域。在一些实例中,所述抗原结合片段的氨基酸序列包括seqidno:16的残基25-269、seqidno:18的残基25-140或seqidno:20的残基25-144。在具体实例中,car的氨基酸序列与seqidno:16的残基25-491、seqidno:18的残基25-362或seqidno:20的残基25-366有至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%或至少99%的同一性,并且huegfrt的氨基酸序列与seqidno:14有至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%或至少99%的同一性。在其他具体的实例中,car的氨基酸序列相对于seqidno:16的残基25-491、seqidno:18的残基25-362或seqidno:20的残基25-366包括不超过10、不超过9、不超过8、不超过7、不超过6、不超过5、不超过4、不超过3、不超过2或不超过1个氨基酸替代,并且huegfrt的氨基酸序列相对于seqidno:14包括不超过10、不超过9、不超过8、不超过7、不超过6、不超过5、不超过4、不超过3、不超过2或不超过1个氨基酸替代。在具体的非限制性实例中,car的氨基酸序列包括seqidno:16的残基25-491、seqidno:18的残基25-362或seqidno:20的残基25-366,且huegfrt的氨基酸序列包括seqidno:14。iv.对肿瘤抗原特异的抗体本文公开的car可以通过选择适当的肿瘤抗原特异性单克隆抗体或其抗原结合片段靶向表达或过表达特定抗原的肿瘤细胞。在一些实施方式中,car的抗原结合部分是单克隆抗体的抗原结合片段。在具体实例中,该抗原结合片段是scfv或单域(vh结构域)抗体。尽管本文公开的car可以与任何抗原-特异性抗体(或其抗原结合片段)一起使用,示例性抗体包括gpc3-特异性、gpc2-特异性和间皮素-特异性单克隆抗体。a.gpc3-特异性抗体可以改造本文公开的car构建体以包括任何gpc3-特异性单克隆抗体或其抗原结合片段。几种gpc3-特异性单克隆抗体是本领域已知的,包括,但不限于在pct公开号wo2013/181543中公开的yp6、yp7、yp8、yp9和yp9.1,和在wo2012/145469中公开的hn3,这些专利通过引用整体并入本文。在本文的一些实施方式中,所述car包括抗原结合片段,所述抗原结合片段包括gpc3-特异性单克隆抗体yp7(公开于wo2013/181543)或其人源化形式的cdr序列。下面提供了yp7和人源化的yp7(hyp7)的核苷酸和氨基酸序列。表1a-1d显示yp7和hyp7的cdr1、cdr2和cdr3。在其他实施方式中,所述car包括单域单克隆抗体,所述单域单克隆抗体包括gpc3-特异性抗体hn3(公开于wo2012/145469)的cdr序列。下面提供了hn3的核苷酸和氨基酸序列。表2a-2b显示在hn3中cdr1、cdr2和cdr3的位置。yp7vh核苷酸序列(seqidno:21)gaggtgcagcttgttgagactggtggaggaatggtgcagcctgaagggtcattgaaactctcatgtgcagcctctggattcaccttcaataagaatgccatgaattgggtccgccaggctccaggaaagggtttggaatgggttgctcgcataagaaataaaactaataattatgcaacatattatgccgattcagtgaaagccaggtttaccatctccagagatgattcacaaagcatgctctatctgcaaatgaacaacttgaaaattgaggacacagccatgtactattgtgtggctggtaactcgtttgcttactggggccaagggactctggtcactgtctctgcayp7vh氨基酸序列(seqidno:22)evqlvetgggmvqpegslklscaasgftfnknamnwvrqapgkglewvarirnktnnyatyyadsvkarftisrddsqsmlylqmnnlkiedtamyycvagnsfaywgqgtlvtvsayp7vl核苷酸序列(seqidno:23)gacattgtgatgtcacagtctccatcctccctagttgtgtcaattggagagaaggttactatgacctgcaagtccagtcagagccttttatatagcagcaatcaaaagaactacttggcctggtaccaacagaaaccagggcagtctcctaaactgctgatttactgggcatccagtagggaatctggggtccctgatcgcttcacaggcagtggatctgggacagatttcactctcaccatcagcagtgtgaaggctgaagacctggcagtttattactgtcagcaatattataactatccgctcacgttcggtgctgggaccaagttggagctgaaayp7vl氨基酸序列(seqidno:24)divmsqspsslvvsigekvtmtckssqsllyssnqknylawyqqkpgqspklliywassresgvpdrftgsgsgtdftltissvkaedlavyycqqyynypltfgagtklelkhyp7vh核苷酸序列(seqidno:25)gaggtgcagcttgttgagtctggtggaggattggtgcagcctggagggtcattgagactctcatgtgcagcctctggattcaccttcaataagaatgccatgaattgggtccgccaggctccaggaaagggtttggaatgggttggccgcataagaaataaaactaataattatgcaacatattatgccgattcagtgaaagccaggtttaccatctccagagatgattcaaagaactcactctatctgcaaatgaacagcttgaaaaccgaggacacagccgtgtactattgtgtggctggtaactcgtttgcttactggggccaagggactctggtcactgtctctgcahyp7vh氨基酸序列(seqidno:26)evqlvesggglvqpggslrlscaasgftfnknamnwvrqapgkglewvgrirnktnnyatyyadsvkarftisrddsknslylqmnslktedtavyycvagnsfaywgqgtlvtvsahyp7vl核苷酸序列(seqidno:27)gacattgtgatgacccagtctccagactccctagctgtgtcactgggagagagggccactatcaactgcaagtccagtcagagccttttatatagcagcaatcaaaagaactacttggcctggtaccaacagaaaccagggcagcctcctaaactgctgatttactgggcatccagtagggaatctggggtccctgatcgcttcagtggcagtggatctgggacagatttcactctcaccatcagcagtctgcaggctgaagacgtggcagtttattactgtcagcaatattataactatccgctcacgttcggtcaggggaccaagttggagatcaaahyp7vl氨基酸序列(seqidno:28)divmtqspdslavslgeratinckssqsllyssnqknylawyqqkpgqppklliywassresgvpdrfsgsgsgtdftltisslqaedvavyycqqyynypltfgqgtkleik表1a.在yp7/hyp7vh序列中cdr的位置(根据kabat)表1b.在yp7/hyp7vh序列中cdr的位置(根据imgt)表1c.在yp7/hyp7vl序列中cdr的位置(根据kabat)表1d.在yp7/hyp7vl序列中cdr的位置(根据imgt)hn3dna序列(seqidno:29)caggtgcagctggtgcagtctgggggaggcttggtacagcctggagggtccctgagactctcctgtgcagcctcttatttcgatttcgattcttatgaaatgagctgggtccgccaggctccagggaagggcctagagtggattgggagtatctatcatagtgggagcacctactacaacccgtccctcaagagtcgagtcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacaccctgagagccgaggacacagccacgtattactgtgcgagagtaaatatggaccgatttgactactggggccagggaaccctggtcaccgtctcctcaagthn3蛋白质序列(seqidno:30)qvqlvqsggglvqpggslrlscaasyfdfdsyemswvrqapgkglewigsiyhsgstyynpslksrvtisrdnskntlylqmntlraedtatyycarvnmdrfdywgqgtlvtvsss表2a.在hn3序列中cdr的位置(根据kabat)cdrdna序列(seqidno:29)蛋白质序列(seqidno:30)cdr1核苷酸91-105氨基酸31-35cdr2核苷酸148-195氨基酸50-65cdr3核苷酸286-315氨基酸96-105表2b.在hn3序列中cdr的位置(根据imgt)cdrdna序列(seqidno:29)蛋白质序列(seqidno:30)cdr1核苷酸76-99氨基酸26-33cdr2核苷酸151-171氨基酸51-57cdr3核苷酸286-315氨基酸96-105b.gpc2-特异性抗体还可以改造本文公开的car构建体以包括任何gpc2-特异性单克隆抗体或其抗原结合片段。在本文的一些实施方式中,所述car包括抗原结合片段,所述抗原结合片段包括gpc2-特异性单域单克隆抗体lh7、lh4、lh6、lh1、lh2或lh3的cdr序列(公开于lietal.,procnatlacadsciusa114(32):e6623-e6631,2017)。下面提供了lh7、lh4和lh6的核苷酸和氨基酸序列。表3a-5b显示h7、lh4和lh6的cdr1、cdr2和cdr3的位置。lh7dna(seqidno:31)caggtgcagctggtgcagtctgggggaggcttggtacagcctggagggtccctgagactctcctgtgcagcctctgatttctatttctatgattatgaaatgagctgggtccgccaggctccagggaagggtctggagtggattgggactgtctcctatagtgggagcacctactacaacccgtccctcaagagtcgagtcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacaccctaagagccgaggacacagccatgtattactgtgcgagaggttacagctatgatgactcccgatattttgactactggggccagggaaccctggtcaccgtctcctcalh7蛋白(seqidno:32)qvqlvqsggglvqpggslrlscaasdfyfydyemswvrqapgkglewigtvsysgstyynpslksrvtisrdnskntlylqmntlraedtamyycargysyddsryfdywgqgtlvtvss表3a.在lh7序列中cdr的位置(根据kabat)cdrdna序列(seqidno:31)蛋白质序列(seqidno:32)cdr191-10531-35cdr2148-19550-65cdr3286-32796-109表3b.在lh7序列中cdr的位置(根据imgt)cdrdna序列(seqidno:31)蛋白质序列(seqidno:32)cdr176-9926-33cdr2151-17151-57cdr3286-32796-109lh4dna(seqidno:33)caggtgcagctggtgcagtctgggggaggcttggtacagcctggagggtccctgagactctcctgtgcagcctcttctttctatttcgatgattatgaaatgagctgggtccgccaggctccagggaaggccctggagtggattgggcgtatctataccagtgggagcaccaactacaacccctccctcaagagtcgagtcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacaccctgagagccgaggacacagccacgtattactgtgcgaggggatattgtagtggtggtagctgctactttgactactggggccagggaaccctggtcaccgtctcctcalh4蛋白(seqidno:34)qvqlvqsggglvqpggslrlscaassfyfddyemswvrqapgkalewigriytsgstnynpslksrvtisrdnskntlylqmntlraedtatyycargycsggscyfdywgqgtlvtvss表4a.在lh4序列中cdr的位置(根据kabat)cdrdna序列(seqidno:33)蛋白质序列(seqidno:34)cdr191-10531-35cdr2148-19550-65cdr3286-32796-109表4b.在lh4序列中cdr的位置(根据imgt)cdrdna序列(seqidno:33)蛋白质序列(seqidno:34)cdr176-9926-33cdr2151-17151-57cdr3286-32796-109lh6dna(seqidno:35)caggtgcagctggtgcagtctgggggaggcttggtacagcctggagggtccctgagactctcctgtgcagcctctgatttctatttcgatgattatgaaatgagctgggtccgccaggctccagggaaggggctggagtgggtctcaactattagtggtagtggtggtggcacatactacgcagactcagtgaagggccgattcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacaccctgagagccgaggacacagccacatattactgtgcgagaggttacagttatgacgactcccgatattttgactactggggccagggaaccctggtcaccgtctcctcalh6蛋白(seqidno:36)qvqlvqsggglvqpggslrlscaasdfyfddyemswvrqapgkglewvstisgsgggtyyadsvkgrftisrdnskntlylqmntlraedtatyycargysyddsryfdywgqgtlvtvss表5a.在lh6序列中cdr的位置(根据kabat)cdrdna序列(seqidno:35)蛋白质序列(seqidno:36)cdr191-10531-35cdr2148-19850-66cdr3289-33097-110表5b.在lh6序列中cdr的位置(根据imgt)cdrdna序列(seqidno:35)蛋白质序列(seqidno:36)cdr176-9926-33cdr2151-17451-58cdr3289-33097-110c.间皮素-特异性抗体还可以改造本文公开的car构建体以包括任何间皮素-特异性单克隆抗体或其抗原结合片段。几种间皮素-特异性单克隆抗体是本领域已知的,包括,但不限于在pct公开号wo2014/031476中公开的yp218、yp223、yp3、yp158和yp187,在pct公开号wo2014/052064中公开的sd1,在美国专利第8,460,660号中公开的hn1,在美国专利第6,809,184号中公开的ss,和在美国专利第7,081,518号中公开的ss1,这些专利各自通过引用整体并入本文。下面提供了yp218和sd1的核苷酸和氨基酸序列。表6a-7b显示yp218和sd1的cdr的位置。yp218vh核苷酸序列(seqidno:37)cagcagcagctggaggagtccgggggaggcctggtcaagcctgagggatccctgacactcacctgcaaagcctctggattcgacctcggtttctacttttacgcctgttgggtccgccaggctccagggaagggcctggagtggatcgcatgcatttatactgctggtagtggtagcacgtactacgcgagctgggcgaaaggccgattcaccatctccaaagcctcgtcgaccacggtgactctgcaaatgaccagtctggcagccgcggacacggccacctatttctgtgcgagatctactgctaatactagaagtacttattatcttaacttgtggggcccaggcaccctggtcaccgtctcctcayp218vh氨基酸序列(seqidno:38)qqqleesggglvkpegsltltckasgfdlgfyfyacwvrqapgkglewiaciytagsgstyyaswakgrftiskassttvtlqmtslaaadtatyfcarstantrstyylnlwgpgtlvtvssyp218vl核苷酸序列(seqidno:39)gacgtcgtgatgacccagactccagcctccgtgtctgaacctgtgggaggcacagtcaccatcaagtgccaggccagtcagaggattagtagttacttatcctggtatcagcagaaaccagggcagcgtcccaagctcctgatctttggtgcatccactctggcatctggggtcccctcgcggttcaaaggcagtggatctgggacagaatacactctcaccatcagcgacctggagtgtgccgatgctgccacttactactgtcagagttatgcttattttgatagtaataattggcatgctttcggcggagggaccgaggtggtggtcyp218vl氨基酸序列(seqidno:40)dvvmtqtpasvsepvggtvtikcqasqrissylswyqqkpgqrpkllifgastlasgvpsrfkgsgsgteytltisdlecadaatyycqsyayfdsnnwhafgggtevvv表6a.在yp218vh序列中cdr的位置(根据kabat)cdrdna序列(seqidno:37)蛋白质序列(seqidno:38)cdr1核苷酸91-108氨基酸31-36cdr2核苷酸101-204氨基酸51-68cdr3核苷酸298-336氨基酸100-112表6b.在yp218vh序列中cdr的位置(根据imgt)cdrdna序列(seqidno:37)蛋白质序列(seqidno:38)cdr1核苷酸79-102氨基酸27-34cdr2核苷酸154-177氨基酸52-59cdr3核苷酸292-336氨基酸98-112表6c.在yp218vl序列中cdr的位置(根据kabat)cdrdna序列(seqidno:39)蛋白质序列(seqidno:40)cdr1核苷酸70-102氨基酸24-34cdr2核苷酸148-168氨基酸50-56cdr3核苷酸265-303氨基酸89-101表6d.在yp218vl序列中cdr的位置(根据imgt)cdrdna序列(seqidno:39)蛋白质序列(seqidno:40)cdr1核苷酸79-96氨基酸27-32cdr2核苷酸148-156氨基酸50-52cdr3核苷酸265-303氨基酸89-101sd1核苷酸序列(seqidno:41):caggtgcagctggtgcagtctgggggaggcttggtacagcctggagggtccctgagactctcctgtgcagcctctgatttcgatttcgctgcttatgaaatgagctgggtccgccaggctccaggacaaggccttgagtgggtggcaattatatcacatgatggaatcgataaatactacacagactccgtgaagggccgattcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacaccctgagagccgaggacacagccacgtattactgtttaaggcttggtgctgtaggccagggaaccctggtcaccgtctcctcaagtsd1氨基酸序列(seqidno:42):qvqlvqsggglvqpggslrlscaasdfdfaayemswvrqapgqglewvaiishdgidkyytdsvkgrftisrdnskntlylqmntlraedtatyyclrlgavgqgtlvtvsss表7a.在sd1序列中cdr的位置(根据kabat)cdrdna序列(seqidno:41)蛋白质序列(seqidno:42)cdr191-10531-35cdr2151-19851-66cdr3295-30699-102表7b.在sd1序列中cdr的位置(根据imgt)cdrdna序列(seqidno:41)蛋白质序列(seqidno:42)cdr178-10526-35cdr2151-17451-58cdr3289-30997-103v.嵌合抗原受体(car)本文公开的是car(也称为嵌合t细胞受体、人工t细胞受体或嵌合免疫受体)和经改造以表达car的t细胞。通常,car包括结合部分、细胞外铰链/间隔子元件、跨膜区和执行信号功能的细胞内结构域(cartellierietal.,jbiomedbiotechnol2010:956304,2010;daietal.,jnatlcancerinst108(7):djv439,2016)。在许多情况下,结合部分是单克隆抗体的抗原结合片段,例如scfv或单域抗体。间隔子/铰链区通常包括来自igg亚类的序列,例如igg1、igg4、igd和cd8结构域。跨膜结构域可以衍生自各种不同的t细胞蛋白质,例如cd3ζ、cd4、cd8或cd28。尽管在car中可以采用整个胞内t细胞信号传导结构域,在许多情况下没有必要使用整条链。就使用细胞内t细胞信号传导结构域的截短部分而言,只要其转导相关的t细胞效应功能信号,就可以使用该截短部分代替完整链。用于car的细胞内t细胞信号传导结构域的例子包括t细胞受体(tcr)的细胞质序列和在抗原受体结合后共同启动信号转导的共刺激分子,以及这些序列的衍生物或变体和具有相同功能能力的任何合成序列。已经使用几种不同的胞内结构域来产生car。例如,胞内结构域可以由具有itam的信号传导链例如cd3ζ或fcεriγ组成。在一些情况下,胞内结构域还包括至少一个另外的共刺激结构域的胞内部分。共刺激结构域是指包含共刺激分子的胞内结构域的car的一部分。共刺激分子是除抗原受体或其配体以外的淋巴细胞有效应答抗原所需的细胞表面分子。共刺激分子包括,例如cd28、4-1bb(cd137,tnfrsf9)、ox-40(cd134)、icos、cd27和/或dap10。car还可以包括信号肽序列,例如,抗原结合结构域的n-端。信号肽序列可以是任何适合的信号肽序列,例如来自粒细胞-巨噬细胞集落刺激因子受体(gmcsfr)、免疫球蛋白轻链κ或il-2的信号序列。尽管信号肽序列可以促进car在细胞表面上的表达,但是在表达的car中信号肽序列的存在对于为了使car起作用不是必需的。当car在细胞表面上表达时,信号肽序列可以从car上切割下来。因此,在一些实施方式中,car缺少信号肽序列。本文公开的car表达自构建体(例如,自慢病毒载体),所述构建体还表达截短形式的人egfr(huegfrt;如在下面vi节中详细讨论的)。car和huegfrt被自裂解肽序列(例如t2a)分开,使得在转导细胞中表达后,car从huegfrt上切割下来(参见图1)。在本文公开的一些实施方式中,car构建体以n-端至c-端的方向编码以下氨基酸序列:gmcsfrss:mlllvtslllcelphpafllip(seqidno:2)ndei:hm抗原结合:scfv或单域抗体序列spei:tscd8α铰链:tttpaprpptpaptiasqplslrpeacrpaaggavhtrgldfacd(seqidno:4)cd8αtm:iyiwaplagtcgvlllslvit(seqidno:6)4-1bb:krgrkkllyifkqpfmrpvqttqeedgcscrfpeeeeggcel(seqidno:8)cd3ζ:rvkfsrsadapayqqgqnqlynelnlgrreeydvldkrrgrdpemggkprrknpqeglynelqkdkmaeayseigmkgerrrgkghdglyqglstatkdtydalhmqalppr(seqidno:10)t2a:egrgslltcgdveenpgp(seqidno:12)gmcsfrss:mlllvtslllcelphpafllip(seqidno:2)huegfrt:rkvcngigigefkdslsinatnikhfknctsisgdlhilpvafrgdsfthtppldpqeldilktvkeitgflliqawpenrtdlhafenleiirgrtkqhgqfslavvslnitslglrslkeisdgdviisgnknlcyantinwkklfgtsgqktkiisnrgensckatgqvchalcspegcwgpeprdcvscrnvsrgrecvdkcnllegeprefvenseciqchpeclpqamnitctgrgpdnciqcahyidgphcvktcpagvmgenntlvwkyadaghvchlchpnctygctgpglegcptngpkipsiatgmvgalllllvvalgiglfm(seqidno:14)本文公开的表达car的t细胞可用于靶向特定的细胞类型,如肿瘤细胞,例如gpc3-阳性、gpc2-阳性或间皮素-阳性肿瘤细胞。与标准的基于ctl的免疫疗法相比,表达car的t细胞的使用更为普遍,因为表达car的ctl是hla非限制性的,因此可用于任何患有表达靶抗原的肿瘤的患者。因此,本文提供了car,其包括肿瘤-特异性抗体(或其结合片段),例如gpc3-特异性抗体、gpc2-特异性抗体或间皮素-特异性抗体。还提供了编码car的分离的核酸分子和载体,和表达car的宿主细胞,例如t淋巴细胞。表达包括gpc3-特异性、gpc2-特异性或间皮素-特异性单克隆抗体的car的t细胞可用于治疗分别表达gpc3、gpc2和间皮素的癌症。vi.截短的人egfr(huegfrt)人表皮生长因子受体由四个细胞外结构域、一个跨膜结构域和三个细胞内结构域组成。发现egfr结构域按以下n-端至c-端的顺序:结构域i–结构域ii–结构域iii–结构域iv–跨膜(tm)结构域–近膜结构域–酪氨酸激酶结构域–c-端尾。结构域i和结构域iii为参与配体结合的亮氨酸-富含结构域。结构域ii和结构域iv为半胱氨酸-富含结构域并且不与egfr配体接触。结构域ii介导与来自其他egfr家族成员的类似结构域形成同型或异型二聚体,并且结构域iv可以与结构域ii形成二硫键。egfrtm结构域单次通过细胞膜,且可能在蛋白质二聚化中起作用。胞内结构域包括介导egfr信号传导的近膜结构域、酪氨酸激酶结构域和c-端尾(weeandwang,cancers9(52),doi:10.3390/cancers9050052;ferguson,annurevbiophys37:353-373,2008;wangetal.,blood118(5):1255-1263,2011)。截短形式的人egfr,在本文也称为“huegfrt”仅包括结构域iii、结构域iv和tm结构域。因此,huegfrt缺少结构域i、结构域ii和所有三个胞内结构域。huegfrt不能结合egf并且缺乏信号传导活性。但是,该分子保留了结合特定egfr-特异性单克隆抗体的能力,例如fda批准的西妥昔单抗(pct公开号wo2011/056894,其通过引用并入本文)。用本文公开的编码huegfrt和肿瘤抗原特异性car的构建体(例如慢病毒载体)转导t细胞允许使用标记的egfr单克隆抗体西妥昔单抗(erbituxtm)选择转导的t细胞。例如,西妥昔单抗可以用生物素标记,并且转导的t细胞可以使用可商购的抗生物素磁珠(例如,来自miltenyibiotec)进行选择。huegfrt的共表达还允许体内跟踪过继转移的car-表达t细胞。此外,西妥昔单抗与表达huegfrt的t细胞的结合诱导adcc效应细胞的细胞毒性,由此提供在体内消除转导t细胞的机制(wangetal.,blood118(5):1255-1263,2011),例如在治疗结束时。在本文的一些实施方式中,编码huegfrt的核酸分子与seqidno:13有至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%或至少99%的同一性。在一些实例中,编码huegfrt的核酸分子包括seqidno:13的核苷酸序列或由其组成。在一些实施方式中,huegfrt的氨基酸序列与seqidno:14有至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%或至少99%的同一性。在一些实例中,huegfrt的氨基酸序列包括seqidno:14或由其组成。在其他实施方式中,huegfrt的氨基酸序列相对于seqidno:14包括不超过10、不超过9、不超过8、不超过7、不超过6、不超过5、不超过4、不超过3、不超过2或不超过1个氨基酸替代。在一些实例中,氨基酸取代是保守替代。vii.car-表达细胞组合物提供了组合物,其包括与一种或多种药学上或生理上可接受的载体、稀释剂或赋形剂组合的car-表达细胞。car-表达细胞可以是t细胞,例如cd3+t细胞,例如cd4+和/或cd8+t细胞,和/或nk细胞。这样的组合物可以包括缓冲剂,例如中性缓冲盐溶液、磷酸缓冲盐溶液等;碳水化合物,例如葡萄糖、甘露糖、蔗糖、葡聚糖或甘露醇;蛋白质;多肽或氨基酸,例如甘氨酸;抗氧化剂;螯合剂,如edta或谷胱甘肽;佐剂(例如氢氧化铝);和防腐剂。细胞对接受者而言可以是自体的。但是,细胞也可以是异源的(同种异体的)。关于细胞,可以使用各种水性载体,例如缓冲盐溶液等来引入细胞。这些溶液是无菌的,并且通常没有不想要的物质。这些组合物可以通过常规的、众所周知的灭菌技术进行灭菌。所述组合物可包含近似生理条件所需的药学上可接受的辅助物质,例如ph调节剂和缓冲剂、毒性调节剂等,例如乙酸钠、氯化钠、氯化钾、氯化钙、乳酸钠等。这些制剂中的浓度可以在很大范围内变化,并且将主要根据所选择的具体施用方式和受试者的需要,根据流体体积、粘度、体重等进行选择。可以由医师考虑年龄、体重、肿瘤大小、转移程度和患者(受试者)状况的个体差异来确定待施用组合物的精确量。一般而言,本文所述的包含car-表达t细胞(和/或nk细胞)的药物组合物可以以下述剂量施用:104至109细胞/kg体重,例如105至106细胞/kg体重,包括这些范围内的所有整数值。示例性剂量为106细胞/kg至约108细胞/kg,例如约5x106细胞/kg至约7.5x107细胞/kg,例如约2.5x107细胞/kg或约5.0x107细胞/kg。可以以这些剂量一次或多次施用组合物,例如2、3、4、5、6、7、8、9或10次。可以通过使用免疫疗法中众所周知的输注技术来施用组合物(参见,例如rosenbergetal.,neweng.j.ofmed.319:1676,1988)。组合物可以每天、每周、每半个月(bimonthly)或每月施用。在一些非限制性实例中,将组合物配制成用于静脉内施用并且多次施用。施用的数量和频率由受试者的病情、受试者疾病的类型和严重性等因素确定,尽管适当的剂量可以通过临床试验确定。在一些实施方式中,将car-编码核酸分子引入到细胞例如t细胞或nk细胞中,并且受试者接受初始细胞给药,以及随后的一次或多次细胞给药,其中随后的一次或多次给药在之前给药后小于15天,例如14、13、12、11、10、9、8、7、6、5、4、3或2天给予。在一个实施方式中,每周向受试者(例如,人)给予多于一次的car-表达细胞给药,例如,每周给予2、3或4次本公开的car-表达细胞给药。在一个实施方式中,受试者每周接受多于一次的car-表达t细胞给药(例如,每周2、3或4次给药)(也称为周期),随后一周不进行car表达细胞给药,然后向受试者给予一次或更多次额外的car-表达细胞给药(例如,每周多于一次的cart细胞给药)。在另一个实施方式中,受试者(例如人受试者)接受一个以上的car-表达细胞周期,并且每个周期之间的时间少于10、9、8、7、6、5、4或3天。在一个实施方式中,每隔一天施用一次car-表达细胞,每周3次。在另一个实施方式中,施用car-表达细胞至少两周、三周、四周、五周、六周、七周、八周或更长时间。施用给患者的上述治疗的剂量将随所治疗病症的确切性质和治疗接受者而变化。可以根据本领域公认的实践来缩放用于人类施用的剂量。在一些实施方式中,car-表达t细胞能够在体内复制,导致长期持续存在,从而可以持续控制肿瘤。在各个方面,给受试者施用t细胞后,施用给受试者的t细胞或这些细胞的后代在受试者中持续至少4个月、5个月、6个月、7个月、8个月、9个月、10个月、11个月、12个月、13个月、14个月、15个月、16个月、17个月、18个月、19个月、20个月、21个月、22个月、23个月或数年。在其他实施方式中,给受试者施用car-表达t细胞后,该细胞及其后代的存在持续少于6个月、5个月、4个月、3个月、2个月或1个月,例如3周、2周、1周。主题组合物的施用可以任何方便的方式进行,包括通过注射、摄入、输液、植入或移植。公开的组合物可以通过静脉注射经动脉、皮下、皮内、肿瘤内、结节内、髓内、肌内或腹膜内施用给患者。在一些实施方式中,组合物通过皮内或皮下注射施用给患者。在其他实施方式中,通过静脉注射施用本发明的组合物。也可以将组合物直接注射到肿瘤或淋巴结中。在一些实施方式中,受试者可以经历白细胞分离(leukapheresis),其中白细胞在体外被收集、富集或耗尽,以选择和/或分离感兴趣的细胞,例如t细胞和/或nk细胞。这些细胞分离物可以通过本领域已知的方法扩增并进行处理,使得可以引入一个或更多个car构建体,从而产生表达car的自体细胞。在本文的一些实施方式中,使用表达car和截短形式的人egfr(huegfrt)的慢病毒载体来产生car-表达细胞。huegfrt的共表达允许使用识别huegfrt的抗体(例如西妥昔单抗,参见pct公开号wo2011/056894,其通过引用并入本文)来选择和纯化car-表达t细胞,如在上面第vi节中所描述的。在一些实施方式中,通过裂解红细胞并耗尽单核细胞,例如通过percolltm梯度离心或对流离心淘析(counterflowcentrifugalelutriation),从外周血淋巴细胞中分离t细胞。可以通过阳性或阴性选择技术进一步分离t细胞的特定亚群,例如cd3+、cd28+、cd4+、cd8+、cd45ra+和cd45ro+t细胞。例如,通过与抗-cd3/抗-cd28(例如3×28)-缀合珠子例如m-450cd3/cd28t孵育足以进行阳性选择所需t细胞的时间,可以分离t细胞,参见美国公开申请第us20140271635a1号。在非限制性实例中,所述时间为约30分钟。在其他非限制性实例中,所述时间的范围为30分钟至36小时或更长时间,以及介于两者之间整数值。在另外的非限制性实例中,所述时间是至少1、2、3、4、5、6个小时,10至24个小时,24个小时或更长时间。在与其他细胞类型相比t细胞很少的任何情况下(例如从免疫受损个体中分离),可以使用更长的孵育时间来分离t细胞。此外,使用更长的孵育时间可以提高捕获cd8+t细胞的效率。因此,通过简单地缩短或延长允许t细胞结合cd3/cd28珠子的时间和/或通过增加或降低珠子与t细胞的比例(如本文进一步描述的),可以在培养起始时或该过程中的其他时间点优先选择或不选择t细胞的亚群。另外,通过增加或减少珠子或其他表面上的抗-cd3和/或抗-cd28抗体的比例,可以在培养开始时或在其他所需时间点优先选择或不选择t细胞的亚群。也可以使用多轮选择。通过阴性选择富集t细胞群可以采用针对阴性选择细胞特有的表面标志物的抗体的组合来完成。一种方法是通过负磁性免疫粘附或流式细胞术进行细胞分选和/或选择,其使用针对阴性选择的细胞上存在的细胞表面标志物的单克隆抗体混合物。例如,为了通过阴性选择来富集cd4+细胞,单克隆抗体混合物通常包括针对cd14、cd20、cd11b、cd16、hla-dr和cd8的抗体。可以选择表达一种或更多种细胞因子的t细胞群。针对细胞表达进行筛选的方法公开于pct公开号wo2013/126712。为了通过阳性或阴性选择分离期望的细胞群,可以改变细胞浓度和表面(例如,颗粒如珠子)以确保细胞和珠子的最大接触。在一些实施方式中,使用1×109细胞/ml的浓度。在其他实施方式中,使用大于1×108细胞/ml。在其他实施方式中,使用10、15、20、25、30、35、40、45、50、65、70、75、80、85、90、95或100×106细胞/ml的细胞浓度。不受理论的束缚,使用高浓度可以导致细胞产量增加、细胞活化和细胞扩增。也可以使用较低浓度的细胞。不受理论的束缚,显著稀释t细胞和表面(例如,颗粒如珠子)的混合物,使颗粒与细胞之间的相互作用最小化。这样可以选择表达大量所需抗原的细胞与颗粒结合。例如,cd4+t细胞表达更高水平的cd28并且在稀释浓度下比cd8+t细胞更有效地被捕获。在一些实施方式中,使用的细胞浓度是5×106/ml。在其他实施方式中,使用的浓度可以为约1×105/ml至1×106/ml,以及介于两者之间的任何整数值。viii.治疗方法本文提供了通过给受试者施用治疗有效量的本文公开的肿瘤-靶向cart细胞来治疗受试者中癌症的方法。本文还提供通过给受试者施用治疗有效量的本文公开的肿瘤-靶向cart细胞来抑制受试者中肿瘤生长或转移的方法。具体提供了治疗受试者中gpc3-阳性癌症的方法。在一些实施方式中,该方法包括给受试者施用治疗有效量的包括编码gpc3-靶向car和huegfrt的核酸分子的分离的宿主细胞,或施用治疗有效量的共表达gpc3-靶向car和huegfrt的分离的宿主细胞。在一些实施方式中,gpc3-阳性癌症是hcc、黑色素瘤、卵巢透明细胞癌、yst、神经母细胞瘤、肝母细胞瘤或wilms肿瘤。还提供治疗受试者中gpc2-阳性癌症的方法。在一些实施方式中,该方法包括给受试者施用治疗有效量的包括编码gpc2-靶向car和huegfrt的核酸分子的分离的宿主细胞,或施用治疗有效量的共表达gpc2-靶向car和huegfrt的分离的宿主细胞。在一些实施方式中,gpc2-阳性癌症为神经母细胞瘤、急性淋巴细胞白血病、胚胎横纹肌肉瘤、腺泡状横纹肌肉瘤、尤文氏肉瘤、促结缔组织增生性小圆细胞肿瘤或骨肉瘤。进一步提供了治疗受试者中间皮素-阳性癌症的方法。在一些实施方式中,该方法包括给受试者施用治疗有效量的包括编码间皮素-靶向car和huegfrt的核酸分子的分离的宿主细胞,或施用治疗有效量的共表达间皮素-靶向car和huegfrt的分离的宿主细胞。在一些实施方式中,间皮素-阳性癌症是间皮瘤、前列腺癌、肺癌、胃癌、鳞状细胞癌、胰腺癌、胆管癌、三阴性乳腺癌或卵巢癌。在本文公开方法的一些实施方式中,分离的宿主细胞是t淋巴细胞。在一些实例中,t淋巴细胞是自体t淋巴细胞。car-表达t细胞的治疗有效量将取决于疾病的严重程度,疾病的类型以及患者的总体健康状况。car-表达t细胞及其组合物的治疗有效量是如临床医生或其他合格观察者所指出的那样,其提供症状的主观缓解或客观可确定的改善(例如肿瘤体积或转移的减少)。本文公开的car-表达t细胞和组合物的施用也可以伴有其他抗癌药或治疗性治疗(例如肿瘤的手术切除)的施用。任何合适的抗癌药可以与本文公开的组合物组合施用。示例性的抗癌药包括但不限于,化疗剂,例如有丝分裂抑制剂、烷基化剂、抗代谢物、嵌入抗生素(intercalatingantibiotics)、生长因子抑制剂、细胞周期抑制剂、酶、拓扑异构酶抑制剂、抗存活剂、生物反应调节剂、抗激素(例如抗雄激素)和抗血管生成剂。其他抗癌治疗包括放射治疗和其他特异性靶向癌细胞的抗体。烷基化剂的非限制性实例包括氮芥(如二氯甲基二乙胺(mechlorethamine)、环磷酰胺、美法仑、尿嘧啶氮芥或苯丁酸氮芥)、烷基磺酸盐(如白消安)、亚硝基脲(如卡莫司汀、洛莫司汀(lomustine)、司莫司汀(semustine)、链脲霉素或达卡巴嗪)。抗代谢物的非限制性实例包括叶酸类似物(例如甲氨蝶呤)、嘧啶类似物(例如5-fu或阿糖胞苷)和嘌呤类似物,例如巯基嘌呤或硫代鸟嘌呤。天然产物的非限制性实例包括长春碱类(例如长春碱、长春新碱或长春地辛)、表鬼臼毒素(epipodophyllotoxins)(例如依托泊苷或替尼泊苷)、抗生素(例如放线菌素d、柔红霉素、阿霉素、博来霉素、普卡霉素或丝裂霉素c)和酶(l-天冬酰胺酶)。杂类试剂的非限制性实例包括铂配位络合物(例如顺式二胺-二氯铂ii,也称为顺铂)、取代的脲(例如羟基脲)、甲基肼衍生物(例如丙卡巴肼(procarbazine))和肾上腺皮质抑制剂(例如米托坦(mitotane)和氨鲁米特(aminoglutethimide))。激素和拮抗剂的非限制性实例包括肾上腺皮质类固醇(如泼尼松)、孕激素(如己酸羟孕酮、醋酸甲羟孕酮和醋酸甲地孕酮(magestrolacetate))、雌激素(如己烯雌酚和炔雌醇)、抗雌激素(如他莫昔芬(tamoxifen))和雄激素(如丙酸睾丸酮(testeroneproprionate)和氟甲睾酮)。最常用的化疗药物的例子包括阿霉素、美法仑(alkeran)、阿糖胞苷(ara-c)、bicnu、白消安、ccnu,卡铂(carboplatinum)、顺铂、环磷酰胺(cytoxan)、道诺霉素、dtic、5-fu、氟达拉滨、羟基脲(hydrea)、伊达比星(idarubicin)、异环磷酰胺、氨甲喋呤、光辉霉素(mithramycin)、丝裂霉素、米托蒽醌、氮芥、紫杉醇(或其他紫杉烷、如多西紫杉醇)、长春花碱(velban)、长春新碱、vp-16、而一些更新的药物包括吉西他滨(gemzar)、赫赛汀(herceptin)、伊立替康(irinotecan)(camptosar,cpt-11)、克拉屈滨(leustatin)、诺维苯(navelbine)、利妥昔单抗sti-571、泰索帝、拓扑替康(hycamtin)、希罗达(卡培他滨)、泽文林(zevelin)和骨化三醇。可以使用的免疫调节剂的非限制性实例包括as-101(wyeth-ayerstlabs.)、溴匹立明(bropirimine)(upjohn)、γ-干扰素(genentech)、gm-csf(粒细胞巨噬细胞集落刺激因子;geneticsinstitute)、il-2(cetusorhoffman-laroche)、人免疫球蛋白(cutterbiological)、imreg(来自imregofneworleans,la.)、sk&f106528和tnf(肿瘤坏死因子;genentech)。某些类型的癌症的另一种常见治疗是外科手术治疗,例如外科手术切除癌症或其一部分。治疗的另一实例是放射疗法,例如在手术切除之前向肿瘤部位施用放射性物质或能量(例如外射束疗法(externalbeamtherapy))以帮助根除肿瘤或使其缩小。提供以下实施例以说明某些具体特征和/或实施方式。这些实施例不应被解释为将本公开限制为所描述具体特征或实施方式。实施例实施例1:car-表达慢病毒构建体本实施例描述了三种编码肿瘤-靶向嵌合抗原受体(car)和截短的人egfr(huegfrt)的慢病毒载体的产生。使用pwpt骨架慢病毒载体(addgene)来产生三种靶向gpc3或gpc2的car构建体。所述载体还编码能被fda-批准的抗-egfr抗体西妥昔单抗识别的huegfrt,使得能够进行cart细胞标记和清除。图1提供了用于产生肿瘤-靶向car的慢病毒构建体的示意图。该慢病毒构建体包括car编码区和编码huegfrt的区域,各自之前都有一个粒细胞-巨噬细胞集落刺激因子受体信号序列(gmcsfrss)。所述两个区域被自裂解t2a序列分开,使得在该构建体表达后,car从huegfrt上切割下来。该构建体的表达由人延伸因子1α(ef1α)启动子驱动。所述car包括抗原结合区、cd8α铰链区、cd8α跨膜(tm)结构域、4-1bb共刺激区和cd3ζ信号传导结构域。该huegfrt包括两个胞外结构域(结构域iii和结构域iv)和一个tm结构域。所述car构建体中有两种靶向gpc3。慢病毒载体pmh228编码gpc3-特异性单域单克隆抗体hn3(公开于wo2012/145469,其通过引用并入本文)。pmh288的载体图谱显示于图2a,car.hn3的核苷酸和氨基酸序列在本文中如seqidno:17和18所示。第二gpc3-特异性car表达自慢病毒载体pmh289,其编码小鼠gpc3-特异性抗体yp7的人源化scfv。小鼠抗体yp7公开于wo2013/18154,其通过引用并入本文。pmh289的载体图谱显示于图2b;car.hyp7的核苷酸和氨基酸序列在本文中如seqidno:15和16所示。第三car构建体靶向gpc2并由慢病毒载体pmh290编码,所述慢病毒载体pmh290编码gpc2-特异性单域单克隆抗体lh7(公开于lietal.,procnatlacadsciusa114(32):e6623-e6631,2017)。pmh290的载体图谱显示于图2c;car.lh7的核苷酸和氨基酸序列在本文中如seqidno:19和20。在上面的第iv节中提供了hn3、人源化yp7(hyp7)和lh7的核苷酸和氨基酸序列,它们如seqidno:25-32所示。实施例2:材料和方法该实施例描述了慢病毒生产和滴定方法,t细胞活化和转导方法以及与实施例3中所述研究相关的功能测定。慢病毒生成、浓缩和滴定在转染前18-24小时将293t细胞接种到10cm的培养皿(7.0×106细胞/培养皿)中,使得在转染时单层细胞密度达到最佳90%融合度。转染前大约30-60分钟,移出细胞上清液,并用5ml加有血清和抗生素的完全培养基(dulbecco改良的eagle培养基;dmem)代替。对于每个培养皿,将总共16μg的dna(8μg慢病毒载体质粒、2μg包膜质粒md2g和6μg包装质粒pax28)稀释到500μl无血清dmem培养基中并轻轻涡旋混合。另外,将48μlcalfectintm(signagenlaboratories;dna:calfectintm=1:3)添加到500μl无血清dmem中并轻轻混合。然后将稀释的calfectintm试剂立即与稀释的dna溶液混合并涡旋,以形成calfectintm-dna复合物。将混合物在室温孵育10分钟。接下来,将calfectintm-dna复合物逐滴添加到每个培养皿的培养基中,并通过轻轻旋转板使其均匀。转染后48-72小时从细胞上清液收集病毒。将上清液以500xg离心5分钟,然后通过0.45μm过滤器过滤。为了浓缩慢病毒,将澄清的上清液转移到无菌容器中,并将3体积的澄清上清液与1体积的lenti-xconcentrator(clontech)合并。将混合物在4℃下孵育30分钟至过夜。然后将样品在4℃下以1,500xg离心45分钟,形成灰白色沉淀物。除去上清液,并使用完整的dmem将沉淀物重悬于原始体积的1/10至1/100。慢病毒滴定将293t细胞以1至5x105细胞/孔的密度接种在12孔板中,每孔含1ml生长培养基(补充有10%fbs的dmem)。一个孔用于计数细胞,和另一个孔用作非转导对照(ni)。其他孔一式两份地用500μl、100μl、50μl、20μl或10μl的粗(未浓缩的)上清液转导。用生长培养基使每个孔的体积增加到500μl。对于浓缩的病毒样品,在500μl新鲜生长培养基中用1μl、0.1μl、0.01μl、0.001μl或0.0001μl的载体转导细胞。三天后,将细胞用1mlpbs洗涤,并每孔加入200μl胰蛋白酶/edta使其脱落(detached),并在37℃下孵育1分钟。将生长培养基(800μl)添加到每个孔中以重悬细胞。此步骤使胰蛋白酶和edta失活。然后将细胞转移到5mlfacs管中,在500xg和4℃下离心5分钟,并除去上清液。于冰上在facs缓冲液(pbs中5%bsa,0.01%叠氮化钠)中用1μg/ml西妥昔单抗对细胞进行染色1小时。用pbs1x洗涤细胞,在500xg和4℃下离心5分钟,并将沉淀物重悬于具有适当荧光染料的第二抗体中。所述第二抗体也在facs缓冲液中稀释。在冰上对细胞进行染色1小时,然后在pbs中洗涤一次,然后在500xg和4℃下离心5分钟。将沉淀物重悬于500μl含1%甲醛的pbs中,并在室温下孵育5分钟。该步骤固定细胞并使载体颗粒失活。在pbs1x中洗涤固定的细胞,并在500xg和4℃下离心5分钟。将沉淀物重悬于1mlpbs中。使用流式细胞仪分析细胞的car表达。t细胞活化和转导方案将dynabeadstm人t-激活剂cd3/cd28(dynabeadstmhumant-activatorcd3/cd28,lifetechnologies)重悬于小瓶中,并将所需体积转移至试管。加入1mlpbs或生长培养基,并通过涡旋混合5秒。离心小瓶并弃去上清液。经洗涤的dynabeadstm重悬于培养基(rpmi1640+10%fbs),所述培养基的体积与从小瓶中取出的珠子的初始体积相同。将冷冻的外周血单个核细胞(pbmc)解冻并重悬于生长培养基(rpmi1640+10%fbs)中。使用台盼蓝试剂对细胞计数。将细胞(1×106)接种在24孔板的1ml培养基中。以珠子和细胞的比例为2:1加入dynabeadstm人t-激活剂cd3/cd28,并加入50u/mlil-2。孵育24小时后,在10μg/ml硫酸鱼精蛋白的存在下,将pbmc用慢病毒(moi为5)在1000g旋转接种60分钟。然后将细胞重悬于病毒上清液中并孵育过夜,维持50u/mlil-2。第二天,将细胞离心并重悬于新鲜的rpmi1640+10%fbs培养基中,加入100iu/mlil-2。在接下来的10天里,每天检查培养物,注意细胞大小和形状。通常在耗竭的细胞培养物中观察到细胞萎缩和增殖率降低。彻底重悬后,每隔一天对细胞计数一次。当细胞密度超过2.0×106细胞/ml或当培养基变成黄色时,在含有100u/mlil-2的培养基中使培养物分开至密度为0.5-1×106细胞/ml。当细胞生长动力学和体积表明细胞已由激活静止下来时,它们可用于功能测定或冷冻保存。功能测定将表达荧光素酶的靶细胞(2×103)接种在96孔板各孔的50μl培养基中。以不同的效应子(e)/靶(t)比例制备cart细胞(效应细胞)。将50μl的cart细胞添加至每个孔,并在37℃下孵育过夜。每个效应子:靶(e:t)比例一式三份进行。第二天,收集上清液以通过elisa测量细胞因子水平,并在-20℃下保存。将稳定的glo荧光素酶试剂(promega)添加到每个孔中,以裂解肿瘤细胞,并在室温下避光孵育平板至少5分钟。在victor(perkinelmer)上读取发光。结果分析为基于单独肿瘤细胞的孔中荧光素酶活性的杀死百分数:[%杀死=100-((来自具有效应子和靶细胞的孔的rlu)/(来自具有靶细胞的孔的rlu)×100)]。实施例3:gpc3-靶向car诱导gpc3-表达细胞系的细胞毒性并降低动物模型中gpc3-阳性肿瘤的肿瘤体积该实施例描述了表达car.hn3和car.hyp7的t细胞的体外和体内细胞毒性。慢病毒载体pmh288(表达car.hn3)和pmh289(表达car.hyp7)的t细胞转导效率通过流式细胞术使用抗huegfrt抗体西妥昔单抗来评估。编码car.hn3(图3a)和car.hyp7(图3b)的慢病毒载体分别转导65%和45.4%的t细胞。人血清igg用作对照(图3c)。测试了car.hyp7t细胞对几种人细胞系的细胞毒性,包括gpc3+g1细胞、hep3b细胞、hepg2细胞和huh7细胞以及gpc3-a431细胞、t3m4细胞和imr32细胞。对于每个细胞系,使用效应子:靶标比例为1:2、1.5:1、5:1和16:1。car.hyp7t细胞对所有gpc3阳性细胞系具有细胞毒性(图4a-4d),但对gpc3阴性细胞系没有细胞毒性(图4e-4g)。进行了另一项研究以确定用car.hyp7t细胞的处理是否诱导培养物中gpc3阳性细胞的ifn-γ产生。hep3b、huh7和g1细胞经空白处理(mock-treated)或用car.hyp7t细胞处理。如图5所示,car.hyp7t细胞诱导所有三个靶gpc阳性肿瘤细胞的ifn-γ分泌。进行了一项研究,以测试gpc3-靶向cart细胞抑制小鼠gpc3阳性肿瘤生长的能力。小鼠在第0天腹膜内注射了400万个hep3b细胞。在第10天,对小鼠进行空白注射或注射pbs、1000万个car.hn3t细胞(hn3-10m)、1000万个car.hyp7t细胞(hyp7-10m),2000万个car.hyp7t细胞(hyp7-20m)或4000万个car.hyp7t细胞(hyp7-40m)。通过生物发光成像测量肿瘤大小。与pbs治疗和空白治疗的动物相比,用car.hyp7t细胞的治疗(在所有测试剂量下)导致肿瘤体积明显减少(图6)。还评估了car.hyp7t细胞对小鼠hep3b异种移植肿瘤的抗肿瘤作用的持久性。小鼠在第0天腹膜内注射了400万个hep3b细胞。在第10天,对小鼠进行空白注射或注射pbs,1000万个car.hn3t细胞(hn3-10m)、1000万个car.hyp7t细胞(hyp7-10m)、2000万个car.hyp7t细胞(hyp7-20m)或4000万个car.hyp7t细胞(hyp7-40m)。首先,在治疗后测量肿瘤体积长达3周。在pbs治疗、空白治疗和car.hn3治疗的小鼠中肿瘤随时间稳定增加。相反,在1000万和2000万个car.hyp7t细胞的剂量下,肿瘤体积在三周内几乎保持不变,而在4000万个car.hyp7t细胞的剂量下,肿瘤体积明显降低(图7a)。第二项研究评估了在7周的过程中荷hep3b肿瘤的小鼠的抗肿瘤活性。该研究评估了用pbs、1000万个car.hn3t细胞、1000万个car.hyp7t细胞或4000万个car.hyp7t细胞治疗的小鼠。结果表明,在7周的研究中,4000万个car.hyp7t细胞的剂量导致肿瘤体积减小并维持减小的肿瘤体积(图7b)。在hep3b细胞接种后还评估了荷hep3b肿瘤的小鼠的存活率长达70天。这项研究追踪了注射了pbs、1000万个car.hyp7t细胞或4000万个car.hyp7t细胞的小鼠。用1000万和4000万个car.hyp7t细胞的治疗分别导致50%和100%的存活率。pbs治疗的小鼠均未存活(图7c)。接下来,在另一个gpc3阳性肿瘤模型中测试car.hyp7t细胞。将hepg2异种移植nsg小鼠进行空白治疗或用1000万个car.hyp7t细胞或4000万个car.hyp7t细胞进行治疗。如图8a-8d所示,在20天的研究期内,使用任一剂量的car.hyp7t细胞进行治疗均可导致肿瘤体积减小。实施例4:gpc3-靶向car研究的材料和方法该实施例提供了实施例5中描述的研究的实验过程。细胞培养人hcc细胞系hep3b获自美国马里兰州贝塞斯达市国家癌症研究所(nci)(nationalcancerinstitute(nci),bethesda,maryland)。hepg2(肝母细胞瘤)、a431(表皮癌)和hek-293t细胞系购自美国菌种保藏中心(americantypeculturecollection,atcc)。g1是稳定表达人gpc3的转染a431细胞系。用获自ncifrederick的表达的萤火虫荧光素酶的慢病毒转导hep3b和hepg2(dayetal.,pigmentcellmelanomares22:283-295,2009)。表达荧光素酶的huh-7细胞系(一种hcc细胞系)获自贝勒医学院(baylorcollegeofmedicine)。将上述细胞系在补充有10%fbs、1%l-谷氨酰胺和1%青霉素-链霉素的dmem中于5%co2的潮湿气氛中在37℃下培养。t3m4(人胰腺癌细胞系)细胞获自nci并经改造以表达荧光素酶。根据制造商的说明,通过ficoll(gehealthcare)从健康供体的血液中分离出外周血单个核细胞(pbmc)。hcc患者的pbmc获自nci。jurkat细胞也购自atcc。这些细胞在补充有10%fbs、1%l-谷氨酰胺和1%青霉素-链霉素的rpmi-1640培养基中,于含5%co2的潮湿环境中在37℃下生长。所有细胞系均通过形态学和生长速率进行验证,并且不含支原体。gpc3-靶向car的产生将来自hyp7的gpc3-特异性scfv和单域抗体片段hn3分别在框内(inframe)亚克隆至基于ef-1α启动子的慢病毒表达载体pwpt(addgene)中。该构建体包含编码cd8α铰链和跨膜区、4-1bb共刺激结构域、细胞内cd3ζ、自裂解t2a序列和huegfrt的表达盒,如图10b所示(也参见实施例1)。终产物通过序列分析进行确认。慢病毒产生、t-细胞转导和扩增使用calfectin(signagen),通过用获自addgene的包装质粒pspax2和包膜质粒pmd2.g共转染至hek-293t细胞中来产生重组gpc3-car慢病毒载体。转染72小时后,从上清液中收集慢病毒颗粒,并按照制造商的说明,用lenti-x浓缩剂(clontech)浓缩100倍。pbmc购自奥克拉荷马血液研究所(oklahomabloodinstitute),并且根据制造商的说明,在il-2的存在下以2:1的珠子/细胞比例使用抗-cd3/抗-cd28抗体-包被的珠子(invitrogen)刺激pbmc24小时。如前所述产生cart细胞(lietal.,procnatlacadsciusa114:e6623-e6631,2017)。为了随时间追踪t细胞数量,使用台盼蓝对存活细胞进行计数。流式细胞术gpc3car对t细胞的转导效率通过抗-egfr人单克隆抗体西妥昔单抗(erbitux)和山羊-抗-人igg-藻红蛋白(pe)或别藻蓝蛋白(apc)-缀合的抗体(jacksonimmunoresearch)进行检测。t细胞上的car表达使用gpc3-hfc融合蛋白和山羊-抗-人igg-pe-缀合抗体进行测量。pe-缀合的抗-cd3、抗-cd4和抗-cd8抗体获自ebioscience。使用facscantoii(bdbiosciences)进行数据采集并使用flowjo软件(treestar)进行分析。细胞毒性测定用gpc3-特异性car转导的t细胞的细胞毒性通过之前所述的基于荧光素酶的测定来确定(lietal.,procnatlacadsciusa114:e6623-e6631,2017)。简而言之,cart细胞和表达荧光素酶的gpc3-阳性(g1,hep3b,hepg2,huh-7)和gpc3-阴性(a431,t3m4)肿瘤细胞在不同的效应子/靶(e:t)比例下孵育24小时。荧光素酶活性使用荧光素酶测定系统(promega)在victor(perkinelmer)上进行测量。car-表达t细胞的细胞毒性也通过使用incucyte-flr-platform(essenbioscience)进行测试。简而言之,以2:1的e:t比将t细胞添加到表达gfp的hepg2肿瘤细胞中。每2分钟拍摄一次图像,长达140小时。基于gfp表达对活细胞的数量进行定量。使用incucytezoom肝细胞成像系统分析细胞的杀伤活性。细胞因子测定t细胞和肿瘤细胞共培养24小时后收集的上清液中的细胞因子水平使用人细胞因子22-plexpanel(粒酶b、gm-csf、ifn-γ、tnf-α、il-1β、il-2、il-4、il-5、il-6、il-7、il-10、il-12、il-13、il-15、il-21、ccl-3、ccl-4、ccl-19、ccl-20、cx3cl1、cxcl-11、cxcl-8)在luminex系统(thermofisherscientific)上进行分析。通过单细胞细胞因子分析评估t细胞多功能性将冷冻保存的cart细胞产物解冻,并在含有il-2(10ng/ml)的完全rpmi1640培养基中培养。过夜恢复后,使用ficoll富集活cart细胞。使用抗cd4或抗cd8微珠(miltenyibiotec)分离cd4+/cd8+t细胞亚群,并以1:1的比例用hep3b或g1细胞刺激20小时。接下来,使用之前描述的方法确定单细胞功能谱(functionalprofiles)(maetal.,cancerdiscov3:418-429,2013;rossietal.,blood132:804-814,2018;xueetal.,jimmunothercancer5:85,2017)。将功能谱分类为效应子组(粒酶b、ifn-γ、ccl-3、穿孔素(perforin)、tnf-α、tnf-β)、刺激性组(gm-csf、il-2、il-5、il-7、il-8、il-9、il-12、il-15、il-21)、调节性组(il-4、il-10、il-13、il-22、tgf-β1、scd137、scd40l)、趋化(chemoattractive)组(ccl-11、ip-10、ccl-4、rantes)和炎性组(il-1β、il-6、il-17a、il-17f、mcp-1、mcp-4)。多功能car产物t细胞定义为,结合产生的每种蛋白质的量,从每个细胞预先指定的panel共分泌至少2种蛋白质的细胞。此外,使用预先指定的公式计算每个样品的psi(maetal.,cancerdiscov3:418-429,2013),定义为多功能细胞的百分比乘以这些细胞分泌的蛋白质的平均荧光强度(mfi)。免疫组织化学人hcc组织和正常组织微阵列购自usbiomax,并用抗-gpc3抗体yp7进行免疫染色。将所有的组织样品送至histoservinc.(germantown,md)进行染色。人正常组织cdna阵列人类正常组织阵列购自origene。该panel包含48个样品,覆盖了不同位置的所有主要人类正常组织。gpc3引物和rt2sybrgreenqpcrmastermix购自qiagen。实时定量在appliedbiosystems7900ht实时pcr系统上进行。使用2-δδct方法分析结果。蛋白质印迹用冰冷裂解缓冲液(cellsignalingtechnology)裂解细胞,并通过在4℃下以10,000g离心10分钟使细胞澄清。按照制造商的说明,使用双辛可宁酸测定(pierce)测量蛋白质浓度。将20μg细胞裂解物上样至4-20%sds-page凝胶上进行电泳。抗gpc3抗体yp7描述于pct公开号wo2013/181543和phungetal.(mabs4(5):592-599,2012)中。抗活性-β-连环蛋白抗体获自millipore。所有其他抗体均获自cellsignalingtechnology。crispr/cas9-介导的gpc3编辑下表列出了靶向gpc3的不同外显子的sgrna。lenticrisprv2表达载体是addgene的产品(质粒#52961)。简而言之,用bsmbi消化载体,并使用凝胶提取试剂盒(qiagen)进行凝胶纯化。遵循先前描述的方案(sanjanaetal.,natmethods11:783-784,2014;shalemetal.,science343:84-87,2014),将每个靶向位点的一对寡核苷酸退火并连接到线性化的lenticrisprv2载体中,以生成表达grna的质粒。靶向gpc2的sgrna用作对照。根据制造商的说明,使用lipofectamine2000(thermofisherscientific)用表达grna的质粒转染hep3b。然后在转染后将细胞在37℃下孵育72小时。使用结晶紫测定法(crystalvioletassay)测定不同的grna表达质粒对细胞增殖的影响。sgrnaseqidno:序列gpc3外显子sgrna1-143gcagtctctggaagaaggag1sgrna1-244tggtgacaggtggcgtccgg1sgrna245cggttttccaaggtgagttc2sgrna3-146ggtcacgtcttgctcctcgg3sgrna3-247gacatcaatgagtgcctccg3sgrna448gataataagcagatctatat4sgrna5-149cgttttccgccacagggcta5sgrna5-250agggtgtcgttttccgccac5对照sgrna51gaggcagagcaggtagtcaggpc2afp测定根据制造商的说明,使用酶联免疫吸附测定(genwaybiotech)确定血清afp水平。微滴式数字pcr使用flexigenedna试剂盒(qiagen)从t细胞分离基因组dna。根据制造商的说明,在qx200微滴式数字pcr系统(bio-rad)上进行了微滴式数字pcr。动物研究饲养五周大的雌性nod/scid(nsg)小鼠(ncifrederick),并按照批准的规程进行处理。对于已建立的腹膜内(i.p.)模型hep3b和hepg2,将300万个表达荧光素酶的hep3b或200万个表达荧光素酶的hepg2肿瘤细胞腹膜内(i.p.)注射给小鼠。为了耗尽宿主淋巴细胞区室(lymphocytecompartment),在cart细胞输注前24小时给所有小鼠腹膜内注射200mg/kg环磷酰胺。对于hep3b模型,将小鼠随机分为六组,并分别仅通过腹膜内注射不同的cart细胞一次,如下所示:(a)仅生理盐水而无t细胞(pbs);(b)500万个未转导的t细胞(空白);(c)500万个car(hn3)t细胞;(d)500万个car(hyp7)t细胞;(e)1000万个car(hyp7)t细胞;或(f)2000万个car(hyp7)t细胞。对于hepg2模型,将小鼠随机分为两组,包括空白和car(hyp7)。对于已建立的原位hep3b模型,在小鼠肝脏中用50万个表达荧光素酶的hep3b细胞进行接种。肿瘤形成3周后,向小鼠腹膜内或静脉内输注car(hyp7)t细胞。为了检测带有hcc异种移植物的小鼠的肿瘤生长和存活率,所有小鼠均腹膜内注射3mgd-荧光素(perkinelmer),10分钟后每周使用xenogenivislumina(perkinelmer)进行成像。使用livingimage软件分析每只小鼠的生物发光信号通量,以光子/秒/平方厘米/球面度(photons/s/cm2/sr)表示。当生物发光信号达到5×1010时,对小鼠实施安乐死。为了测量gpc3敲除对hcc肿瘤细胞生长的影响,将200万个hep3b细胞皮下注射到裸鼠中。在肿瘤形成并达到100mm3的大小后,每隔一天肿瘤内注射表达sgrna5-2的质粒或空载体开始治疗,总共5次注射。用卡尺每周测定肿瘤尺寸两次。通过以下公式计算以立方毫米为单位的肿瘤体积:(a)×(b2)×0.5,其中“a”是肿瘤长度,“b”是肿瘤宽度,以毫米为单位。毒理学分析从每组中选择三只nsg小鼠进行毒理学研究。对样品进行全血细胞计数(cbc),全面的血清化学分析(vetscan,abaxisveterinarydiagnostics,unioncity,ca)和内部器官重量。这些分析由位于马里兰州nci-frederick的病理学/历史技术实验室(pathology/histotechnologylaboratoryinnci-frederick,md)进行。统计学所有实验至少重复三遍,以确定结果的可重复性。使用prism(graphpadsoftware)软件分析数据,并以平均值±sem表示。使用未配对的学生t检验(2-尾)分析结果。p<0.05被认为具有统计学意义。所有统计分析均使用prism软件进行。实施例5:glypican3-靶向嵌合抗原受体t细胞用于治疗肝细胞癌该实施例描述了这样的发现:在肝细胞癌的小鼠模型中,表达huegfrt和抗-gpc3抗体hyp7的结合片段的t细胞表现出明显的抗癌活性以及强大的t细胞激活和扩增能力。在hcc和正常组织中的gpc3表达为了分析肿瘤和正常组织中的gpc3表达,使用yp7抗体通过免疫组织化学检查了hcc患者的46对肿瘤组织和邻近的非肿瘤组织(肝硬化或肝炎)。与匹配的肿瘤邻近组织中仅2%(1/46)相比,gpc3蛋白在50%(23/46)的原发性hcc中高度表达。在另外40例hcc组织中,有24例2%(1/46)发现了强gpc3染色,但在任何正常肝组织中均未发现。cart细胞疗法的一个问题是由于抗原在正常组织上的表达而可能导致中靶的肿瘤外毒性(on-target,off-tumortoxicities)。在这里,分析了30种人类正常组织中的gpc3表达。值得注意的是,gpc3蛋白在包括大脑、心脏、肺、胃、小肠、结肠、肾脏、胰腺、脾脏、神经和皮肤在内的所有基本正常组织中均不存在。在所有正常组织中,仅在睾丸中检测到低水平的gpc3蛋白表达。还通过定量实时pcr在人正常组织阵列中测量了gpc3mrna水平。与蛋白质谱一致,除胎盘外,在大多数正常组织中未发现gpc3mrna表达(图9),这与之前在人胎盘中表达gpc3的报道一致(khanetal.,histolhistopathol16:71-78,2001)。然后通过流式细胞术在一组癌细胞系上检查gpc3的细胞表面表达。yp7显示与hcc细胞系(hep3b、hepg2和huh-7)以及过表达gpc3的a431细胞系(g1)的强结合。相比之下,yp7没有显示与a431和t3m4细胞的结合,这表明yp7抗体检测到的gpc3表达具有高度的肿瘤特异性。gpc3-特异性cart细胞的产生人单域抗体hn3识别gpc3的n-小叶(图10a)(fengetal.,procnatlacadsciusa110:e1083-1091,2013)。靶向gpc3的c-小叶的yp7抗体(hyp7)也被人源化以降低免疫原性的风险(zhangandho,scireports6:33878,2016)。将hn3或hyp7抗体的可变区同框克隆到包含具有4-1bb和cd3ζ胞内域的表达盒的慢病毒载体中(图10b)(另请参见实施例1)。为了促进细胞追踪和消融(ablation),将截短的人表皮生长因子受体(huegfrt)掺入构建体中并通过t2a核糖体跳跃序列与car分开。huegfrt缺少配体结合和酪氨酸激酶活性所必需的结构域,但保留了抗egfr单克隆抗体西妥昔单抗的结合表位(wangetal.,blood118:1255-1263,2011)。如图2b所示,将car质粒转导到来自健康供体或hcc患者的原代t细胞中,在体外扩增10-12天,然后在hcc细胞和动物模型中进行测试。通过流式细胞术确定gpc3-靶向car的表达。如重组gpc3-人fc(hfc)融合蛋白所检测的,两种car在人jurkatt细胞的表面均被有效表达。另外,通过细胞表面huegfrt表达证明了car在人原代t细胞上的表达。如图10c所示,car(hn3)和car(hyp7)的转导效率分别为76%和58%。在体外扩增11天后,超过99%的健康供体来源的car(hyp7)t细胞变为cd3阳性,其包括相似频率的cd4+(43%)和cd8+(56%)t细胞亚群(图10d)。值得注意的是,在hcc患者来源的car(hyp7)t细胞中,cd4+t细胞的比例(14.9%)明显低于cd8+t细胞(57.2%)。令人惊讶的是,在hcc患者来源的car(hyp7)t细胞中检测到27.9%的cd3+cd4-cd8-t细胞,称为“双阴性”(dn)t细胞。如图10e所示,最初用抗-cd3/cd28珠子致敏11天后,来自8个不同健康供体的car(hyp7)t细胞显示出15至60倍的扩增。相比之下,来自4个hcc患者的car(hyp7)t细胞在激活后第11天仅扩增了5到25倍。综上所述,基于hn3和hyp7抗体的cart细胞在表达水平、对gpc3的结合亲和力、转导效率和cd4+/cd8+t细胞比例方面具有相似的值。源自hcc患者的原代t细胞能够表达car并在培养中扩增。gpc3-靶向cart细胞的体外抗肿瘤活性为了确定靶向gpc3的t细胞是否可以特异性识别并杀死gpc3阳性肿瘤细胞,使用表达荧光素酶的肿瘤细胞建立了细胞溶解测定(cytolyticassay)。如图11a所示,过表达gpc3的a431(g1)细胞被car(hn3)和car(hyp7)t细胞有效地溶解,呈剂量依赖性。相比之下,两种cart细胞对包括a431和t3m4在内的gpc3阴性细胞均表现出最小的细胞溶解活性,表明cart细胞的特异性。还比较了来自健康供体和hcc患者的gpc3-靶向cart细胞的细胞溶解能力。在hep3b细胞上不同的e:t比下,car(hyp7)t细胞似乎比car(hn3)t细胞具有更高的溶解活性(图11b和图11c)。在e:t比为5时,hcc患者来源的car(hyp7)t细胞对hep3b细胞的溶解活性为30%至70%,平均为50%(图11c),其低于健康的供体来源的car(hyp7)t细胞的平均活性(90%)(图11b)。相比之下,在用任何来源的空白t细胞治疗的hep3b细胞中观察到的细胞溶解程度最低。有趣的是,car(hyp7)t细胞能够通过用抗cd3/cd28珠子的初始激活而进行长期扩增(图11d)。car(hyp7)t细胞在第14天或第28天在hep3b细胞中诱导相似水平的细胞死亡(图11e)。除hep3b细胞外,还在包括hepg2和huh-7的其他hcc细胞系中测试了gpc3cart细胞。如图11f所示,hepg2和huh-7细胞被car(hn3)和car(hyp7)t细胞溶解的程度比表达高水平gpc3的hep3b细胞的程度低。为了确定在长期共培养期间gpc3-靶向cart细胞是否会导致肿瘤细胞溶解增加,将car(hn3)或car(hyp7)cart细胞与hepg2细胞以2:1的e:t比孵育超过140小时。最初,与空白t细胞相比,任一种cart细胞都没有杀死hepg2细胞。在40小时及以后的时间里,car(hyp7)t细胞在清除hepg2细胞方面比car(hn3)t细胞(图11g)更有效。总的来说,与gpc3阳性肿瘤细胞共培养时,car(hyp7)t细胞显示出比car(hn3)t细胞更好的细胞溶解能力。gpc3-靶向cart细胞的体外多重细胞因子和趋化因子谱已经确定gpc3-靶向cart细胞可以抗原特异性方式识别gpc3阳性肿瘤细胞后,进行研究以确定4-1bb对暴露于hep3b或hepg2细胞后的car(hn3)或car(hyp7)t细胞的细胞因子和趋化因子谱的影响。如图12a所示,在gpc3-阳性肿瘤细胞的存在下,与空白-转导的t细胞相比,gpc3-特异性cart细胞产生了大量的所有细胞因子。当与hep3b或hepg2细胞共培养24小时时,两种cart细胞均分泌显著高水平的粒酶b(2500-13000pg/ml)(图12a)。还确定了4-1bb共刺激诱导高水平的th1细胞因子(gm-csf、ifn-γ、tnf-α和il-12)和th2细胞因子(il-5和il-13),这与th1/th2表型一致。然而,gpc3-特异性cart细胞仅表达低水平的il-21(<10pg/ml),其由th17细胞产生(图18)。除细胞因子外,car(hn3)和car(hyp7)t细胞分泌非常高水平的ccl-3和ccl-4趋化因子(400-2225pg/ml),所述趋化因子可能促进淋巴细胞浸润到肿瘤中(图12a)。本研究中分析的细胞因子和趋化因子的完整谱图如图18所示。总体而言,car(hyp7)t细胞比car(hn3)t细胞产生更多的细胞因子和趋化因子,这与两种car之间抗肿瘤活性的差异是一致的。gpc3-靶向cart细胞的基于单细胞的多功能性分析最近的研究表明,能够在单个细胞水平上共同产生多种细胞因子/趋化因子的t细胞,称为“多功能”t细胞,是有助于发展出针对癌症的有效且持久的细胞免疫力的关键效应细胞(ahmadzadehetal.,blood114:1537-1544,2009;baitschetal.,jclininvest121:2350-2360,2011)。为了确定我们的cart细胞产物的多功能性,应用了高含量的单细胞多重细胞因子分析(luetal.,procnatlacadsciusa112:e607-615,2015;maetal.,cancerdiscov3:418-429,2013),其使得能在体外用gpc3抗原鉴定刺激后会产生2种或更多种细胞因子的多功能t细胞亚群。32-plexpanel包括t细胞的关键免疫元件。与空白t细胞相比,hep3b细胞-刺激的cart细胞显示多功能细胞的百分比增加。当用hep3b细胞刺激时,car(hyp7)t细胞比car(hn3)t细胞具有更高的多功能性。类似地,与a431细胞刺激相比,当受g1细胞刺激时,在cd4+和cd8+cart细胞中均观察到增强的多功能性。还注意到,cd8+t细胞比cd4+t细胞更具多功能性。此外,先前描述的多功能强度指数(psi)用于量化多功能t细胞的集合效应(collectiveimpact)(maetal.,cancerdiscov3:418-429,2013)。样品的psi定义为多功能细胞的百分比乘以这些细胞分泌的细胞因子的平均信号强度。将psi按细胞因子功能-效应子、刺激性、调节性和炎性进行分解,以突显各组对样品整体多功能性的贡献。尽管效应细胞因子有助于cd4+car(hyp7)t细胞和大多数cd8+car(hyp7)t细胞的所有多功能性,但在受hep3b细胞刺激的cd8+car(hyp7)t细胞中观察到小比例的调节性细胞因子scd137。此外,cd8+car(hyp7)t细胞与g1细胞共培养时,会产生更多的效应子和趋化分子(例如ccl-3和ccl-4),这与使用luminex测定测量的细胞因子释放是一致的。为了区分样品中的所有多功能亚群,使用了多功能热图可视化。与受hep3b或g1细胞刺激后的car(hn3)t细胞相比,car(hyp7)t细胞具有更高频率的表达最多的功能组。在hep3b刺激下,由cd8+car(hyp7)t细胞表达了含有粒酶b、inf-γ、穿孔素和scd137的4-plex组。g1刺激的cd8+car(hyp7)t细胞更具多功能性,并分泌了含颗粒酶b、inf-γ、ccl-3、ccl-4、穿孔素、tnf-α和scd137的7-plex组。综上所述,car(hyp7)刺激了多功能t细胞(尤其是cd8+细胞毒性t细胞)更强大的激活和扩增。wnt信号传导对gpc3-靶向cart-细胞治疗的影响以前的研究表明,gpc3与wnt配体相互作用,并通过促进wnt/frizzled结合来促进hcc细胞增殖(capurroetal.,cancerres65:6245-6254,2005;gaoetal.,hepatology60:576-587,2014;gaoetal.,natcomm6:6536,2015)。为了确定gpc3-靶向cart细胞是否能影响wnt信号传导,测量了活性和总β-连环蛋白的水平。如图13a所示,与hep3b细胞以10:1的e:t比例共培养6小时后与空白t细胞相比,car(hyp7)t细胞明显降低了活性-β-连环蛋白和总β-连环蛋白的表达。活性-β-连环蛋白表达的降低甚至开始于hep3b细胞中car(hyp7)t细胞治疗的2小时(图13b)。此外,car(hyp7)t细胞诱导了hep3b细胞的凋亡,如裂解的多聚(adp核糖)聚合酶(parp)和裂解的半胱天冬酶9的表达升高所证明的。然而,在孵育6小时后,car(hn3)t细胞既不抑制β-连环蛋白的表达,也不诱导hep3b细胞凋亡。为了进一步研究靶向gpc3是否通过抑制wnt信号传导来下调hcc肿瘤生长,使用了crispr/cas9技术对hep3b细胞中的gpc3基因进行基因编辑。用编码靶向gpc3的不同外显子的小向导rna(smallguidingrna,sgrna)的构建体转染细胞会导致gpc3蛋白的大量减少,尤其是外显子5-靶向的sgrna(5-2)降低了超过95%的gpc3表达(图13c)。该5-2sgrna处理下调了活性β-连环蛋白和总β-连环蛋白的表达(图13c),并导致显著的细胞死亡。因此,car(hyp7)t细胞和crispr/cas9-介导的gpc3的基因编辑均抑制hcc细胞中的wnt/β-连环蛋白信号传导。在体外实验之后,测试了用crispr/cas9平台靶向gpc3是否会阻碍小鼠中的hcc肿瘤生长。将hep3b细胞皮下接种到裸鼠中。肿瘤接种后四周,给小鼠静脉内注射空质粒或编码5-2sgrna的质粒。如图13d所示,与用空质粒治疗的小鼠相比,用5-2sgrna质粒治疗的小鼠中肿瘤生长明显得到抑制。重要的是,5-2sgrna治疗还导致肿瘤中活性-β-连环蛋白和总β-连环蛋白水平的下降(图13e)。甲胎蛋白(afp)在过去的几十年中一直是hcc最广泛使用的生物标志物(ius,voprmedkhim10:90-91,1964)。20ng/ml的血清浓度是区分hcc患者和健康成人的常用临界值(trevisanietal.,jhepatol34:570-575,2001)。具有高afp浓度(≥400ng/ml)的hcc患者往往具有更大的肿瘤尺寸,肿块或弥漫性类型,和较低的中位生存率(fujiokaetal.,hepatology34:1128-1134,2001;tangkijvanichetal.,jclingastroenterol31:302-308,2000)。在crispr/cas9-介导的gpc3编辑之前和之后测量血清afp。如图13f所示,注射5-2sgrna的小鼠的afp血清水平明显低于注射空质粒的小鼠,表明afp水平与肿瘤尺寸呈正相关。总体而言,该数据表明靶向gpc3可能通过抑制wnt/β-连环蛋白信号传导来抑制hcc肿瘤生长。car(hyp7)t细胞使异种移植小鼠模型中的hcc肿瘤退缩(regression)为了评估gpc3-特异性cart细胞的体内抗肿瘤活性,向nsg小鼠腹膜内注射表达荧光素酶的hep3b细胞(hep3b-luc)。12天后,腹膜内单次输注空白或cart细胞(图14a)。治疗后两周,与空白t细胞治疗组相比,不同剂量car(hyp7)t细胞治疗的组均显示出减轻的肿瘤负荷(图14b和图14c)。尽管car(hn3)t细胞在体外对hep3b细胞表现出适度的细胞溶解活性,但在用car(hn3)t细胞治疗的小鼠中未观察到明显的肿瘤生长抑制作用。值得注意的是,到第70天,接受2000万个car(hyp7)t细胞的nsg小鼠100%存活且没有复发,而500万个car(hyp7)t细胞治疗组的存活仅为50%(图14d)。尽管gpc3-靶向cart细胞最初引起体重减轻,但小鼠体重逐渐增加(图19a)。此外,治疗两周后,用500万个(平均:400ng/ml)或1000万个(平均:300ng/ml)car(hyp7)t细胞治疗的小鼠的血清afp水平显著低于空白t细胞治疗的小鼠(平均值:20000ng/ml)(图14e)。值得注意的是,用2000万car(hyp7)t细胞治疗的小鼠的afp水平在25-78ng/ml范围内,接近成人的临界值(20ng/ml)。遗传修饰的t细胞的强大的体内扩增和持久性也被认为是癌症患者中持久临床缓解的关键预测指标。为了了解输注的cart细胞的持久性,使用利用car特异性扩增物的微滴式数字pcr(ddpcr)来评估cart细胞的百分比。如图14f所示,治疗3周后,在500万个car(hyp7)组中发现22.1%的car表达,而在500万个car(hn3)组中仅检测到1.3%的car。此外,在1000万个car(hyp7)组中检测到26.5%的car整合,表明随着时间的推移,肿瘤负荷与t细胞持久性之间呈负相关。相比之下,在1000万cd19cart细胞治疗的小鼠中,仅检测到2.2%的car表达,这表明肿瘤抗原识别驱动了体内输注的t细胞的存活。从hep3b腹膜播散小鼠模型中,发现小鼠在肝脏以及腹腔中的其他组织器官上发生了肿瘤病变。有趣的是,在用500万car(hyp7)t细胞治疗的小鼠中hep3b肿瘤局部生长,并局限于远离小鼠肝脏的脂肪组织,这表明car(hyp7)t细胞可阻止肿瘤在肝脏中播种和生长以及扩散到例如肾脏,肺和心脏的其他器官。还在hepg2腹膜播散异种移植小鼠模型中评估了car(hyp7)t细胞的功效。向nsg小鼠腹膜内注射表达荧光素酶的hepg2细胞(hepg2-luc),然后在12天后注射2000万个car(hyp7)t细胞(图15a)。如图15b和图15c所示,car(图15a)t细胞将肿瘤负荷降低至背景水平,且在第21天时肿瘤通量远低于接受空白t细胞治疗的小鼠,进一步证明了car(hyp7)t细胞具有优越的抗肿瘤功效。用car(hyp7)t细胞治疗的小鼠体重暂时下降,但体重恢复到基线水平,此后保持稳定(图19b),这与瞬时细胞因子释放综合征相一致。经过5周的car(图19b)t细胞治疗后,ddpcr在来自肿瘤和小鼠脾脏的基因组dna中分别检测到35.6%和19.5%的car表达(图15d)。相反,cd19cart细胞在任一组织中均未显示基因整合的迹象。此外,人hepg2细胞迁移到小鼠肝脏,car(hyp7)t细胞限制了肿瘤细胞的扩散,类似于在hep3b腹膜异种移植小鼠模型中观察到的情况。在原位hcc小鼠模型中进一步检查了car(hyp7)t细胞的抗肿瘤活性,因为它与临床更为相关。将50万个hep3b-luc细胞注射到nsg小鼠的肝脏中,并通过生物发光成像确认了肿瘤的植入(图16a)。在第21天,将car(hyp7)t细胞腹膜内或静脉内输注入小鼠中。尽管与对照组相比,car(hyp7)t细胞的两种施用途径均导致肿瘤尺寸减小并明显抑制了肿瘤生长,但在小鼠中静脉注射cart细胞(hyp7iv)导致的肿瘤退缩比腹膜内注射cart细胞(hyp7ip)更强(图16b-16c)。在该研究结束时,hyp7iv组的4只小鼠中有3只无肝肿瘤,而空白t细胞组的所有小鼠都带有大肿瘤。如图16d所示,cart细胞输注后5周,在来自hyp7iv组的小鼠脾脏中检出30.3%的car,在来自hyp7ip组的小鼠脾脏中仅存在8.6%的car,这与小鼠中每一种施用途径的抗肿瘤功效一致。此外,发光成像表明hep3b细胞在小鼠肝脏中生长,并且car(hyp7)t细胞的输注可显著抑制小鼠中的肿瘤生长或完全消除肿瘤细胞。最后,进行了毒理学研究,以评估car(hyp7)t细胞治疗的副作用。hyp7静脉注射组的小鼠显示白细胞和中性粒细胞增加,这可能与体内快速免疫反应有关(表8)。此外,通过尾静脉接受car(hyp7)t细胞的一只小鼠的丙氨酸氨基转移酶(alt)活性升高。但是,在小鼠尸检后未发现任何明显的肝损害证据。除肺外,治疗小鼠的所有器官重量均与对照组相似。在任何其他测量参数中均未检测到显著差异。总之,这些结果表明,car(hyp7)t细胞可以诱导小鼠hcc肿瘤的完全退缩。表8.hep3b原位异种移植小鼠中car(hyp7)t细胞的毒性鉴于该公开发明的原理可以应用于许多可能的实施方式,应当认识到,所阐述的实施方式仅是本发明的实例,并不应当理解成限定本发明的范围。相反,本发明的范围由所附权利要求限定。因此,我们要求保护所有落入这些权利要求的范围和精神内的发明。序列表<110>美国政府(由卫生和人类服务部的部长所代表)<120>靶向肿瘤抗原的嵌合抗原受体<130>4239-99836-02<150>us62/584,421<151>2017-11-10<160>51<170>patentinversion3.5<210>1<211>66<212>dna<213>人工序列<220><223>合成多核苷酸(gmcsfrss)<400>1atgcttctcctggtgacaagccttctgctctgtgagttaccacacccagcattcctcctg60atccca66<210>2<211>22<212>prt<213>人工序列<220><223>合成肽(gmcsfrss)<400>2metleuleuleuvalthrserleuleuleucysgluleuprohispro151015alapheleuleuilepro20<210>3<211>135<212>dna<213>人工序列<220><223>合成多核苷酸(cd8alpha铰链)<400>3accacgacgccagcgccgcgaccaccaacaccggcgcccaccatcgcgtcgcagcccctg60tccctgcgcccagaggcgtgccggccagcggcggggggcgcagtgcacacgagggggctg120gacttcgcctgtgac135<210>4<211>45<212>prt<213>人工序列<220><223>合成多肽(cd8alpha铰链)<400>4thrthrthrproalaproargproprothrproalaprothrileala151015serglnproleuserleuargproglualacysargproalaalagly202530glyalavalhisthrargglyleuaspphealacysasp354045<210>5<211>63<212>dna<213>人工序列<220><223>合成多核苷酸(cd8alphatm)<400>5atctacatctgggcgcccttggccgggacttgtggggtccttctcctgtcactggttatc60acc63<210>6<211>21<212>prt<213>人工序列<220><223>合成多肽(cd8alphatm)<400>6iletyriletrpalaproleualaglythrcysglyvalleuleuleu151015serleuvalilethr20<210>7<211>126<212>dna<213>人工序列<220><223>合成多核苷酸(4-1bb)<400>7aaacggggcagaaagaaactcctgtatatattcaaacaaccatttatgagaccagtacaa60actactcaagaggaagatggctgtagctgccgatttccagaagaagaagaaggaggatgt120gaactg126<210>8<211>42<212>prt<213>人工序列<220><223>合成多肽(4-1bb)<400>8lysargglyarglyslysleuleutyrilephelysglnprophemet151015argprovalglnthrthrglnglugluaspglycyssercysargphe202530proglugluglugluglyglycysgluleu3540<210>9<211>336<212>dna<213>人工序列<220><223>合成多核苷酸(cd3zeta信号传导结构域)<400>9agagtgaagttcagcaggagcgcagacgcccccgcgtaccagcagggccagaaccagctc60tataacgagctcaatctaggacgaagagaggagtacgatgttttggacaagagacgtggc120cgggaccctgagatggggggaaagccgagaaggaagaaccctcaggaaggcctgtacaat180gaactgcagaaagataagatggcggaggcctacagtgagattgggatgaaaggcgagcgc240cggaggggcaaggggcacgatggcctttaccagggtctcagtacagccaccaaggacacc300tacgacgcccttcacatgcaggccctgccccctcgc336<210>10<211>112<212>prt<213>人工序列<220><223>合成多肽(cd3zeta信号传导结构域)<400>10argvallyspheserargseralaaspalaproalatyrglnglngly151015glnasnglnleutyrasngluleuasnleuglyargarggluglutyr202530aspvalleuasplysargargglyargaspproglumetglyglylys354045proargarglysasnproglngluglyleutyrasngluleuglnlys505560asplysmetalaglualatyrsergluileglymetlysglygluarg65707580argargglylysglyhisaspglyleutyrglnglyleuserthrala859095thrlysaspthrtyraspalaleuhismetglnalaleuproproarg100105110<210>11<211>54<212>dna<213>人工序列<220><223>合成多核苷酸(t2a)<400>11gagggcagaggaagtcttctaacatgcggtgacgtggaggagaatcccggccct54<210>12<211>18<212>prt<213>人工序列<220><223>合成肽(t2a)<400>12gluglyargglyserleuleuthrcysglyaspvalglugluasnpro151015glypro<210>13<211>1005<212>dna<213>人工序列<220><223>合成多核苷酸(huegfrt)<400>13cgcaaagtgtgtaacggaataggtattggtgaatttaaagactcactctccataaatgct60acgaatattaaacacttcaaaaactgcacctccatcagtggcgatctccacatcctgccg120gtggcatttaggggtgactccttcacacatactcctcctctggatccacaggaactggat180attctgaaaaccgtaaaggaaatcacagggtttttgctgattcaggcttggcctgaaaac240aggacggacctccatgcctttgagaacctagaaatcatacgcggcaggaccaagcaacat300ggtcagttttctcttgcagtcgtcagcctgaacataacatccttgggattacgctccctc360aaggagataagtgatggagatgtgataatttcaggaaacaaaaatttgtgctatgcaaat420acaataaactggaaaaaactgtttgggacctccggtcagaaaaccaaaattataagcaac480agaggtgaaaacagctgcaaggccacaggccaggtctgccatgccttgtgctcccccgag540ggctgctggggcccggagcccagggactgcgtctcttgccggaatgtcagccgaggcagg600gaatgcgtggacaagtgcaaccttctggagggtgagccaagggagtttgtggagaactct660gagtgcatacagtgccacccagagtgcctgcctcaggccatgaacatcacctgcacagga720cggggaccagacaactgtatccagtgtgcccactacattgacggcccccactgcgtcaag780acctgcccggcaggagtcatgggagaaaacaacaccctggtctggaagtacgcagacgcc840ggccatgtgtgccacctgtgccatccaaactgcacctacggatgcactgggccaggtctt900gaaggctgtccaacgaatgggcctaagatcccgtccatcgccactgggatggtgggggcc960ctcctcttgctgctggtggtggccctggggatcggcctcttcatg1005<210>14<211>335<212>prt<213>人工序列<220><223>合成多肽(huegfrt)<400>14arglysvalcysasnglyileglyileglygluphelysaspserleu151015serileasnalathrasnilelyshisphelysasncysthrserile202530serglyaspleuhisileleuprovalalapheargglyaspserphe354045thrhisthrproproleuaspproglngluleuaspileleulysthr505560vallysgluilethrglypheleuleuileglnalatrpprogluasn65707580argthraspleuhisalaphegluasnleugluileileargglyarg859095thrlysglnhisglyglnpheserleualavalvalserleuasnile100105110thrserleuglyleuargserleulysgluileseraspglyaspval115120125ileileserglyasnlysasnleucystyralaasnthrileasntrp130135140lyslysleupheglythrserglyglnlysthrlysileileserasn145150155160argglygluasnsercyslysalathrglyglnvalcyshisalaleu165170175cysserprogluglycystrpglyprogluproargaspcysvalser180185190cysargasnvalserargglyargglucysvalasplyscysasnleu195200205leugluglygluproarggluphevalgluasnserglucysilegln210215220cyshisproglucysleuproglnalametasnilethrcysthrgly225230235240argglyproaspasncysileglncysalahistyrileaspglypro245250255hiscysvallysthrcysproalaglyvalmetglygluasnasnthr260265270leuvaltrplystyralaaspalaglyhisvalcyshisleucyshis275280285proasncysthrtyrglycysthrglyproglyleugluglycyspro290295300thrasnglyprolysileproserilealathrglymetvalglyala305310315320leuleuleuleuleuvalvalalaleuglyileglyleuphemet325330335<210>15<211>2601<212>dna<213>人工序列<220><223>合成构建体(pmh289)<400>15atgcttctcctggtgacaagccttctgctctgtgagttaccacacccagcattcctcctg60atcccacatatggaggtgcagcttgttgagtctggtggaggattggtgcagcctggaggg120tcattgagactctcatgtgcagcctctggattcaccttcaataagaatgccatgaattgg180gtccgccaggctccaggaaagggtttggaatgggttggccgcataagaaataaaactaat240aattatgcaacatattatgccgattcagtgaaagccaggtttaccatctccagagatgat300tcaaagaactcactctatctgcaaatgaacagcttgaaaaccgaggacacagccgtgtac360tattgtgtggctggtaactcgtttgcttactggggccaagggactctggtcactgtctct420gcaggcggaggcggatcaggtggtggcggatctggaggtggcggaagcgacattgtgatg480acccagtctccagactccctagctgtgtcactgggagagagggccactatcaactgcaag540tccagtcagagccttttatatagcagcaatcaaaagaactacttggcctggtaccaacag600aaaccagggcagcctcctaaactgctgatttactgggcatccagtagggaatctggggtc660cctgatcgcttcagtggcagtggatctgggacagatttcactctcaccatcagcagtctg720caggctgaagacgtggcagtttattactgtcagcaatattataactatccgctcacgttc780ggtcaggggaccaagttggagatcaaaactagtaccacgacgccagcgccgcgaccacca840acaccggcgcccaccatcgcgtcgcagcccctgtccctgcgcccagaggcgtgccggcca900gcggcggggggcgcagtgcacacgagggggctggacttcgcctgtgacatctacatctgg960gcgcccttggccgggacttgtggggtccttctcctgtcactggttatcaccaaacggggc1020agaaagaaactcctgtatatattcaaacaaccatttatgagaccagtacaaactactcaa1080gaggaagatggctgtagctgccgatttccagaagaagaagaaggaggatgtgaactgaga1140gtgaagttcagcaggagcgcagacgcccccgcgtaccagcagggccagaaccagctctat1200aacgagctcaatctaggacgaagagaggagtacgatgttttggacaagagacgtggccgg1260gaccctgagatggggggaaagccgagaaggaagaaccctcaggaaggcctgtacaatgaa1320ctgcagaaagataagatggcggaggcctacagtgagattgggatgaaaggcgagcgccgg1380aggggcaaggggcacgatggcctttaccagggtctcagtacagccaccaaggacacctac1440gacgcccttcacatgcaggccctgccccctcgcgagggcagaggaagtcttctaacatgc1500ggtgacgtggaggagaatcccggccctatgcttctcctggtgacaagccttctgctctgt1560gagttaccacacccagcattcctcctgatcccacgcaaagtgtgtaacggaataggtatt1620ggtgaatttaaagactcactctccataaatgctacgaatattaaacacttcaaaaactgc1680acctccatcagtggcgatctccacatcctgccggtggcatttaggggtgactccttcaca1740catactcctcctctggatccacaggaactggatattctgaaaaccgtaaaggaaatcaca1800gggtttttgctgattcaggcttggcctgaaaacaggacggacctccatgcctttgagaac1860ctagaaatcatacgcggcaggaccaagcaacatggtcagttttctcttgcagtcgtcagc1920ctgaacataacatccttgggattacgctccctcaaggagataagtgatggagatgtgata1980atttcaggaaacaaaaatttgtgctatgcaaatacaataaactggaaaaaactgtttggg2040acctccggtcagaaaaccaaaattataagcaacagaggtgaaaacagctgcaaggccaca2100ggccaggtctgccatgccttgtgctcccccgagggctgctggggcccggagcccagggac2160tgcgtctcttgccggaatgtcagccgaggcagggaatgcgtggacaagtgcaaccttctg2220gagggtgagccaagggagtttgtggagaactctgagtgcatacagtgccacccagagtgc2280ctgcctcaggccatgaacatcacctgcacaggacggggaccagacaactgtatccagtgt2340gcccactacattgacggcccccactgcgtcaagacctgcccggcaggagtcatgggagaa2400aacaacaccctggtctggaagtacgcagacgccggccatgtgtgccacctgtgccatcca2460aactgcacctacggatgcactgggccaggtcttgaaggctgtccaacgaatgggcctaag2520atcccgtccatcgccactgggatggtgggggccctcctcttgctgctggtggtggccctg2580gggatcggcctcttcatgtga2601<210>16<211>866<212>prt<213>人工序列<220><223>合成多肽(hyp7-car)<400>16metleuleuleuvalthrserleuleuleucysgluleuprohispro151015alapheleuleuileprohismetgluvalglnleuvalglusergly202530glyglyleuvalglnproglyglyserleuargleusercysalaala354045serglyphethrpheasnlysasnalametasntrpvalargglnala505560proglylysglyleuglutrpvalglyargileargasnlysthrasn65707580asntyralathrtyrtyralaaspservallysalaargphethrile859095serargaspaspserlysasnserleutyrleuglnmetasnserleu100105110lysthrgluaspthralavaltyrtyrcysvalalaglyasnserphe115120125alatyrtrpglyglnglythrleuvalthrvalseralaglyglygly130135140glyserglyglyglyglyserglyglyglyglyseraspilevalmet145150155160thrglnserproaspserleualavalserleuglygluargalathr165170175ileasncyslysserserglnserleuleutyrserserasnglnlys180185190asntyrleualatrptyrglnglnlysproglyglnproprolysleu195200205leuiletyrtrpalaserserarggluserglyvalproaspargphe210215220serglyserglyserglythraspphethrleuthrileserserleu225230235240glnalagluaspvalalavaltyrtyrcysglnglntyrtyrasntyr245250255proleuthrpheglyglnglythrlysleugluilelysthrserthr260265270thrthrproalaproargproprothrproalaprothrilealaser275280285glnproleuserleuargproglualacysargproalaalaglygly290295300alavalhisthrargglyleuaspphealacysaspiletyriletrp305310315320alaproleualaglythrcysglyvalleuleuleuserleuvalile325330335thrlysargglyarglyslysleuleutyrilephelysglnprophe340345350metargprovalglnthrthrglnglugluaspglycyssercysarg355360365pheproglugluglugluglyglycysgluleuargvallyspheser370375380argseralaaspalaproalatyrglnglnglyglnasnglnleutyr385390395400asngluleuasnleuglyargarggluglutyraspvalleuasplys405410415argargglyargaspproglumetglyglylysproargarglysasn420425430proglngluglyleutyrasngluleuglnlysasplysmetalaglu435440445alatyrsergluileglymetlysglygluargargargglylysgly450455460hisaspglyleutyrglnglyleuserthralathrlysaspthrtyr465470475480aspalaleuhismetglnalaleuproproarggluglyargglyser485490495leuleuthrcysglyaspvalglugluasnproglyprometleuleu500505510leuvalthrserleuleuleucysgluleuprohisproalapheleu515520525leuileproarglysvalcysasnglyileglyileglygluphelys530535540aspserleuserileasnalathrasnilelyshisphelysasncys545550555560thrserileserglyaspleuhisileleuprovalalaphearggly565570575aspserphethrhisthrproproleuaspproglngluleuaspile580585590leulysthrvallysgluilethrglypheleuleuileglnalatrp595600605progluasnargthraspleuhisalaphegluasnleugluileile610615620argglyargthrlysglnhisglyglnpheserleualavalvalser625630635640leuasnilethrserleuglyleuargserleulysgluileserasp645650655glyaspvalileileserglyasnlysasnleucystyralaasnthr660665670ileasntrplyslysleupheglythrserglyglnlysthrlysile675680685ileserasnargglygluasnsercyslysalathrglyglnvalcys690695700hisalaleucysserprogluglycystrpglyprogluproargasp705710715720cysvalsercysargasnvalserargglyargglucysvalasplys725730735cysasnleuleugluglygluproarggluphevalgluasnserglu740745750cysileglncyshisproglucysleuproglnalametasnilethr755760765cysthrglyargglyproaspasncysileglncysalahistyrile770775780aspglyprohiscysvallysthrcysproalaglyvalmetglyglu785790795800asnasnthrleuvaltrplystyralaaspalaglyhisvalcyshis805810815leucyshisproasncysthrtyrglycysthrglyproglyleuglu820825830glycysprothrasnglyprolysileproserilealathrglymet835840845valglyalaleuleuleuleuleuvalvalalaleuglyileglyleu850855860phemet865<210>17<211>2214<212>dna<213>人工序列<220><223>合成构建体(pmh288)<400>17atgcttctcctggtgacaagccttctgctctgtgagttaccacacccagcattcctcctg60atcccacatatgcaggtgcagctggtgcagtctgggggaggcttggtacagcctggaggg120tccctgagactctcctgtgcagcctcttatttcgatttcgattcttatgaaatgagctgg180gtccgccaggctccagggaagggcctagagtggattgggagtatctatcatagtgggagc240acctactacaacccgtccctcaagagtcgagtcaccatctccagagacaattccaagaac300acgctgtatctgcaaatgaacaccctgagagccgaggacacagccacgtattactgtgcg360agagtaaatatggaccgatttgactactggggccagggaaccctggtcaccgtctcctca420actagtaccacgacgccagcgccgcgaccaccaacaccggcgcccaccatcgcgtcgcag480cccctgtccctgcgcccagaggcgtgccggccagcggcggggggcgcagtgcacacgagg540gggctggacttcgcctgtgacatctacatctgggcgcccttggccgggacttgtggggtc600cttctcctgtcactggttatcaccaaacggggcagaaagaaactcctgtatatattcaaa660caaccatttatgagaccagtacaaactactcaagaggaagatggctgtagctgccgattt720ccagaagaagaagaaggaggatgtgaactgagagtgaagttcagcaggagcgcagacgcc780cccgcgtaccagcagggccagaaccagctctataacgagctcaatctaggacgaagagag840gagtacgatgttttggacaagagacgtggccgggaccctgagatggggggaaagccgaga900aggaagaaccctcaggaaggcctgtacaatgaactgcagaaagataagatggcggaggcc960tacagtgagattgggatgaaaggcgagcgccggaggggcaaggggcacgatggcctttac1020cagggtctcagtacagccaccaaggacacctacgacgcccttcacatgcaggccctgccc1080cctcgcgagggcagaggaagtcttctaacatgcggtgacgtggaggagaatcccggccct1140atgcttctcctggtgacaagccttctgctctgtgagttaccacacccagcattcctcctg1200atcccacgcaaagtgtgtaacggaataggtattggtgaatttaaagactcactctccata1260aatgctacgaatattaaacacttcaaaaactgcacctccatcagtggcgatctccacatc1320ctgccggtggcatttaggggtgactccttcacacatactcctcctctggatccacaggaa1380ctggatattctgaaaaccgtaaaggaaatcacagggtttttgctgattcaggcttggcct1440gaaaacaggacggacctccatgcctttgagaacctagaaatcatacgcggcaggaccaag1500caacatggtcagttttctcttgcagtcgtcagcctgaacataacatccttgggattacgc1560tccctcaaggagataagtgatggagatgtgataatttcaggaaacaaaaatttgtgctat1620gcaaatacaataaactggaaaaaactgtttgggacctccggtcagaaaaccaaaattata1680agcaacagaggtgaaaacagctgcaaggccacaggccaggtctgccatgccttgtgctcc1740cccgagggctgctggggcccggagcccagggactgcgtctcttgccggaatgtcagccga1800ggcagggaatgcgtggacaagtgcaaccttctggagggtgagccaagggagtttgtggag1860aactctgagtgcatacagtgccacccagagtgcctgcctcaggccatgaacatcacctgc1920acaggacggggaccagacaactgtatccagtgtgcccactacattgacggcccccactgc1980gtcaagacctgcccggcaggagtcatgggagaaaacaacaccctggtctggaagtacgca2040gacgccggccatgtgtgccacctgtgccatccaaactgcacctacggatgcactgggcca2100ggtcttgaaggctgtccaacgaatgggcctaagatcccgtccatcgccactgggatggtg2160ggggccctcctcttgctgctggtggtggccctggggatcggcctcttcatgtga2214<210>18<211>737<212>prt<213>人工序列<220><223>合成多肽(hn3-car)<400>18metleuleuleuvalthrserleuleuleucysgluleuprohispro151015alapheleuleuileprohismetglnvalglnleuvalglnsergly202530glyglyleuvalglnproglyglyserleuargleusercysalaala354045sertyrpheasppheaspsertyrglumetsertrpvalargglnala505560proglylysglyleuglutrpileglyseriletyrhisserglyser65707580thrtyrtyrasnproserleulysserargvalthrileserargasp859095asnserlysasnthrleutyrleuglnmetasnthrleuargalaglu100105110aspthralathrtyrtyrcysalaargvalasnmetaspargpheasp115120125tyrtrpglyglnglythrleuvalthrvalserserthrserthrthr130135140thrproalaproargproprothrproalaprothrilealasergln145150155160proleuserleuargproglualacysargproalaalaglyglyala165170175valhisthrargglyleuaspphealacysaspiletyriletrpala180185190proleualaglythrcysglyvalleuleuleuserleuvalilethr195200205lysargglyarglyslysleuleutyrilephelysglnprophemet210215220argprovalglnthrthrglnglugluaspglycyssercysargphe225230235240proglugluglugluglyglycysgluleuargvallyspheserarg245250255seralaaspalaproalatyrglnglnglyglnasnglnleutyrasn260265270gluleuasnleuglyargarggluglutyraspvalleuasplysarg275280285argglyargaspproglumetglyglylysproargarglysasnpro290295300glngluglyleutyrasngluleuglnlysasplysmetalagluala305310315320tyrsergluileglymetlysglygluargargargglylysglyhis325330335aspglyleutyrglnglyleuserthralathrlysaspthrtyrasp340345350alaleuhismetglnalaleuproproarggluglyargglyserleu355360365leuthrcysglyaspvalglugluasnproglyprometleuleuleu370375380valthrserleuleuleucysgluleuprohisproalapheleuleu385390395400ileproarglysvalcysasnglyileglyileglygluphelysasp405410415serleuserileasnalathrasnilelyshisphelysasncysthr420425430serileserglyaspleuhisileleuprovalalapheargglyasp435440445serphethrhisthrproproleuaspproglngluleuaspileleu450455460lysthrvallysgluilethrglypheleuleuileglnalatrppro465470475480gluasnargthraspleuhisalaphegluasnleugluileilearg485490495glyargthrlysglnhisglyglnpheserleualavalvalserleu500505510asnilethrserleuglyleuargserleulysgluileseraspgly515520525aspvalileileserglyasnlysasnleucystyralaasnthrile530535540asntrplyslysleupheglythrserglyglnlysthrlysileile545550555560serasnargglygluasnsercyslysalathrglyglnvalcyshis565570575alaleucysserprogluglycystrpglyprogluproargaspcys580585590valsercysargasnvalserargglyargglucysvalasplyscys595600605asnleuleugluglygluproarggluphevalgluasnserglucys610615620ileglncyshisproglucysleuproglnalametasnilethrcys625630635640thrglyargglyproaspasncysileglncysalahistyrileasp645650655glyprohiscysvallysthrcysproalaglyvalmetglygluasn660665670asnthrleuvaltrplystyralaaspalaglyhisvalcyshisleu675680685cyshisproasncysthrtyrglycysthrglyproglyleuglugly690695700cysprothrasnglyprolysileproserilealathrglymetval705710715720glyalaleuleuleuleuleuvalvalalaleuglyileglyleuphe725730735met<210>19<211>2226<212>dna<213>人工序列<220><223>合成构建体(pmh290)<400>19atgcttctcctggtgacaagccttctgctctgtgagttaccacacccagcattcctcctg60atcccacatatgcaggtgcagctggtgcagtctgggggaggcttggtacagcctggaggg120tccctgagactctcctgtgcagcctctgatttctatttctatgattatgaaatgagctgg180gtccgccaggctccagggaagggtctggagtggattgggactgtctcctatagtgggagc240acctactacaacccgtccctcaagagtcgagtcaccatctccagagacaattccaagaac300acgctgtatctgcaaatgaacaccctaagagccgaggacacagccatgtattactgtgcg360agaggttacagctatgatgactcccgatattttgactactggggccagggaaccctggtc420accgtctcctcaactagtaccacgacgccagcgccgcgaccaccaacaccggcgcccacc480atcgcgtcgcagcccctgtccctgcgcccagaggcgtgccggccagcggcggggggcgca540gtgcacacgagggggctggacttcgcctgtgacatctacatctgggcgcccttggccggg600acttgtggggtccttctcctgtcactggttatcaccaaacggggcagaaagaaactcctg660tatatattcaaacaaccatttatgagaccagtacaaactactcaagaggaagatggctgt720agctgccgatttccagaagaagaagaaggaggatgtgaactgagagtgaagttcagcagg780agcgcagacgcccccgcgtaccagcagggccagaaccagctctataacgagctcaatcta840ggacgaagagaggagtacgatgttttggacaagagacgtggccgggaccctgagatgggg900ggaaagccgagaaggaagaaccctcaggaaggcctgtacaatgaactgcagaaagataag960atggcggaggcctacagtgagattgggatgaaaggcgagcgccggaggggcaaggggcac1020gatggcctttaccagggtctcagtacagccaccaaggacacctacgacgcccttcacatg1080caggccctgccccctcgcgagggcagaggaagtcttctaacatgcggtgacgtggaggag1140aatcccggccctatgcttctcctggtgacaagccttctgctctgtgagttaccacaccca1200gcattcctcctgatcccacgcaaagtgtgtaacggaataggtattggtgaatttaaagac1260tcactctccataaatgctacgaatattaaacacttcaaaaactgcacctccatcagtggc1320gatctccacatcctgccggtggcatttaggggtgactccttcacacatactcctcctctg1380gatccacaggaactggatattctgaaaaccgtaaaggaaatcacagggtttttgctgatt1440caggcttggcctgaaaacaggacggacctccatgcctttgagaacctagaaatcatacgc1500ggcaggaccaagcaacatggtcagttttctcttgcagtcgtcagcctgaacataacatcc1560ttgggattacgctccctcaaggagataagtgatggagatgtgataatttcaggaaacaaa1620aatttgtgctatgcaaatacaataaactggaaaaaactgtttgggacctccggtcagaaa1680accaaaattataagcaacagaggtgaaaacagctgcaaggccacaggccaggtctgccat1740gccttgtgctcccccgagggctgctggggcccggagcccagggactgcgtctcttgccgg1800aatgtcagccgaggcagggaatgcgtggacaagtgcaaccttctggagggtgagccaagg1860gagtttgtggagaactctgagtgcatacagtgccacccagagtgcctgcctcaggccatg1920aacatcacctgcacaggacggggaccagacaactgtatccagtgtgcccactacattgac1980ggcccccactgcgtcaagacctgcccggcaggagtcatgggagaaaacaacaccctggtc2040tggaagtacgcagacgccggccatgtgtgccacctgtgccatccaaactgcacctacgga2100tgcactgggccaggtcttgaaggctgtccaacgaatgggcctaagatcccgtccatcgcc2160actgggatggtgggggccctcctcttgctgctggtggtggccctggggatcggcctcttc2220atgtga2226<210>20<211>741<212>prt<213>人工序列<220><223>合成多肽(lh7-car)<400>20metleuleuleuvalthrserleuleuleucysgluleuprohispro151015alapheleuleuileprohismetglnvalglnleuvalglnsergly202530glyglyleuvalglnproglyglyserleuargleusercysalaala354045seraspphetyrphetyrasptyrglumetsertrpvalargglnala505560proglylysglyleuglutrpileglythrvalsertyrserglyser65707580thrtyrtyrasnproserleulysserargvalthrileserargasp859095asnserlysasnthrleutyrleuglnmetasnthrleuargalaglu100105110aspthralamettyrtyrcysalaargglytyrsertyraspaspser115120125argtyrpheasptyrtrpglyglnglythrleuvalthrvalserser130135140thrserthrthrthrproalaproargproprothrproalaprothr145150155160ilealaserglnproleuserleuargproglualacysargproala165170175alaglyglyalavalhisthrargglyleuaspphealacysaspile180185190tyriletrpalaproleualaglythrcysglyvalleuleuleuser195200205leuvalilethrlysargglyarglyslysleuleutyrilephelys210215220glnprophemetargprovalglnthrthrglnglugluaspglycys225230235240sercysargpheproglugluglugluglyglycysgluleuargval245250255lyspheserargseralaaspalaproalatyrglnglnglyglnasn260265270glnleutyrasngluleuasnleuglyargarggluglutyraspval275280285leuasplysargargglyargaspproglumetglyglylysproarg290295300arglysasnproglngluglyleutyrasngluleuglnlysasplys305310315320metalaglualatyrsergluileglymetlysglygluargargarg325330335glylysglyhisaspglyleutyrglnglyleuserthralathrlys340345350aspthrtyraspalaleuhismetglnalaleuproproargglugly355360365argglyserleuleuthrcysglyaspvalglugluasnproglypro370375380metleuleuleuvalthrserleuleuleucysgluleuprohispro385390395400alapheleuleuileproarglysvalcysasnglyileglyilegly405410415gluphelysaspserleuserileasnalathrasnilelyshisphe420425430lysasncysthrserileserglyaspleuhisileleuprovalala435440445pheargglyaspserphethrhisthrproproleuaspproglnglu450455460leuaspileleulysthrvallysgluilethrglypheleuleuile465470475480glnalatrpprogluasnargthraspleuhisalaphegluasnleu485490495gluileileargglyargthrlysglnhisglyglnpheserleuala500505510valvalserleuasnilethrserleuglyleuargserleulysglu515520525ileseraspglyaspvalileileserglyasnlysasnleucystyr530535540alaasnthrileasntrplyslysleupheglythrserglyglnlys545550555560thrlysileileserasnargglygluasnsercyslysalathrgly565570575glnvalcyshisalaleucysserprogluglycystrpglyproglu580585590proargaspcysvalsercysargasnvalserargglyargglucys595600605valasplyscysasnleuleugluglygluproarggluphevalglu610615620asnserglucysileglncyshisproglucysleuproglnalamet625630635640asnilethrcysthrglyargglyproaspasncysileglncysala645650655histyrileaspglyprohiscysvallysthrcysproalaglyval660665670metglygluasnasnthrleuvaltrplystyralaaspalaglyhis675680685valcyshisleucyshisproasncysthrtyrglycysthrglypro690695700glyleugluglycysprothrasnglyprolysileproserileala705710715720thrglymetvalglyalaleuleuleuleuleuvalvalalaleugly725730735ileglyleuphemet740<210>21<211>351<212>dna<213>人工序列<220><223>合成多核苷酸(yp7vh结构域)<400>21gaggtgcagcttgttgagactggtggaggaatggtgcagcctgaagggtcattgaaactc60tcatgtgcagcctctggattcaccttcaataagaatgccatgaattgggtccgccaggct120ccaggaaagggtttggaatgggttgctcgcataagaaataaaactaataattatgcaaca180tattatgccgattcagtgaaagccaggtttaccatctccagagatgattcacaaagcatg240ctctatctgcaaatgaacaacttgaaaattgaggacacagccatgtactattgtgtggct300ggtaactcgtttgcttactggggccaagggactctggtcactgtctctgca351<210>22<211>117<212>prt<213>人工序列<220><223>合成多肽(yp7vh结构域)<400>22gluvalglnleuvalgluthrglyglyglymetvalglnproglugly151015serleulysleusercysalaalaserglyphethrpheasnlysasn202530alametasntrpvalargglnalaproglylysglyleuglutrpval354045alaargileargasnlysthrasnasntyralathrtyrtyralaasp505560servallysalaargphethrileserargaspaspserglnsermet65707580leutyrleuglnmetasnasnleulysilegluaspthralamettyr859095tyrcysvalalaglyasnserphealatyrtrpglyglnglythrleu100105110valthrvalserala115<210>23<211>339<212>dna<213>人工序列<220><223>合成多核苷酸(yp7vl结构域)<400>23gacattgtgatgtcacagtctccatcctccctagttgtgtcaattggagagaaggttact60atgacctgcaagtccagtcagagccttttatatagcagcaatcaaaagaactacttggcc120tggtaccaacagaaaccagggcagtctcctaaactgctgatttactgggcatccagtagg180gaatctggggtccctgatcgcttcacaggcagtggatctgggacagatttcactctcacc240atcagcagtgtgaaggctgaagacctggcagtttattactgtcagcaatattataactat300ccgctcacgttcggtgctgggaccaagttggagctgaaa339<210>24<211>113<212>prt<213>人工序列<220><223>合成多肽(yp7vl结构域)<400>24aspilevalmetserglnserproserserleuvalvalserilegly151015glulysvalthrmetthrcyslysserserglnserleuleutyrser202530serasnglnlysasntyrleualatrptyrglnglnlysproglygln354045serprolysleuleuiletyrtrpalaserserarggluserglyval505560proaspargphethrglyserglyserglythraspphethrleuthr65707580ileserservallysalagluaspleualavaltyrtyrcysglngln859095tyrtyrasntyrproleuthrpheglyalaglythrlysleugluleu100105110lys<210>25<211>351<212>dna<213>人工序列<220><223>合成多核苷酸(hyp7vh结构域)<400>25gaggtgcagcttgttgagtctggtggaggattggtgcagcctggagggtcattgagactc60tcatgtgcagcctctggattcaccttcaataagaatgccatgaattgggtccgccaggct120ccaggaaagggtttggaatgggttggccgcataagaaataaaactaataattatgcaaca180tattatgccgattcagtgaaagccaggtttaccatctccagagatgattcaaagaactca240ctctatctgcaaatgaacagcttgaaaaccgaggacacagccgtgtactattgtgtggct300ggtaactcgtttgcttactggggccaagggactctggtcactgtctctgca351<210>26<211>117<212>prt<213>人工序列<220><223>合成多肽(hyp7vh结构域)<400>26gluvalglnleuvalgluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrpheasnlysasn202530alametasntrpvalargglnalaproglylysglyleuglutrpval354045glyargileargasnlysthrasnasntyralathrtyrtyralaasp505560servallysalaargphethrileserargaspaspserlysasnser65707580leutyrleuglnmetasnserleulysthrgluaspthralavaltyr859095tyrcysvalalaglyasnserphealatyrtrpglyglnglythrleu100105110valthrvalserala115<210>27<211>339<212>dna<213>人工序列<220><223>合成多核苷酸(hyp7vl结构域)<400>27gacattgtgatgacccagtctccagactccctagctgtgtcactgggagagagggccact60atcaactgcaagtccagtcagagccttttatatagcagcaatcaaaagaactacttggcc120tggtaccaacagaaaccagggcagcctcctaaactgctgatttactgggcatccagtagg180gaatctggggtccctgatcgcttcagtggcagtggatctgggacagatttcactctcacc240atcagcagtctgcaggctgaagacgtggcagtttattactgtcagcaatattataactat300ccgctcacgttcggtcaggggaccaagttggagatcaaa339<210>28<211>113<212>prt<213>人工序列<220><223>合成多肽(hyp7vl结构域)<400>28aspilevalmetthrglnserproaspserleualavalserleugly151015gluargalathrileasncyslysserserglnserleuleutyrser202530serasnglnlysasntyrleualatrptyrglnglnlysproglygln354045proprolysleuleuiletyrtrpalaserserarggluserglyval505560proaspargpheserglyserglyserglythraspphethrleuthr65707580ileserserleuglnalagluaspvalalavaltyrtyrcysglngln859095tyrtyrasntyrproleuthrpheglyglnglythrlysleugluile100105110lys<210>29<211>351<212>dna<213>人工序列<220><223>合成多核苷酸(hn3)<400>29caggtgcagctggtgcagtctgggggaggcttggtacagcctggagggtccctgagactc60tcctgtgcagcctcttatttcgatttcgattcttatgaaatgagctgggtccgccaggct120ccagggaagggcctagagtggattgggagtatctatcatagtgggagcacctactacaac180ccgtccctcaagagtcgagtcaccatctccagagacaattccaagaacacgctgtatctg240caaatgaacaccctgagagccgaggacacagccacgtattactgtgcgagagtaaatatg300gaccgatttgactactggggccagggaaccctggtcaccgtctcctcaagt351<210>30<211>117<212>prt<213>人工序列<220><223>合成多肽(hn3)<400>30glnvalglnleuvalglnserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalasertyrpheasppheaspsertyr202530glumetsertrpvalargglnalaproglylysglyleuglutrpile354045glyseriletyrhisserglyserthrtyrtyrasnproserleulys505560serargvalthrileserargaspasnserlysasnthrleutyrleu65707580glnmetasnthrleuargalagluaspthralathrtyrtyrcysala859095argvalasnmetaspargpheasptyrtrpglyglnglythrleuval100105110thrvalserserser115<210>31<211>360<212>dna<213>人工序列<220><223>合成多核苷酸(lh7)<400>31caggtgcagctggtgcagtctgggggaggcttggtacagcctggagggtccctgagactc60tcctgtgcagcctctgatttctatttctatgattatgaaatgagctgggtccgccaggct120ccagggaagggtctggagtggattgggactgtctcctatagtgggagcacctactacaac180ccgtccctcaagagtcgagtcaccatctccagagacaattccaagaacacgctgtatctg240caaatgaacaccctaagagccgaggacacagccatgtattactgtgcgagaggttacagc300tatgatgactcccgatattttgactactggggccagggaaccctggtcaccgtctcctca360<210>32<211>120<212>prt<213>人工序列<220><223>合成多肽(lh7)<400>32glnvalglnleuvalglnserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaseraspphetyrphetyrasptyr202530glumetsertrpvalargglnalaproglylysglyleuglutrpile354045glythrvalsertyrserglyserthrtyrtyrasnproserleulys505560serargvalthrileserargaspasnserlysasnthrleutyrleu65707580glnmetasnthrleuargalagluaspthralamettyrtyrcysala859095argglytyrsertyraspaspserargtyrpheasptyrtrpglygln100105110glythrleuvalthrvalserser115120<210>33<211>360<212>dna<213>人工序列<220><223>合成多核苷酸(lh4)<400>33caggtgcagctggtgcagtctgggggaggcttggtacagcctggagggtccctgagactc60tcctgtgcagcctcttctttctatttcgatgattatgaaatgagctgggtccgccaggct120ccagggaaggccctggagtggattgggcgtatctataccagtgggagcaccaactacaac180ccctccctcaagagtcgagtcaccatctccagagacaattccaagaacacgctgtatctg240caaatgaacaccctgagagccgaggacacagccacgtattactgtgcgaggggatattgt300agtggtggtagctgctactttgactactggggccagggaaccctggtcaccgtctcctca360<210>34<211>120<212>prt<213>人工序列<220><223>合成多肽(lh4)<400>34glnvalglnleuvalglnserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserserphetyrpheaspasptyr202530glumetsertrpvalargglnalaproglylysalaleuglutrpile354045glyargiletyrthrserglyserthrasntyrasnproserleulys505560serargvalthrileserargaspasnserlysasnthrleutyrleu65707580glnmetasnthrleuargalagluaspthralathrtyrtyrcysala859095argglytyrcysserglyglysercystyrpheasptyrtrpglygln100105110glythrleuvalthrvalserser115120<210>35<211>363<212>dna<213>人工序列<220><223>合成多核苷酸(lh6)<400>35caggtgcagctggtgcagtctgggggaggcttggtacagcctggagggtccctgagactc60tcctgtgcagcctctgatttctatttcgatgattatgaaatgagctgggtccgccaggct120ccagggaaggggctggagtgggtctcaactattagtggtagtggtggtggcacatactac180gcagactcagtgaagggccgattcaccatctccagagacaattccaagaacacgctgtat240ctgcaaatgaacaccctgagagccgaggacacagccacatattactgtgcgagaggttac300agttatgacgactcccgatattttgactactggggccagggaaccctggtcaccgtctcc360tca363<210>36<211>121<212>prt<213>人工序列<220><223>合成多肽(lh6)<400>36glnvalglnleuvalglnserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaseraspphetyrpheaspasptyr202530glumetsertrpvalargglnalaproglylysglyleuglutrpval354045serthrileserglyserglyglyglythrtyrtyralaaspserval505560lysglyargphethrileserargaspasnserlysasnthrleutyr65707580leuglnmetasnthrleuargalagluaspthralathrtyrtyrcys859095alaargglytyrsertyraspaspserargtyrpheasptyrtrpgly100105110glnglythrleuvalthrvalserser115120<210>37<211>369<212>dna<213>人工序列<220><223>合成多核苷酸(yp218vh结构域)<400>37cagcagcagctggaggagtccgggggaggcctggtcaagcctgagggatccctgacactc60acctgcaaagcctctggattcgacctcggtttctacttttacgcctgttgggtccgccag120gctccagggaagggcctggagtggatcgcatgcatttatactgctggtagtggtagcacg180tactacgcgagctgggcgaaaggccgattcaccatctccaaagcctcgtcgaccacggtg240actctgcaaatgaccagtctggcagccgcggacacggccacctatttctgtgcgagatct300actgctaatactagaagtacttattatcttaacttgtggggcccaggcaccctggtcacc360gtctcctca369<210>38<211>123<212>prt<213>人工序列<220><223>合成多肽(yp218vh结构域)<400>38glnglnglnleuglugluserglyglyglyleuvallysproglugly151015serleuthrleuthrcyslysalaserglypheaspleuglyphetyr202530phetyralacystrpvalargglnalaproglylysglyleuglutrp354045ilealacysiletyrthralaglyserglyserthrtyrtyralaser505560trpalalysglyargphethrileserlysalaserserthrthrval65707580thrleuglnmetthrserleualaalaalaaspthralathrtyrphe859095cysalaargserthralaasnthrargserthrtyrtyrleuasnleu100105110trpglyproglythrleuvalthrvalserser115120<210>39<211>330<212>dna<213>人工序列<220><223>合成多核苷酸(yp218vl结构域)<400>39gacgtcgtgatgacccagactccagcctccgtgtctgaacctgtgggaggcacagtcacc60atcaagtgccaggccagtcagaggattagtagttacttatcctggtatcagcagaaacca120gggcagcgtcccaagctcctgatctttggtgcatccactctggcatctggggtcccctcg180cggttcaaaggcagtggatctgggacagaatacactctcaccatcagcgacctggagtgt240gccgatgctgccacttactactgtcagagttatgcttattttgatagtaataattggcat300gctttcggcggagggaccgaggtggtggtc330<210>40<211>110<212>prt<213>人工序列<220><223>合成多肽(yp218vl结构域)<400>40aspvalvalmetthrglnthrproalaservalsergluprovalgly151015glythrvalthrilelyscysglnalaserglnargilesersertyr202530leusertrptyrglnglnlysproglyglnargprolysleuleuile354045pheglyalaserthrleualaserglyvalproserargphelysgly505560serglyserglythrglutyrthrleuthrileseraspleuglucys65707580alaaspalaalathrtyrtyrcysglnsertyralatyrpheaspser859095asnasntrphisalapheglyglyglythrgluvalvalval100105110<210>41<211>339<212>dna<213>人工序列<220><223>合成多核苷酸(sd1)<400>41caggtgcagctggtgcagtctgggggaggcttggtacagcctggagggtccctgagactc60tcctgtgcagcctctgatttcgatttcgctgcttatgaaatgagctgggtccgccaggct120ccaggacaaggccttgagtgggtggcaattatatcacatgatggaatcgataaatactac180acagactccgtgaagggccgattcaccatctccagagacaattccaagaacacgctgtat240ctgcaaatgaacaccctgagagccgaggacacagccacgtattactgtttaaggcttggt300gctgtaggccagggaaccctggtcaccgtctcctcaagt339<210>42<211>113<212>prt<213>人工序列<220><223>合成多肽(sd1)<400>42glnvalglnleuvalglnserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserasppheaspphealaalatyr202530glumetsertrpvalargglnalaproglyglnglyleuglutrpval354045alaileileserhisaspglyileasplystyrtyrthraspserval505560lysglyargphethrileserargaspasnserlysasnthrleutyr65707580leuglnmetasnthrleuargalagluaspthralathrtyrtyrcys859095leuargleuglyalavalglyglnglythrleuvalthrvalserser100105110ser<210>43<211>20<212>dna<213>人工序列<220><223>合成寡核苷酸<400>43gcagtctctggaagaaggag20<210>44<211>20<212>dna<213>人工序列<220><223>合成寡核苷酸<400>44tggtgacaggtggcgtccgg20<210>45<211>20<212>dna<213>人工序列<220><223>合成寡核苷酸<400>45cggttttccaaggtgagttc20<210>46<211>20<212>dna<213>人工序列<220><223>合成寡核苷酸<400>46ggtcacgtcttgctcctcgg20<210>47<211>20<212>dna<213>人工序列<220><223>合成寡核苷酸<400>47gacatcaatgagtgcctccg20<210>48<211>20<212>dna<213>人工序列<220><223>合成寡核苷酸<400>48gataataagcagatctatat20<210>49<211>20<212>dna<213>人工序列<220><223>合成寡核苷酸<400>49cgttttccgccacagggcta20<210>50<211>20<212>dna<213>人工序列<220><223>合成寡核苷酸<400>50agggtgtcgttttccgccac20<210>51<211>20<212>dna<213>人工序列<220><223>合成寡核苷酸<400>51gaggcagagcaggtagtcag20当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1