一种水稻MYB转录因子及其重组表达载体的应用的制作方法
本发明属于基因工程技术领域,涉及一种水稻myb转录因子及其重组表达载体的应用。
背景技术:
植物转录因子myb是近年来发现的一类与调控植物生长发育、生理代谢、细胞的形态和模式建成等生理过程有关的一类转录因子,在植物中普遍存在,同时也是植物中最大的转录家族之一,myb转录因子在植物的代谢和调控中发挥重要作用。大多数myb蛋白在n端含有一段氨基酸残基组成的myb结构域,根据这个高度保守的结构域的结构特征可将myb转录因子分为四类:1r-myb/myb-related;r2r3-myb;3r-myb;4r-myb(4个r1/r2的重复)。myb转录因子具有多种生物学功能,广泛参与植物根、茎、叶、花的生长发育,与此同时,myb基因家族对干旱、盐渍、冷害等非生物胁迫过程也有响应,此外myb转录因子还与某些经济作物的品质好坏密切相关。
控制人口过快增长与粮食增产是全球亟待解决的重大问题。水稻是重要的粮食作物以及单子叶植物的模式植物,以作物矮秆基因的发现和杂交育种利用为代表的第一次农业绿色革命中,矮杆水稻的应用提高了水稻的抗倒伏能力,是保证其高产的重要基础(jiaoetal.,2010;miuraetal.,2010)。另一方面,水稻的分蘖数和有效分蘖是评价水稻的重要指标,其对于产量有巨大的贡献作用,增加分蘖数在一定程度上可以有效的增加水稻产量,是近年来培育高产品种的一个重要指标。
技术实现要素:
本发明的目的在于提供水稻myb转录因子基因osmyb110的应用。
本发明的另一目的是提供该基因的过表达载体。
本发明的又一目的是提供该过表达载体的应用。
本发明的目的通过如下技术方案实现:
水稻myb类转录因子osmyb110在降低植物株高、增加分蘖、缩短花期和增加稻米品质等方面的应用,该基因在水稻注释计划数据库(rap_db)的序列号为os10g0478300。
其中,所述的植物优选单子叶植物,进一步优选水稻、玉米或小麦,特别优选水稻。
水稻myb类转录因子基因osmyb110的编码产物,水稻myb转录因子蛋白osmyb110降低植物株高、增加分蘖、缩短花期和增加稻米品质等方面的应用。
其中,所述的植物优选单子叶植物,进一步优选水稻、玉米或小麦,特别优选水稻。
含有水稻水稻myb类转录因子osmyb110的过表达载体,所述的过表达载体包含水稻水稻myb类转录因子osmyb110。
所述的过表达载体,优选以pcambia1305为出发载体,在ecori和saci酶切位点插入2x35scamv启动子,在限制性内切酶位点kpni和bamhi间插入所述的水稻水稻myb类转录因子osmyb110开放阅读框,在psti和hindiii酶切位点插入35s终止子所得。
所述的水稻水稻myb类转录因子osmyb110的过表达载体在降低植物株高、增加分蘖、缩短花期和/或增加稻米品质方面的应用。
本发明的有益效果:
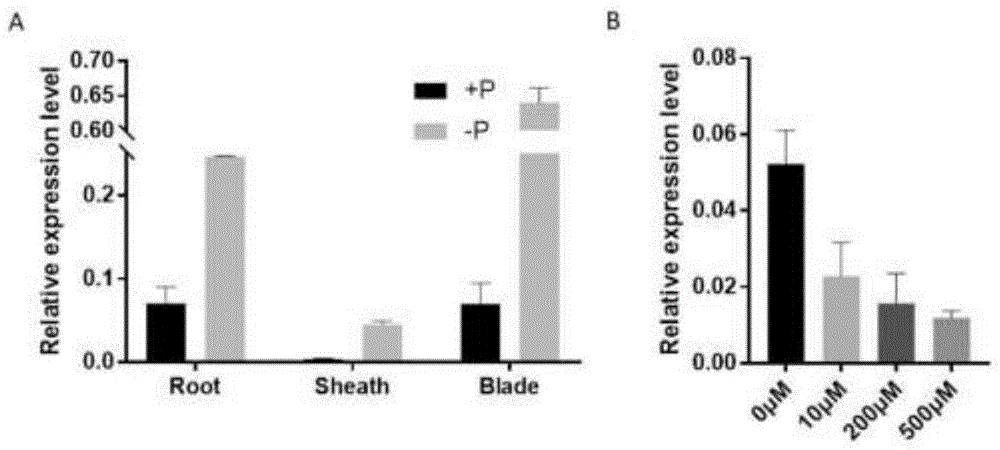
1、本发明利用特异引物研究了osmyb110在水稻中的表达,发现osmyb110在缺磷时表达上调,且在根、叶片、叶鞘均有表达(图1)。
2、本发明构建的osmyb110过量表达植株的株高显著降低、分蘖显著增多(图2),有利于改善水稻株型,提高其抗倒伏性继而增产。
3、osmyb110过量表达植株的花期提前(图3),有利于水稻提前成熟,有利于作物的轮作(譬如夏天种水稻,紧接着冬天种油菜或小麦,水稻提前收获,则可以早种油菜或者小麦,或者反过来,水稻可以迟一点播种或移栽,赶在深秋降温或者小麦油菜播种移栽之前收获)和作物之间的间作(譬如旱作水稻与西瓜,旱作水稻与豆科作物,玉米与豆科作物等等,缩短水稻生育前有可能使得间作的作物同时收获,或者提高光能、空间利用效率等),提高光能、空间利用效率等。
4、osmyb110过量表达植株的籽粒中各项品质数据提高(表1),有利于提高籽粒的口感和营养。
本发明从水稻中鉴定了一个表达受供磷水平影响的myb转录因子osmyb110,提出了利用osmyb110进行水稻株型改良、花期提前以实现高产的方法,并可利用osmyb110进行水稻稻米品质改良的方法。
本专利发明亮点:①该osmyb110转录因子表达受磷素水平调控且其超表达转基因植株呈现株高降低、分蘖增多、提前开花的现象。对水稻产量及作物的轮作、间作有改善作用。该项发明已有专利中未发现。②超表达材料的籽粒口感品质增加,在已有专利中暂未发现相关报道。
附图说明
图1rt-qpcr分析osmyb110在5叶苗期的根、叶鞘及叶片中的表达;在缺磷条件下的表达
图2生育后期osmyb110过量表达植株的表型,其株高与野生型植株相比显著降低,其分蘖与野生型植株相比显著增高
图3生育后期osmyb110过量表达植株的开花时间表型,其开花时间与野生型植株相比显著提前。图为同一日期拍摄的野生型与osmyb110过量表达植株照片,图中野生型植株在该日期开始抽穗、扬花,超表达植株已扬花完成进行灌浆。
具体实施方式
实施例1水稻myb转录因子基因osmyb110的分子克隆
1)总rna提取和cdna的合成
选用的水稻为粳稻,品种为日本晴(nipponbare,水稻基因组测序品种)。将水稻籽粒去除颖壳后于10%h2o2中进行表面消毒30min,再用去离子水洗三次后置于底部有孔洞的塑料筐中,并将塑料筐架于桶沿上,加水至没过种子一半后于37℃黑暗条件下浸种。待种子露白(1-2天)继续用水培养,当水稻幼苗长至两叶一心时改用1/2国际水稻所营养液培养。待水稻幼苗长至四叶一心后,对水稻地上部和根部进行混合采样,加液氮后用研钵裂解组织和细胞,转入2ml离心管中用trizolreagent(invitrogen,usa)抽提总rna。用琼脂糖凝胶电泳和nanodrop仪器检测所提取总rna的完整性和浓度,继而用hiscriptiiqselectrtsupermixforqpcr试剂盒(vazyme,nanjing,china)进行cdna的合成。
2)osmyb110cdna的扩增
以osmyb110的基因号os10g0478300在ncbi数据库获得其cdna序列,并根据该序列设计扩增osmyb110cdna的一对引物,myb110-cdna-f:aaaaagaagaaggaagaaaaaaaca(seqidno.1)和myb110-cdna-r:tgcatctaattctgaagccag(seqidno.2)。pcr反应程序为:94℃预变性5分钟;94℃变性30s,55℃复性30s,72℃延伸82s,30个循环后72℃7min。最终将pcr产物进行克隆、测序获得了水稻osmyb110的序列。测序结果与ncbi数据库中的序列完全一致,表明osmyb110确为一个编码myb家族转录因子的基因。
实施例2osmyb110的序列信息与特性分析
本发明osmyb110的cdna全长为1360bp,其开放阅读框位于286-1116bp处,编码277个氨基酸,其启动子区域含有磷响应启动子元件(p1bs元件、w-box等)。osmyb110含有2个myb结构,属于r2r3-myb转录因子。
实施例3osmyb110的表达研究
根据实例1中获得的osmyb110cdna序列,在其3’非翻译区(3’utr)设计特异引物进行定量rt-pcr(rt-qpcr)分析,引物序列如下:myb110-q-f:ctcttctctgttgtttttttttaccttct(seqidno.3)和myb110-q-r:cctgaatttcccatcgttttct(seqidno.4)。所使用的内参基因为水稻actin基因osactin1(mcelroyetal.,1990),其扩增引物:act-q-f:caacacccctgctatgtacg(seqidno.5)和act-q-r:catcaccagagtccaacacaa(seqidno.6)。结果表明,osmyb110的表达丰度在5叶期幼苗中根和叶片中表达相当,在叶鞘中表达含量稍低;osmyb110在缺磷条件下表达增加,并随着磷浓度的降低表达量增加(图1)。
实施例4osmyb110过量表达水稻植株的获得
委托生物公司合成2x35scamv启动子序列(seqidno.9)和35s终止子序列(seqidno.10),在2x35scamv启动子序列上下游分别引入ecori和saci酶切位点,在35s终止子序列上下游分别引入psti和hindiii酶切位点。通过两对酶切位点将获得的启动子和终止子序列连接到表达载体pcambia1305,获得最终的过量表达载体pcambia1305-35st。根据实例1得到的osmyb110的cdna全长序列,设计扩增出完整开放阅读框的引物,并在上游和下游引物上分别引入限制性内切酶位点kpni和bamhi,上游引物为myb110-orf-f:atggggagggcgccgtgctg(seqidno.7),下游引物为myb110-orf-r:ttacaagacggccaaatccc(seqidno.8),以实施例1中的cdna模板进行pcr,继而通过这两个酶切位点将osmyb110的开放阅读框连接到pcambia1305-35st载体中,由此获得2x35scamvpromoter::osmyb110::35sterminator表达盒。然后通过根癌农杆菌介导的水稻转基因方法将其转入水稻品种武运粳。待获得的转基因植株进行一系列分子鉴定后对其进行生长发育指标的评价。结果表明,osmyb110过量表达植株的株高与野生型植株相比显著降低,分蘖数与野生型植株相比显著增加,花期较野生型植株相比提前,而其籽粒中rva的各项指标与野生型相比差异较大(图2-图3,表1)。上述实施例表明本发明中克隆的myb转录因子基因osmyb110在水稻中可改变地上部株型及籽粒中直链淀粉含量的变化。
表1生育后期osmyb110过量表达植株的籽粒rva测定数据表
注:1.糊化温度(pastingtemperature),即rva仪测量曲线开始上升时的温度;2.峰值粘度(peakviscosity),即rna仪测量曲线中出现峰值的粘度大小:3.热浆粘度(holdingviscosity),即峰谷粘度的大小:4.最终粘度(finalviscosity),即rva仪结束后的粘度大小:5.崩解值(breakdown),即峰值粘度减去热浆粘度得到的数值:6.消减值(setback),即最终粘度减去峰值粘度得到的数值:7.回复值(consistence),即最终粘度减去热浆粘度得到的数值
本实验材料中osmyb110超表达材料与野生型wt材料的米相比,崩解值(bdv)显著增高,消减值(sbv)显著降低且为负数。据己有研究可知,味道较好的米饭,崩解值大多在100rvu以上,消减值在25rvu以下,且多数为负值:反之,味道较差的米饭,崩解值低于36rvu,消减值高于80rva。消减值小的米仮一般比较软糯粘口,消减值太大的米饭则又硬又糙。由此可见,在野生型材料中超表达osmyb110之后在原有品质的基础上又增加了大米的口感。
本发明人从单子叶植物水稻(oryzasativa)中克隆了一个cdna,其编码r2r3型myb转录因子osmyb110。rt-qpcr分析表明其在水稻根部、叶片、叶鞘中均有表达,并受缺磷诱导表达。转基因研究表明,将osmyb110基因转入水稻,转基因植株在正常条件下的长势与对照相比其株高降低、分蘖数增多、花期提前。该基因可作为目的基因导入植株,改变植株株型,提高植株抗倒伏能力及光合作用能力,以进行植物品种改良。在本发明的方法中,可利用osmyb110基因作为目的基因构建植物表达载体,其中可用任何一种启动子例如花椰菜花叶病毒(camv)35s启动子、ubiquitin启动子或其它启动子,该表达载体中必要时可包括增强子,不论是转录增强子或翻译增强子。
序列表
<110>南京农业大学
<120>一种水稻myb转录因子及其重组表达载体的应用
<160>10
<170>siposequencelisting1.0
<210>1
<211>25
<212>dna
<213>人工序列(artificialsequence)
<400>1
aaaaagaagaaggaagaaaaaaaca25
<210>2
<211>21
<212>dna
<213>人工序列(artificialsequence)
<400>2
tgcatctaattctgaagccag21
<210>3
<211>29
<212>dna
<213>人工序列(artificialsequence)
<400>3
ctcttctctgttgtttttttttaccttct29
<210>4
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>4
cctgaatttcccatcgttttct22
<210>5
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>5
caacacccctgctatgtacg20
<210>6
<211>21
<212>dna
<213>人工序列(artificialsequence)
<400>6
catcaccagagtccaacacaa21
<210>7
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>7
atggggagggcgccgtgctg20
<210>8
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>8
ttacaagacggccaaatccc20
<210>9
<211>892
<212>dna
<213>人工序列(artificialsequence)
<400>9
accggtcaacatgtggagcacgacacacttgtctactccaaaaatatcaaagatacagtc60
tcagaagaccaaagggcaattgagacttttcaacaaagggtaatatccggaaacctcctc120
ggattccattgcccagctatctgtcactttattgtgaagatagtggaaaaggaaggtggc180
tcctacaaatgccatcattgcgataaaggaaaggccatcgttgaagatgcctctgccgac240
agtggtcccaaagatggacccccacccacgaggagcatcgtggaaaaagaagacgttcca300
accacgtcttcaaagcaagtggattgatgtgataacatggtggagcacgacacacttgtc360
tactccaaaaatatcaaagatacagtctcagaagaccaaagggcaattgagacttttcaa420
caaagggtaatatccggaaacctcctcggattccattgcccagctatctgtcactttatt480
gtgaagatagtggaaaaggaaggtggctcctacaaatgccatcattgcgataaaggaaag540
gccatcgttgaagatgcctctgccgacagtggtcccaaagatggacccccacccacgagg600
agcatcgtggaaaaagaagacgttccaaccacgtcttcaaagcaagtggattgatgtgat660
atctccactgacgtaagggatgacgcacaatcccactatccttcgcaagacccttcctct720
atataaggaagttcatttcatttggagaggacgtcgagagttctcaacacaacatataca780
aaacaaacgaatctcaagcaatcaagcattctacttctattgcagcaatttaaatcattt840
cttttaaagcaaaagcaattttctgaaaattttcaccatttacgaacgatag892
<210>10
<211>219
<212>dna
<213>人工序列(artificialsequence)
<400>10
gtccgcaaaaatcaccagtctctctctacaaatctatctctctctatttttctccagaat60
aatgtgtgagtagttcccagataagggaattagggttcttatagggtttcgctcatgtgt120
tgagcatataagaaacccttagtatgtatttgtatttgtaaaatacttctatcaataaaa180
tttctaattcctaaaaccaaaatccagtgacgcggccgc219
- 还没有人留言评论。精彩留言会获得点赞!