靶核酸序列上的核苷酸变异的检测的制作方法与工艺
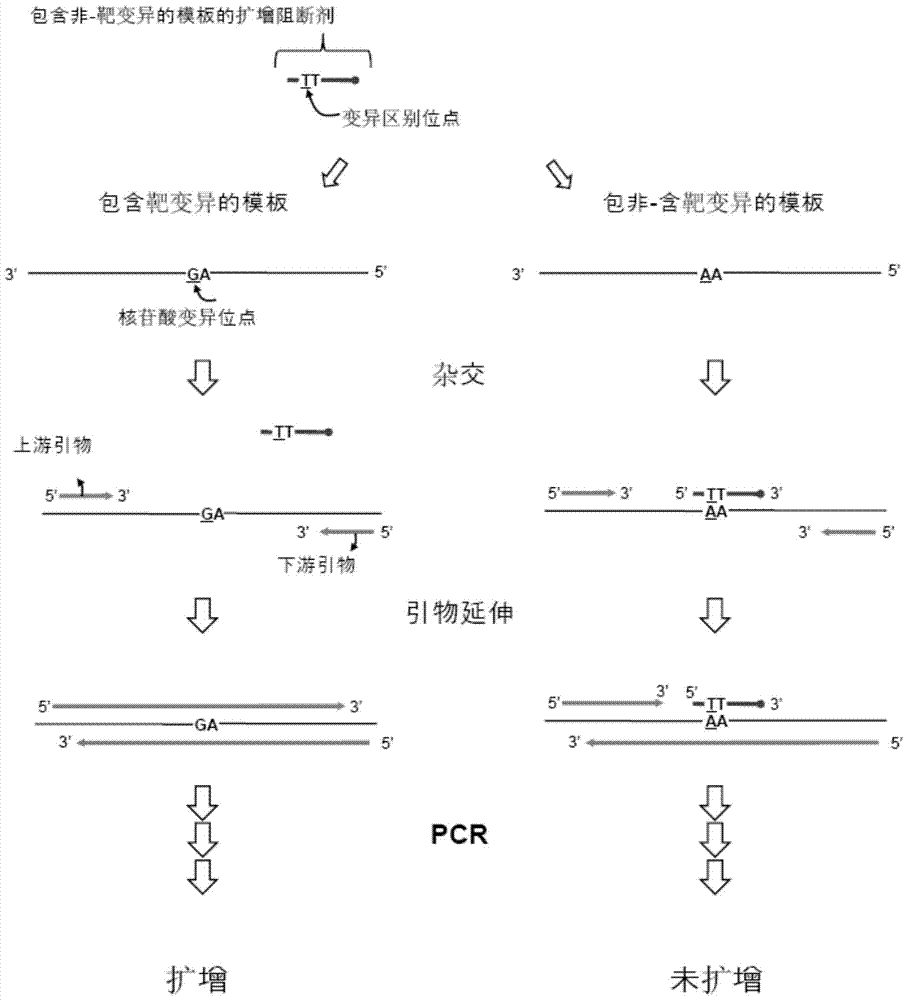
靶核酸序列上的核苷酸变异的检测【相关申请】本申请基于申请号为第61/827966、申请日为2013年5月28日的美国临时专利申请提出,并要求该专利申请的优先权,上述专利申请的公开事项在此引入本申请作为参考。【技术领域】本发明涉及利用扩增阻断剂及基于探测和标记寡核苷酸切割及延伸的变异检测(VD-PTOCE,VariationDetectionbyPTOCleavageandExtension)分析的靶核酸序列上的核苷酸变异的检测。
背景技术:
脱氧核糖核酸(DNA)杂交(hybridization)是分子生物学的基本的过程,受离子强度、碱基结构、减少后的核酸片段的长度、错配程度以及变性剂的存在的影响。基于脱氧核糖核酸杂交的技术将成为决定特定核酸序列的非常有用的用具,将会明确有助于临床诊断、基因研究及法医学上的实验分析。但是,在仅依赖杂交的现有的方法以及过程中,发生因探针和非-靶序列之间的非-特异性杂交引起的假阳性结果的可能性高。因此,现有的方法以及过程存在改善可靠性的问题。除探针杂交过程以外还提出了利用酶反应的几种接近法,例如,TaqManTM探针方法。在TaqManTM探针方法中,与靶核酸序列杂交的标记探针基于上游引物-依赖性脱氧核糖核酸聚合酶的5’核酸酶活性被切割,来产生表示靶序列存在的信号(美国专利第5210015号、第5538848号以及第6326145号)。TaqManTM探针方法提出用于信号产生的两个接近法:聚合-依赖性切割(polymerization-dependentcleavage)以及聚合-独立性切割(polymerization-independentcleavage)。在聚合-依赖性切割中,上游引物的延伸必须在核酸聚合酶与标记探针的5’-末端接触之前发生。随着延伸反应的进行,聚合酶逐渐地切割标记探针的5’-末端。在聚合-独立切割中,上游引物以及标记探针非常邻近,由此与靶核酸序列杂交,上游引物的3’-末端和核酸聚合酶的结合使上述核酸聚合酶与标记探针的5’-末端接触来放出标记。并且,TaqManTM探针方法公开,通过在其5’-末端部位具有不与靶序列杂交的5’-尾巴部位的标记探针被切割来形成包含5’-尾巴部位的片段。也有对具有对靶序列非-互补的5’-尾巴部位的探针被5’核酸酶切割,放出包含5’-尾巴部位的片段的几种方法的报告。例如,美国专利第5691142号就公开了对基于脱氧核糖核酸聚合酶的5’核酸酶活性来被切割的切割结构。公开中例示了有对包含对模板(template)非-互补的5’部位以及对模板互补的3’部位的寡核苷酸与模板杂交,并且,上游寡核苷酸与模板非常邻近而由此与模板杂交的切割结构。上述切割结构通过被具有5’核酸酶活性的脱氧核糖核酸聚合酶或具有减少的合成活性的变形脱氧核糖核酸聚合酶切割,来放出与模板非-互补的5’部位。之后,被放出的5’部位与具有发夹结构的寡核苷酸杂交,形成切割结构。由此,诱导渐进性的切割反应来检测靶序列。美国专利第7381532号公开了具有3’-末端被阻断的上游寡核苷酸的切割结构通过被具有5’核酸酶活性的脱氧核糖核酸聚合酶或瓣状核酸内切酶切割,来放出上述非-互补的5’皮瓣部位,被放出的上述5’皮瓣部位基于大小分析或相互作用性双重标记来被检测的过程。美国专利第6893819号公开了能够检测的被放出的皮瓣基于核酸合成依赖性的、皮瓣-媒介连续性扩增方法而生成的内容。在此方法中,从第一个切割结构放出的皮瓣,以核酸合成依赖性方式切割第二切割结构来从第二切割结构放出皮瓣,并检测被放出的上述皮瓣。美国专利第7309573号公开了包括基于核酸合成生成的被放出的皮瓣的生成、被放出的皮瓣的延伸、在皮瓣延伸期间的寡核苷酸的切割及切割寡核苷酸而生成的信号检测的方法。借助液相中的荧光-标记探针的杂交,即使利用一种荧光标记,也能够基于解链曲线分析来同时检测多个靶核酸序列。但是,在基于相互作用性-双重标记探针的5’核酸酶介导的切割来检测靶序列的现有技术中,在检测多重靶时,对相互不同的靶序列需要相互不同的种类的荧光标记,由于这种荧光标记的种类数量有限,因而检测出的靶序列的数量受限。美国申请公开号第2008-0241838号中公开了利用在靶核酸序列具有非-互补的5’部位的探针的切割以及捕捉(capture)探针的杂交的靶检测方法。标记位于非-互补的5’部位。杂交于靶序列的标记探针被切割而放出片段,之后,片段与捕捉探针杂交而检测靶序列的存在。在此方法中,非切割的/完好无损的(uncleaved/intact)探针不与捕捉探针杂交是必须的。为此,需要长度短的捕捉探针在固相底物实现固定化。但是,这种限制会降低固相底物上的杂交效率,并且使反应条件的最优化变得困难。因此,对开发以更加便利、具有可靠性以及再现性的方式,不仅基于杂交,还基于5’核酸切割反应(5’nucleolyticreaction)等的酶反应,在液相以及固相中检测靶序列,更优选地检测多个靶序列的新颖的接近法的要求正在抬头。并且,在本发明所属领域中正需求一种不受限于标记(尤其,荧光标记)种类的数量的新颖的靶检测方法。另一方面,核苷酸变异在研究及临床领域很重要。其中,单核苷酸多态性(SNP,singlenucleotidepolymorphism)在人类基因组中最常发现,并起到对于疾病相关基因座(loci)及药物遗传学(pharmacogenetics)的标记作用(Landegren等,1998;Roses,2000)。单核苷酸多态性在人类基因组中以每1000bp约一个的比率被发现,总数推定为约300万个。据报告,有用于检测如单核苷酸多态性、缺失、插入及移位(translocation)之类的核苷酸变异的各种等位基因区别技术。等位基因-特异性TaqMan探针设计成仅在聚合酶链式反应(PCR)的延伸步骤中与靶序列完美匹配的情况下杂交。TaqMan探针具有报道分子及可从报道分子对荧光信号进行猝灭的猝灭分子。与靶序列杂交的TaqMan探针借助TaqDNA聚合酶的5’核酸酶活性而被切割,报道分子及猝灭分子通过分离来生成靶信号。为了区别等位基因,使用与小沟结合物(MGB,minorgroovebinder)相结合的13-20mer的探针(Livak等,etal.,Genet.Anal.14:143-149(1999))。利用TaqMan探针的等位基因区别方法不仅利用杂交反应,而且利用5’核酸酶活性的酶反应,使得特异性增加。但是,上述方法存在如难以设计等位基因-特异性探针、难以将可区别基于一个错配的差异的反应条件最优化之类的严重的问题。并且,与小沟结合物的结合为对等位基因-特异性TaqMan探针的问题的解决方法之一。已公开有用于利用肽核酸(PNA)或锁核酸(LNA)夹子(clamp)并通过突变等位基因的选择性的扩增来检测少数突变群体(minoritymutantpopulation)的聚合酶链式反应箝术(clamping)。利用肽核酸的代表性的聚合酶链式反应箝术已公开于文献[Henrik等,NucleicAcidResearch21:5332-5336(1993)及Luo等,NucleicAcidResearchVol.34,No2e12(2006)]。但上述聚合酶链式反应箝术有可能无法完全阻断(blocking)野生型等位基因的扩增。因此,要求开发一种克服现有技术的缺点,并且能够以更加便利、具有可靠性、再现性的方式检测核苷酸变异的新颖的接近法。本说明书全文中,参照了多个专利及文献,并用括号来表示了其引用。这种专利及文献的公开内容全部包括在本说明书作为参照,从而更加明确说明本发明所属的技术领域的水平及本发明的内容。【发明概述】本发明人为了开发具有更加得到改善的准确性以及便利性,尤其以多重方式检测靶序列的新颖的接近法而锐意研究努力。结果,本发明人定立了用于改善由本发明人研发的基于探测和标记寡核苷酸切割及延伸的变异检测分析(参照PCT/KR2013/001492)来检测少量的靶核酸上的靶核苷酸变异的方案。本发明的方案不仅优秀地适用于固相反应,而且也能够优秀地适用于液相反应,能够以具有更加得到改善的准确性以及便利性的方式检测多个靶序列。因此,本发明的目的在于,提供利用扩增阻断剂及基于探测和标记寡核苷酸切割及延伸的变异检测分析来检测靶核酸序列上的靶核苷酸变异的方法。本发明的另一目的在于,提供用于利用扩增阻断剂及基于探测和标记寡核苷酸切割及延伸的变异检测分析来检测靶核酸序列上的靶核苷酸变异的试剂盒。通过所附的发明要求保护范围、附图及以下的详细说明,本发明的其他目的及优点将变得更加明确。【附图说明】图1示出利用于探测和标记寡核苷酸切割及延伸分析(PTOcleavageandextensionassay,PTOCEassay)的探测和标记寡核苷酸(ProbingandTaggingOligonucleotide)及捕捉和模板化寡核苷酸(CapturingandTemplatingOligonucleotide)的图示性的结构。优选地,探测和标记寡核苷酸及捕捉和模板化寡核苷酸的3’-末端被阻断,以防止上述探测和标记寡核苷酸及捕捉和模板化寡核苷酸的3’-末端延伸。探测和标记寡核苷酸-核苷酸变异作为探测和标记寡核苷酸的变形,包含对靶核酸上的靶核苷酸变异互补的序列,还包含位于3’-靶向部位的5’-末端一部分的核苷酸变异区别位点。图2以图示性的方式示出使用扩增阻断剂来选择性地扩增包含靶核苷酸变异的靶核酸序列(即,包含靶变异的模板)。图3以图示性的方式示出基于本发明的扩增阻断剂-参与变异检测的靶核苷酸变异的选择性的检测。图4以图示性的方式示出用于检测靶核酸序列上的靶核苷酸变异的扩增阻断剂及探测和标记寡核苷酸-核苷酸变异的协同工作流程(co-workingprocess)。上述扩增阻断剂及探测和标记寡核苷酸-核苷酸变异被设计成位于靶核酸序列的相同链。图5以图示性的方式示出基于使用被相互作用性双重标记标记的捕捉和模板化寡核苷酸的本发明的扩增阻断剂-参与变异检测的靶核苷酸变异的选择性的检测。图6以图示性的方式示出基于使用信号寡核苷酸的本发明的扩增阻断剂-参与变异检测的靶核苷酸变异的选择性的检测。图7以图示性的方式示出基于使用杂交寡核苷酸的本发明的扩增阻断剂-参与变异检测的靶核苷酸变异的选择性的检测。图8A及图8B示出基于本发明的扩增阻断剂-参与变异检测的少数检测界限(minoritydetectionlimit)的改善结果。图9A及图9B示出基于使用信号寡核苷酸的本发明的扩增阻断剂-参与变异检测的少数检测界限的改善结果。图10A及图10B示出基于使用杂交寡核苷酸的本发明的扩增阻断剂-参与变异检测的少数检测界限的改善结果。【发明详述】在本发明的一实施方式中,本发明提供利用扩增阻断剂及基于探测和标记寡核苷酸切割及延伸的变异检测分析来检测靶核酸序列上的靶核苷酸变异的方法,其特征在于,包括:步骤(a),使上述靶核酸序列与包含用于使上述靶核酸扩增的上游引物及下游引物的引物对、对5’核酸酶切割具有耐性的上述扩增阻断剂及探测和标记寡核苷酸-核苷酸变异(PTO-NV,ProbingandTaggingOligonucleotideforNucleotideVariation)杂交,上述上游引物及下游引物分别包含与上述靶核酸序列互补的杂交核苷酸序列,上述扩增阻断剂包含与非-靶核苷酸变异互补的序列,上述非-靶核苷酸变异不同于上述靶核酸序列上的上述靶核苷酸变异,上述探测和标记寡核苷酸-核苷酸变异包含:(i)3’-靶向部位,包含与上述靶核酸序列互补的杂交核苷酸序列,(ii)5’-标记部位,包含与上述靶核酸序列非-互补的核苷酸序列,以及(iii)核苷酸变异区别位点,包含与上述靶核酸上的核苷酸变异互补的序列,并位于上述3’-靶向部位的5’-末端一部分的核苷酸变异区别位点(nucleotidevariationdiscriminationsite),上述扩增阻断剂与具有上述非-靶核苷酸变异的靶核酸序列杂交,不与具有上述靶核苷酸变异的靶核酸序列杂交,上述探测和标记寡核苷酸-核苷酸变异的3’-靶向部位与上述靶核酸序列杂交,上述探测和标记寡核苷酸-核苷酸变异的5’-标记部位不与上述靶核酸序列杂交,上述上游引物位于上述探测和标记寡核苷酸-核苷酸变异的上游,上述扩增阻断剂位于上述上游引物或下游引物的下游,上述扩增阻断剂及上述探测和标记寡核苷酸-核苷酸变异位于上述上游引物和下游引物之间;步骤(b),在用于切割上述探测和标记寡核苷酸-核苷酸变异的条件下,使上述步骤(a)的产物与具有5’核酸酶活性的酶相接触,上述上游引物通过其延伸链来诱导基于上述具有5’核酸酶活性的酶的上述探测和标记寡核苷酸-核苷酸变异的切割,上述扩增阻断剂和具有上述非-靶核苷酸变异的靶核酸序列的杂交抑制位于上述扩增阻断剂的上游的上述引物的延伸,由此,阻断具有上述非-靶核苷酸变异的靶核酸序列的扩增;若上述探测和标记寡核苷酸-核苷酸变异与具有对上述核苷酸变异区别位点互补的上述靶核苷酸变异的上述靶核酸序列杂交,则上述3’-靶向部位的5’-末端一部分与上述靶核酸序列形成双链,来从第一初期切割位点诱导切割,并放出第一片段,若上述探测和标记寡核苷酸-核苷酸变异与具有对上述核苷酸变异区别位点非-互补的上述非-靶核苷酸变异的上述靶核酸序列杂交,则上述3’-靶向部位的5’-末端一部分不与上述靶核酸序列形成双链,而从位于上述第一初期位点的下游的第二初期切割位点诱导切割,并放出第二片段,上述第二片段包含追加的3’-末端部位,用于使上述第二片段与上述第一片段不同;步骤(c),使从上述探测和标记寡核苷酸-核苷酸变异放出的上述片段与捕捉和模板化寡核苷酸(CTO,CapturingandTemplatingOligonucleotide)杂交,上述捕捉和模板化寡核苷酸沿着3’→5’方向包含:(i)捕捉部位,包含与上述探测和标记寡核苷酸-核苷酸变异的5’-标记部位或5’-标记部位的一部分互补的核苷酸序列,以及(ii)模板化部位,包含与上述探测和标记寡核苷酸-核苷酸变异的5’-标记部位及3’-靶向部位非-互补的核苷酸序列,从上述探测和标记寡核苷酸-核苷酸变异放出的上述第一片段或上述第二片段与上述捕捉和模板化寡核苷酸的捕捉部位杂交;步骤(d),利用上述步骤(c)的产物及模板-依赖性核酸聚合酶来实施延伸反应,若上述第一片段与上述捕捉和模板化寡核苷酸的捕捉部位杂交,则上述第一片段被延伸,来生成包含与上述捕捉和模板化寡核苷酸的模板化部位互补的延伸序列的延伸链,若上述第二片段与上述捕捉和模板化寡核苷酸的捕捉部位杂交,则上述第二片段不被延伸;以及步骤(e),检测上述延伸链的存在,上述延伸链的存在表示上述靶核苷酸变异的存在。本发明人为了开发检测少量的靶核酸上的靶核苷酸变异的新的接近法而进行了不断的努力。结果,本本发明人定立了用于改善由本发明人研发的基于探测和标记寡核苷酸切割及延伸的变异检测分析(参照PCT/KR2013/001492)来检测少量的靶核酸上的靶核苷酸变异的方案。上述基于探测和标记寡核苷酸切割及延伸的变异检测分析为用于检测核苷酸变异的由本发明人开发的探测和标记寡核苷酸切割及延伸(PTOCE)分析(参照WO2012/096523)的特定实例。本发明的方案不仅良好地适用于固相反应,而且也良好地适用于液相反应,而且能够以更加改善的准确性及方便性检测少量的多个核苷酸变异。本发明的目的在于,在利用探测和标记寡核苷酸-核苷酸变异(ProbingandTaggingOligonucleotideforNucleotideVariation)的基于探测和标记寡核苷酸切割及延伸的变异检测分析中适用利用扩增阻断剂的特定序列的选择性的扩增,从而有效地检测少量的靶核酸上的靶核苷酸变异。本发明利用扩增阻断剂及探测和标记寡核苷酸-核苷酸变异(ProbingandTaggingOligonucleotideforNucleotideVariation)的杂交、探测和标记寡核苷酸-核苷酸变异的切割及延伸、核苷酸变异–依赖性延伸链的生成及延伸链检测等连续步骤。因此,本发明被命名为基于探测和标记寡核苷酸(PTO)切割及延伸的扩增阻断剂-参与变异检测(AmplificationBlocker-InvolvedVariationDetectionbyPTOCleavageandExtension:AB-VDPTOCE)分析。临床样品通常在过量的野生型等位基因内包含少量的突变等位基因。过量的野生型等位基因具有在扩增过程中消耗必备试剂,且阻隔突变等位基因的信号的倾向。为了克服这种问题,揭示了一边抑制野生型等位基因的扩增,一边使突变等位基因选择性地扩增的多种方法。作为代表性的例,报告出使用包含肽核酸或锁核酸的寡核苷酸作为扩增阻断剂的方法(美国申请公开第2004/0014105号、美国专利第7803543号、美国专利第8206929号、H.Orum.,NucleicAcidsResearch21:5332-5336(1993),A.Senescau等.,JournalofClinicalMicrobiology,3304-3308(2005),Y.Nagai等.,CancerRes65:7276-7282(2005),Henrik等.,NucleicAcidResearch21:5332-5336(1993)及Luo等.,NucleicAcidResearchVol.34,No2e12(2006))。通常,上述扩增阻断剂被设计成只有在相同条件下具有与扩增阻断剂完全互补的序列的模板中杂交,哪怕只有一个错配,也无法与模板杂交。与用于抑制引物的退火或链的延伸的扩增阻断剂杂交的模板无法扩增,只有与扩增阻断剂未杂交的模板才会扩增。由于肽核酸及锁核酸之类的核酸类似物仅利用一个碱基差异也可以呈现出显著的Tm差异,因此有利于用作扩增阻断剂。在所使用的聚合酶具有核酸酶活性的情况下,扩增阻断剂需要对上述核酸酶活性具有耐性。并且,上述方法通常需要用于发生信号的追加的探针。扩增阻断剂可具有标记。在靶核酸序列上的核苷酸变异区域具有两个相区别的变异的情况下,扩增阻断剂虽然对具有受关注的变异的靶核酸序列进行扩增,但抑制具有其他变异的靶核酸序列的扩增,从而有效地检测受关注的变异。优选地,扩增阻断剂非常有利于在包含过量的野生型等位基因及少量的突变等位基因的临床样品中检测少量的突变等位基因。然而值得关注的是,野生型等位基因的扩增无法被扩增阻断剂完全防止。根据对利用两种技术,即,扩增阻断剂及探测和标记寡核苷酸-核苷酸变异的基于探测和标记寡核苷酸切割及延伸的变异检测分析进行组合的本发明,可以检测原本无法使用现有技术来检测的非常少量的突变等位基因。本发明的基于探测和标记寡核苷酸切割及延伸的变异检测分析为了对特定核苷酸变异的探测和标记寡核苷酸的选择性而使用具有位于3’-靶向部位的5’-末端一部分的核苷酸变异区别位点的探测和标记寡核苷酸-核苷酸变异(参照图1)。若上述探测和标记寡核苷酸-核苷酸变异与具有对核苷酸变异区别位点互补的核苷酸变异的靶核酸序列(即,匹配模板)杂交,则3’-靶向部位的5’-末端一部分与匹配模板形成双链。相反,若上述探测和标记寡核苷酸-核苷酸变异与具有对核苷酸变异区别位点非-互补的核苷酸变异的靶核酸序列(即,错配模板)杂交,则3’-靶向部位的5’-末端一部分不与错配模板形成双链。在本发明中,具有对探测和标记寡核苷酸-核苷酸变异的核苷酸变异区别位点互补的核苷酸变异的靶核酸序列还被表达为对探测和标记寡核苷酸-核苷酸变异的“匹配模板(matchtemplate)”。具有对探测和标记寡核苷酸的核苷酸变异区别位点非-互补的核苷酸变异的靶核酸序列还被表达为对探测和标记寡核苷酸-核苷酸变异的“错配模板(mismatchtemplate)”。只要没有不同的表述,本说明书所使用的术语“匹配模板”及“错配模板”与探测和标记寡核苷酸-核苷酸变异相关地进行定义。值得瞩目的是,受关注的核苷酸变异中的这种相区别的杂交模式在探测和标记寡核苷酸-核苷酸变异的初期切割位点中提供差异,由此,生成两种探测和标记寡核苷酸-核苷酸变异片段,并根据是否存在受关注的核苷酸变异的来发生信号差异。通过探测和标记寡核苷酸-核苷酸变异和匹配模板之间的杂交物的切割来生成第一片段。通过探测和标记寡核苷酸-核苷酸变异和错配模板之间的杂交物的切割来生成第二片段。上述第二片段在3’-末端部位包含比第一片段追加的核苷酸。可通过捕捉和模板化寡核苷酸中的延伸反应来以区别的方式检测第一片段或第二片段的生成。通常,在严格条件下,引物的3’-末端一部分和模板之间的杂交对引物的延伸非常重要。在本发明中,第一片段及第二片段分别在捕捉和模板化寡核苷酸的相同位置中杂交。如上所述,与第一片段相比,第二片段包含追加的3’-末端部位。通过调节杂交条件及第二片段的追加的3’-末端部位的对面的捕捉和模板化寡核苷酸序列,可以只使上述第一片段延伸。可通过多种方法来检测基于上述第一片段的延伸的延伸链的生成。图2及图3以图示性的方式示出本发明的一实例。图2及图3只为了成为本发明的基本的实施原理的理解而提供于本说明书。图2示出因扩增阻断剂而防止包含非-靶变异的模板扩增,并使包含靶变异的模板选择性地扩增。图3示出延伸链的选择性的形成。实际上,也可在包含靶变异的模板扩增的期间内引起延伸链的选择性的形成。如实施例3所示,在进行几次扩增循环之后,观察到可检测的信号。在图2中,上游引物及下游引物使包含靶变异的模板扩增。然而,由于包含与非-靶核苷酸变异互补的序列的扩增阻断剂,包含非-靶变异的模板无法扩增。在图3中,探测和标记寡核苷酸-核苷酸变异的核苷酸变异区别位点包含对靶核苷酸变异互补的序列。上述探测和标记寡核苷酸-核苷酸变异与包含靶变异的模板(匹配模板)杂交,在上游引物延伸的过程中,上述探测和标记寡核苷酸-核苷酸变异在第一初期切割位点被切割,来形成第一片段,由此,在捕捉和模板化寡核苷酸上形成延伸链。相反,与包含非-靶变异的模板(错配模板)杂交的探测和标记寡核苷酸-核苷酸变异在上游引物进行延伸的过程中,在第二初期切割位点被切割,由此,无法在捕捉和模板化寡核苷酸上形成延伸链。如图3所示,探测和标记寡核苷酸-核苷酸变异可以与具有分别区别的变异的两个靶核酸序列杂交。在包含非-靶变异的变异量显著大于包含受关注的靶变异的变异量的情况下,探测和标记寡核苷酸-核苷酸变异有可能被不必要地切割来消耗。扩增阻断剂-参与变异检测分析可以适用扩增阻断剂来改善基于探测和标记寡核苷酸切割及延伸的变异检测分析,从而能够以更加改善的准确性及方便性在少量中检测多个核苷酸变异。对上述扩增阻断剂-参与变异检测分析进行更加详细的说明如下。【步骤(a):引物对、扩增阻断剂、探测和标记寡核苷酸-核苷酸变异及靶核酸序列的杂交】根据本发明,首先使靶核酸序列与引物对、扩增阻断剂及探测和标记寡核苷酸-核苷酸变异(ProbingandTaggingOligonucleotideforNucleotideVariation)杂交。在本说明书中所使用的术语“靶核酸”、“靶核酸序列”或“靶序列”意味着所要检测的核酸序列,且在杂交、退火或扩增条件下与探针或引物进行退火或杂交。在本说明书中所使用的术语“引物(primer)”意味着在诱导与核酸链(模板)互补的引物延伸产物的合成的条件,即,如核苷酸和脱氧核糖核酸聚合酶之类的聚合剂的存在时,以及适合的温度与pH的条件下,可起到合成的起始点的作用的寡核苷酸。在本说明书中所使用的术语“探针(probe)”意味着与靶核酸序列实质上互补的部位或包含这些部位的单链核酸分子。优选地,探针以及引物为单链脱氧核糖核苷酸分子。在本发明中利用的探针或引物可包含天然(naturallyoccurring)脱氧单磷酸核苷(dNMP)(即,dAMP(脱氧腺苷酸)、dGMP(脱氧鸟苷酸)、dCMP(脱氧胞苷酸)及dTMP(脱氧胸苷酸))、变形的核苷酸或非-天然核苷酸。并且,探针或引物还可包含核糖核苷酸。就引物而言,应充分长,以便能够在聚合剂的存在之下引发延伸产物的合成。引物的适合的长度取决于包括例如,温度、应用领域及引物的根源(source)的多个因素。在本说明书中使用的术语“退火”或“引发”意味着在模板核酸并置(apposition)寡脱氧核苷酸或核酸,就上述并置而言,通过聚合酶对核苷酸进行聚合而在模板核酸或其一部分形成互补的核酸分子。在本说明书中使用的术语“杂交(hybridization)”意味着互补的单链核酸形成双链核酸。可在两个核酸链之间的互补性完全匹配(perfectmatch)时发生杂交,或即使存在一部分错配(mismatch)碱基也能发生杂交。杂交所需的互补性的程度可根据杂交条件而不同,尤其是可根据温度来进行调节。靶核酸序列和引物对、扩增阻断剂及探测和标记寡核苷酸-核苷酸变异之间的杂交可在通常基于最佳化步骤而决定的合适的杂交条件下实施。温度、成分的浓度、杂交及清洗次数、缓冲液成分及它们的pH及离子强度之类的条件可根据包含寡核苷酸(引物以及探测和标记寡核苷酸)的长度及糖皮质激素(GC)含量以及靶核苷酸序列的各种因子而不同。例如,在利用相对短的寡核苷酸的情况下,优选地,选择低的严格条件(stringentcondition)。用于杂交的详细条件可以在文献[JosephSambrook等,MolecularCloning,ALaboratoryManual,ColdSpringHarborLaboratoryPress,ColdSpringHarbor,N.Y.(2001);及M.L.M.Anderson,NucleicAcidHybridization,Springer-VerlagNewYorkInc.N.Y.(1999)]中确认。术语“退火”和“杂交”没有区别,在本说明书中混用。上游引物及下游引物分别包含与靶核酸序列互补的杂交核苷酸序列。探测和标记寡核苷酸-核苷酸变异的3’-靶向部位包含与靶核酸序列互补的杂交核苷酸序列。在本说明书中使用的术语“互补的”意味着在预定的退火条件或严格条件下,引物或探针以选择性地与靶核酸序列杂交的程度充分地互补,并意味着全部包括术语“实质上互补的(substantiallycomplementary)”及“完全互补的(perfectlycomplementary)”,优选地,意味着完全互补的意思。探测和标记寡核苷酸的5’-标记部位具有与靶核酸序列非-互补的核苷酸序列。捕捉和模板化寡核苷酸(CTO,CapturingandTemplatingOligonucleotide)的模板化部位具有与探测和标记寡核苷酸的5’-标记部位以及3’-靶向部位非-互补的核苷酸序列。在本说明书中使用的术语“非-互补的”意味着在预定的退火条件或严格条件下引物或探针不以选择性地与靶核酸序列杂交的程度充分地非-互补,并意味着全部包括术语“实质上非互补的(substantiallynon-complementary)”及“完全非-互补的(perfectlynon-complementary)”,优选地,意味着完全非-互补的意思。例如,与探测和标记寡核苷酸-核苷酸变异的5’-标记部位相关而所使用的术语“非-互补的”意味着在预定的退火条件或严格条件下,上述5’-标记部位不以选择性地与靶核酸序列杂交的程度充分地非-互补,并意味着全部包括术语“实质上非互补的(substantiallynon-complementary)”及“完全非-互补的(perfectlynon-complementary)”,优选地,意味着完全非-互补的意思。扩增阻断剂包含与不同于靶核酸序列上的靶核苷酸变异的其他非-靶核苷酸变异互补的序列。上述扩增阻断剂与具有非-靶核苷酸变异的靶核酸序列杂交,且不与具有靶核苷酸变异的靶核酸序列杂交,由此,扩增阻断剂使具有靶核苷酸变异的靶核酸序列选择性地扩增。上述扩增阻断剂位于上游引物或下游引物的下游,上述扩增阻断剂及探测和标记寡核苷酸-核苷酸变异位于上游引物和下游引物之间。与存在于靶核酸序列上的变异-发生部位(variation-occurringregion)的核苷酸变异相关地,本说明书所使用的术语“靶核苷酸变异(targetnucleotidevariation)”意味着通过本发明来确认的核苷酸变异。在本发明的方法中,探测和标记寡核苷酸-核苷酸变异的核苷酸变异区别位点包含对靶核酸上的靶核苷酸变异互补的序列。本说明书所使用的术语“包含靶核苷酸变异的模板(targetnucleotidevariationcontainingtemplate)”或“包含靶变异的模板(targetvariationcontainingtemplate)”意味着包含通过本发明来确认的核苷酸变异的靶核酸分子。与存在于靶核酸序列上的变异-发生部位的核苷酸变异相关地,本说明书所使用的术语“非-靶核苷酸变异(non-targetnucleotidevariation)”意味着不同于靶核苷酸变异的核苷酸变异。本说明书所使用的术语“包含非-靶核苷酸变异的模板(non-targetnucleotidevariationcontainingtemplate)”或“包含非-靶变异的模板(non-targetvariationcontainingtemplate)”意味着包含不同于靶核苷酸变异的核苷酸变异的靶核酸分子。例如,在通过本发明的方法来在过量的野生型等位基因中检测少量的突变等位基因的过程中,上述野生型等位基因为具有非-靶核苷酸变异的靶核酸,上述突变等位基因为具有靶核苷酸变异的靶核酸。术语“核苷酸变异(nucleotidevariation)”包含在核酸序列的特定位置中的野生型及所有突变型。在本发明的方法中,扩增阻断剂包含对非-靶核苷酸变异互补的序列。存在于变异-发生部位的上述非-靶核苷酸变异可以是一种或一种以上。在这种情况下,上述扩增阻断剂包含对非-靶核苷酸变异互补的序列,用于抑制上述非-靶核苷酸变异的扩增。为了抑制多个非-靶核苷酸变异的扩增,可以使用多个扩增阻断剂。本说明书的术语“靶核苷酸变异”及“非-靶核苷酸变异”为了明确和简洁地表示与探测和标记寡核苷酸-核苷酸变异及扩增阻断剂杂交的核酸分子而使用。包含对用于抑制扩增的非-靶核苷酸变异互补的序列的扩增阻断剂与具有非-靶核苷酸变异的靶核酸序列杂交,来抑制位于扩增阻断剂的上游的引物的延伸,由此阻断靶核酸序列的扩增。在相同条件下,包含对非-靶核苷酸变异互补的序列的扩增阻断剂因错配序列的存在而无法在具有靶核苷酸变异的靶核酸序列中杂交,由此,靶核酸序列的扩增不会被阻断。根据一实例,在包含对野生型脱氧核糖核酸互补的序列的扩增阻断剂与野生型脱氧核糖核酸杂交,来抑制引物的延伸,由此抑制野生型脱氧核糖核酸的扩增。包含与野生型脱氧核糖核酸互补的序列的扩增阻断剂不会与突变脱氧核糖核酸杂交,而突变脱氧核糖核酸扩增。由于本发明利用5’核酸酶活性,因此,上述扩增阻断剂需要为了防止扩增阻断剂的切割而在5’核酸酶活性中具有耐性。在特定实例中,被5’核酸酶活性攻击的扩增阻断剂的至少一个位点被设计成在5’核酸酶活性中具有耐性。在特定实例中,上述扩增阻断剂对5’核酸酶切割具有耐性。在特定实例中,扩增阻断剂为可以与核酸序列杂交的寡核苷酸。在一实例中,上述扩增阻断剂包含天然(natural)核苷/核苷酸、核苷/核苷酸类似物或它们的组合。在一实例中,上述扩增阻断剂为具有小沟结合物之类的对5’核酸酶具有耐性的化合物(compound)的寡核苷酸。根据一实例,上述扩增阻断剂包含具有对5’核酸酶活性具有耐性的主链的核苷/核苷酸。包含对5’核酸酶活性具有耐性的主链的核苷/核苷酸包含本发明所属技术领域的普通技术人员公知的任何核苷/核苷酸。例如,上述核苷/核苷酸包含各种硫代磷酸酯连接、膦酸酯连接、磷酰胺酯连接及2’-碳水化合物变形。根据更优选的一实例,包含对5’核酸酶具有耐性的主链的核苷酸包含硫代磷酸酯连接、烷基磷酸三酯连接、芳基磷酸三酯连接、膦酸烷基酯连接、膦酸芳基酯连接、氢膦酸酯连接、烷基磷酰胺酯连接、芳基磷酰胺酯连接、硒代磷酸酯连接、2’-O-氨基丙基变形、2’-O-烷基变形、2’-O-烯丙基变形、2’-O-丁基变形、α-异头寡脱氧核苷酸及1-(4’-硫代-β-D-呋喃核糖)变形。根据一实例,上述扩增阻断剂包含肽核酸、锁核酸、吗啉(morpholino)、甘油核酸(GNA),苏糖核酸(TNA)、桥核酸(BNA)、N3’-P5’氨基磷酸酯(NP)低聚物、连接有小沟结合物的寡核苷酸(MGB-linkedoligonucleotide)、硫代磷酸酯(PS)低聚物、C1-C4膦酸烷基酯低聚物、氨基磷酸酯、β-磷酸二酯寡核苷酸、α-磷酸二酯寡核苷酸或它们的组合。在特定实例中,上述扩增阻断剂对5’核酸酶具有耐性,还通过一个错配来呈现出显著的Tm值的变化,其代表性的例有包含肽核酸或锁核酸的扩增阻断剂。上述扩增阻断剂可具有任何长度。例如,上述扩增阻断剂可具有5-100核苷酸、5-80核苷酸、5-50核苷酸、5-40核苷酸、5-30核苷酸、10-100核苷酸、10-80核苷酸、10-50核苷酸、10-40核苷酸、10-30核苷酸、15-100核苷酸、15-80核苷酸、15-50核苷酸、15-40核苷酸、15-30核苷酸、20-100核苷酸、20-80核苷酸、20-50核苷酸、20-40核苷酸或20-30核苷酸的长度。根据一实例,扩增上述阻断剂的3'-末端被“阻断”,以防止上述扩增上述阻断剂的3'-末端的延伸。只要上述扩增阻断剂对具有非-靶核苷酸变异的靶核酸序列的扩增进行抑制,且不会抑制具有靶核苷酸变异的靶核酸序列的扩增,位于靶核酸序列上的核苷酸变异部位的对面的扩增阻断剂的核苷酸变异区别位点(即,与非-靶核苷酸变异互补的部位)就可以位于扩增阻断剂的任何位点。在特定实例中,扩增阻断剂的核苷酸变异区别位点可位于扩增阻断剂的5’-末端部位、中间部位或3’-末端部位。上述扩增阻断剂位于上游引物或下游引物的下游,并位于上述上游引物和下游引物之间。位于上述扩增阻断剂的5’-末端和扩增阻断剂的上游的引物的3’-末端之间的距离至少可以为300核苷酸、200核苷酸、100核苷酸、50核苷酸、30核苷酸、20核苷酸、10核苷酸、5核苷酸、2核苷酸或1核苷酸。在特定实例中,本发明根据不对称聚合酶链式反应(PierceKE等.,MethodsMolMed.MethodsinMolecularMedicine132:65-85(2007))来实施。过量的引物或受限的引物可位于上述扩增阻断剂的上游。优选地,过量的引物可位于上述扩增阻断剂的上游。上述探测和标记寡核苷酸-核苷酸变异及扩增阻断剂可以被设计成位于靶核酸序列的相同链或不同链。根据一实例,通过本发明来检测的核苷酸变异为置换变异、缺失变异或插入变异。根据一实例,具有通过本发明来检测的(多个)核苷酸变异靶核酸序列包含如下基因:K-ras、H-ras、N-ras、p53(TP53)、CDKN2A(p16)、PIC3K、PTEN、RB1、表皮生长因子受体基因、BRAF、BRCA1、BRCA2、STK11及VHL;NF1、FBN1、MSH2、常染色体显性遗传病相关基因(autosomaldominantdisorder-associatedgene;MLH1);CFTR、血红蛋白β基因(Hemoglobinbetagene)、HEXA、SMN1、常染色体隐性遗传病相关基因(autosomalrecessivedisorder-associatedgene;VAPB);X连锁显性遗传病相关基因(X-linkeddominantdisorder-associatedgene;PHEX);VIII因子(factorVIII)、抗肌萎缩蛋白基因(dystrophingene)、CNGA3、CNGB3、GNAT2、雄激素受体(androgenreceptor;AR)基因(X-连锁隐性遗传疾病的相关基因;X-linkedrecessivedisorder-associatedgene);Y染色体连锁疾病相关基因(Y-linkeddisorder-associatedgene;USP9Y);MT-ND1、MT-ND4、MT-ND4L、线粒体疾病相关基因(mitochondrialdisease-associatedgene;MT-ND6);与用于治疗肺癌的药物(gefitnib)相关地对表皮生长因子受体(EGFR)进行加密的表皮生长因子受体基因、与用于治疗卵巢癌的药物相关地对多药耐性相关蛋白质(MRP)进行加密的多药耐性相关蛋白质基因、与用于治疗卵巢癌的药物相关的肺耐性蛋白质(LRP)基因及cagPAI、vacA、iceA、babA、erp、spvC、spuB、cnf1、cnf2、eaeA、eagg、einv、stx1、stx2及vt2e等。本说明书所使用的术语“探测和标记寡核苷酸-核苷酸变异(ProbingandTaggingOligonucleotideforNucleotideVariation)”意味着包含:(i)起到探针作用的3’-靶向部位;(ii)5’-标记部位,其包含与靶核酸序列非-互补的核苷酸序列;以及(iii)核苷酸变异区别位点,其包含与靶核酸的核苷酸变异互补的序列,并位于3’-靶向部位的5’-末端一部分。上述5’-标记部位与靶核酸序列杂交后,核酸切割(nucleolytically)方式从探测和标记寡核苷酸-核苷酸变异放出。在上述探测和标记寡核苷酸-核苷酸变异中,5’-标记部位及3’-靶向部位必须以5’→3’方向定位。图1及图3中以图示性的方式例示了上述探测和标记寡核苷酸-核苷酸变异。上述探测和标记寡核苷酸-核苷酸变异可以视为用于检测核苷酸变异的探测和标记寡核苷酸的一适用形态,向3’-靶向部位的5’-末端一部分导入核苷酸变异区别位点来制作。上述探测和标记寡核苷酸-核苷酸变异包含核苷酸变异区别位点,上述核苷酸变异区别位点包含与核苷酸变异互补的序列,上述核苷酸变异区别位点位于3’-靶向部位的5’-末端一部分。上述在探测和标记寡核苷酸-核苷酸变异与具有对变异区别位点互补的核苷酸变异的靶核酸序列杂交的情况下,3’-靶向部位的5’-末端一部分与靶核酸序列形成双链。上述探测和标记寡核苷酸-核苷酸变异与具有对变异区别位点非-互补的核苷酸变异的靶核酸序列杂交的情况下,3’-靶向部位的5’-末端一部分不与靶核酸序列形成双链。对于受关注的核苷酸变异的区别的杂交模式提供在探测和标记寡核苷酸-核苷酸变异的切割位置的差异,由此生成两种探测和标记寡核苷酸-核苷酸变异片段,来根据受关注的核苷酸变异的存在与否产生信号差异。并且,与具有对变异区别位点非-互补的核苷酸的靶核酸序列杂交的情况下,上述探测和标记寡核苷酸-核苷酸变异的3’-靶向部位的5’-末端一部分可以被记载为形成单链(singlestrand-forming)的上述探测和标记寡核苷酸-核苷酸变异的3’-靶向部位的5’-末端一部分。位于上述探测和标记寡核苷酸-核苷酸变异的3’-靶向部位的5’-末端一部分的核苷酸变异区别位点包含与核苷酸变异互补的序列。根据一实例,上述核苷酸变异区别位点位于从探测和标记寡核苷酸-核苷酸变异的3’-靶向部位的5’-末端隔开10核苷酸,更优选为8核苷酸,尤其优选为6核苷酸,进而优选为4核苷酸、3核苷酸、2核苷酸1核苷酸或0核苷酸以内的位置。优选地,核苷酸变异区别位点位于探测和标记寡核苷酸-核苷酸变异的3’-靶向部位的5’-末端。可以考虑检测的序列、核酸酶类型及反应条件来决定核苷酸变异区别位点的位置。揭示探针的核苷酸变异区别位点或靶序列的核苷酸变异位点而使用的术语“位点(site)”不仅包含单一核苷酸,还包含多个核苷酸。优选地,在上述步骤(a)中,杂交是在3’-靶向部位与靶核酸序列杂交,而5’-标记部位不与靶核酸序列杂交的严格条件下实施。上述探测和标记寡核苷酸-核苷酸变异无需具有某种特定的长度。例如,探测和标记寡核苷酸-核苷酸变异的长度可以为15-150核苷酸、15-100核苷酸、15-80核苷酸、15-60核苷酸、15-40核苷酸、20-150核苷酸、20-100核苷酸、20-80核苷酸、20-60核苷酸、20-50核苷酸、30-150核苷酸、30-100核苷酸、30-80核苷酸、30-60核苷酸、30-50核苷酸、35-100核苷酸、35-80核苷酸、35-60核苷酸或35-50核苷酸。只要与其靶核酸序列特异性地杂交,上述探测和标记寡核苷酸-核苷酸变异的3’-靶向部位可以具有任何长度。例如,探测和标记寡核苷酸-核苷酸变异的3’-靶向部位的长度可以为10-100核苷酸、10-80核苷酸、10-50核苷酸、10-40核苷酸、10-30核苷酸、15-100核苷酸、15-80核苷酸、15-50核苷酸、15-40核苷酸、15-30核苷酸、20-100核苷酸、20-80核苷酸、20-50核苷酸、20-40核苷酸或20-30核苷酸。上述5’-标记部位只要在其片段与捕捉和模板化寡核苷酸的捕捉(capturing)部位特异性地杂交来延伸,就可以具有任何长度。例如,上述探测和标记寡核苷酸-核苷酸变异的5’-标记部位的长度可以为5-50核苷酸、5-40核苷酸、5-30核苷酸、5-20核苷酸、10-50核苷酸、10-40核苷酸、10-30核苷酸、10-20核苷酸、15-50核苷酸、15-40核苷酸、15-30核苷酸或15-20核苷酸。根据一实例,上述探测和标记寡核苷酸-核苷酸变异的3’-末端被阻断,以防止上述探测和标记寡核苷酸-核苷酸变异的3’-末端的延伸。阻断可以根据现有方法而有所不同。例如,阻断可通过在最后核苷酸的3’-羟基上追加如生物素、标记、磷酸基、烷基、非-核苷酸连接肽、硫代磷酸酯或烷烃-二醇残基之类的化学组成部分(moiety)来实施。择一性地,阻断可通过去除最后核苷酸的3’-羟基或利用如双脱氧核苷酸之类的没有3’-羟基的核苷酸来实施。择一性地,上述探测和标记寡核苷酸可以被设计成具有细销结构。上述上游引物位于上述探测和标记寡核苷酸-核苷酸变异的上游。上述上游引物通过其延伸链来诱导基于具有5’核酸酶活性的酶的探测和标记寡核苷酸-核苷酸变异的切割。在本发明的方法中,与探测和标记寡核苷酸-核苷酸变异的位置相关地决定术语“上游引物”,因此,上述上游引物位于探测和标记寡核苷酸-核苷酸变异的上游。根据一实例,上述上游引物,下游引物和/或探测和标记寡核苷酸-核苷酸变异的5’-标记部位具有由本发明人开发的双重引发寡核苷酸(dualprimingoligonucleotide;DPO)结构。具有上述双重引发寡核苷酸结构的寡核苷酸与现有引物以及探针相比表现出相当得到改善的靶特异度(参照WO2006/095981;Chun等,DualprimingoligonucleotidesystemforthemultiplexdetectionofrespiratoryvirusesandSNPgenotypingofCYP2C19gene,NucleicAcidResearch,35:6e40(2007))。根据一实例,上述探测和标记寡核苷酸-核苷酸变异的3’-靶向部位具有由本发明人开发的变形双重特异性寡核苷酸(modifieddualspecificityoligonucleotide;mDSO)结构。上述变形双重特异性寡核苷酸结构与现有的探针相比表现出相当得到改善的靶特异性(参照WO2011/028041)。上述探测和标记寡核苷酸-核苷酸变异及扩增阻断剂可以被设计成位于靶核酸序列的相同链或不同链。在图4中,探测和标记寡核苷酸-核苷酸变异及扩增阻断剂位于靶核酸序列的相同链。上述探测和标记寡核苷酸-核苷酸变异的核苷酸变异区别位点包含对靶核酸上的靶核苷酸变异互补的序列,上述扩增阻断剂包含对靶核酸上的非-靶核苷酸变异互补的序列。上述探测和标记寡核苷酸-核苷酸变异与包含靶核苷酸变异的模板杂交,上游引物延伸并切割上述探测和标记寡核苷酸-核苷酸变异,在捕捉和模板化寡核苷酸上形成延伸链。另一方面,上述扩增阻断剂与包含非-靶核苷酸变异的模板杂交,由扩增阻断剂防止上游引物的延伸。并且,包含非-靶核苷酸变异的模板上的扩增阻断剂的存在抑制探测和标记寡核苷酸-核苷酸变异的杂交,从而防止探测和标记寡核苷酸-核苷酸变异与包含非-靶核苷酸变异的模板杂交来不必要地被切割。可以调节反应条件及扩增阻断剂的序列,使得扩增阻断剂和野生型脱氧核糖核酸之间的杂交更优于探测和标记寡核苷酸-核苷酸变异和野生型脱氧核糖核酸之间的杂交。择一性地,上述探测和标记寡核苷酸-核苷酸变异及上述扩增阻断剂可以被设计成位于靶核酸序列的互不相同的链。【步骤(b):从探测和标记寡核苷酸-核苷酸变异放出片段】接着,在用于探测和标记寡核苷酸切割的条件下,使上述步骤(a)的产物与具有5’核酸酶活性的酶相接触。上游引物通过其延伸来诱导基于具有5’核酸酶活性的酶的探测和标记寡核苷酸-核苷酸变异的切割。本说明书所使用的术语“用于探测和标记寡核苷酸-核苷酸变异切割的条件”意味着对基于具有5’核酸酶活性的酶发生与靶核酸序列杂交的探测和标记寡核苷酸-核苷酸变异被切割的充分的条件,例如,温度、pH、离子强度、缓冲液、寡核苷酸的长度及序列和酶。例如,在利用作为具有5’核酸酶活性的酶的TaqDNA聚合酶的情况下,用于探测和标记寡核苷酸切割的条件包含Tris-HCl缓冲液、KCl、MgCl2及温度。具有非-靶核苷酸变异的靶核酸序列和扩增阻断剂之间的杂交抑制位于上述扩增阻断剂的上游的引物的延伸,由此阻断具有非-靶核苷酸变异的靶核酸序列的扩增。若上述探测和标记寡核苷酸-核苷酸变异与具有对变异区别位点互补的靶核苷酸变异的靶核酸序列(即,匹配模板)杂交,上述3’-靶向部位的5’-末端一部分与靶核酸序列形成双链,来诱导从第一初期切割位点的切割,则放出第一片段。若上述探测和标记寡核苷酸-核苷酸变异与具有对上述变异区别位点非-互补的核苷酸变异的靶核酸序列(即,错配模板)杂交,且3’-靶向部位的5’-末端一部分不与上述靶核酸序列形成双链,而从位于第一初期切割位置的下游的第二初期切割位置诱导切割,则放出第二片段,上述第二片段包含追加的3’-末端部位,这使第二片段与第一片段不同。在样品内不存在靶核酸序列的情况下,不发生探测和标记寡核苷酸-核苷酸变异切割。像这样,切割位点及所生成的探测和标记寡核苷酸-核苷酸变异片段的种类的差异可以根据靶核酸序列上是否存在受关注核苷酸变异来呈现不同的延伸模式,并且可以区别靶核酸序列上的核苷酸变异来检测。上述探测和标记寡核苷酸-核苷酸变异的初期切割位点受到5’核酸酶的种类、上游引物的杂交位点及切割条件的影响。通常,与上述上游引物的延伸一同,基于具有5’核酸酶活性的模板依赖性聚合酶的初期切割位点沿着5’→3’方向位于包含单链及双链的结构中的双链的起始核苷酸(即,分叉位点(bifurcationsite))或从起始核苷酸隔开1-2核苷酸的方位。借助切割反应生成包含5’-标记部位及3’-靶向部位的一部分的片段。与探测和标记寡核苷酸-核苷酸变异相关地使用的术语“第一初期切割位置(afirstinitialcleavagesite)”意味着具有对变异区别位点互补的核苷酸变异的靶核酸序列与探测和标记寡核苷酸-核苷酸变异杂交的情况下,首先被切割的探测和标记寡核苷酸-核苷酸变异的切割位置。与探测和标记寡核苷酸-核苷酸变异相关地使用的术语“第二初期切割位置(asecondinitialcleavagesite)”意味着具有对变异区别位点非-互补的核苷酸变异的靶核酸序列与探测和标记寡核苷酸杂交的情况下,首先被切割的探测和标记寡核苷酸切割位置。在本发明中使用的术语“第一片段(afirstfragment)”意味着在第一初期切割位点借助切割生成的片段。“第一片段”可更换为“第一分段(afirstsegment)”及“探测和标记寡核苷酸-核苷酸变异第一片段(aPTO-NVfirstfragment)”相互交换着使用。术语“第二片段(asecondfragment)”意味着在第二初期切割位置借助切割生成的片段。“第二片段”可更换为“第二分段(asecondsegment)”及“探测和标记寡核苷酸-核苷酸变异第二片段(aPTO-NVsecondfragment)”相互交换着使用。尤其,上述第一片段及第二片段分别包含5’-标记部位或5’-标记部位的一部分。可根据所利用的切割方法,在上述第一初期切割位置(或第二初期切割位置)的切割之后也可以连续进行上述切割。例如,与上游引物的延伸一同利用5’核酸酶切割反应的情况下,初期切割位置及与其连续的序列被切割。根据一实例,依赖于上游引物延伸的初期切割位置可沿着5’→3’方向位于双链的起始核苷酸(即,分叉位点)。作为本发明的一例,如图3所示,上述核苷酸变异区别位点位于探测和标记寡核苷酸-核苷酸变异的3’-靶向部位的5’-末端一部分的5’-末端。这种情况下,第一初期切割位置沿着5’→3’方向正好与3’-靶向部位的5’-末端一部分相邻。即,上述第一初期切割位置沿着3’方向正好与核苷酸变异区别位点相邻。通常,第二初期切割位置位于从核苷酸变异区别位点沿着3’方向隔开1核苷酸的位置。择一性地,上述核苷酸变异区别位点可从3’-靶向部位的5’-末端一部分的5’-末端隔开1核苷酸。在这种情况下,第一初期切割位置沿着5’方向正好与核苷酸变异区别位点相邻。通常,第二初期切割位置位于从核苷酸变异区别位点沿着3’方向隔开1核苷酸的位置。根据一实例,上述探测和标记寡核苷酸-核苷酸变异具有阻断剂部位,上述阻断剂部位包含对基于具有5’核酸酶活性的酶的切割具有耐性的一个阻断剂,上述阻断剂部位用于调节初期切割位点和/或连续的切割。根据一实例,上述探测和标记寡核苷酸-核苷酸变异具有阻断剂部位,上述阻断剂部位包含对基于具有5’核酸酶活性的酶的切割具有耐性的至少一个核苷酸作为阻断剂。例如,未来在上述探测和标记寡核苷酸-核苷酸变异的杂交部位(3’-靶向部位)和非-杂交部位(5’-标记部位)之间的连接位置(junctionsite)诱导切割,探测和标记寡核苷酸的3’-靶向部位的5’-末端一部分可以由阻断剂阻断。上述阻断剂部位中所包含的阻断剂的数量不受限制,优选为1-10阻断剂,更优选为2-10阻断剂,尤其优选为3-8阻断剂,最优选为3-6阻断剂。存在于探测和标记寡核苷酸的阻断剂能够以连续或不连续的方式存在,优选为连续的方式。作为阻断剂的核苷酸,即包含对5’核酸酶活性具有耐性的主链的核苷酸包含本领域中公知的任何核苷酸。例如,上述核苷酸包含各种硫代磷酸酯连接、膦酸酯连接、磷酰胺酯连接及2’-碳水化合物变形。根据本发明的更优选的一实例,包含对5’核酸酶具有耐性的主链的核苷酸包含硫代磷酸酯连接、烷基磷酸三酯连接、芳基磷酸三酯连接、膦酸烷基酯连接、膦酸芳基酯连接、氢膦酸酯连接、烷基磷酰胺酯连接、芳基磷酰胺酯连接、硒代磷酸酯连接、2’-O-氨基丙基变形、2’-O-烷基变形、2’-O-烯丙基变形、2’-O-丁基变形、α-异头寡脱氧核苷酸及1-(4’-硫代-β-D-呋喃核糖)变形。根据一实例,核苷酸包含锁核酸(LNA,lockednulceicacid)作为阻断剂。包含核苷酸变异区别位点的5’-末端一部分能够包含可与靶核酸序列杂交的序列。择一性地,一部分5’-末端的一部分可以包含非-杂交序列。上述探测和标记寡核苷酸-核苷酸变异与具有对核苷酸变异区别位点非-互补的核苷酸变异的靶核酸序列杂交的情况下,向上述5’-末端一部分导入非-杂交序列,这对上述5’-末端一部分形成单链很有用。根据一实例,上述探测和标记寡核苷酸-核苷酸变异的3’-靶向部位的5’-末端一部分包含位于从核苷酸变异区别位点隔开1-10核苷酸(更优选为1-5核苷酸)的位置以内的非-碱基对部分(non-basepairingmoiety)。上述探测和标记寡核苷酸-核苷酸变异与具有对变异区别位点非-互补的核苷酸变异的靶核酸序列杂交的情况下,上述非-碱基对部分防止3’-靶向部位的5’-末端一部分与靶核苷酸序列形成双链。根据一实例,具有对核苷酸变异区别位点互补的核苷酸变异的靶核酸序列与探测和标记寡核苷酸-核苷酸变异杂交的情况下,非-碱基对部分不抑制5’-末端一部分与靶核酸序列形成双链。根据一实例,非-碱基对部分扩大第一初期切割位置及第二初期切割位置之间的差异(differentiation)。例如,探测和标记寡核苷酸-核苷酸变异的3’-靶向部位的5’-末端一部分的杂交模式没有差异,因而变异区别位点的差异引起的匹配模板及错配模板的切割位置没有差异的情况下,非-碱基对部分的使用对杂交模式产生差异。并且,探测和标记寡核苷酸-核苷酸变异的3’-靶向部位的5’-末端一部分在变异区别位点的差异引起的匹配模板及错配模板的杂交模式存在差异的情况下,非-碱基对部分的使用能够使第二片段的3’-末端部位长于第一片段的3’-末端部位,由此在捕捉和模板化寡核苷酸完全防止第二片段的延伸。非-碱基对部分的使用可以改善扩增阻断剂-参与变异检测分析。根据一实例,非-碱基对部分(例如,人为错配核苷酸)的使用增加对于核苷酸变异的探测和标记寡核苷酸-核苷酸变异的区别可能性。根据一实例,基于具有5’核酸酶活性的酶的第一初期切割位置及第二初期切割位置之间的差异识别根据由非-碱基对部分赋予的差异得到改善。上述差异可根据基于非-碱基对部分的第一初期切割位置及第二初期切割位置之间的距离扩大。根据一实例,非-碱基对部分扩大第一初期切割位置及第二初期切割位置之间的距离。根据一实例,非-碱基对部分序列的导入能够调节第二初期切割位置。优选地,非-碱基对部分位于核苷酸变异区别位点的下游。例如,作为非-碱基对部分的错配核苷酸向沿着3’方向从核苷酸变异区别位点隔开2核苷酸的部位导入的情况下,第二初期切割位置调整为从核苷酸变异区别位点隔开2核苷酸的位置。在不使用错配核苷酸的情况下,第二初期切割位置位于从核苷酸变异区别位点隔开1核苷酸的位置。即,非-碱基对部分扩大第一初期切割位置及第二初期切割位置之间的距离。非-碱基对部分包含在靶核酸序列之间不形成碱基对的任何部分。优选地,非-碱基对部分为:(i)包含人为错配碱基、变形为无法碱基配对的非-碱基对碱基或通用碱基的核苷酸;(ii)变形为无法碱基配对的非-碱基对核苷酸;或(iii)非-碱基配对化合物(non-basepairingchemicalcompound)。例如,非-碱基对部分包含亚烷基(alkylenegroup)、呋喃核糖萘(ribofuranosylnaphthalene)、脱氧呋喃核糖萘(deoxyribofuranosylnaphthalene)、偏磷酸盐(metaphosphate)、硫代磷酸酯连接、烷基磷酸三酯连接、芳基磷酸三酯连接、膦酸烷基酯连接、膦酸芳基酯连接、氢膦酸酯连接、烷基磷酰胺酯连接及芳基磷酰胺酯连接。现有的碳间隔区也被利用为非-碱基对部分。作为非-碱基对部分的通用碱基对调节探测和标记寡核苷酸-核苷酸变异的切割位置有用。包含如脱氧肌苷(deoxyinosine)、1-(2’-脱氧-β-D-呋喃核糖)-3-硝基吡咯(1-(2’-deoxybeta-D-ribofuranosyl)-3-nitropyrrole)以及5-硝基吲哚(5-nitroindole)之类的通用碱基的碱基对的结合力低于天然碱基间的结合力,通用碱基在特定杂交条件下可利用为非-碱基对部分。导入5’-末端一部分的非-碱基对部分优选地具有1~5个部分,更优选地具有1~2个部分。5’-末端一部分的多个非-碱基对部分能够连续或不连续地存在。优选地,非-碱基对部分具有2~5个连续的部分。优选地,非-碱基对部分为非-碱基对形成化合物。根据一实例,上述探测和标记寡核苷酸-核苷酸变异的核苷酸变异区别位点及非-碱基对部分位于从3’-靶向部位的5’-末端隔开10核苷酸(更优选为8核苷酸、7核苷酸、6核苷酸、5核苷酸、4核苷酸、3核苷酸、2核苷酸或1核苷酸,尤其优选为1核苷酸)的位置以内。根据一实例,在上述探测和标记寡核苷酸-核苷酸变异与错配模板杂交的情况下,第二初期切割位置在包括单链及双链的结构中包括双链的初期位点(即,分叉位点)。根据一实例,上述探测和标记寡核苷酸-核苷酸变异具有阻断剂部位,该阻断剂部位包含对具有5’核酸酶活性的酶引起的切割具有耐性的至少一个核苷酸作为阻断剂,上述阻断剂部分调节初期切割位置或在一个位置或多个位置防止切割。在本说明书中,如探测和标记寡核苷酸-核苷酸变异的5’-标记部位的一部分、探测和标记寡核苷酸-核苷酸变异的3’-靶向部位的5’-末端一部分及捕捉和模板化寡核苷酸的捕捉部位的5’-末端一部分等与探测和标记寡核苷酸-核苷酸变异或捕捉和模板化寡核苷酸相关地使用的术语“一部分(part)”意味着由1-40核苷酸、1-30核苷酸、1-20核苷酸、1-15核苷酸、1-10核苷酸或1-5核苷酸,优选地,由1核苷酸、2核苷酸、3核苷酸或4核苷酸形成的核苷酸序列。根据一实例,具有5’核酸酶活性的酶为具有5’核酸酶活性的脱氧核糖核酸聚合酶或瓣状核酸内切酶,更优选为具有5’核酸酶活性的热稳定性脱氧核糖核酸聚合酶或瓣状核酸内切酶。适合于本发明的具有5’核酸酶活性的脱氧核糖核酸聚合酶为从各种细菌种得到的热稳定性脱氧核糖核酸聚合酶,其包含Thermusaquaticus(Taq)、Thermusthermophilus(Tth)、Thermusfiliformis、Thermisflavus、Thermococcusliteralis、Thermusantranikianii、Thermuscaldophilus、Thermuschliarophilus、Thermusflavus、Thermusigniterrae、Thermuslacteus、Thermusoshimai、Thermusruber、Thermusrubens、Thermusscotoductus、Thermussilvanus、ThermusspeciesZ05、Thermusspeciessps17、Thermusthermophilus、Thermotogamaritima、Thermotoganeapolitana、Thermosiphoafricanus、Thermococcuslitoralis、Thermococcusbarossi、Thermococcusgorgonarius、Thermotogamaritima、Thermotoganeapolitana、Thermosiphoafricanus、Pyrococcuswoesei、Pyrococcushorikoshii、Pyrococcusabyssi、Pyrodictiumoccultum、Aquifexpyrophilus及Aquifexaeolieus。最优选地,热稳定性脱氧核糖核酸聚合酶为Taq聚合酶。根据一实例,模板-依赖性聚合酶用于上游及下游引物的延伸。根据一实例,用于引物的延伸的模板-依赖性聚合酶与具有5’核酸酶活性的酶相同或不同于具有5’核酸酶活性的酶。【步骤(c):从探测和标记寡核苷酸-核苷酸变异放出的片段与捕捉和模板化寡核苷酸的杂交】从探测和标记寡核苷酸-核苷酸变异放出的片段与捕捉和模板化寡核苷酸(CTO,CapturingandTemplatingOligonucleotide)杂交。上述捕捉和模板化寡核苷酸沿着3’→5’方向包含:(i)捕捉部位,其包含与探测和标记寡核苷酸的5’-标记部位或5’-标记部位的一部分互补的核苷酸序列,以及(ii)模板化部位,其包含与探测和标记寡核苷酸的5’-标记部位以及3’-靶向部位非-互补的核苷酸序列。通常,第一片段及第二片段包含可以与捕捉和模板化寡核苷酸的捕捉部位杂交的序列,由此第一片段及第二片段中的一个与捕捉和模板化寡核苷酸杂交。与错配模板杂交而生成的第二片段与杂交于匹配模板而生成的第一片段不同地包含追加的3’-末端部位。根据一实例,在上述第二片段与上述捕捉和模板化寡核苷酸的捕捉部位杂交的情况下,上述捕捉和模板化寡核苷酸不与上述第二片段的上述追加的3’-末端部位杂交,因此,选择捕捉和模板化寡核苷酸的序列,以防止上述第二片段的延伸。例如,捕捉和模板化寡核苷酸的序列能够以在第二片段的追加的3’-末端部位的对面捕捉和模板化寡核苷酸具有错配核苷酸的方式被选择。择一性地,通用碱基可根据反应条件替代错配核苷酸。在某种条件下,第一初期切割位置(或第二初期切割位置)可以不固定,并且是多个。例如,初期切割位置可沿着5’→3’方向位于包含单链及双链的结构中的双链的起始核苷酸(即,分叉位点)及从上述起始核苷酸隔开1-2核苷酸的位置。这种情况下,优选地,在本发明中,选择捕捉和模板化寡核苷酸的序列,来使借助第一初期切割放出的片段中的最短的片段选择性地被延伸,从而生成表示核苷酸变异的存在的延伸链。只要上述捕捉和模板化寡核苷酸的模板化部位具有与探测和标记寡核苷酸-核苷酸变异的5’-标记部位以及3’-靶向部位非-互补的序列,那么就可以包含任何序列。并且,只要模板化部位可以起到用于延伸从探测和标记寡核苷酸-核苷酸变异放出的片段的模板的作用,那么就可以包含任何序列。上述捕捉和模板化寡核苷酸的长度可以多样。例如,上述捕捉和模板化寡核苷酸具有7-1000核苷酸、7-500核苷酸、7-300核苷酸、7-100核苷酸、7-80核苷酸、7-60核苷酸、7-40核苷酸、15-1000核苷酸、15-500核苷酸、15-300核苷酸、15-100核苷酸、15-80核苷酸、15-60核苷酸、15-40核苷酸、20-1000核苷酸、20-500核苷酸、20-300核苷酸、20-100核苷酸、20-80核苷酸、20-60核苷酸、20-40核苷酸、30-1000核苷酸、30-500核苷酸、30-300核苷酸、30-100核苷酸、30-80核苷酸、30-60核苷酸或30-40核苷酸的长度。只要与从探测和标记寡核苷酸放出的片段特异性地杂交,上述捕捉和模板化寡核苷酸的捕捉部位就可以具有任何长度。例如,上述捕捉和模板化寡核苷酸的捕捉部位可以具有5-100核苷酸、5-60核苷酸、5-40核苷酸、5-30核苷酸、5-20核苷酸、10-100核苷酸、10-60核苷酸、10-40核苷酸、10-30核苷酸、10-20核苷酸、15-100核苷酸、15-60核苷酸、15-40核苷酸、15-30核苷酸或15-20核苷酸的长度。并且,只要上述捕捉和模板化寡核苷酸的模板化部位在从探测和标记寡核苷酸放出的片段的延伸反应中可起到模板的作用,上述捕捉和模板化寡核苷酸的模板化部位就可以具有任何长度。例如,上述捕捉和模板化寡核苷酸的模板化部位可以具有1-900核苷酸、1-400核苷酸、1-300核苷酸、1-100核苷酸、1-80核苷酸、1-60核苷酸、1-40核苷酸、1-20核苷酸、2-900核苷酸、2-400核苷酸、2-300核苷酸、2-100核苷酸、2-80核苷酸、2-60核苷酸、2-40核苷酸、2-20核苷酸、5-900核苷酸、5-400核苷酸、5-300核苷酸、5-100核苷酸、5-80核苷酸、5-60核苷酸、5-40核苷酸、5-30核苷酸、10-900核苷酸、10-400核苷酸、10-300核苷酸、15-900核苷酸、15-100核苷酸、15-80核苷酸、15-60核苷酸、15-40核苷酸或15-20核苷酸的长度。上述捕捉和模板化寡核苷酸的3’-末端可以具有3’-OH末端。根据一实例,捕捉和模板化寡核苷酸的3’-末端被阻断,以防止上述捕捉和模板化寡核苷酸的3’-末端的延伸。通过现有方法就可以实现阻断捕捉和模板化寡核苷酸的非-延伸。从上述探测和标记寡核苷酸-核苷酸变异放出的第一片段与捕捉和模板化寡核苷酸杂交,并提供适合于第一片段的延伸的形态。并且,即使非切割探测和标记寡核苷酸-核苷酸变异通过其5’-标记部位与捕捉和模板化寡核苷酸的捕捉部位杂交,由于探测和标记寡核苷酸-核苷酸变异的3’-靶向部位不与捕捉和模板化寡核苷酸杂交,因而防止延伸二聚物的形成。参照对步骤(a)中的杂交的说明,就可以详细地说明步骤(c)中的杂交。【步骤(d):片段的延伸】利用上述步骤(c)的产物及模板-依赖性核酸聚合酶来实施延伸反应。若第一片段与捕捉和模板化寡核苷酸的捕捉部位杂交,则第一片段延伸来形成包含与捕捉和模板化寡核苷酸的模板化部位互补的延伸序列的延伸链。在上述第二片段与捕捉和模板化寡核苷酸的捕捉部位杂交的情况下,第二片段不延伸。在本说明书中与延伸链相关地使用的术语“延伸序列(extendedsequence)”仅意味着在延伸链中除了第一片段之外的重新延伸的序列。上述延伸链包含上述第一片段及上述延伸序列。在特定实例中,第一片段的延伸链及捕捉和模板化寡核苷酸在步骤(d)中形成延伸二聚物。在本说明书中使用的术语“延伸二聚物”意味着基于将模板-依赖性核酸聚合酶和捕捉和模板化寡核苷酸的模板化部位利用为模板使与捕捉和模板化寡核苷酸的捕捉部位杂交的第一片段被延伸的延伸反应而形成的二聚物。上述延伸二聚物的Tm值与非切割探测和标记寡核苷酸-核苷酸变异以及捕捉和模板化寡核苷酸之间的杂交物(hybrid)的Tm值不同。优选地,上述延伸二聚物的Tm值大于非切割探测和标记寡核苷酸和捕捉和模板化寡核苷酸之间的杂交物的Tm值。上述延伸二聚物的Tm值可根据(i)第一片段的序列和/或长度、(ii)捕捉和模板化寡核苷酸的序列和/或长度,或(iii)第一片段的序列和/或长度以及捕捉和模板化寡核苷酸的序列和/或长度来调节。在步骤(e)中,为了基于延伸二聚物的解链来提供表示延伸二聚物的存在的靶信号,可以使用延伸二聚物的可调节的Tm值。在本说明书中使用的术语“Tm”意味着双链核酸分子的群体(population)的一半解离为单链分子的解链温度。Tm值可根据杂交的核苷酸的长度及G/C含量来决定。Tm值可利用如Wallacerule(R.B.Wallace等,NucleicAcidsResearch,6:3543-3547(1979))及nearest-neighbor方法(SantaLuciaJ.Jr.等,Biochemistry,35:3555-3562(1996));SugimotoN.等,NucleicAcidsRes.,24:4501-4505(1996))之类的现有的方法来计算。根据一实例,Tm值意味着实际使用的反应条件下的实际Tm值。在步骤(d)中利用的模板-依赖性核酸聚合酶包含任何核酸聚合酶,例如,E.coliDNA聚合酶I的“克列诺(Klenow)”片段、热稳定性DNA聚合酶以及噬菌体T7DNA聚合酶。优选地,聚合酶为能够从各种细菌种得到的热稳定性DNA聚合酶,其包含Thermusaquaticus(Taq)、Thermusthermophilus(Tth)、Thermusfiliformis、Thermisflavus、Thermococcusliteralis、Thermusantranikianii、Thermuscaldophilus、Thermuschliarophilus、Thermusflavus、Thermusigniterrae、Thermuslacteus、Thermusoshimai、Thermusruber、Thermusrubens、Thermusscotoductus、Thermussilvanus、ThermusspeciesZ05、Thermusspeciessps17、Thermusthermophilus、Thermotogamaritima、Thermotoganeapolitana、Thermosiphoafricanus、Thermococcuslitoralis、Thermococcusbarossi、Thermococcusgorgonarius、Thermotogamaritima、Thermotoganeapolitana、Thermosiphoafricanus、Pyrococcusfuriosus(Pfu)、Pyrococcuswoesei、Pyrococcushorikoshii、Pyrococcusabyssi、Pyrodictiumoccultum、Aquifexpyrophilus及Aquifexaeolieus。最优选地,模板-依赖性核酸聚合酶为Taq聚合酶。根据一实例,在步骤(b)中利用的具有5’核酸酶活性的酶与在步骤(d)中利用的模板-依赖性核酸聚合酶相同。更优选地,在上述步骤(b)中利用的具有5’核酸酶活性的酶、为了使上游引物延伸而利用的模板-依赖性核酸聚合酶以及在步骤(d)中利用的模板-依赖性核酸聚合酶相同。通常,可通过引物的3’-末端一部分和模板之间的杂交来调节引物的延伸。可通过调节引物序列及反应条件(例如,退火温度)来延伸在3’-末端一部分具有1-3错配核苷酸的引物。择一性地,只有在引物具有与靶序列完全互补的序列时,才可以进行引物的延伸。根据一实例,上述捕捉和模板化寡核苷酸的序列被选择为使第一片段或第二片段中的一个选择性地延伸。根据一实例,在上述片段的延伸为片段的3’-末端一部分中只存在一个错配的情况下,在未延伸的条件下进行上述片段的延伸。【步骤(e):延伸链的检测】在延伸之后检测延伸链。上述延伸链的存在表示的探测和标记寡核苷酸-核苷酸变异的核苷酸区别位点互补的核苷酸变异的存在。在本发明中,可形成非切割探测和标记寡核苷酸-核苷酸变异和捕捉和模板化寡核苷酸之间的杂交物或第二片段和捕捉和模板化寡核苷酸之间杂交物。以下所述的非切割探测和标记寡核苷酸-核苷酸变异和捕捉和模板化寡核苷酸之间的杂交物与延伸二聚物的差异同样可适用于第二片段和捕捉和模板化寡核苷酸之间的杂交物和延伸二聚物的差异。【基于解链或杂交分析的延伸二聚物的检测】根据一实例,可根据包括利用延伸链和捕捉和模板化寡核苷酸之间的延伸二聚物的信号的解链分析的探测和标记寡核苷酸切割及延伸分析来实施步骤(e)的检测(参照WO2012/096523)。根据一实例,第一片段的延伸链和捕捉和模板化寡核苷酸在上述步骤(d)中形成延伸二聚物,上述延伸二聚物具有能够基于如下条件而调节的Tm值:(i)上述第一片段的序列和/或长度,(ii)上述捕捉和模板化寡核苷酸的序列和/或长度或(iii)上述第一片段的序列和/或长度及上述捕捉和模板化寡核苷酸的序列和/或长度,上述延伸二聚物提供基于如下标记的靶信号:(i)与上述第一片段和/或上述捕捉和模板化寡核苷酸相连接的至少一个标记,(ii)在延伸反应期间向上述延伸二聚物内插入的标记,(iii)与上述第一片段和/或捕捉和模板化寡核苷酸相连接的至少一个标记及在上述延伸反应期间向上述延伸二聚物内插入的标记或(iv)嵌入标记(intercalatinglabel),根据对于上述延伸二聚物的解链分析或杂交分析,从上述延伸二聚物测定上述靶信号,来检测上述延伸链的存在。本说明书所使用的术语“解链分析(meltinganalysis)”意味着通过延伸二聚物的解链得到表示延伸二聚物的存在的靶信号的方法,包括在两个不同的温度条件下测定信号的方法、解链曲线分析、解链模式分析及解链峰分析。优选地,解链分析为解链曲线分析。根据一实例,在步骤(e)中通过解链分析检测延伸链的存在,延伸二聚物在提供表示延伸二聚物的存在的靶信号的规定范围的温度下被解链。择一性地,在步骤(e)中通过杂交分析检测延伸链的存在。优选地,在步骤(e)中通过杂交分析检测延伸链的存在,且延伸二聚物被解链,由此得到的产物在提供表示延伸二聚物的存在的靶信号的规定范围的温度下杂交。根据一实例,在步骤(e)的解链之后实施杂交,来提供表示上述延伸二聚物的存在的靶信号。这种情况下,借助杂交曲线分析检测延伸二聚物的存在。解链曲线或杂交曲线可通过例如美国专利第6174670号及第5789167号、Drobyshev等、Gene188:45(1997);Kochinsky及MirzabekovHumanMutation19:343(2002);Livehits等,J.Biomol.StructureDynam.11:783(1994);以及Howell等,NatureBiotechnology17:87(1999)中所述的现有技术来获得。例如,解链曲线或杂交曲线可由对于杂交严格度(stringency)的参数的输出信号(outputsignal)变化的图表绘制(graphicplot)或显示(display)构成。输出信号可直接对杂交参数进行绘图。典型地,例如,解链曲线或杂交曲线在Y-轴绘图表示二聚物结构的程度(即,杂交程度)的输出信号,例如荧光,在X-轴绘图(plotting)有杂交参数。荧光vs.温度的一阶导数(derivative)的绘图,即在荧光vs.温度(dF/dTvs.T)或(-dF/dTvs.T)的变化率绘图提供解链峰。根据解链或杂交分析来实施的步骤(e)能够以与如下多种标记系统一同进行详细说明。【(i)与第一片段和/或捕捉和模板化寡核苷酸相连接的标记】根据一实例,上述靶信号根据与第一片段和/或捕捉和模板化寡核苷酸相连接的至少一个标记来提供。由于延伸二聚物形成于第一片段和捕捉和模板化寡核苷酸之间,因此,第一片段或捕捉和模板化寡核苷酸上的标记存在于延伸二聚物上,并在解链步骤中提供靶信号。上述标记包含相互作用性双重标记及单一标记。【(i-1)相互作用性双重标记】作为相互作用性标记系统的代表性例,荧光共振能量转移(FRET,fluorescenceresonanceenergytransfer)标记系统包含荧光报道分子(供体分子)及猝灭分子(受体分子)。在荧光共振能量转移中,能量供体为荧光性,但是能量受体可以为荧光性或非-荧光性。在相互作用性标记系统的再一例中,能量供体为非-荧光性,例如为发色团(chromophore),能量受体为荧光性。在相互作用性标记系统的另一例中,能量供体为发光性(luminescent),例如为生物发光性、化学发光性或电化学发光性,能量受体为荧光性。在本发明中,可将供体分子及受体分子分别记述为报道分子及猝灭分子。相互作用性双重标记包含用于提供可基于接触-介导后的淬灭来检测的信号的标记对(Salvatore等.,NucleicAcidsResearch,2002(30)no.21e122及Johansson等.,J.AM.CHEM.SOC2002(124)pp6950-6956)。在本发明中,上述相互作用性标记系统包括用于诱导基于至少两个分子(例如,染料)之间的相互作用的信号变化的所有情况。尤其,表示延伸二聚物的存在(即,靶核酸序列的存在)的信号借助相互作用性标记系统产生,更优选地,借助荧光共振能量转移标记系统(即,相互作用性双重标记系统)产生。【实例1(链间相互作用性-双重标记)】在相互作用性双重标记系统的实例1中,第一片段或捕捉和模板化寡核苷酸具有包含报道分子及猝灭分子的相互作用性双重标记,在上述步骤(e)中的延伸二聚物的解链诱导来自相互作用性双重标记的信号的变化,来在步骤(e)中提供靶信号。在图5中以图示性的方式地示出相互作用性双重标记系统的实例1。实例1被命名为链间相互作用性-双重标记。【实例1-1(捕捉和模板化寡核苷酸上的链间相互作用性-双重标记)】参照图5,对例示的实例进行说明。捕捉和模板化寡核苷酸的模板化部位具有报道分子及猝灭分子。与靶核酸序列杂交的探测和标记寡核苷酸-核苷酸变异被切割并放出第一片段,且上述第一片段与捕捉和模板化寡核苷酸的捕捉部位杂交并被延伸,来形成延伸二聚物。在上述步骤(d)中形成延伸二聚物的情况下,捕捉和模板化寡核苷酸的报道分子及猝灭分子在结构上(conformationally)相互隔开,使得猝灭分子不对来自报道分子的信号进行猝灭,在步骤(e)中延伸二聚物被解链的情况下,报道分子及猝灭分子在结构上相互接近,使得猝灭分子对来自报道分子的信号进行猝灭,由此提供靶信号,并在步骤(e)中表示延伸二聚物的存在。在本说明书中使用的表达“报道分子及猝灭分子在结构上接近”意味着第一片段或捕捉和模板化寡核苷酸的形态结构,例如,意味着借助无规卷曲(coli)及发夹结构使报道分子及猝灭分子以三维方式相邻。在本说明书中使用的表达“报道分子及猝灭分子在结构上隔开”意味着基于双链的形成的第一片段或捕捉和模板化寡核苷酸的形态结构的变化,使报道分子及猝灭分子以三维方式隔开。优选地,在步骤(e)中提供的靶信号包含测定在上述步骤(d)中产生的荧光信号的变化来得到的解链曲线、解链模式或Tm值。根据一实例,只要与延伸二聚物解链对来自报道分子的信号进行猝灭或不对来自报道分子的信号进行猝灭,那么报道分子及猝灭分子可以位于捕捉和模板化寡核苷酸的任何位置。根据一实例,报道分子及猝灭分子均与捕捉和模板化寡核苷酸的模板化部位或捕捉部位相连接。根据一实例,报道分子及猝灭分子位于捕捉和模板化寡核苷酸的5’-末端及3’-末端。根据一实例,捕捉和模板化寡核苷酸的报道分子及猝灭分子中的一个位于其5’-末端或从其5’-末端隔开1-5核苷酸的位置,另一个位于根据捕捉和模板化寡核苷酸的形态来对来自报道分子的信号进行猝灭和不对来自报道分子的信号进行猝灭的位置。根据一实例,捕捉和模板化寡核苷酸的报道分子及猝灭分子中的一个位于其3’-末端或从其3’-末端隔开1-5核苷酸的位置,另一个位于根据捕捉和模板化寡核苷酸的形态来对来自报道分子的信号进行猝灭和不对来自报道分子的信号进行猝灭的位置。根据一实例,报道分子及猝灭分子相互最大隔开80核苷酸,更优选为最大60核苷酸,尤其优选为最大30核苷酸,最优选为最大25核苷酸。根据一实例,报道分子及猝灭分子隔开至少4核苷酸,更优选为至少6核苷酸,尤其优选为至少10核苷酸,最优选为至少15核苷酸。在本发明中,可形成非切割探测和标记寡核苷酸-核苷酸变异和捕捉和模板化寡核苷酸之间的杂交物。如图5所示,捕捉和模板化寡核苷酸的模板化部位由相互作用性双重标记标记的情况下,不会从非切割探测和标记寡核苷酸-核苷酸变异以及捕捉和模板化寡核苷酸之间的杂交物的标记诱导信号的变化。因此,上述杂交物不提供非-靶信号。捕捉和模板化寡核苷酸的捕捉部位由相互作用性双重标记标记的情况下,非切割探测和标记寡核苷酸以及捕捉和模板化寡核苷酸之间的杂交物提供非-靶信号。这种情况下,可根据延伸二聚物及杂交物的Tm值的差异来区别延伸二聚物的靶信号和杂交物的非-靶信号。【实例1-2(探测和标记寡核苷酸-核苷酸变异上的链间相互作用性-双重标记)】探测和标记寡核苷酸-核苷酸变异的5’-标记部位可具有报道分子及猝灭分子。与靶核酸序列杂交的探测和标记寡核苷酸-核苷酸变异被切割,来放出包含具有报道分子及猝灭分子的5’-标记部位的第一片段。上述第一片段与捕捉和模板化寡核苷酸的捕捉部位杂交。在上述步骤(d)中形成延伸二聚物的情况下,上述第一片段上的报道分子及猝灭分子在结构上相互隔开,使得猝灭分子不对来自报道分子的信号进行猝灭,在步骤(e)中延伸二聚物被解链的情况下,报道分子及猝灭分子在结构上相互接近,使得猝灭分子对来自报道分子的信号进行猝灭,由此提供靶信号,并在步骤(e)中表示延伸二聚物的存在。根据一实例,只要与延伸二聚物解链对来自报道分子的信号进行猝灭或不对来自报道分子的信号进行猝灭,报道分子及猝灭分子可以位于第一片段的任何位置。根据一实例,第一片段上的报道分子及猝灭分子中的一个位于其5’-末端或从其5’-末端隔开1-5核苷酸的位置,另一个位于根据第一片段的形态(conformation)来对来自报道分子的信号进行猝灭和不对来自报道分子的信号进行猝灭的位置。根据一实例,报道分子及猝灭分子相互最大隔开50核苷酸,更优选为最大40核苷酸,尤其优选为最大30核苷酸,最优选为最大20核苷酸。根据一实例,报道分子及猝灭分子隔开至少4核苷酸,更优选为至少6核苷酸,尤其优选为至少10核苷酸,最优选为至少15核苷酸。非切割探测和标记寡核苷酸-核苷酸变异以及捕捉和模板化寡核苷酸之间的杂交物可以在解链步骤中提供非-靶信号。在这种情况下,可通过延伸二聚物及上述杂交物的Tm值的差异来区别延伸二聚物的靶信号和上述杂交物的非-靶信号。【实例2(链间相互作用性-双重标记)】在相互作用性标记系统的实例2中,第一片段具有包含报道分子及猝灭分子的相互作用性双重标记中的一个,捕捉和模板化寡核苷酸具有相互作用性双重标记的另一个;在上述步骤(e)中的延伸二聚物的解链诱导来自相互作用性双重标记的信号的变化,并在步骤(e)中提供靶信号。例如,在上述步骤(d)中形成延伸二聚物的情况下,借助与探测和标记寡核苷酸-核苷酸变异相连接的猝灭分子对与捕捉和模板化寡核苷酸相连接的来自报道分子的信号进行猝灭。在步骤(e)中延伸二聚物被解链的情况下,报道分子及猝灭分子相互隔开,使得猝灭分子不对来自报道分子的信号进行猝灭,由此提供靶信号,并在步骤(e)中表示延伸二聚物的存在。优选地,在步骤(e)中提供的靶信号包含测定来自相互作用性双重标记的荧光信号的变化来得到的解链曲线、解链模式或Tm值。只要借助延伸二聚物的猝灭分子对来自报道分子的信号进行猝灭,报道分子及猝灭分子可以位于第一片段及捕捉和模板化寡核苷酸的任何位置。根据一实例,探测和标记寡核苷酸-核苷酸变异片段上的报道分子或猝灭分子位于5’-标记部位的5’-末端。根据一实例,捕捉和模板化寡核苷酸的报道分子或猝灭分子位于其3’-末端。非切割探测和标记寡核苷酸以及捕捉和模板化寡核苷酸之间的杂交物可在解链步骤中提供非-靶信号。这种情况下,可根据延伸二聚物及杂交物的Tm值的差异来区别延伸二聚物的靶信号和杂交物的非-靶信号。在本发明中有用的报道分子及猝灭分子可包含在本发明所属技术领域中公知的任何分子。具体例如下:Cy2TM(506)、YO-PROTM-1(509)、YOYOTM-1(509)、Calcein(517)、FITC(518)、FluorXTM(519)、AlexaTM(520)、Rhodamine110(520)、OregonGreenTM500(522)、OregonGreenTM488(524)、RiboGreenTM(525)、RhodamineGreenTM(527)、Rhodamine123(529)、MagnesiumGreenTM(531)、CalciumGreenTM(533)、TO-PROTM-1(533)、TOTO1(533)、JOE(548)、BODIPY530/550(550)、Dil(565)、BODIPYTMR(568)、BODIPY558/568(568)、BODIPY564/570(570)、Cy3TM(570)、AlexaTM546(570)、TRITC(572)、MagnesiumOrangeTM(575)、PhycoerythrinR&B(575)、RhodaminePhalloidin(575)、CalciumOrangeTM(576)、PyroninY(580)、RhodamineB(580)、TAMRA(582)、RhodamineRedTM(590)、Cy3.5TM(596)、ROX(608)、CalciumCrimsonTM(615)、AlexaTM594(615)、TexasRed(615)、NileRed(628)、YO-PROTM-3(631)、YOYOTM-3(631)、Rphycocyanin(642)、C-Phycocyanin(648)、TO-PROTM-3(660)、TOTO3(660)、DiDDilC(5)(665)、Cy5TM(670)、Thiadicarbocyanine(671)、Cy5.5(694)、HEX(556)、TET(536)、BiosearchBlue(447)、CALFluorGold540(544)、CALFluorOrange560(559)、CALFluorRed590(591)、CALFluorRed610(610)、CALFluorRed635(637)、FAM(520)、Fluorescein(520)、Fluorescein-C3(520)、Pulsar650(566)、Quasar570(667)、Quasar670(705)及Quasar705(610)。括号内的数字为以纳米为单位表示的最大的发光波长。优选地,报道分子及猝灭分子包含JOE、FAM、TAMRA、ROX及基于荧光素的标记。适合的一对报道分子-猝灭分子公开在许多文献中:Pesce等,editors,FluorescenceSpectroscopy(MarcelDekker,NewYork,1971);White等,FluorescenceAnalysis:APracticalApproach(MarcelDekker,NewYork,1970);Berlman,HandbookofFluorescenceSpectraofAromaticMolecules,2ndEdition(AcademicPress,NewYork,1971);Griffiths,ColorANDConstitutionofOrganicMolecules(AcademicPress,NewYork,1976);Bishop,editor,Indicators(PergamonPress,Oxford,1972);Haugland,HandbookofFluorescentProbesandResearchChemicals(MolecularProbes,Eugene,1992);Pringsheim,FluorescenceandPhosphorescence(IntersciencePublishers,NewYork,1949);Haugland,R.P.,HandbookofFluorescentProbesandResearchChemicals,6thEdition(MolecularProbes,Eugene,Oreg.,1996);美国专利第3996345号及第4351760号。在本发明中,值得关注的是,可以利用能够对广范围波长或特定波长的荧光进行猝灭的非-荧光黑猝灭分子。作为非-荧光黑猝灭分子的例,有BHQ及DABCYL。在适用于捕捉和模板化寡核苷酸的荧光共振能量转移标记中,报道分子包含荧光共振能量转移的供体,猝灭分子包含荧光共振能量转移的另一伙伴(受体)。例如,荧光素染料(fluoresceindye)用作报道分子,罗丹明染料(rhodaminedye)用作猝灭分子。上述标记可通过现有的方法与捕捉和模板化寡核苷酸或探测和标记寡核苷酸相连接。优选地,上述标记通过包含碳原子的间隔区(例如,3-碳间隔区、6-碳间隔区或12-碳间隔区)与捕捉和模板化寡核苷酸或探测和标记寡核苷酸相连接。【(i-2)单一标记】并且,本发明利用提供表示靶核酸序列的存在的信号的单一标记系统来优秀地被实施。根据一实例,第一片段或捕捉和模板化寡核苷酸具有单一标记,在步骤(e)中的延伸二聚物的解链诱导来自单一标记的信号的变化,并在步骤(e)中提供靶信号。【实例1(单一标记系统)】捕捉和模板化寡核苷酸的模板化部位可具有单一荧光标记。与靶核酸序列杂交的探测和标记寡核苷酸-核苷酸变异被切割,来放出第一片段。第一片段与捕捉和模板化寡核苷酸的捕捉部位杂交并被延伸,来形成延伸二聚物。延伸二聚物的形成使来自单一荧光标记的荧光强度增加。延伸二聚物在步骤(e)中被解链的情况下,来自上述单一荧光标记的荧光强度减少,由此提供靶信号,并在步骤(e)中表示延伸二聚物的存在。根据一实例,只要根据延伸二聚物的解链使来自上述单一标记的信号的水平发生变化,那么上述单一标记就可以位于捕捉和模板化寡核苷酸的任何位置。根据一实例,上述单一标记与捕捉和模板化寡核苷酸的模板化部位或捕捉部位相连接。捕捉和模板化寡核苷酸的模板化部位由单一标记标记的情况下,不诱导来自非切割测和标记寡核苷酸-核苷酸变异以及捕捉和模板化寡核苷酸之间的杂交物的标记的信号的变化。因此,杂交物不提供非-靶信号。捕捉和模板化寡核苷酸的捕捉部位由单一标记标记的情况下,非切割探测和标记寡核苷酸-核苷酸变异以及捕捉和模板化寡核苷酸之间的杂交物提供非-靶信号。这种情况下,可根据延伸二聚物及上述杂交物的Tm值的差异来区别延伸二聚物的靶信号和杂交物的非-靶信号。【实例2(单一标记系统)】探测和标记寡核苷酸-核苷酸变异的5’-标记部位可具有单一荧光标记。与靶核酸序列杂交的探测和标记寡核苷酸-核苷酸变异被切割,来放出包含具有单一荧光标记的5’-标记部位的第一片段。借助杂交使来自5’-标记部位的上述单一荧光标记的荧光强度增加。延伸二聚物在步骤(e)中被解链的情况下,来自上述单一荧光标记的信号强度减少,由此提供靶信号,并在步骤(e)中表示延伸二聚物的存在。根据一实例,只要根据延伸二聚物的解链使来自上述单一标记的信号的水平发生变化,上述单一标记就可以位于探测和标记寡核苷酸片段的任何位置。非切割探测和标记寡核苷酸-核苷酸变异以及捕捉和模板化寡核苷酸之间的杂交物可在解链步骤中提供非-靶信号。在这种情况下,可根据延伸二聚物及杂交物的Tm值的差异来区别延伸二聚物的靶信号和杂交物的非-靶信号。在本说明书中使用的单一标记应根据存在于双链上或单链上来提供不同的信号。上述单一标记包括荧光标记、发光标记、化学发光标记、电化学标记及金属标记。优选地,上述单一标记包括荧光标记。在本发明中使用的单一荧光标记的种类及优选结合位置公开在美国专利第7537886号及第7348141号中,且相关教导事项包括在本说明书作为参照。优选地,上述单一荧光标记包含JOE、FAM、TAMRA、ROX以及基于荧光素的标记。优选地,被标记的核苷酸残基位于寡核苷酸内的内部核苷酸残基,而不是5’-末端或3’-末端。参照如上所述的报道分子及猝灭分子的说明,对在本发明中有用的单一荧光标记进行说明。尤其,在固相中利用单一标记来实施本发明的情况下,可以利用通常的荧光标记,不需要可根据存在于双链上或单链上提供具有不同的强度的荧光信号的特定的荧光标记。测定在固相底物中提供的靶信号。利用在固相底物实现固定化的捕捉和模板化寡核苷酸的情况下,可以利用化学标记(例如,生物素)或酶标记(例如,碱性磷酸酶、过氧化物酶、β-半乳糖苷酶及β-葡萄糖苷酶)。在利用“与第一片段和/或捕捉和模板化寡核苷酸相连接的标记”的标记系统中,形成非切割探测和标记寡核苷酸-核苷酸变异以及捕捉和模板化寡核苷酸之间的杂交物的情况下,标记位于上述杂交物在步骤(e)中不提供非-靶信号的范围内。择一性地,形成非切割探测和标记寡核苷酸-核苷酸变异以及捕捉和模板化寡核苷酸之间的杂交物的情况下,标记可以位于上述杂交物在步骤(e)中提供非-靶信号的范围内,上述延伸二聚物的Tm值高于非切割探测和标记寡核苷酸-核苷酸变异以及捕捉和模板化寡核苷酸之间的杂交物的Tm值。尤其,在上述标记位于非切割探测和标记寡核苷酸-核苷酸变异以及捕捉和模板化寡核苷酸之间的杂交物不提供非-靶信号的范围内的情况下,包含上述杂交物的Tm值的区间可以利用于选择用于检测靶核酸序列的延伸二聚物的Tm值。【(ii)向延伸二聚物内插入的标记】本发明可以为了提供表示延伸二聚物的存在的靶信号而利用在延伸反应期间向延伸二聚物内插入的标记。即使第一片段或捕捉和模板化寡核苷酸不具有标记,也可利用在延伸反应期间向延伸二聚物内插入的标记来成功标记延伸二聚物。根据一实例,靶信号借助在延伸反应期间向延伸二聚物内插入的单一标记提供,上述插入的单一标记与在延伸反应期间插入的核苷酸相连接,在步骤(e)中的上述延伸二聚物的解链诱导来自单一标记的信号的变化,并在步骤(e)中提供靶信号。例如,与靶核酸序列杂交的探测和标记寡核苷酸-核苷酸变异被切割,来放出第一片段。上述第一片段与在固相底物实现固定化的捕捉和模板化寡核苷酸的捕捉部位杂交,并在由单一荧光标记标记的核苷酸存在的条件下被延伸,来形成延伸二聚物。将捕捉和模板化寡核苷酸固定化的固相底物的点可以检测来自延伸二聚物的荧光信号。延伸二聚物被解链的情况下,放出具有荧光标记的链,而在点上再也检测不到荧光信号。因此,能够借助延伸二聚物的解链在点上提供信号的变化。即,提供靶信号,并在步骤(e)中表示延伸二聚物的存在。在步骤(e)中提供的靶信号包括在捕捉和模板化寡核苷酸-固定化点上测定荧光强度的变化来得到的解链曲线、解链模式或Tm值。根据一实例,在延伸反应期间插入(incorporated)的核苷酸为ddNTP。根据一实例,在延伸反应期间插入的核苷酸具有第一非-天然碱基(non-naturalbase),捕捉和模板化寡核苷酸具有对上述第一非-天然碱基具有特异性结合亲和性的第二非-天然碱基。优选地,具有第二非-天然碱基的核苷酸位于捕捉和模板化寡核苷酸的模板化部位的任何位置。在本说明书中使用的术语“非-天然碱基”意味着如能够形成氢键碱基对的腺嘌呤(A)、鸟嘌呤(G)、胸腺嘧啶(T)、胞嘧啶(C)及尿嘧啶(U)之类的天然碱基的衍生物。在本说明书中使用的术语“非-天然碱基”包含具有与作为母体化合物(mothercompound)的天然碱基不同的碱基对模式的碱基,例如,在美国专利第5432272号、第5965364号、第6001983号及第6037120号中记载。非-天然碱基之间的碱基对与天然碱基类似地包含两个或三个氢键。并且,非-天然碱基之间的碱基对也以特定的方式形成。作为非-天然碱基的特定的例,包含以下碱基的碱基对组合:iso-C/iso-G、iso-dC/iso-dG、K/X、H/J及M/N(参照美国专利第7422850号)。例如,包含具有第二非-天然碱基(例如,iso-dC)的核苷酸的捕捉和模板化寡核苷酸与片段杂交,上述第二非-天然碱基对第一非-天然碱基(例如,iso-dG)具有特异性结合亲和性。具有由单一荧光标记标记的第一非-天然碱基的核苷酸存在的条件下,实施延伸来形成延伸二聚物。在延伸反应中,具有第一非-天然碱基的核苷酸插入于具有第二非-天然碱基的核苷酸的对面位置。在实现固定化的捕捉和模板化寡核苷酸存在的固相底物的点上可检测来自延伸二聚物的荧光信号。延伸二聚物被解链的情况下,放出具有荧光标记的链,而在点上再也检测不到荧光信号。因此,可借助延伸二聚物的解链在点上提供信号的变化。即,提供靶信号,并在步骤(e)中表示延伸二聚物的存在。在利用在延伸反应期间向延伸二聚物内插入的标记的情况下,上述标记不包含于非切割探测和标记寡核苷酸-核苷酸变异以及捕捉和模板化寡核苷酸之间的杂交物内,这是因为杂交物不被延伸。因此,杂交物不提供非-靶信号。可参照在本说明书中所述的利用“与第一片段和/或捕捉和模板化寡核苷酸相连接的标记”的标记系统的说明,对所使用的单一标记的种类及特征进行说明。【(iii)向延伸二聚物内插入的标记和与第一片段或捕捉和模板化寡核苷酸相连接的标记】本发明可使用利用在延伸反应期间向延伸二聚物内插入的标记和与片段和/或捕捉和模板化寡核苷酸相连接的标记的组合的标记系统。根据一实例,根据在延伸反应期间向延伸二聚物内插入的标记和与片段和/或捕捉和模板化寡核苷酸相连接的标记提供靶信号,上述插入的标记与在上述延伸反应期间插入的核苷酸相连接,上述两个标记为报道分子及猝灭分子的相互作用性双重标记,在上述步骤(e)中的延伸二聚物的解链诱导来自相互作用性双重标记的信号的变化,并在步骤(e)中提供靶信号。尤其,在延伸反应期间插入的核苷酸具有第一非-天然碱基,捕捉和模板化寡核苷酸具有对上述第一非-天然碱基存在特异性结合亲和性的第二非-天然碱基。例如,包含具有第二非-天然碱基(例如,iso-dC)的核苷酸的捕捉和模板化寡核苷酸与片段杂交,上述第二非-天然碱基对报道分子或猝灭分子及第一非-天然碱基(例如,iso-dG)存在特异性结合亲和性。具有由猝灭分子或报道分子标记的第一非-天然碱基的核苷酸存在的条件下,实施延伸来形成延伸二聚物,并借助猝灭分子来对来自报道分子的信号进行猝灭。在延伸反应中,具有上述第一非-天然碱基的核苷酸插入于具有上述第二非-天然碱基的核苷酸的对面位置。在步骤(e)中延伸二聚物被解链的情况下,报道分子及猝灭分子相互隔开,猝灭分子不对来自报道分子的信号进行猝灭,由此提供靶信号,并在步骤(e)中表示延伸二聚物的存在。尤其,在步骤(e)中提供的靶信号包括测定来自相互作用性双重标记的信号的变化来得到的解链曲线、解链模式或Tm值。在解链步骤中,捕捉和模板化寡核苷酸上的标记位置及所插入的标记的插入位置在上述两个标记起到诱导信号的变化的相互作用性双重标记作用的范围内决定。尤其,捕捉和模板化寡核苷酸的模板化部位具有核苷酸,上述核苷酸具有报道分子或猝灭分子及第二非-天然碱基。在上述步骤(d)中的延伸反应在具有第一非-天然碱基的核苷酸存在的条件下实施,上述第一非-天然碱基对猝灭分子或报道分子及上述捕捉和模板化寡核苷酸内的第二非-天然碱基存在特异性结合亲和性。步骤(d)的延伸二聚物的上述两个非-天然碱基形成碱基对,并借助猝灭分子对来自报道分子的信号进行猝灭,并诱导信号的变化,由此提供靶信号。择一性地,片段具有报道分子或猝灭分子,捕捉和模板化寡核苷酸的模板化部位具有核苷酸,上述核苷酸具有第二非-天然碱基。在上述步骤(d)中的延伸反应在具有第一非-天然碱基的核苷酸存在的条件下实施,上述第一非-天然碱基对猝灭分子或报道分子及上述捕捉和模板化寡核苷酸内的第二非-天然碱基存在特异性结合亲和性。上述步骤(d)的延伸二聚物的两个非-天然碱基形成碱基对,并借助猝灭分子诱导来自报道分子的信号的变化,由此提供靶信号。作为另一例,包含报道分子或猝灭分子的第一片段与包含具有第二非-天然碱基(例如,iso-dC)的核苷酸的捕捉和模板化寡核苷酸杂交,上述第二非-天然碱基对第一非-天然碱基(例如,iso-dG)具有特异性结合亲和性。在具有由猝灭分子或报道分子标记的第一非-天然碱基的核苷酸存在的条件下实施延伸,来形成延伸二聚物,并借助猝灭分子对来自报道分子的信号进行猝灭。在延伸反应中,具有上述第一非-天然碱基的核苷酸插入于具有上述第二非-天然碱基的核苷酸的对面位置。在上述步骤(d)中形成延伸二聚物的情况下,报道分子及猝灭分子在结构上相互隔开,使得猝灭分子不对来自报道分子的信号进行猝灭,在步骤(e)中延伸二聚物被解链的情况下,报道分子及猝灭分子在结构上相互接近,使得猝灭分子对来自报道分子的信号进行猝灭,由此提供靶信号,并在步骤(e)中表示延伸二聚物的存在。尤其,在步骤(e)中提供的靶信号包括测定来自相互作用性双重标记的信号的变化来得到的解链曲线、解链模式或Tm值。在解链步骤中,探测和标记寡核苷酸-核苷酸变异上的标记位置及插入的标记的插入位置在上述两个标记起到诱导信号的变化的相互作用性双重标记作用的范围内决定。利用在延伸反应期间向延伸二聚物内插入的标记的情况下,上述标记不插入于非切割探测和标记寡核苷酸-核苷酸变异以及捕捉和模板化寡核苷酸之间的杂交物内。这是因为上述杂交物不被延伸。因此,上述杂交物在解链步骤中不提供非-靶信号。【(iv)嵌入标记】本发明可以使用提供表示延伸二聚物的存在的靶信号的嵌入标记。由于在样品内,双链核酸分子可产生信号,因此在利用实现固定化的捕捉和模板化寡核苷酸的固相反应中嵌入标记更有用。在本发明中有用的嵌入染料的例包含:SYBRTMGreenI、PO-PROTM-1、BO-PROTM-1、SYTOTM43、SYTOTM44、SYTOTM45、SYTOXTMBlue、POPOTM-1、POPOTM-3、BOBOTM-1、BOBOTM-3、LO-PROTM-1、JO-PROTM-1、YO-PROTM1、TO-PROTM1、SYTOTM11、SYTOTM13、SYTOTM15、SYTOTM16、SYTOTM20、SYTOTM23、TOTOTM-3、YOYOTM3、GelStarTM及噻唑橙(thiazoleorange)。嵌入染料特异性地插入于双链核酸分子内来产生信号。在特定实例中,在固相底物实现固定化的捕捉和模板化寡核苷酸的捕捉部位与第一片段杂交。在嵌入染料(例如,SYBRTMGreen)存在的条件下实施延伸,来生成包含嵌入染料的延伸二聚物。可利用嵌入荧光染料来检测来自具有实现固定化的捕捉和模板化寡核苷酸的固相底物的点上的延伸二聚物的荧光信号。延伸二聚物被解链的情况下,放出嵌入荧光染料,而在点上再也检测不到荧光信号。因此,提供靶信号,并在步骤(e)中表示延伸二聚物的存在。非切割探测和标记寡核苷酸-核苷酸变异以及捕捉和模板化寡核苷酸之间的杂交物在解链步骤中提供非-靶信号。这种情况下,可根据延伸二聚物及上述杂交物的Tm值的差异来区别延伸二聚物的靶信号和杂交物的非-靶信号。尤其,在步骤(e)中提供的靶信号包括测定在步骤(e)中产生的荧光信号的变化来得到的解链曲线、解链模式或Tm值。【在预先指定的温度中检测延伸二聚物】根据一实例,可根据包括利用延伸链和捕捉和模板化寡核苷酸之间的延伸二聚物的信号的解链分析的探测和标记寡核苷酸切割及延伸分析来实施步骤(e)的检测(参照WO2012/096523)。根据一实例,上述第一片段的延伸链和上述捕捉和模板化寡核苷酸在上述步骤(d)形成延伸二聚物;上述延伸二聚物具有可基于如下条件而调节的Tm值:(i)上述第一片段的序列和/或长度,(ii)上述捕捉和模板化寡核苷酸的序列和/或长度或(iii)上述第一片段的序列和/或长度和上述捕捉和模板化寡核苷酸的序列和/或长度,上述延伸二聚物提供基于如下标记的靶信号:(i)与上述第一片段和/或捕捉和模板化寡核苷酸相连接的至少一个标记,(ii)在上述延伸反应期间向上述延伸二聚物内插入的标记,(iii)与上述第一片段和/或捕捉和模板化寡核苷酸相连接的至少一个标记及在上述延伸反应期间向上述延伸二聚物内插入的标记或(iv)嵌入标记,在上述延伸二聚物维持双链所需的充分的指定温度下,从上述延伸二聚物测定靶信号,来检测上述延伸链的存在。延伸二聚物本身可以提供能够区别延伸二聚物的形成和非-形成的信号,在延伸二聚物维持双重-链形态的预先指定的温度中检测上述信号,由此决定靶核酸序列的存在。本发明为了检测靶核酸序列的存在而测定与延伸二聚物的形成相关的靶信号。在本发明中,延伸二聚物具有标记,由此,上述延伸二聚物提供靶信号。在本发明的方法中,用于通过解链或杂交分析来检测延伸二聚物的标记系统可以提供靶信号。来自延伸二聚物的靶信号的基本工作原理如下。(i)第一片段的延伸诱导来自标记的信号的变化来提供靶信号,或(ii)第一片段及捕捉和模板化寡核苷酸的杂交诱导来自标记的信号的变化来提供靶信号,上述延伸二聚物维持靶信号。例如,利用实现固定化的捕捉和模板化寡核苷酸的情况下,本发明以更加有效的方式检测多个靶核酸序列。实现固定化的捕捉和模板化寡核苷酸的模板化部位具有报道分子及猝灭分子。报道分子及猝灭分子在结构上相互接近,使得猝灭分子对来自报道分子的信号进行猝灭。在第一片段与捕捉和模板化寡核苷酸的捕捉部位进行杂交的情况下,猝灭分子对来自报道分子的信号进行猝灭。随着上述延伸二聚物的形成,报道分子及猝灭分子在结构上相互隔开,使得猝灭分子不对来自报道分子的信号进行猝灭。在延伸步骤中提供靶信号。在特定实例中,探测和标记寡核苷酸-核苷酸变异的5’-标记部位具有报道分子及猝灭分子。报道分子及猝灭分子在结构上相互接近,使得猝灭分子对来自报道分子的信号进行猝灭。与靶核酸序列杂交的探测和标记寡核苷酸-核苷酸变异被切割,来放出包含具有报道分子及猝灭分子的5’-标记部位的第一片段,上述第一片段与捕捉和模板化寡核苷酸的捕捉部位杂交。借助上述杂交,报道分子及猝灭分子在结构上相互隔开,使得猝灭分子不对来自报道分子的信号进行猝灭。在片段的杂交步骤中提供靶信号,且延伸二聚物维持靶信号。在探测和标记寡核苷酸-核苷酸变异的5’-标记部位具有报道分子及猝灭分子的情况下,非切割探测和标记寡核苷酸-核苷酸变异以及捕捉和模板化寡核苷酸之间的杂交物提供非-靶信号,并且有必要为了去除上述非-靶信号而使上述杂交物解离。因此,在上述杂交物解离的温度中决定用于测定靶信号的温度。根据一实例,还考虑杂交物的Tm值,来决定上述温度。根据一实例,可在一部分上述杂交物解离的温度下,检测延伸二聚物。根据一实例,可在上述杂交物充分解离并去除非-靶信号的温度中检测延伸二聚物。根据一实例,预先指定的温度高于从杂交物的Tm值减去10℃的温度,优选地,高于从杂交物的Tm值减去5℃的温度,更优选地,高于杂交物的Tm值,尤其优选地,高于在杂交物的Tm值加上5℃的温度。【利用信号寡核苷酸的检测】根据一实例,可利用公开于PCT/KR2012/005281的信号寡核苷酸(SignalingOligonucleotide:SO)来检测第一片段的延伸链。与延伸链杂交的信号寡核苷酸包含与上述延伸链互补的序列。根据一实例,上述信号寡核苷酸包含与延伸序列互补的序列。根据一实例,上述信号寡核苷酸的至少一个部位包含与延伸序列互补的序列。包含与延伸序列互补的序列的信号寡核苷酸的部位的长度至少为1个核苷酸、2个核苷酸、3个核苷酸、4个核苷酸、5个核苷酸或10个核苷酸长度。在上述信号寡核苷酸的一部位被设计成包含与重新合成的延伸序列的一部位互补的序列的情况下,信号寡核苷酸和延伸链的杂交产物的Tm值不同于信号寡核苷酸和非切割探测和标记寡核苷酸-核苷酸变异的杂交产物的Tm值。Tm值的差异可区别来自上述两个杂交产物的信号。例如,可在实时检测中考虑Tm值来调节检测温度,或者可在解链曲线分析中通过解链峰来排除非-靶信号。上述信号寡核苷酸的整个序列可以包含与延伸序列互补的序列。择一性地,上述信号寡核苷酸可以包含具有与延伸序列互补的序列的部位。例如,上述信号寡核苷酸的一部位可以包含与延伸链互补的序列,另一部位可以包含与片段互补的序列。优选地,上述信号寡核苷酸的整个序列包含与延伸序列互补的序列。上述信号寡核苷酸可以具有任何长度长度。例如,可以具有5-100核苷酸、5-80核苷酸、5-60核苷酸、5-40核苷酸、5-20核苷酸、5-10核苷酸、10-100核苷酸、10-80核苷酸、10-60核苷酸、10-40核苷酸、10-30核苷酸、10-20核苷酸、15-100核苷酸、15-80核苷酸、15-60核苷酸、15-40核苷酸、15-30核苷酸、15-20核苷酸、20-100核苷酸、20-80核苷酸、20-60核苷酸、20-40核苷酸或20-30核苷酸的长度。上述信号寡核苷酸可以具有细销结构。上述信号寡核苷酸的3’-末端被阻断,以防止上述信号寡核苷酸的3’-末端的延伸。择一性地,具有未被阻断的3’-OH基的信号寡核苷酸可以被延伸。根据一实例,可利用信号寡核苷酸(SO)来检测上述第一片段的延伸链,上述信号寡核苷酸包含与上述延伸链互补的序列及至少一个标记,上述信号寡核苷酸提供可通过与延伸链之间的结合(association)或解离(dissociation)来检测的信号。术语“与延伸链之间的结合(association)或解离(dissociation)”与术语“与延伸链之间的杂交或变性”具有相同的含义。根据一实例,通过以下组合来提供表示靶核酸序列的存在的可检测的信号:(i)与信号寡核苷酸相连接的标记;(ii)与信号寡核苷酸相连接的标记及与上述探测和标记寡核苷酸的片段相连接的标记的组合;(iii)与述信号寡核苷酸相连接的标记及在上述步骤(d)的延伸反应期间内插入于延伸链的标记的组合;或者(iv)与上述信号寡核苷酸相连接的标记及嵌入染料(intercalatingdye)。简单地,能够以如下方式说明有利于本发明的标记系统。【(i)与信号寡核苷酸相连接的单一标记】本发明可以提供利用具有单一标记的信号寡核苷酸来表示存在靶核苷酸变异的延伸链的形成相关的信号。根据一实例,上述信号寡核苷酸被标记为单一标记,在步骤(e)中,上述信号寡核苷酸和延伸链之间的杂交可以诱导来自单一标记的信号变化,来提供可检测的信号。在特定实例中,本说明书所使用的单一标记应可以根据其标记存在于双链还是单链来提供不同的信号。【(ii)信号寡核苷酸与相连接的链间相互作用性-双重标记】根据一实例,信号寡核苷酸被标记为包含报道分子及猝灭分子的相互作用性双重标记,在步骤(e)中,上述信号寡核苷酸和延伸链之间的杂交诱导来自相互作用性双重标记的信号变化,来提供可检测的信号(参照图6)。在上述信号寡核苷酸杂交之前,上述信号寡核苷酸上的报道分子及猝灭分子在结构上相互接近,使得猝灭分子对来自报道分子的信号进行淬灭。若完成杂交,则上述信号寡核苷酸上的报道分子及猝灭分子在结构上相隔开,使得猝灭分子不对来自报道分子的信号进行淬灭(unquenching),而这会引起来自相互作用性双重标记的信号变化。在利用具有相互作用性双重标记的信号寡核苷酸的本发明的根据一实例中,从与靶核酸序列杂交的探测和标记寡核苷酸-核苷酸变异放出的第一片段与捕捉和模板化寡核苷酸的捕捉部位杂交并延伸,来形成延伸链。若上述延伸链与信号寡核苷酸杂交,则信号寡核苷酸上的报道分子及猝灭分子在结构上相隔开,使得猝灭分子不对来自报道分子的信号进行淬灭,并引起来自相互作用性双重标记的信号变化(例如,增加来自报道分子的信号)。未参与杂交的信号寡核苷酸上的报道分子及猝灭分子在结构上相互接近,使得猝灭分子对来自报道分子的信号进行淬灭。根据一实例,报道分子及猝灭分子位于信号寡核苷酸的5’-末端(或3’-末端)及3’-末端(或5’-末端)。根据一实例,上述信号寡核苷酸上的报道分子及猝灭分子中的一个位于信号寡核苷酸的5’-末端或位于从5’-末端隔开1-5核苷酸的位置,另一个根据信号寡核苷酸的形态来对来自报道分子的信号进行淬灭或不对来自报道分子的信号进行淬灭。根据一实例,上述信号寡核苷酸上的报道分子及猝灭分子中的一个位于信号寡核苷酸的3’-末端或位于从3’-末端隔开1-5核苷酸的位置,另一个根据信号寡核苷酸的形态来对来自报道分子的信号进行淬灭或不对来自报道分子的信号进行淬灭。【(iii)链间相互作用性-双重标记】根据使用链间相互作用性-双重标记的实例,延伸链具有包含报道分子及猝灭分子的相互作用性双重标记中的一种,信号寡核苷酸具有相互作用性双重标记中的另一种。可根据如下三种方法来实施使用链间相互作用性-双重标记的实例。根据第一方法,上述信号寡核苷酸包含相互作用性双重标记的报道分子和猝灭分子中的一种标记,探测和标记寡核苷酸-核苷酸变异的片段包含报道分子和猝灭分子中的另一种标记,延伸链包含源于上述探测和标记寡核苷酸-核苷酸变异的片段的标记,上述信号寡核苷酸和延伸链之间的杂交诱导从相互作用性双重标记提供的信号的变化,来提供可检测的信号。与上述信号寡核苷酸相连接的标记可以为报道分子或猝灭分子中的一种,与上述片段相连接的标记可以为猝灭分子或报道分子中的一种。可以考虑探测和标记寡核苷酸-核苷酸变异切割位点来决定探测和标记寡核苷酸-核苷酸变异上的标记位点,由此,探测和标记寡核苷酸-核苷酸变异片段可具有标记。在探测和标记寡核苷酸-核苷酸变异片段与信号寡核苷酸杂交的情况下,只要可以与连接在信号寡核苷酸的标记相互作用来诱导信号变化,上述标记就可以与探测和标记寡核苷酸-核苷酸变异片段的任何位点(例如,探测和标记寡核苷酸-核苷酸变异的标记部位)相连接。在信号寡核苷酸与探测和标记寡核苷酸-核苷酸变异片段杂交的情况下,只要可以与连接在探测和标记寡核苷酸-核苷酸变异片段的标记相互作用来诱导信号变化,上述标记就可以与信号寡核苷酸的任何位点(例如,信号寡核苷酸的5’-末端)相连接。根据第二方法,上述信号寡核苷酸包含相互作用性双重标记的报道分子和猝灭分子中的一种标记,捕捉和模板化寡核苷酸的模板化部位包含具有第一非-天然碱基的核苷酸,在存在具有与上述第一非-天然碱基特异性结合亲和性的第二非-天然碱基及报道分子和猝灭分子中的另一种的核苷酸的条件下实施上述步骤(d)的延伸反应,由此,标记向延伸链内插入,上述信号寡核苷酸和延伸链之间的杂交诱导来自相互作用性双重标记的信号的变化,来提供可检测的信号。优选地,在延伸反应期间内插入的标记与核苷酸相连接,更优选地,与三磷酸核苷相连接。优选地,上述标记与三磷酸核苷的碱基相结合。上述片段与包含核苷酸的捕捉和模板化寡核苷酸杂交,上述核苷酸包含具有对非-天然碱基(例如,iso-dG)特异性结合亲和性的非-天然碱基(例如,iso-dC)。在具有通过猝灭来标记的iso-dG的核苷酸的存在条件下实施延伸,来形成延伸链。在上述延伸反应中,与猝灭一同具有iso-dG的核苷酸插入于具有iso-dC的核苷酸对面的位点。在包含猝灭-iso-dG的延伸链和被报道分子标记的信号寡核苷酸之间引起杂交之后,延伸链上的猝灭对来自信号寡核苷酸上的报道的信号进行淬灭,来诱导信号的变化,并提供可检测的信号。相互作用性双重标记中的一个与信号寡核苷酸相连接,另一个在延伸反应期间内从溶液向延伸链内插入。与上述信号寡核苷酸相连接的标记可以为报道分子或猝灭分子中的一种,向延伸链插入的标记可以为猝灭分子或报道分子中的一种。只要可以与信号寡核苷酸杂交并与连接在信号寡核苷酸的标记相互作用来诱导信号变化,向延伸链内插入的标记可以与延伸链的任何位点(例如,延伸链的3’-末端)相连接。只要可以与延伸链杂交并与插入于延伸链内的标记相互作用来诱导信号变化,上述标记可与信号寡核苷酸的任何位点(例如,信号寡核苷酸的5’-末端)相连接。根据第三方法,上述信号寡核苷酸包含相互作用性双重标记的报道分子和猝灭分子中的一种标记,在具有报道分子和猝灭分子中的另一种核苷酸存在的条件下实施上述步骤(d)的延伸反应,由此,标记向延伸链内插入,上述信号寡核苷酸和延伸链之间的杂交诱导来自相互作用性双重标记的信号的变化,来提供可检测的信号。与信号寡核苷酸相连接的标记可以为报道分子或猝灭分子中的一种(优选地,报道分子),向延伸链内插入的标记可以为猝灭分子或报道分子中的一种(优选地,猝灭分子)。【(iv)利用两个信号寡核苷酸的相互作用性-双重标记】在利用两个信号寡核苷酸的相互作用性-双重标记的实例中,本发明的方法利用包含与延伸链互补的序列的追加的信号寡核苷酸,上述两个信号寡核苷酸以接近的方式与延伸链杂交,上述两个信号寡核苷酸分别包含相互作用性双重标记的报道分子及猝灭分子中的一种标记,上述两个信号寡核苷酸和延伸链之间的杂交诱导来自相互作用性双重标记的信号的变化,来提供可检测的信号。尤其,上述两个信号寡核苷酸中的至少一个包含与延伸反应中重新延伸的序列杂交的部位。实施利用上述两个信号寡核苷酸的相互作用性双重标记的实例的基本工作原理如下:与具有靶核苷酸变异的靶核酸序列杂交的探测和标记寡核苷酸-核苷酸变异所放出的第一片段与捕捉和模板化寡核苷酸的捕捉部位杂交并延伸来形成延伸链。之后,两个信号寡核苷酸与延伸链杂交。在上述杂交中,上述两个信号寡核苷酸以接近的方式与延伸链杂交,因此,上述两个信号寡核苷酸上的报道分子及猝灭分子相互接近,使得猝灭分子对来自报道分子的信号进行淬灭,最终引起来自相互作用性双重标记的信号变化(例如,来自报道分子的信号增加)。未参与上述杂交的两个信号寡核苷酸上的报道分子及猝灭分子相互隔开来从报道分子生成信号。根据一实例,只要通过两个信号寡核苷酸和延伸链的杂交使猝灭分子对来自报道分子的信号进行淬灭,上述两个信号寡核苷酸就可以与延伸链的任何位点杂交。优选地,上述两个信号寡核苷酸以直接接近的方式所处或隔开1-5核苷酸。根据一实例,在上述两个信号寡核苷酸接近延伸链来杂交的情况下,只要猝灭分子对来自报道分子的信号进行淬灭,报道分子及猝灭分子就可以与两个信号寡核苷酸的任何位点相连接。例如,报道分子或猝灭分子与一个信号寡核苷酸的5’-末端相连接或从其5’-末端隔开1-5核苷酸来连接,猝灭分子或报道分子与另一个信号寡核苷酸的3’-末端相连接或从其3’-末端隔开1-5核苷酸来连接。【(v)利用嵌入染料的荧光共振能量转移标记】根据本发明,可利用嵌入染料来形成荧光共振能量转移信号。根据一实例,信号寡核苷酸包含荧光共振能量转移的受体,并在步骤(e)杂交的嵌入染料存在的条件下实施,上述信号寡核苷酸和延伸链之间的杂交诱导来自信号寡核苷酸的上述受体的信号变化,来提供可检测的信号。实施利用嵌入染料的荧光共振能量转移标记的实例的基本工作原理如下:与具有靶核苷酸变异的靶核酸序列杂交的探测和标记寡核苷酸-核苷酸变异所放出的第一片段与捕捉和模板化寡核苷酸的捕捉部位杂交并延伸,来形成延伸链。之后,被受体标记的信号寡核苷酸与上述延伸链杂交来形成双重-链核酸分子后,嵌入染料与上述双重-链核酸分子相结合。由于用于激发(excitation)供体的照明(illumination)而起到供体作用的嵌入染料向受体引起能量转移,而这诱导来自受体的信号的变化,来提供可检测的信号。根据一实例,如上所述,与信号寡核苷酸相连接的受体包含多种单一荧光标记,但并不局限于此。有利于本发明的信号寡核苷酸包含可以提供对杂交具有依赖性的信号的所有探针,例如,MolecularbeaconTM(美国专利第5925517号)、HybeaconsTM(D.J.French,等.,MolecularandCellularProbes(2001)13,363–374及美国专利第7348141号)、双重-标记的自我淬灭探针(美国专利第5876930号)、LUXTM(I.A.Nazarenko,等.,NucleicAcidsRes2002,30:2089-2095.及美国专利第7537886号)及杂交探针(BernardPS,等.,ClinChem2000,46,147-148及DeeptiParashar等.,IndianJMedRes124,reviewarticleOctober2006385-398)。根据一实例,可使用提供能够进行实时检测的信号的标记来实时进行利用信号寡核苷酸的检测。择一性地,本发明所使用的标记可以提供可在杂交产物的解链期间内或在杂交产物的解链及杂交期间内检测的信号,因此,可在解链分析或杂交分析中实施利用信号寡核苷酸的检测。根据一实例,可利用探测和标记寡核苷酸-核苷酸变异片段和形成引物对的引物来追加扩增延伸链。根据一实例,信号寡核苷酸的3’-末端被阻断,以防止上述信号寡核苷酸的3’-末端的延伸。【利用杂交寡核苷酸的检测】根据一实例,可利用杂交寡核苷酸(hybridizingoligonucleotide)来检测第一片段的延伸链,上述杂交寡核苷酸包含与捕捉和模板化寡核苷酸互补的杂交核苷酸序列及至少一个标记,上述第一片段的延伸诱导由具有5’核酸酶活性的酶引起的杂交寡核苷酸的切割,来从上述标记中提供可检测的信号。根据一实例,上述杂交寡核苷酸位于捕捉和模板化寡核苷酸上的第一片段的下游。根据一实例,上述杂交寡核苷酸包含与捕捉和模板化寡核苷酸的模板化部位互补的杂交核苷酸序列。根据一实例,用于使上述片段延伸的模板-依赖性核酸聚合酶具有5’核酸酶活性。上述杂交寡核苷酸的长度可以多样。例如,杂交寡核苷酸的长度为5-100核苷酸、5-80核苷酸、5-60核苷酸、5-40核苷酸、5-20核苷酸、5-10核苷酸、10-100核苷酸、10-80核苷酸、10-60核苷酸、10-40核苷酸、10-30核苷酸、10-20核苷酸、15-100核苷酸、15-80核苷酸、15-60核苷酸、15-40核苷酸、15-30核苷酸、15-20核苷酸、20-100核苷酸、20-80核苷酸、20-60核苷酸、20-40核苷酸或20-30核苷酸。在本发明的根据一实例中,杂交寡核苷酸的3’-末端被阻断,以防止上述杂交寡核苷酸的3’-末端的延伸。简单地,能够以如下方式说明有利于本发明的标记系统。【(i)与杂交寡核苷酸相连接的单一标记】本发明可以提供利用具有单一标记的信号寡核苷酸来表示存在靶核苷酸变异的延伸链的形成相关的信号。在特定实例中,本说明书所使用的单一标记应可以根据其标记存在于双链还是单链(例如,杂交寡核苷酸及杂交寡核苷酸的片段)来提供不同的信号。根据一实例,需要在可进行杂交寡核苷酸以及捕捉和模板化寡核苷酸之间的杂交的温度中检测信号。【(ii)与杂交寡核苷酸相连接的相互作用性双重标记】根据一实例,由与杂交寡核苷酸相连接的相互作用性双重标记提供可检测的信号。如图7所示,从记寡核苷酸-核苷酸变异放出的第一片段与捉和模板化寡核苷酸的捕捉部位杂交,被包含灭分子的相互作用性双重标记标记的杂交寡核苷酸与捉和模板化寡核苷酸的模板化部位杂交。上述第一片段的延伸诱导上述杂交寡核苷酸的切割,使得报道分子从猝灭分子分离,由此提供表示延伸链的存在的信号。在这种实例中,在与双重标记-相连接的核苷酸相互以相对性的方式接近的情况下,可用于对杂交寡核苷酸切割之前和切割之后之间的信号变化进行信号检测。在进行双重标记-连接的核苷酸相对较远的情况下,杂交寡核苷酸和捕捉和模板化寡核苷酸之间的杂交诱导相互作用性双重标记在结构上相隔开,从而即使在杂交寡核苷酸未被切割的情况下,也不会对报道分子的信号进行淬灭,由此提供信号的变化。在这种情况下,可以在能够防止杂交寡核苷酸和捕捉和模板化寡核苷酸之间的杂交的更高的温度(例如,95℃)中检测来自杂交寡核苷酸的被切割的片段的信号。根据一实例,只要可以提供能够区别切割的杂交寡核苷酸及非切割的杂交寡核苷酸的信号,报道分子及猝灭分子可位于杂交寡核苷酸的任何位点。在特定实例中,报道分子及猝灭分子分别位于杂交寡核苷酸的两末端。【(iii)与杂交寡核苷酸及捕捉和模板化寡核苷酸相连接的相互作用性双重标记】根据一实例,通过报道分子及猝灭分子的相互作用性双重标记来提供可检测的信号,上述报道分子及猝灭分子中的一个与杂交寡核苷酸相连接,另一个与捕捉和模板化寡核苷酸相连接。在特定实例中,在杂交寡核苷酸与捕捉和模板化寡核苷酸杂交的情况下,报道分子及猝灭分子位于杂交寡核苷酸及捕捉和模板化寡核苷酸上,使得来自报道分子的信号被猝灭分子淬灭。通过第一片段的延伸来被诱导的杂交寡核苷酸的切割使杂交寡核苷酸从捕捉和模板化寡核苷酸放出,使报道分子从猝灭分子隔开,使得猝灭分子不对来自报道分子的信号进行淬灭,由此提供用于表示延伸链的存在的信号。根据一实例,有必要在可使杂交寡核苷酸以及捕捉和模板化寡核苷酸之间进行杂交的温度中检测信号。根据一实例,上述杂交寡核苷酸可以被设计成具有细销结构。在特定实例中,报道分子及猝灭分子在的一个与杂交寡核苷酸的3’-末端相连接,另一个与捕捉和模板化寡核苷酸的5’-末端相连接。根据一实例,可在利用杂交寡核苷酸的本发明的方法中利用使用两个杂交寡核苷酸的相互作用性-双重标记之类的标记系统。只要可以提供能够区别切割的杂交寡核苷酸及非切割的杂交寡核苷酸的信号,相互作用性-双重标记可位于两个杂交寡核苷酸的任何位点。可参照上述信号寡核苷酸的说明来记述标记的种类及位置。根据一实例,可在利用杂交寡核苷酸的本发明的方法中利用使用嵌入染料的荧光共振能量转移之类的标记系统。只要可以提供在嵌入染料条件下也能区别切割的杂交寡核苷酸及非切割的杂交寡核苷酸的信号,荧光共振能量转移标记可位于杂交寡核苷酸的任何位点。可参照上述信号寡核苷酸的说明来记述标记的种类及及位置。【基于延伸链的大小或序列的检测】根据一实例,可以基于延伸链的大小或序列检测第一片段的延伸链。例如,延伸链可以利用电泳或质量分析(例如,电子轰击(EI,electronimpact)、化学电离(CI,chemicalionization)、场解吸(FD,FieldDesoption)、252Cf-等离子解吸(PD,252Cf-Plasmadesoprtion)、解吸化学电离(DCI,desoprtionchemicalionization)、二次离子质谱(SIMS,secondaryionmassspectrometry)、快原子轰击(FAB,fastatombombardment)、电喷射离子化(ESI,electrosprayionization)、基质辅助激光解吸电离(MALDI,matrix-assistedlaserdesoprtionionization)及串联质谱(TandemMassSpectrometry)来测定。探测和标记寡核苷酸-核苷酸变异及捕捉和模板化寡核苷酸可由天然(naturallyoccurring)dNMPs组成。择一性地,探测和标记寡核苷酸以及捕捉和模板化寡核苷酸可包含变形核苷酸或非-天然核苷酸,例如肽核酸(PNA,PeptideNucleicAcid,参照PCT申请第WO92/20702号)及锁核酸(LNA,LockedNucleicAcid,参照PCT申请第WO98/22489号、第WO98/39352号以及第WO99/14226号)。探测和标记寡核苷酸以及捕捉和模板化寡核苷酸可包含如脱氧肌苷(deoxyinosine)、肌苷(inosine)、1-(2’-脱氧-β-D-呋喃核糖)-3-硝基吡咯(1-(2’-deoxybeta-D-ribofuranosyl)-3-nitropyrrole)以及5-硝基吲哚(5-nitroindole)之类的通用碱基。术语“通用碱基”意味着分别对天然脱氧核糖核酸/核糖核酸(RNA)碱基可以几乎没有区别地形成碱基对。根据一实例,上述方法在反复循环之间还包括变性过程,来反复实施上述步骤(a)~步骤(e)的全部或一部分。反应的反复伴随靶核酸序列的扩增。优选地,上述扩增根据在美国专利第4683195号、第4683202号以及第4800159号中记载的聚合酶链式反应(PCR,polymerasechainreaction)来实施。根据一实例,上述方法在反复循环之间还包括变性过程,来反复实施上述步骤(a)~步骤(b)、步骤(a)~步骤(d)或步骤(a)~步骤(f)。例如,上述方法可在反复循环之间包括变性过程,并使上述步骤(a)~步骤(b)、步骤(a)~步骤(d)或步骤(a)~步骤(e)反复实施数循环,例如,2-80循环、2-50循环、2-40循环、10-80循环、10-50循环、10-40循环、20-80循环、20-50循环、20-40循环、30-60循环或40-60循环后,实施上述步骤(e)。例如,上述可在反复循环之间包括变性过程,并使上述步骤(a)~步骤(b)反复实施数循环,例如,2-80循环、2-50循环、2-40循环、10-80循环、10-50循环、10-40循环、20-80循环、20-50循环、20-40循环、30-60循环或40-60循环后,实施上述步骤(c)~步骤(e)。可通过包含热、碱、甲酰胺、尿素及乙二醛处理、酶方法(如解旋酶作用)及结合蛋白质的公知的技术实施上述变性,但并不局限于此。例如,变性可在80~105℃温度范围内进行热处理来达成。在文献[JosephSambrook,等,MolecularCloning,ALaboratoryManual,ColdSpringHarborLaboratoryPress,ColdSpringHarbor,N.Y.(2001)]中公开了用于达成这种处理的常规方法。根据一实例,上述步骤(a)~步骤(e)在一个反应容器或单独的反应容器中实施。例如,可在单独的反应容器中实施步骤(a)~步骤(b)、步骤(c)~步骤(d)或步骤(e)。根据一实例,步骤(a)~步骤(e)可根据反应条件(尤其,温度)在一个反应容器中同时或单独实施。例如,上述步骤(a)~步骤(b)、步骤(c)~步骤(e)可根据反应条件(尤其,温度)在一个反应容器中同时或单独实施。根据一实例,利用包含上游引物和下游引物的引物对及扩增阻断剂的选择性的扩增以及利用探测和标记寡核苷酸-核苷酸变异的切割的靶核苷酸变异的检测可根据反应条件(尤其,温度)来在单独的反应容器中单独实施或在一个反应溶器中实施。在单独的反应容器中实施的情况下,探测和标记寡核苷酸-核苷酸变异可独立地被上游寡核苷酸切割或以依赖性的方式被上游探针切割。本发明并不需要所要进行检测和/或扩增的靶核酸序列具有某种特定的序列或长度,上述靶核酸序列包含所有脱氧核糖核酸(gDNA及cDNA)及核糖核酸分子。在利用mRNA作为初期物质的情况下,需要在实施退火步骤之前进行逆转录步骤,其相关详细内容公开于文献[JosephSambrook,等,MolecularCloning,ALaboratoryManual,ColdSpringHarborLaboratoryPress,ColdSpringHarbor,N.Y.(2001);以及Noonan,K.F.等,NucleicAcidsRes.16:10366(1988)]。为了实现逆转录反应,可利用与随机六聚体或mRNA杂交的寡核苷酸dT引物。可检测和/或扩增的靶核酸序列还都包含任何天然(naturallyoccurring)原核细胞核酸、真核细胞(例如,原生动物和寄生动物、菌类、酵母、高等植物、低等动物及包含哺乳动物和人类的高等动物)核酸、病毒(例如,疱疹病毒、人类免疫缺陷病毒(HIV,HumanImmunodeficiencyVirus)、流感病毒、EB病毒(Epstein-Barrvirus)、肝炎病毒以及脊髓灰质炎病毒等)核酸或类病毒核酸。可通过本发明来检测的靶核酸序列包含各种核酸序列,例如,基因组内序列、人为分离或片段化的序列及合成序列(例如,cDNA序列及条形码序列)。例如,靶核酸序列包含用于免疫PCR(IPCR,Immuno-PCR)的核酸标记序列。IPCR与PCR一同使用核酸标记序列及抗体间的接合体,这广泛适用于检测包含蛋白质的各种靶(Sano等,Science258pp:120-122(1992),美国专利第5665539号、Niemeyer等,TrendsinBiotechnology23pp:208-216(2005)、美国申请公开号第2005/0239108号及Ye等,JournalofEnvironmentalScience22pp:796-800(2010))。同时检测至少两种靶核酸序列(多重),这使本发明的优点更加突出。根据一实例,上述方法为了检测至少两种(更优选为至少3种,尤其优选为至少5种)靶核酸序列而实施,上述上游寡核苷酸包含至少两种(更优选为至少3种,尤其优选为至少5种)上游引物及下游引物,上述扩增阻断剂包含至少两种(更优选为至少3种,尤其优选为至少5种)扩增阻断剂,上述探测和标记寡核苷酸-核苷酸变异包含至少两种(更优选为至少3种,尤其优选为至少5种)探测和标记寡核苷酸-核苷酸变异。【利用在固相中实现固定化的寡核苷酸的核苷酸变异的检测】本发明在如微阵列之类的固相中也可以有效地检测靶核酸序列。根据一实例,本发明在固相中实施,寡核苷酸(例如,捕捉和模板化寡核苷酸、信号寡核苷酸或杂交寡核苷酸)通过其5’-末端或3’-末端来在固相底物上实现固定化。在固相中测定固相底物上的靶信号。可通过两种方法来实现上述捕捉和模板化寡核苷酸、信号寡核苷酸或杂交寡核苷酸的固定化。在第一方法中,已在固相底物上固定化的捕捉和模板化寡核苷酸、信号寡核苷酸或杂交寡核苷酸参与反应步骤。在第二方法中,捕捉和模板化寡核苷酸、信号寡核苷酸或杂交寡核苷酸以非-固定化的形态参与之后,在反应步骤中,在固相底物上实现固定化。根据一实例,在固相反应中,无需具有根据包含单一标记的核酸序列存在于单链内还是双链来使单一标记提供不同强度的信号的能力。单一标记包含化学标记(例如,生物素)、酶标记(例如,碱性磷酸酶、过氧化物酶、β-半乳糖苷酶及β-葡萄糖苷酶)、放射性同位素标记(例如,I125及C14)、荧光标记、发光标记、化学发光标记及金属标记(例如,金),但并不局限于此。为了实现固相反应,上述捕捉和模板化寡核苷酸、信号寡核苷酸或杂交寡核苷酸通过其5’-末端或3’-末端(优选为3’-末端)来在固相底物的表面直接或间接(优选为间接)实现固定化。并且,上述捕捉和模板化寡核苷酸、信号寡核苷酸或杂交寡核苷酸能够以共价键或非共价键方式在固相底物的表面实现固定化。实现固定化的寡核苷酸在固相底物的表面间接性地实现固定化的情况下,利用合适的连接肽。在本发明中有用的连接肽可包含利用于固相底物的表面的探针固定化的任何连接肽。例如,具有胺基管能性的烷基或芳基化合物或具有硫醇基的烷基或芳基化合物可作为连接肽使用于捕捉和模板化寡核苷酸的固定化。并且,可使用聚胸腺嘧啶尾(polyTtail)或聚腺苷酸尾(polyAtail)作为连接肽,这显著降低作为酶作用(例如,酶的切割反应)的抑制因素的空间妨碍因素,从而贡献于杂交效率的增加。不考虑使用聚胸腺嘧啶尾或聚腺苷酸尾作为探针的序列。根据一实例,上述捕捉和模板化寡核苷酸、信号寡核苷酸或杂交寡核苷酸可通过结合伙伴(例如,生物素/链霉亲和素)之间的相互作用来在固相底物上实现固定化。例如,具有结合伙伴(生物素及链霉亲和素)中的一个的捕捉和模板化寡核苷酸、信号寡核苷酸或杂交寡核苷酸可在变形为表面不同的结合伙伴的固相底物上实现固定化。根据一实例,上述捕捉和模板化寡核苷酸、信号寡核苷酸或杂交寡核苷酸可通过用于实现固定化的核苷酸序列来在固相底物上实现固定化。例如,为了在用于固定化的核苷酸序列固定具有互补的追加序列的捕捉和模板化寡核苷酸、信号寡核苷酸或杂交寡核苷酸,可以使用表面变形为用于固定化的核苷酸序列的固相底物。根据一实例,在本发明中利用的固相底物为微阵列。在本发明中提供反应环境的微阵列可以使用本发明所属的技术领域中公知的任何微阵列。本发明的所有过程,即与靶核酸序列的杂交、切割、延伸、解链以及荧光的检测在微阵列实施。在微阵列实现固定化的捕捉和模板化寡核苷酸利用为杂交阵列因素(hybridizablearrayelement)。用于制备微阵列的固相底物包括金属(例如,金、金与铜的合金、铝)、金属氧化物、玻璃、陶瓷、石英、硅、半导体、Si/SiO2晶圆、锗、砷化镓、碳、碳纳米管、聚合物(例如,聚苯乙烯、聚乙烯、聚丙烯以及聚丙烯酰胺)、琼脂糖凝胶(sepharose)、琼脂糖(agarose)及胶体(colloids),但并不局限于此。上述固相底物可以为试纸、板、颗粒(例如,珠)亲和性柱及膜(membrane)的形态。本发明的实现固定化的多个寡核苷酸可在固相底物的一个寻址(addressable)部位或多个寻址部位实现固定化,固相底物可包括2~1000000个寻址部位。可借助如光刻法、喷墨打印法、机械点样法及与其类似的方法之类的现有制备技术,构建(fabricate)为了生产阵列或用于特定应用的阵列而实现固定化的寡核苷酸。由于实现固定化的寡核苷酸上的标记以物理方式相互隔开,因此,可在固相中实施的本发明即使利用一种标记也可以同时检测多个靶核酸序列。因此,在固相中,可根据本发明来检测的靶核酸序列的数量不受限制。可利用共聚点检测设备来与残留于液相的标记无关地只检测固相底物上的信号。【用于检测靶核苷酸变异的试剂盒】在本发明的另一实施方式中,本发明提供利用扩增阻断剂及基于探测和标记寡核苷酸切割及延伸的变异检测分析来检测靶核酸序列上的靶核苷酸变异的试剂盒,其特征在于,包含:(a)包含用于扩增靶核酸的上游引物及下游引物的引物对,上述上游引物及下游引物分别包含与靶核酸序列互补的杂交核苷酸序列;(b)对5’核酸酶切割具有耐性的扩增阻断剂,上述扩增阻断剂包含与不同于上述靶核酸序列上的上述靶核苷酸变异的其他非-靶核苷酸变异互补的序列;(c)探测和标记寡核苷酸-核苷酸变异,上述探测和标记寡核苷酸-核苷酸变异包含:(i)3’-靶向部位,包含与上述靶核酸序列互补的杂交核苷酸序列,(ii)5’-标记部位,包含与上述靶核酸序列非-互补的核苷酸序列,以及(iii)核苷酸变异区别位点,包含与上述靶核酸上的上述靶核苷酸变异互补的序列,并位于上述3’-靶向部位的5’-末端一部分;(d)捕捉和模板化寡核苷酸,上述捕捉和模板化寡核苷酸沿着3’→5’方向包含:(i)捕捉部位,包含与上述探测和标记寡核苷酸-核苷酸变异的5’-标记部位或5’-标记部位的一部分互补的核苷酸序列,以及(ii)模板化部位,包含与上述探测和标记寡核苷酸-核苷酸变异的5’-标记部位及3’-靶向部位非-互补的核苷酸序列,从上述探测和标记寡核苷酸-核苷酸变异放出的第一片段或第二片段与上述捕捉和模板化寡核苷酸的捕捉部位杂交,上述扩增阻断剂与具有上述非-靶核苷酸变异的靶核酸序列杂交,不与具有上述靶核苷酸变异的靶核酸序列杂交,上述探测和标记寡核苷酸-核苷酸变异的3’-靶向部位与上述靶核酸序列杂交,上述探测和标记寡核苷酸-核苷酸变异的5’-标记部位不与上述靶核酸序列杂交,上述上游引物位于上述探测和标记寡核苷酸-核苷酸变异的上游,上述扩增阻断剂位于上述上游引物或下游引物的下游,上述扩增阻断剂及上述探测和标记寡核苷酸-核苷酸变异位于上述上游引物和下游引物之间,上述上游引物通过其延伸链来诱导基于上述具有5’核酸酶活性的酶的上述探测和标记寡核苷酸-核苷酸变异的切割,上述扩增阻断剂和具有上述非-靶核苷酸变异的靶核酸序列的杂交抑制位于上述扩增阻断剂的上游的上述引物的延伸,由此阻断具有上述非-靶核苷酸变异的靶核酸序列的扩增,若上述探测和标记寡核苷酸-核苷酸变异与具有对上述核苷酸变异区别位点互补的上述靶核苷酸变异的上述靶核酸序列杂交,则上述3’-靶向部位的5’-末端一部分与上述靶核酸序列形成双链,来从第一初期切割位点诱导切割,并放出第一片段,若上述探测和标记寡核苷酸-核苷酸变异与具有对上述核苷酸变异区别位点非-互补的上述非-靶核苷酸变异的上述靶核酸序列杂交,则上述3’-靶向部位的5’-末端一部分不与上述靶核酸序列形成双链,而从位于上述第一初期位点的下游的第二初期切割位点诱导切割,并放出第二片段,上述第二片段包含追加的3’-末端部位,用于使上述第二片段与上述第一片段不同;若上述第一片段与上述捕捉和模板化寡核苷酸的捕捉部位杂交,则上述第一片段延伸并形成与上述捕捉和模板化寡核苷酸的模板化部位互补的延伸序列的延伸链,若上述第二片段与上述捕捉和模板化寡核苷酸的捕捉部位杂交,则上述第二片段不延伸。本发明的试剂盒以用于实施上述本发明的检测方法的方式构成,因此,为了避免有可能导致本说明书的复杂性的过度重复而省略它们之间的共同的内容。根据一实例,上述试剂盒还包含上述具有5’核酸酶活性的酶、模板-依赖性核酸聚合酶或它们的组合。根据一实例,上述扩增阻断剂包含核苷/核苷酸,实施核苷/核苷酸包含对5’核酸酶活性具有耐性的主链(backbone)。根据一实例,上述扩增阻断剂包含肽核酸、锁核酸、吗啉、甘油核酸、苏糖核酸、桥核酸、N3’-P5’氨基磷酸酯(NP)低聚物、连接有小沟结合物的寡核苷酸、硫代磷酸酯低聚物、C1-C4膦酸烷基酯低聚物、氨基磷酸酯、β-磷酸二酯寡核苷酸、α-磷酸二酯寡核苷酸或它们的组合。根据一实例,在上述第二片段与上述捕捉和模板化寡核苷酸的捕捉部位杂交的情况下,由于上述捕捉和模板化寡核苷酸不与上述第二片段的上述追加的3’-末端部位杂交,因此,上述捕捉和模板化寡核苷酸具有以防止上述第二片段的延伸的方式选择的序列。根据一实例,上述核苷酸变异区别位点与上述探测和标记寡核苷酸-核苷酸变异的3’-靶向部位的5’-末端隔开10核苷酸以内。根据一实例,上述探测和标记寡核苷酸-核苷酸变异的3’-靶向部位的5’-末端一部分包含与上述核苷酸变异区别位点隔开1核苷酸至5核苷酸以内的非-碱基对部分(moiety),上述非-碱基对部分增加上述第一初期切割位点及上述第二初期切割位点之间的差异。根据一实例,上述非-碱基对部分为:(i)包含人为错配碱基、变形为无法碱基配对的非-碱基对碱基或通用碱基的核苷酸;(ii)变形为无法碱基配对的非-碱基对核苷酸;或(iii)非-碱基配对化合物(non-basepairingchemicalcompound)。根据一实例,上述核苷酸变异为置换变异、缺失变异或插入变异。根据一实例,上述第一片段和上述捕捉和模板化寡核苷酸在形成延伸二聚物,上述延伸二聚物具有能够基于如下条件而调节的Tm值:(i)上述第一片段的序列和/或长度,(ii)上述捕捉和模板化寡核苷酸的序列和/或长度或(iii)上述第一片段的序列和/或长度及上述捕捉和模板化寡核苷酸的序列和/或长度,上述延伸二聚物提供基于如下标记的靶信号:(i)与上述第一片段和/或上述捕捉和模板化寡核苷酸相连接的至少一个标记,(ii)在上述延伸反应期间向上述延伸二聚物内插入的标记,(iii)与上述第一片段和/或上述捕捉和模板化寡核苷酸相连接的至少一个标记及在上述延伸反应期间向上述延伸二聚物内插入的标记或(iv)嵌入标记,根据对于上述延伸二聚物的解链分析或杂交分析,从上述延伸二聚物测定上述靶信号,来检测上述延伸链的存在。根据一实例,上述第一片段的延伸链和上述捕捉和模板化寡核苷酸形成延伸二聚物,上述延伸二聚物具有能够基于如下条件而调节的Tm值:(i)上述第一片段的序列和/或长度,(ii)上述捕捉和模板化寡核苷酸的序列和/或长度或(iii)上述第一片段的序列和/或长度和上述捕捉和模板化寡核苷酸的序列和/或长度,上述延伸二聚物提供基于如下标记的靶信号:(i)与上述第一片段和/或捕捉和模板化寡核苷酸相连接的至少一个标记,(ii)在上述延伸反应期间向上述延伸二聚物内插入的标记,(iii)与上述第一片段和/或捕捉和模板化寡核苷酸相连接的至少一个标记及在上述延伸反应期间向上述延伸二聚物内插入的标记或(iv)嵌入标记,在上述延伸二聚物维持双链所需的充分的指定温度下,从上述延伸二聚物测定靶信号,来检测上述延伸链的存在。根据一实例,上述试剂盒还包含用于检测上述第一片段的延伸链的信号寡核苷酸,上述信号寡核苷酸包含与上述延伸链互补的序列及至少一个标记,上述信号寡核苷酸提供能够通过与上述延伸链之间的结合或解离来检测的信号。根据一实例,通过如下组合来提供上述可检测的信号:(i)与上述信号寡核苷酸相连接的标记;(ii)与上述信号寡核苷酸相连接的标记及与上述探测和标记寡核苷酸的片段相连接的标记的组合;(iii)与信号寡核苷酸相连接的标记及在延伸反应期间内插入于延伸链的标记的组合;或者(iv)与信号寡核苷酸相连接的标记及嵌入染料。根据一实例,上述试剂盒还包含用于检测第一片段的延伸链的杂交寡核苷酸(HO,hybridizingoligonucleotide),上述杂交寡核苷酸包含与上述捕捉和模板化寡核苷酸互补的杂交核苷酸序列及至少一个标记,上述第一片段的延伸诱导由具有5’核酸酶活性的酶引起的上述杂交寡核苷酸的切割,从而从上述标记生成可检测的信号。根据一实例,通过如下相互作用性双重标记来提供上述可检测的信号:(i)与上述杂交寡核苷酸相连接的相互作用性双重标记;或者(ii)相互作用性双重标记中的一个与上述杂交寡核苷酸相连接而另一个与上述捕捉和模板化寡核苷酸相连接的包含报道分子及猝灭分子的相互作用性双重标记。根据一实例,上述扩增阻断剂、探测和标记寡核苷酸-核苷酸变异、捕捉和模板化寡核苷酸、信号寡核苷酸和/或杂交寡核苷酸的3’-末端被阻断,以防止上述扩增阻断剂、探测和标记寡核苷酸-核苷酸变异、捕捉和模板化寡核苷酸、信号寡核苷酸和/或杂交寡核苷酸的3’-末端延伸。根据一实例,上述试剂盒用于检测至少两种核苷酸变异,上述上游引物及上述下游引物包含至少两种上游引物及下游引物,上述扩增阻断剂包含至少两种扩增阻断剂,上述探测和标记寡核苷酸-核苷酸变异包含至少两种探测和标记寡核苷酸-核苷酸变异。根据一实例,上述模板-依赖性核酸聚合酶为与上述具有5’核酸酶活性的酶相同的酶。根据一实例,上述具有5’核酸酶活性的酶为具有5’核酸酶活性的热稳定性脱氧核糖核酸聚合酶或瓣状核酸内切酶。上述本发明的所有试剂盒可以包含缓冲液、脱氧核糖核酸聚合酶辅助因子及脱氧核糖核苷酸-5-三磷酸等实施靶扩增聚合酶链式反应(例如,聚合酶链式反应)所需的多种试剂。选择性地,上述试剂盒还可以包含多种多核苷酸分子、逆转录酶、多种缓冲液、试剂及用于抑制脱氧核糖核酸聚合酶活性的抗体。并且上述试剂盒可以包含用于实施阳性对照组及阴性对照组反应所需的试剂。本发明所属技术领域的普通技术人员可容易地决定在特定反应中所使用的试剂的最佳量。典型地,本发明的试剂盒以在额外的包装及划分内包含上述组成成分的方式采用。简要说明本发明的特征及优点如下:(a)本发明为对本发明人所开发的基于探测和标记寡核苷酸切割及延伸的变异检测分析进行改善的内容,少本发明的目的在于,有效检测少量的等位基因。(b)本发明非常有利于在过量的野生型脱氧核糖核酸内检测少数突变。扩增阻断剂可通过突变脱氧核糖核酸的选择性的扩增来限制对野生型脱氧核糖核酸的探测和标记寡核苷酸-核苷酸变异的消耗。或者,扩增阻断剂可在对野生型脱氧核糖核酸的杂交中与探测和标记寡核苷酸-核苷酸变异竞争,而这可以在野生型脱氧核糖核酸上防止探测和标记寡核苷酸-核苷酸变异的切割。(c)根据本发明,可以看出探针(PTO-NV)根据是否存在受关注的核苷酸变异来相区别的不同的杂交模式。(d)对受关注的核苷酸变异这种区别的杂交模式提供探测和标记寡核苷酸-核苷酸变异的初期切割位点的差异,由此,生成两种探测和标记寡核苷酸-核苷酸变异片段,从而根据是否存在受关注核苷酸变异来发生信号差异。(e)值得关注的是,可以无需考虑靶核酸序列而选择探测和标记寡核苷酸-核苷酸变异的5’-标记部位序列及捕捉和模板化寡核苷酸的序列。由此可预先设计用于探测和标记寡核苷酸-核苷酸变异的5’-标记部位及捕捉和模板化寡核苷酸的序列的库(pool)。探测和标记寡核苷酸-核苷酸变异的3’-靶向部位需考虑靶核酸序列来制备,但捕捉和模板化寡核苷酸无需考虑靶核酸序列的信息或靶核酸序列,能够以现成的方法制备。这种特征在多重靶检测,尤其,利用在固相底物实现固定化的捕捉和模板化寡核苷酸的微阵列提供显著的优点。以下,通过实施例对本发明进行更为详细的说明。这些实施例仅用于更加具体地说明本发明,根据本发明的要旨,本发明的范围不局限于这些实施例,这对于本发明所属技术领域的普通技术人员来说是显而易见的。【实施例】【实施例1:基于利用扩增阻断剂的基于探测和标记寡核苷酸切割及延伸的变异检测分析的少数突变的检测】通过实验来确认是否可以通过扩增阻断剂和基于探测和标记寡核苷酸切割及延伸的变异检测的组合来确认过量的野生型脱氧核糖核酸内的少数突变。为了上游引物和下游引物的延伸、探测和标记寡核苷酸-核苷酸变异切割及探测和标记寡核苷酸-核苷酸变异片段的延伸而使用具有5’核酸酶活性的TaqDNA聚合酶。探测和标记寡核苷酸-核苷酸变异、扩增阻断剂及捕捉和模板化寡核苷酸的3’-末端被碳间隔区(carbonspacer)阻断,以防止上述探测和标记寡核苷酸-核苷酸变异、扩增阻断剂及捕捉和模板化寡核苷酸的3’-末端的延伸。使用BRAF(V600E)野生型(T)及突变(A)人类基因组脱氧核糖核酸作为靶核酸序列。突变脱氧核糖核酸与具有靶核苷酸变异的靶核酸序列相对应,野生型脱氧核糖核酸与具有非-靶核苷酸变异的靶核酸序列相对应。为了调查扩增阻断剂的效果,准备了具有不同比率的突变及野生型BRAF脱氧核糖核酸一系列的混合物(突变100%、突变10%、突变1%、突变0.1%及突变0%)。探测和标记寡核苷酸-核苷酸变异不具有标记。探测和标记寡核苷酸-核苷酸变异的核苷酸变异区别位点具有与感链(sense-strand)的突变(A)脱氧核糖核酸互补核苷酸(T)(序列号:3)。扩增阻断剂包含锁核酸核苷酸,扩增阻断剂的核苷酸变异区别位点具有与反义链(anti-sensestrand)的野生型(A)脱氧核糖核酸互补的核苷酸(T)(序列号:4)。在本实施例的基于探测和标记寡核苷酸切割及延伸的变异检测分析中,随着靶核苷酸变异(即,突变脱氧核糖核酸)的存在来生成的延伸链的存在通过由延伸链和捕捉和模板化寡核苷酸形成的延伸二聚物的解链分析来检测。捕捉和模板化寡核苷酸利用猝灭分子(BHQ-2)及荧光报道分子(CalFluorRed610)来标记其模板化(templating)部位(序列号:5)。本实施例所使用的上游引物、下游引物、探测和标记寡核苷酸-核苷酸变异、扩增阻断剂及捕捉和模板化寡核苷酸的序列如下:BRAF-F5’-CTTCATAATGCTTGCTCTGATAGGIIIIIGAGATCTACT-3’(序列号:1)BRAF-R5’-ATAGCCTCAATTCTTACCATCCAIIIIITGGATCCAGA-3’(序列号:2)BRAF-PTO-NV5’-GGTGGACTTGCGGCTGTAGCTAGACCAAAATCACCTATTTTTACTGTG[C3间隔区]-3’(序列号:3)扩增阻断剂5’-TGATGG[C3间隔区]-3’(序列号:4)BRAF-CTO-15’-[BHQ-2]TTTTTTTTGAGCCAGAGTTA[T(CalFluorRed610)]GGTCACCGCAAGTCCACC[C3间隔区]-3’(序列号:5)(I:脱氧肌苷)(下划线的文字表示探测和标记寡核苷酸-核苷酸变异的5’-标记部位)(粗体字表示核苷酸区别位点)(方框内的文字表示锁核酸核苷酸)利用含有BRAF(V600E)突变(A)及野生型(T)人类基因组脱氧核糖核酸的多种比率的混合物(突变100%、突变10%、突变1%、突变0.1%及突变0%)100ng、上游引物(序列号:1)10pmole、下游引物(序列号:2)10pmole、探测和标记寡核苷酸-核苷酸变异(序列号:3)5pmole、扩增阻断剂(序列号:4)5pmole、片段与捕捉和模板化寡核苷酸(序列号:5)1pmole及2X主混合液(2.5mM的MgCl2、200μM的dNTPs、1.6单位的TaqDNA聚合酶)(韩国索尔杰恩特公司(Solgent,Korea))10μl的20μl的最终体积来实施反应。使含有上述反应混合物的管位于实时热循环仪(CFX96,Bio-Rad)。使上述反应混合物在95℃温度下进行15分钟变性后,在95℃温度下30秒钟、在60℃温度下60秒钟过程反复了50次。反应之后,使上述反应混合物冷却至55℃,并在55℃温度下维持10秒钟后,从55℃慢慢加热至85℃,由此得到解链曲线。在温度上升的过程中,连续测定荧光来监测双链脱氧核糖核酸的解离。解链峰来源于解链曲线数据。如图8A及图8B所示,在存在靶核酸序列的情况下,若没有扩增阻断剂,则在基于探测和标记寡核苷酸切割及延伸的变异检测分析中,到10%突变比率为止检测出与延伸二聚物的预计Tm值相对应的峰,但在具有扩增阻断剂的情况下,到0.1%突变比率为止检测出峰。在不存在目标的情况下,未检测出任何峰。这种结果表示使用扩增阻断剂来改善基于探测和标记寡核苷酸切割及延伸的变异检测分析的能力,从而即使存在过量的野生型脱氧核糖核酸的情况下,也可以检测少数突变。【实施例2:与扩增阻断剂一同利用信号寡核苷酸的基于探测和标记寡核苷酸切割及延伸的变异检测分析的少数突变的检测】本发明人还实验了是否可以通过利用信号寡核苷酸的基于探测和标记寡核苷酸切割及延伸的变异检测和扩增阻断剂的组合来确认在过量的野生型脱氧核糖核酸内的少数突变。在本实施例的基于探测和标记寡核苷酸切割及延伸的变异检测分析中,利用与上述延伸链特异性地杂交的信号寡核苷酸来检测了随着靶核苷酸变异(即,突变脱氧核糖核酸)的存在生成的延伸链的存在。为了解链分析而使用了上述延伸链以及上述信号寡核苷酸之间的杂交物。以与实施例1相同的方式使用了引物、扩增阻断剂、探测和标记寡核苷酸-核苷酸变异、BRAF人类基因组脱氧核糖核酸及TaqDNA聚合酶。捕捉和模板化寡核苷酸不具有标记(序列号:6)。信号寡核苷酸在其5’-末端具有猝灭分子(BHQ-2),在3’-末端具有荧光报道分子(CalFluorRed610)(序列号:7)。本实施例所使用的上游引物、下游引物、探测和标记寡核苷酸-核苷酸变异、扩增阻断剂、捕捉和模板化寡核苷酸及信号寡核苷酸的序列如下:BRAF-F5’-CTTCATAATGCTTGCTCTGATAGGIIIIIGAGATCTACT-3’(序列号:1)BRAF-R5’-ATAGCCTCAATTCTTACCATCCAIIIIITGGATCCAGA-3’(序列号:2)BRAF-PTO-NV5’-GGTGGACTTGCGGCTGTAGCTAGACCAAAATCACCTATTTTTACTGTG[C3间隔区]-3’(序列号:3)扩增阻断剂5’-TGATGG[C3间隔区]-3’(序列号:4)BRAF-CTO-25’-TTTTTTTTGAGCCAGAGTTATGGTCACCGCAAGTCCACC[C3间隔区]-3’(序列号:6)BRAF-SO5’-[BHQ-2]TTTTTTTGAGCCAGAGTTATGGTC[CalFluorRed610]-3’(序列号:7)(I:脱氧肌苷)(下划线的文字表示探测和标记寡核苷酸-核苷酸变异的5’-标记部位)(粗体字表示核苷酸区别位点)(方框内的文字表示锁核酸核苷酸)利用含有BRAF(V600E)突变(A)及野生型(T)人类基因组脱氧核糖核酸的多种比率的混合物(突变100%、突变10%、突变1%、突变0.1%及突变0%)100ng、上游引物(序列号:1)10pmole、下游引物(序列号:2)10pmole、探测和标记寡核苷酸-核苷酸变异(序列号:3)5pmole、扩增阻断剂(序列号:4)5pmole、捕捉和模板化寡核苷酸(序列号:6)0.1pmole、信号寡核苷酸(序列号:7)3pmole及2X主混合液(2.5mM的MgCl2、200μM的dNTPs、1.6单位的TaqDNA聚合酶)(韩国索尔杰恩特公司)10μl的20μl的最终体积来实施反应,使含有上述反应混合物的管位于实时热循环仪(CFX96,Bio-Rad)。使上述反应混合物在95℃温度下进行15分钟变性后,在95℃温度下30秒钟、在60℃温度下60秒钟过程反复了50次。反应之后,使上述反应混合物冷却至40℃,并在40℃温度下维持10秒钟后,从40℃慢慢加热至85℃,由此得到解链曲线。在温度上升的过程中,连续测定荧光来监测双链脱氧核糖核酸的解离。解链峰来源于解链曲线数据。如图9A及图9B所示,在存在靶核酸序列的情况下,以无扩增阻断剂的方式利用信号寡核苷酸的基于探测和标记寡核苷酸切割及延伸的变异检测分析中,在100%突变比率中检测出与延伸链/信号寡核苷酸杂交物的预计Tm值相对应的峰,但在10%突变比率中未检测出峰。然而,在具有扩增阻断剂的情况下,至1%突变比率为止均检测出峰。在不存在目标的情况下,未检测出任何峰。这种结果表示使用扩增阻断剂来改善利用信号寡核苷酸的基于探测和标记寡核苷酸切割及延伸的变异检测分析的能力,从而即使存在过量的野生型脱氧核糖核酸的情况下,也可以检测少数突变。【实施例3:与扩增阻断剂一同利用杂交寡核苷酸的基于探测和标记寡核苷酸切割及延伸的变异检测分析的少数突变的检测】本发明人还实验了是否可以通过利用杂交寡核苷酸的基于探测和标记寡核苷酸切割及延伸的变异检测和扩增阻断剂的组合来确认在过量的野生型脱氧核糖核酸内的少数突变。在本实施例的基于探测和标记寡核苷酸切割及延伸的变异检测分析中,使用与捕捉和模板化寡核苷酸特异性地杂交的杂交寡核苷酸来检测了随着靶核苷酸变异(即,突变脱氧核糖核酸)的存在来生成的延伸链的存在,上述杂交寡核苷酸位于比探测和标记寡核苷酸-核苷酸变异片段更为下游的位置。探测和标记寡核苷酸-核苷酸变异片段在捕捉和模板化寡核苷酸上延伸的过程中,杂交寡核苷酸被切割,并提供信号。在各循环中,通过预先指定的温度中的实时检测来检测了基于杂交寡核苷酸的切割来发生的信号。以与实施例1相同的方式使用了引物、扩增阻断剂、探测和标记寡核苷酸-核苷酸变异、BRAF人类基因组脱氧核糖核酸及TaqDNA聚合酶。捕捉和模板化寡核苷酸不具有标记(序列号:8)。杂交寡核苷酸在其5’-末端具有猝灭分子(BHQ-2),在3’-末端具有荧光报道分子(CalFluorRed610)(序列号:9)。本实施例所使用的上游引物、下游引物、探测和标记寡核苷酸-核苷酸变异、扩增阻断剂、捕捉和模板化寡核苷酸的序列如下:BRAF-F5’-CTTCATAATGCTTGCTCTGATAGGIIIIIGAGATCTACT-3’(序列号:1)BRAF-R5’-ATAGCCTCAATTCTTACCATCCAIIIIITGGATCCAGA-3’(序列号:2)BRAF-PTO-NV5’-GGTGGACTTGCGGCTGTAGCTAGACCAAAATCACCTATTTTTACTGTG[C3间隔区]-3’(序列号:3)扩增阻断剂5’-TGATGG[C3间隔区]-3’(序列号:4)BRAF-CTO-35’-TCCGTCCGAGCCAGAGTGATGGTCACCTCACCGCAAGTCCACC[C3间隔区]-3’(序列号:8)BRAF-HO5’-[BHQ-2]GACCATCACTCTGGCTCGGACGGA[CalFluorRed610]-3’(序列号:9)(I:脱氧肌苷)(下划线的文字表示探测和标记寡核苷酸-核苷酸变异的5’-标记部位)(粗体字表示核苷酸区别位点)(方框内的文字表示锁核酸核苷酸)利用含有BRAF(V600E)突变(A)及野生型(T)人类基因组脱氧核糖核酸的多种比率的混合物(突变100%、突变10%、突变1%、突变0.1%及突变0%)100ng、上游引物(序列号:1)10pmole、下游引物(序列号:2)10pmole、探测和标记寡核苷酸-核苷酸变异(序列号:3)5pmole、扩增阻断剂(序列号:4)5pmole、捕捉和模板化寡核苷酸(序列号:8)1pmole、杂交寡核苷酸(序列号:9)3pmole及2X主混合液(2.5mM的MgCl2、200μM的dNTPs、1.6单位的HTaqDNA聚合酶)(韩国索尔杰恩特公司)10μl的20μl的最终体积来实施反应。将含有上述反应混合物的管放入实时热循环仪(CFX96,Bio-Rad)。使上述反应混合物在95℃温度下进行15分钟变性后,在95℃温度下30秒钟、在55℃温度下60秒钟过程反复了50次。在各循环的变性化步骤(95℃)中检测了所生成的信号。变性化温度(95℃)中的检测支持检测到的信号来自通过杂交寡核苷酸的切割来发生的被标记的片段。如图10A及图10B所示,在存在靶核酸序列的情况下,以无扩增阻断剂的方式利用杂交寡核苷酸的基于探测和标记寡核苷酸切割及延伸的变异检测分析中,到10%突变比率为止检测出荧光信号,但在具有扩增阻断剂的情况下,到0.1%突变比率为止检测出峰。在不存在目标的情况下,未检测出任何峰。这种结果表示,使用扩增阻断剂,来改善利用杂交寡核苷酸的基于探测和标记寡核苷酸切割及延伸的变异检测分析的能力,从而即使在存在过量的野生型脱氧核糖核酸的情况下,也可以检测少数突变。以上,详细记述了本发明优选一实例,对于本发明所属技术领域的普通技术人员来说,能够进行根据本发明的原理的变形及修改是显而易见的,且本发明的范围根据所附的发明要求保护范围及与其等同的技术方案来定义。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1