大豆GmCML8蛋白及其编码基因与应用的制作方法
本发明涉及一种大豆gmcml8蛋白及其编码基因与应用。
背景技术:
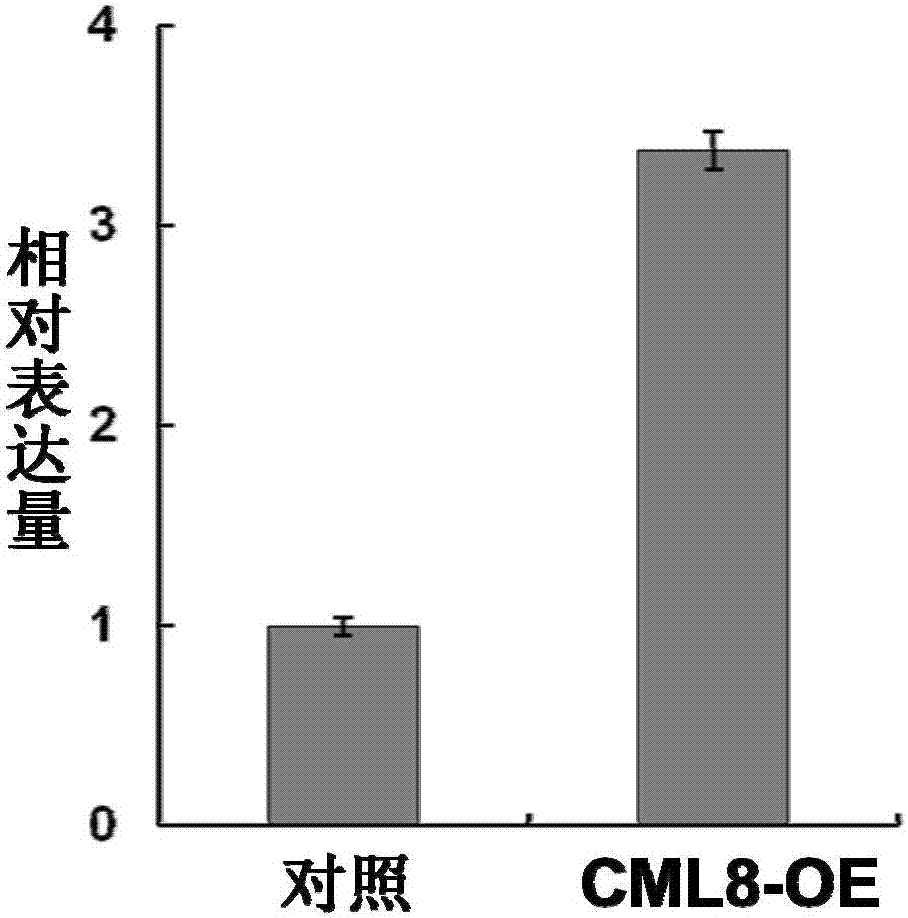
:环境中物理、化学因素的变化,例如干旱、盐碱、冷害、冻害、水涝等胁迫因素以及生物因素,例如病虫害对植物的生长发育具有重要的影响,严重时会造成农作物大规模减产。盐害是农业生产上重要的逆境危害之一,中国盐渍土面积大,分布广泛,类型多样,而且每年盐碱化和次生盐碱化都在不断加重,使农业生产的可持续发展受到威胁。培育耐逆性作物是种植业的主要目标之一,提高作物的耐逆性,除了利用传统的育种方法,目前,分子遗传育种已经成为科技工作者所关注的领域之一。技术实现要素:本发明的目的是提供一种大豆gmcml8蛋白及其编码基因与应用。本发明提供的蛋白质,获自大豆,命名为gmcml8蛋白,是如下(a)或(b):(a)由序列表中序列1所示的氨基酸序列组成的蛋白质;(b)将序列1的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且与植物耐盐性相关的由序列1衍生的蛋白质。序列表中的序列1由150个氨基酸残基组成。为了使(a)中的gmcml8蛋白便于纯化和检测,可在由序列表中序列1所示的氨基酸序列组成的蛋白质的氨基末端或羧基末端连接上如表1所示的标签。表1标签的序列标签残基序列poly-arg5-6(通常为5个)rrrrrpoly-his2-10(通常为6个)hhhhhhflag8dykddddkstrep-tagii8wshpqfekc-myc10eqkliseedl上述(b)中的gmcml8蛋白可人工合成,也可先合成其编码基因,再进行生物表达得到。上述(b)中的gmcml8蛋白的编码基因可通过将序列表中序列2所示的dna序列中缺失一个或几个氨基酸残基的密码子,和/或进行一个或几个碱基对的错义突变,和/或在其5′端和/或3′端连上表1所示的标签的编码序列得到。编码所述gmcml8蛋白的基因(gmcml8基因)也属于本发明的保护范围。所述基因具体可为如下(1)或(2)或(3)的dna分子:(1)编码区如序列表中序列2所示的dna分子;(2)在严格条件下与(1)限定的dna序列杂交且编码与植物耐盐性相关的蛋白质的dna分子;(3)与(1)或(2)限定的dna序列至少具有70%、至少具有75%、至少具有80%、至少具有85%、至少具有90%、至少具有95%、至少具有96%、至少具有97%、至少具有98%或至少具有99%同源性且编码植物耐盐性相关蛋白的dna分子。上述严格条件可为用0.1×sspe(或0.1×ssc),0.1%sds的溶液,在dna或者rna杂交实验中65℃下杂交并洗膜。含有所述gmcml8基因的重组表达载体、表达盒、转基因细胞系或重组菌均属于本发明的保护范围。可用现有的植物表达载体构建含有gmcml8基因的重组表达载体。所述植物表达载体包括双元农杆菌载体和可用于植物微弹轰击的载体等。使用gmcml8基因构建重组表达载体时,可在其转录起始核苷酸前加上任何一种增强型、组成型、组织特异型或诱导型启动子,它们可单独使用或与其它的植物启动子结合使用;此外,使用gmcml8基因构建重组表达载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是atg起始密码子或邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的,也可以是合成的。翻译起始区域可以来自转录起始区域或结构基因。为了便于对转基因植物细胞或植物进行鉴定及筛选,可对所用植物表达载体进行加工,如加入在植物中表达可产生颜色变化的酶或发光化合物的基因、具有抗性的抗生素标记物或是抗化学试剂标记基因等。从转基因植物的安全性考虑,也可不加入任何筛选基因,直接通过逆境胁迫进行筛选。所述重组表达载体具体可为将植物表达载体prokii的bamhi和kpni酶切位点之间的小片段取代为了序列表的序列2的自5′端第1-453位核苷酸所示的dna分子。扩增所述基因全长或其任意片段的引物对也属于本发明的保护范围。所述引物对具体可由序列表的序列3所示的单链dna分子和序列表的序列4所示的单链dna分子组成。本发明还保护所述gmcml8蛋白或其编码基因在调控植物耐盐性中的应用。所述植物为单子叶植物或双子叶植物。所述双子叶植物可为豆目植物。所述豆目植物可为豆科植物。所述豆科植物可为大豆属植物。所述大豆属植物具体可为大豆,例如大豆科丰1号。所述gmcml8蛋白或其编码基因在培育耐盐植物中的应用也属于本发明的保护范围。所述应用的具体方法如下:将gmcml8基因通过所述重组表达载体导入目的植物中,得到转基因毛状根;所述转基因毛状根的增长率大于转空载体毛状根,转空载体毛状根为将植物表达载体prokii转入目的植物得到的转空载体毛状根。所述重组表达载体可通过ti质粒、ri质粒、植物病毒载体、直接dna转化、显微注射、电导、农杆菌介导等常规生物学方法转化到植物细胞或组织中。所述植物为单子叶植物或双子叶植物。所述双子叶植物可为豆目植物。所述豆目植物可为豆科植物。所述豆科植物可为大豆属植物。所述大豆属植物具体可为大豆,例如大豆科丰1号。以上任一所述耐盐性具体体现为:在盐胁迫环境中的毛状根相对增长率高。本发明提供了gmcml8蛋白及其编码基因,gmcml8基因的过量表达提高了转基因植株的耐盐性,对培育耐盐植物品种,特别是培育耐盐的作物、林草等新品种具有重要意义,可用于农牧业和生态环境治理所需的耐盐性植物品种的培育和鉴定。附图说明图1为重组表达载体prokii-gmcml8的部分结构示意图。图2为gmcml8基因表达量分析图。图3为正常培养10天的毛状根表型观察结果图。图4为100mmnacl水溶液处理后的毛状根表型观察结果图。图5为100mmnacl水溶液处理后的毛状根相对增长率统计结果图。具体实施方式以下的实施例便于更好地理解本发明,但并不限定本发明。下述实施例中的实验方法,如无特殊说明,均为常规方法。下述实施例中所用的试验材料,如无特殊说明,均为自常规生化试剂商店购买得到的。以下实施例中的定量试验,均设置三次重复实验,结果取平均值。大豆科丰1号(glycinemaxl.merr.kefeng1):参考文献:w.k.zhang,y.j.wang,g.z.luo,j.s.zhang,c.y.he,x.l.wu,j.y.gai,s.y.chen,qtlmappingoftenagronomictraitsonthesoybean(glycinemaxl.merr.)geneticmapandtheirassociationwithestmarkers,theor.appl.genet,2004,108:1131-1139.;公众可从中科院遗传与发育生物学研究所获得。植物表达载体prokii:参考文献:d.c.baulcombe,g.r.saunders,m.w.bevan,m.a.mayoandb.d.hartison,expressionofbiologicallyactiveviralsatelliternafromthenucleargenomeoftransformedplants.nature321(1986),pp.446-449;公众可从中科院遗传与发育生物学研究所获得。发根农杆菌k599:参考文献:attilakereszt,etal.,agrobacteriumrhizogenes-mediadedtransformationofsoybeantostudyofrootbiology,natureprotocols,2007,2(4),549-552);公众可从petermgressnon教授,theuniversityofqueensland,stlucia,queensland4072,australia,获得,或经petermgressnon教授同意(书面同意书)后由中科院遗传与发育生物学研究所获得。实施例1、gmcml8基因的获得1、取大豆科丰1号3周龄幼苗经200mmnacl水溶液处理6小时后,取2克新鲜叶片,在液氮中研碎,悬于4mol/l硫氢酸胍水溶液中,混合物用酸性苯酚、氯仿抽提,上清液中加入无水乙醇沉淀得到总rna。2、将步骤1得到的总rna反转录为cdna,以cdna为模板,采用f和r组成的引物对进行pcr扩增,得到pcr产物。f:5′-gagggatccatggcagatgttctgagtgaag-3′;r:5′-gagggtacctcatccaatagtcatcatcatc-3′;f和r中,下划线分别标注bamhi和kpni酶切位点。pcr扩增体系(50μl):cdna1μl,10×buffer5μl,dntp(10mm)1μl,引物f1μl,引物r1μl,pfudnapolymerase(takara)1μl,ddh2o40μl。pcr扩增条件为:94℃3min;94℃30s,50℃30s,72℃30s,30个循环;72℃10min。3、将步骤2中得到的pcr产物与pgem-teasy(promega)连接,得到重组质粒。对pcr产物进行测序,测序结果如序列表的序列2所示。将序列表的序列1所示的蛋白质命名为蛋白gmcml8,由150个氨基酸残基组成。将蛋白gmcml8的编码基因命名为gmcml8基因。gmcml8基因的编码区如序列表的序列2所示(453bp)。实施例2、重组表达载体的构建1、提取大豆科丰1号3周龄幼苗叶片的总rna并反转录为cdna,以cdna为模板,采用f和r组成的引物对进行pcr扩增,得到pcr产物。f(序列表的序列3):5′-gagggatccatgaagattcagtgtgatgtg-3′;r(序列表的序列4):5′-gaggagctcttatccgagatccgggacag-3′;f和r中,下划线分别标注bamhi和kpni酶切位点。2、用限制性内切酶bamhi和kpni双酶切步骤1得到的pcr产物,回收酶切产物。3、用限制性内切酶bamhi和kpni双酶切植物表达载体prokii,回收约12.8kb的载体骨架。4、将步骤1的酶切产物和步骤2的载体骨架连接,得到重组表达载体prokii-cmcml8。根据测序结果,对重组表达载体prokii-gmcml8进行结构描述如下:将植物表达载体prokii的bamhi和kpni酶切位点之间的小片段取代为了序列表的序列2的自5′端第1-453位核苷酸所示的dna分子。重组表达载体prokii-gmcml8的部分结构示意图如图1所示。实施例3、过表达gmcml8毛状根的耐逆性鉴定一、过量表达gmcml8基因毛状根的获得1、将重组表达载体prokii-gmcml8通过电击法转化发根农杆菌k599,获得重组农杆菌,命名为k599/prokii-gmcml8。2、将植物表达载体prokii通过电击法转化发根农杆菌k599,获得重组农杆菌,命名为k599/prokii。3、向生长6天含两片真叶的大豆科丰1号幼苗的每片叶片内注射重组农杆菌k599/prokii-gmcml8,在光照16小时黑暗18小时,温度25℃,湿度50%的条件下培养2周。2周后大豆科丰1号幼苗长出毛状根即为转化的毛状根。共获得36个转gmcml8基因毛状根。4、向生长6天含两片真叶的大豆科丰1号幼苗的每片叶片内注射重组农杆菌k599/prokii,在光照16小时黑暗18小时,温度25℃,湿度50%的条件下培养2周。2周后大豆科丰1号幼苗长出毛状根即为转化的毛状根。共获得34个转空载体毛状根。5、分别取步骤3获得的36个转gmcml8基因毛状根和步骤4获得的34个转空载体毛状根,提取总rna,将提取的总rna反转录为cdna。6、以步骤5得到的cdna为模板,采用f2和r2组成的引物对进行gmcml8基因表达量分析。f2:5′-atggcagatgttctgagtgaag-3′;r2:5′-tcatccaatagtcatcatcatc-3′;real-timepcr反应使用toyobo公司的realtimepcrmastermix试剂盒,并按照说明进行操作。采用大豆gmtubulin基因为内标,引物为tf和tr。tf:5′-aactccatttcgtccattccttc-3′;tr:5′-ttgagtggattcccaacaacg-3′;实验重复三次,结果取平均值±标准差。结果如图2所示。图2中,对照为转空载体毛状根,cml8-oe为转gmcml8基因毛状根,转gmcml8基因毛状根中gmcml8基因的相对表达量为3.4±1.1。结果表明,转gmcml8基因毛状根中gmcml8基因的表达量远高于转空载体毛状根中gmcml8基因的表达量。二、耐盐性鉴定待测样本为大豆科丰1号幼苗毛状根(对照)、转空载体毛状根、转gmcml8基因毛状根(cml8-oe)。1、在水中正常培养待测样本10天,结果如图3所示。大豆科丰1号幼苗毛状根(对照)的表型和转gmcml8毛状根(cml8-oe)的表型没有明显差异,转空载体毛状根的表型与大豆科丰1号幼苗毛状根(对照)的表型没有明显差异。2、将各待测样本浸入100mmnacl水溶液中25℃处理3天,观察表型变化并计算毛状根相对增长率,毛状根相对增长率计算方法如下:测量处理前和处理后的毛状根长度,毛状根的相对增长率=(处理后毛状根长-处理前毛状根长)/处理前毛状根长。进行三次重复试验,每次重复试验中每组待测样本取10个毛状根,结果取平均值±标准差。结果如图4和图5所示。图4为表型观察结果图,图5为毛状根相对增长率统计结果图。经100mmnacl水溶液处理前转gmcml8基因毛状根和大豆科丰1号幼苗毛状根的长度分别为4.7±2.3cm和4.3±2.1cm,两者间没有显著差异,转空载体毛状根的长度与二者均没有显著差别,经100mmnacl水溶液处理后转gmcml8基因毛状根和大豆科丰1号幼苗毛状根的长度分别为5.6±2.0cm和4.9±2.1cm,转空载体毛状根的长度与大豆科丰1号幼苗毛状根的长度无显著性差别。经过计算,经100mmnacl水溶液处理的转gmcml8基因毛状根的相对增长率为19.5±3.9%,经100mmnacl水溶液处理的大豆科丰1号幼苗毛状根的相对增长率为14.1±3.6%,两者有显著差异,转空载体毛状根的长度增长率与大豆科丰1号幼苗毛状根的长度增长率无显著差别。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 蛋白质及其编码基因在调控植物抗黄萎病性中的应用的制造方法与工艺
- 甘薯耐逆相关蛋白IbFBA2、编码基因、重组载体与应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 降低种子重量的白桦AP2基因及其编码蛋白的制造方法与工艺
- 可编码ArtRD蛋白基因及其蛋白和应用的制造方法与工艺
- 蝎神经毒素AaIT及其编码基因与应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种花狭口蛙的基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种泽陆蛙基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺