一种酶活及热稳定性改善的凝乳酶突变体的制作方法
本发明涉及一种酶活及热稳定性改善的凝乳酶突变体,属于酶工程技术领域。
背景技术:
凝乳酶是一种最早在未断奶的小牛胃中发现的天门冬氨酸蛋白酶,可专一地切割乳中κ-酪蛋白的Phe105-Met106之间的肽键,破坏酪蛋白胶束使牛奶凝结,凝乳酶的凝乳能力及蛋白水解能力使其成为干酪生产中形成质构和特殊风味的关键性酶,被广泛地应用于奶酪和酸奶的制作。
增强凝乳酶的凝乳能力,降低对蛋白的水解能力以及降低耐热性一直是凝乳酶蛋白改造的热门方向。Chitpinityol等(1998)利用定点突变技术对77位点进行苏氨酸到天冬氨酸的改变,导致突变子较重组野生型凝乳酶活力有所下降;目前为止,对于小牛凝乳酶等动物凝乳酶及微小毛霉等真菌凝乳酶研究较多,对于细菌源凝乳酶定点突变的研究国内暂未发现。
技术实现要素:
本发明的第一个目的是提供一种凝乳酶突变体,其氨基酸序列如SEQ ID NO.1~3所示。
本发明的第二个目的是提供编码所述凝乳酶突变体的基因。
本发明的第三个目的是提供表达所述凝乳酶突变体的基因工程菌,所述基因工程菌以pPIC9K-cMCE为载体,以毕赤酵母为宿主,表达SEQ ID NO.1~3任一所示的凝乳酶突变体。
在本发明的一种实施方式中,所述基因工程菌以毕赤酵母GS115为宿主。
本发明的第四个目的是提供构建所述基因工程菌的方法,所述方法是将pPIC9K-cMCE与编码所述凝乳酶突变体的基因连接,转化至毕赤酵母中。
在本发明的一种实施方式中,所述pPIC9K-cMCE的构建方法公开于专利申请号为201611157248.4的发明专利申请中;所述毕赤酵母是毕赤酵母GS115。
本发明的第五个目的是提供一种提高凝乳酶酶活的方法,是将SEQ ID NO.4所示的凝乳酶第75位亮氨酸突变为谷氨酰胺获得SEQ ID NO.1所示突变体,或将第222位的蛋氨酸替换为苏氨酸获得SEQ ID NO.2所示突变体。
在本发明的一种实施方式中,所述方法是将SEQ ID NO.4所示的凝乳酶第75位亮氨酸突变为谷氨酰胺,并将第222位的蛋氨酸替换为苏氨酸获得SEQ ID NO.3所示突变体。
本发明的第六个目的是提供一种产凝乳酶的方法,是将所述基因工程菌接种至发酵培养基中,25~30℃培养24~96h。
在本发明的一种实施方式中,所述发酵通过流加甲醇高效诱导毕赤酵母胞外表达凝乳酶。
本发明还提供所述凝乳酶突变体在食品、生物、医药领域的应用。
在本发明的一种实施方式中,所述应用具体为制备奶酪等乳制品。
有益效果:(1)本发明首次在解淀粉芽孢杆菌源凝乳酶基因上进行定点突变,采用PCR介导的一步法,进行单位点突变以及双重突变,整个过程简洁清晰,成功率达到90%以上。(2)Leu75Gln与Met222Thr双重突变效果最好,粗酶液凝乳酶活,C/P值均提升了155.6%和195.0%,热稳定性也得到了降低,相比出发菌株降低了13.6%。提升了此类凝乳酶的商业价值,拓宽了商业领域,为获得优良的工业凝乳酶产品奠定了重要基础。
附图说明
图1突变重组质粒双酶切验证;其中,M,15000bp Marker;0,出发菌株P-Cmce;1~6分别对应重组质粒pPIC9K-cMCE-Leu75Gln,pPIC9K-cMCE-Val81Asp,pPIC9K-cMCE-Met222Thr,pPIC9K-cMCE-Leu75Gln-Val81Asp,pPIC9K-cMCE-Leu75Gln-Met222Thrp,和PIC9K-cMCEVal81Asp-Met222Thr;
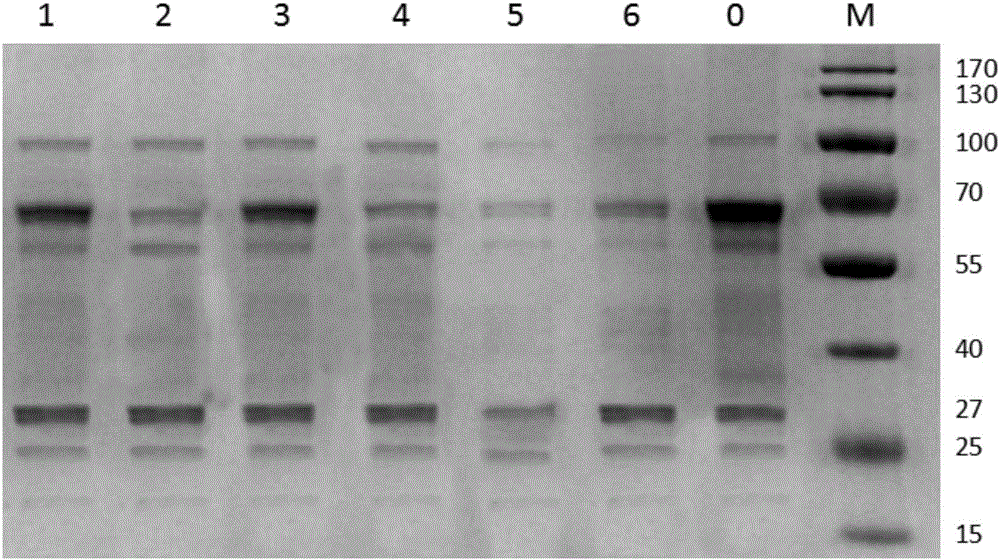
图2为突变转化子摇瓶培养96h粗酶液的SDS-PAGA图;其中,M,Marker;0,出发菌株P-Cmce;1~6,分别对应重组质粒pPIC9K-cMCE-Leu75Gln,pPIC9K-cMCE-Val81Asp,pPIC9K-cMCE-Met222Thr,pPIC9K-cMCE-Leu75Gln-Val81Asp,pPIC9K-cMCE-Leu75Gln-Met222Thrp,和PIC9K-cMCEVal81Asp-Met222Thr;
图3为突变体与对照的热稳定性比较。
具体实施方式
1)凝乳活力测定
根据MAGDA方法,将脱脂乳粉用0.01mol/L CaCl2配制10%复原脱脂乳溶液35℃条件下取待测酶液0.2mL加入2mL复原脱脂乳中并混匀,开始计时,至絮状凝集颗粒出现为止(即40min凝固1mL 100g/L的脱脂乳所需的酶量定义为一个酶活力单位)。
酶活力(soxhelt unit,SU)=(脱脂乳量mL/凝乳酶量mL)×D×2400/T
注:D为酶液稀释倍数;T为凝乳时间。
2)蛋白水解活力测定
初酶液和酪蛋白溶液40℃水浴中预热2min,取1mL酪蛋白溶液与1mL酶液混匀,40℃精确保温10min,时间终止立即加入0.4mol/L三氯乙酸2mL,终止反应,继续使其在水浴中保温20min,离心去除沉淀,取上清1min,加入0.4mol/L碳酸钠5mL 0.67moL/L的福林试剂1mL,摇匀,反应20min后光密度(OD660)测定。空白在加酪蛋白之前加入0.4mol/L三氯乙酸2mL,使酶失活,再加入酪蛋白。
3)热稳定性测定
将待测样品分别放在40℃,45℃,50℃,55℃,60℃,65℃温度下保温120min,每隔10min取样一次,冰浴冷却后在35℃下测定凝乳酶的剩余凝乳活力,以未经热处理的凝乳活力为100%。
4)培养基
BMGY培养基:10g/L酵母粉,20g/L蛋白陈,3g/L磷酸氢二钾,11.8g/L磷酸二氢钾,13.4g/L YNB,4×10-4g/L生物素,10g/L甘油。
BMMY培养基:10g/L酵母粉,20g/L蛋白陈,3g/L磷酸氢二钾,11.8g/L磷酸二氢钾,13.4g/L YNB,4×10-4g/L生物素,5ml/L无水甲醇。
MM培养基:13.4g/L YNB,4×10-4g/L生物素,5ml/L甲醇,15g/L琼脂。
MD培养基:13.4g/L YNB,4×10-4g/L生物素,20g/L葡萄糖,0.4g/L组氨酸,15g/L琼脂。
实施例1
(1)确定解淀粉芽孢杆菌源凝乳酶基因定点突变位点
通过计算机对微小毛霉凝乳酶基因进行三级结构模拟,同时与牛凝乳酶三级结构及核酸序列对比,选定3个关键位点进行定点突变,分别选定可能与水解或热稳定性相关的位点75、81位点和222位点。
75位点原为亮氨酸,81位点原为缬氨酸,222位点原为蛋氨酸,本发明预将75位点突变为谷氨酰胺,81位点突变为天冬氨酸,222位点突变为苏氨酸,通过PCR介导的一步法进行点突变。
通过DNAMAN对选定的突变位点设计引物,如表1所示。
表1定点突变所用的引物序列
(2)PCR介导的一步法定点突变
用杭州宝赛生物科技有限公司的试剂盒(PE01)提取大肠杆菌JM109-cMCE的重组质粒pPIC9K-cMCE(公开于申请号为201611157248.4的发明专利申请中)。以Leu75Gln-F和Leu75Gln-R为引物对凝乳酶第75位亮氨酸进行突变,构建重组质粒pPIC9K-cMCE-Leu75Gln;以Val81Asp-F和Val81Asp-R为引物,对凝乳酶第81位缬氨酸进行突变,构建重组质粒pPIC9K-cMCE-Val81Asp;以Met222Thr-F和Met222Thr-R为引物,对凝乳酶第222位蛋氨酸进行突变,构建重组质粒pPIC9K-cMCE-Met222Thr。
PCR反应体系如下:super pfu PCR Master Mix 50μL,ddH2O 50μL,20μM/L上、下游引物各2μL,模板DNA 2μL,总体积100μL,分装10管,每管10μL。反应条件:94℃预变性3min;98℃变性40s,60℃退火1min,68℃延伸11min;18次circle;72℃延伸10min。将PCR产物进行琼脂糖凝胶电泳,结果如图1所示。
在单位点突变成功后,以单位点突变质粒为模板,用另一对突变引物,按照同样的PCR体系,引入第二个突变位点,即构建双重突变位点Leu75Gln-Val81Asp,Leu75Gln-Met222Thr,Val81Asp-Met222Thr。
PCR反应结束后,直接在PCR反应体系中加入1μL Dpn I,混匀后37℃金属浴孵育1h。
用杭州宝赛生物科技有限公司的试剂盒(PE03)纯化PCR产物,重组载体转化到大肠杆菌JM109感受态细胞,获得重组菌,挑取重组菌单克隆,提取质粒pPIC9K-cMCE-Leu75Gln,pPIC9K-cMCE-Val81Asp,pPIC9K-cMCE-Met222Thr,pPIC9K-cMCE-Leu75Gln-Val81Asp,pPIC9K-cMCE-Leu75Gln-Met222Thrp,和PIC9K-cMCEVal81Asp-Met222Thr。
用EcoR I和Not I双酶切重组载体pPIC9K-cMCE-Leu75Gln,pPIC9K-cMCE-Val81Asp,pPIC9K-cMCE-Met222Thr,pPIC9K-cMCE-Leu75Gln-Val81Asp,pPIC9K-cMCE-Leu75Gln-Met222Thrp,和PIC9K-cMCEVal81Asp-Met222Thr后,进行琼脂糖凝胶电泳,如图1,载体和目的片段的长度分别都相等。
挑选酶切鉴定正确的重组载体送生工生物工程(上海)股份有限公司测序,扩增的凝乳酶基因的核苷酸序列以及与原序列比对结果的一致性达到99.82%,除突变位点之外的核苷酸序列完全一致。。
实施例2
(1)凝乳酶突变体在毕赤酵母里的表达
凝乳酶突变体在毕赤酵母里的表达步骤参照申请号为201611157248.4的专利申请实施例一的相关操作步骤。
重组载体pPIC9K-cMCE-Leu75Gln,pPIC9K-cMCE-Val81Asp,pPIC9K-cMCE-Met222Thr,pPIC9K-cMCE-Leu75Gln-Val81Asp,pPIC9K-cMCE-Leu75Gln-Met222Thrp,和PIC9K-cMCEVal81Asp-Met222Thr用Sac I酶切线性化后,2500V,5ms电转化毕赤酵母GSll5感受态细胞先后涂布到MM和MD平板上,筛选出His+和Mut+型转化子P-cMCE。
挑选转化子进行菌落PCR验证,将PCR产物进行琼脂糖凝胶电泳,挑选转化子于BMGY培养基中,30℃/300rpm培养至0D600=2~6;收集菌体,用BMMY重悬菌体,使OD600=5.0左右,向培养基中添加无水甲醇至终浓度为0.5%(v/v,5mL甲醇/L培养基);每12h添加终浓度0.5%(v/v)的甲醇,96h后结束诱导,浓缩诱导96h后发酵液上清。
(2)重组凝乳酶的纯化
取毕赤酵母P-cMCE诱导96小时的发酵液100mL,然后加入饱和过硫酸铵溶液至过硫酸铵终浓度为60%,4℃放置过夜。10000rpm离心10min,收集蛋白沉淀,真空冷冻干燥,得到P-cMCE酶制剂0.566g。10mL磷酸缓冲盐溶液复溶,取20ul样品进行SDS-PAGE电泳检测。
SDS-PAGE电泳结果如图2所示,在分子量27kDa处有明显的条带,基本符合原菌凝乳酶蛋白大小。
实施例3
(1)凝乳酶活,蛋白酶活及C/P值性质的比较
C/P值是指单位质量的纯酶粉其凝乳酶活性与蛋白酶活性的比率。能切实准确的表征凝乳酶的凝乳能力。
突变质粒Leu75Gln,Val81Asp,Met222Thr,Leu75Gln-Val81Asp,Leu75Gln-Met222Thr,Val81Asp-Met222Thr转化毕赤酵母GS115,随机挑取30个转化子,接种量0.5%,以未进行突变的出发菌株P-cMCE作为对照,经BMGY培养和甲醇诱导,pH处于6.5~6.7,发酵温度30℃在BMMY培养基中发酵96h,离心取上清,测定粗酶液凝乳酶活力,蛋白水解活力,计算C/P比值,结果如表2所示。
由表2可以看出,在单位点突变中,Leu75Gln突变凝乳酶凝乳活性增高最多,达到25.0SU/mL,相比出发菌株提高了138.9%,Val81Asp突变凝乳酶蛋白水解活性明显降低最为显著,但凝乳活性极低,不利于凝乳作用;在双重突变位点中,Leu75Gln-Met222Thr双重突变凝乳酶凝乳活性增高最多,达到28.0SU/mL,蛋白水解活性降低,同时,C/P比值大幅有提高,相比出发菌株提高了188.2%。
表2为三种定点突变转化子与出发菌株的酶活力比较
(2)热稳定性的比较
对凝乳酶突变体Leu75Gln和Met222Thr以及这两个突变位点组合的突变体Leu75Gln-Met222Thr进行热稳定性的复筛,结果如图3所示。
由图3可以看出,Leu75Gln和Met222Thr两个位点的突变,均使突变凝乳酶热稳定性变弱,均弱于出发菌株。受到凝乳酶在应用方面的特性影响,当凝乳酶的热稳定性变差,其在较低温度下发生失活的现象,从而使乳制品(例如奶酪等)在存放过程中不被水解成为乳清。相对于Leu75Gln位突变凝乳酶,T218S位突变凝乳酶具有更弱的热稳定性。Met222Thr位突变凝乳酶45℃处理60min后,相对凝乳活力为62.0%。Leu75Gln与Met222Thr双重突变凝乳酶热稳定性弱化程度虽不如Met222Thr单位点突变凝乳酶,但相比出发菌株P-cMCE弱化了16%。
虽然本发明已以较佳实施例公开如上,但其并非用以限定本发明,任何熟悉此技术的人,在不脱离本发明的精神和范围内,都可做各种的改动与修饰,因此本发明的保护范围应该以权利要求书所界定的为准。
SEQUENCE LISTING
<110> 江南大学
<120> 一种酶活及热稳定性改善的凝乳酶突变体
<160> 10
<170> PatentIn version 3.3
<210> 1
<211> 275
<212> PRT
<213> 人工序列
<400> 1
Ala Gln Ser Val Pro Tyr Gly Val Ser Gln Ile Lys Ala Pro Ala Leu
1 5 10 15
His Ser Gln Gly Tyr Thr Gly Ser Asn Val Lys Val Ala Val Ile Asp
20 25 30
Ser Gly Ile Asp Ser Ser His Pro Asp Leu Lys Val Ala Gly Gly Ala
35 40 45
Ser Met Val Pro Ser Glu Thr Asn Pro Phe Gln Asp Asn Asn Ser His
50 55 60
Gly Thr His Val Ala Gly Thr Val Ala Ala Gln Asn Asn Ser Ile Gly
65 70 75 80
Val Leu Gly Val Ala Pro Ser Ala Ser Leu Tyr Ala Val Lys Val Leu
85 90 95
Gly Ala Asp Gly Ser Gly Gln Tyr Ser Trp Ile Ile Asn Gly Ile Glu
100 105 110
Trp Ala Ile Ala Asn Asn Met Asp Val Ile Asn Met Ser Leu Gly Gly
115 120 125
Pro Ser Gly Ser Ala Ala Leu Lys Ala Ala Val Asp Lys Ala Val Ala
130 135 140
Ser Gly Val Val Val Val Ala Ala Ala Gly Asn Glu Gly Thr Ser Gly
145 150 155 160
Ser Ser Ser Thr Val Gly Tyr Pro Gly Lys Tyr Pro Ser Val Ile Ala
165 170 175
Val Gly Ala Val Asp Ser Ser Asn Gln Arg Ala Ser Phe Ser Ser Val
180 185 190
Gly Pro Glu Leu Asp Val Met Ala Pro Gly Val Ser Ile Gln Ser Thr
195 200 205
Leu Pro Gly Asn Lys Tyr Gly Ala Tyr Asn Gly Thr Ser Met Ala Ser
210 215 220
Pro His Val Ala Gly Ala Ala Ala Leu Ile Leu Ser Lys His Pro Asn
225 230 235 240
Trp Thr Asn Thr Gln Val Arg Ser Ser Leu Glu Asn Thr Thr Thr Lys
245 250 255
Leu Gly Asp Ser Phe Tyr Tyr Gly Lys Gly Leu Ile Asn Val Gln Ala
260 265 270
Ala Ala Gln
275
<210> 2
<211> 275
<212> PRT
<213> 人工序列
<400> 2
Ala Gln Ser Val Pro Tyr Gly Val Ser Gln Ile Lys Ala Pro Ala Leu
1 5 10 15
His Ser Gln Gly Tyr Thr Gly Ser Asn Val Lys Val Ala Val Ile Asp
20 25 30
Ser Gly Ile Asp Ser Ser His Pro Asp Leu Lys Val Ala Gly Gly Ala
35 40 45
Ser Met Val Pro Ser Glu Thr Asn Pro Phe Gln Asp Asn Asn Ser His
50 55 60
Gly Thr His Val Ala Gly Thr Val Ala Ala Leu Asn Asn Ser Ile Gly
65 70 75 80
Val Leu Gly Val Ala Pro Ser Ala Ser Leu Tyr Ala Val Lys Val Leu
85 90 95
Gly Ala Asp Gly Ser Gly Gln Tyr Ser Trp Ile Ile Asn Gly Ile Glu
100 105 110
Trp Ala Ile Ala Asn Asn Met Asp Val Ile Asn Met Ser Leu Gly Gly
115 120 125
Pro Ser Gly Ser Ala Ala Leu Lys Ala Ala Val Asp Lys Ala Val Ala
130 135 140
Ser Gly Val Val Val Val Ala Ala Ala Gly Asn Glu Gly Thr Ser Gly
145 150 155 160
Ser Ser Ser Thr Val Gly Tyr Pro Gly Lys Tyr Pro Ser Val Ile Ala
165 170 175
Val Gly Ala Val Asp Ser Ser Asn Gln Arg Ala Ser Phe Ser Ser Val
180 185 190
Gly Pro Glu Leu Asp Val Met Ala Pro Gly Val Ser Ile Gln Ser Thr
195 200 205
Leu Pro Gly Asn Lys Tyr Gly Ala Tyr Asn Gly Thr Ser Thr Ala Ser
210 215 220
Pro His Val Ala Gly Ala Ala Ala Leu Ile Leu Ser Lys His Pro Asn
225 230 235 240
Trp Thr Asn Thr Gln Val Arg Ser Ser Leu Glu Asn Thr Thr Thr Lys
245 250 255
Leu Gly Asp Ser Phe Tyr Tyr Gly Lys Gly Leu Ile Asn Val Gln Ala
260 265 270
Ala Ala Gln
275
<210> 3
<211> 275
<212> PRT
<213> 人工序列
<400> 3
Ala Gln Ser Val Pro Tyr Gly Val Ser Gln Ile Lys Ala Pro Ala Leu
1 5 10 15
His Ser Gln Gly Tyr Thr Gly Ser Asn Val Lys Val Ala Val Ile Asp
20 25 30
Ser Gly Ile Asp Ser Ser His Pro Asp Leu Lys Val Ala Gly Gly Ala
35 40 45
Ser Met Val Pro Ser Glu Thr Asn Pro Phe Gln Asp Asn Asn Ser His
50 55 60
Gly Thr His Val Ala Gly Thr Val Ala Ala Gln Asn Asn Ser Ile Gly
65 70 75 80
Val Leu Gly Val Ala Pro Ser Ala Ser Leu Tyr Ala Val Lys Val Leu
85 90 95
Gly Ala Asp Gly Ser Gly Gln Tyr Ser Trp Ile Ile Asn Gly Ile Glu
100 105 110
Trp Ala Ile Ala Asn Asn Met Asp Val Ile Asn Met Ser Leu Gly Gly
115 120 125
Pro Ser Gly Ser Ala Ala Leu Lys Ala Ala Val Asp Lys Ala Val Ala
130 135 140
Ser Gly Val Val Val Val Ala Ala Ala Gly Asn Glu Gly Thr Ser Gly
145 150 155 160
Ser Ser Ser Thr Val Gly Tyr Pro Gly Lys Tyr Pro Ser Val Ile Ala
165 170 175
Val Gly Ala Val Asp Ser Ser Asn Gln Arg Ala Ser Phe Ser Ser Val
180 185 190
Gly Pro Glu Leu Asp Val Met Ala Pro Gly Val Ser Ile Gln Ser Thr
195 200 205
Leu Pro Gly Asn Lys Tyr Gly Ala Tyr Asn Gly Thr Ser Thr Ala Ser
210 215 220
Pro His Val Ala Gly Ala Ala Ala Leu Ile Leu Ser Lys His Pro Asn
225 230 235 240
Trp Thr Asn Thr Gln Val Arg Ser Ser Leu Glu Asn Thr Thr Thr Lys
245 250 255
Leu Gly Asp Ser Phe Tyr Tyr Gly Lys Gly Leu Ile Asn Val Gln Ala
260 265 270
Ala Ala Gln
275
<210> 4
<211> 275
<212> PRT
<213> 人工序列
<400> 4
Ala Gln Ser Val Pro Tyr Gly Val Ser Gln Ile Lys Ala Pro Ala Leu
1 5 10 15
His Ser Gln Gly Tyr Thr Gly Ser Asn Val Lys Val Ala Val Ile Asp
20 25 30
Ser Gly Ile Asp Ser Ser His Pro Asp Leu Lys Val Ala Gly Gly Ala
35 40 45
Ser Met Val Pro Ser Glu Thr Asn Pro Phe Gln Asp Asn Asn Ser His
50 55 60
Gly Thr His Val Ala Gly Thr Val Ala Ala Leu Asn Asn Ser Ile Gly
65 70 75 80
Val Leu Gly Val Ala Pro Ser Ala Ser Leu Tyr Ala Val Lys Val Leu
85 90 95
Gly Ala Asp Gly Ser Gly Gln Tyr Ser Trp Ile Ile Asn Gly Ile Glu
100 105 110
Trp Ala Ile Ala Asn Asn Met Asp Val Ile Asn Met Ser Leu Gly Gly
115 120 125
Pro Ser Gly Ser Ala Ala Leu Lys Ala Ala Val Asp Lys Ala Val Ala
130 135 140
Ser Gly Val Val Val Val Ala Ala Ala Gly Asn Glu Gly Thr Ser Gly
145 150 155 160
Ser Ser Ser Thr Val Gly Tyr Pro Gly Lys Tyr Pro Ser Val Ile Ala
165 170 175
Val Gly Ala Val Asp Ser Ser Asn Gln Arg Ala Ser Phe Ser Ser Val
180 185 190
Gly Pro Glu Leu Asp Val Met Ala Pro Gly Val Ser Ile Gln Ser Thr
195 200 205
Leu Pro Gly Asn Lys Tyr Gly Ala Tyr Asn Gly Thr Ser Met Ala Ser
210 215 220
Pro His Val Ala Gly Ala Ala Ala Leu Ile Leu Ser Lys His Pro Asn
225 230 235 240
Trp Thr Asn Thr Gln Val Arg Ser Ser Leu Glu Asn Thr Thr Thr Lys
245 250 255
Leu Gly Asp Ser Phe Tyr Tyr Gly Lys Gly Leu Ile Asn Val Gln Ala
260 265 270
Ala Ala Gln
275
<210> 5
<211> 23
<212> DNA
<213> 人工序列
<400> 5
agttgcggct caaaataact caa 23
<210> 6
<211> 23
<212> DNA
<213> 人工序列
<400> 6
ttgagttatt ttgagccgca act 23
<210> 7
<211> 23
<212> DNA
<213> 人工序列
<400> 7
ctcaatcggt aacttaggcg ttg 23
<210> 8
<211> 23
<212> DNA
<213> 人工序列
<400> 8
caacgcctaa gttaccgatt gag 23
<210> 9
<211> 23
<212> DNA
<213> 人工序列
<400> 9
cggtacgtca atggcatctc cgc 23
<210> 10
<211> 23
<212> DNA
<213> 人工序列
<400> 10
gcggagatgc cattgacgta ccg 23
- 还没有人留言评论。精彩留言会获得点赞!