一种重组融合碱性磷酸酶-过敏原蛋白及其制备方法和应用与流程
本发明涉及体外免疫诊断技术领域,具体涉及一种重组融合碱性磷酸酶-过敏原蛋白及其制备方法和应用,特别是涉及重组融合碱性磷酸酶-过敏原蛋白在ige体外检测试剂盒方面的应用。
背景技术:
过敏症多为ige介导的对自然界中抗原分子产生的不良免疫反应。正常情况下,血液中检测不到对一种过敏原具有特异性的ige,仅在人体被外界抗原物质致敏后产生。引起过敏反应的物质过敏原可诱导产生一种特异性的ige,其只与该物质发生反应。sige(specificige,特异性ige)与过敏原结合后引起细胞释放组胺,组胺引发过敏症,如哮喘、湿疹、皮疹、眼部发痒、鼻炎、流涕等,严重者甚至昏迷、休克。
传统的过敏原体内诊断试验通常具有较高的特异性和敏感性,但易受抗过敏药物和皮肤状况等影响,同时操作程序易诱发严重过敏反应,且需丰富经验的专业医生诊断,因此安全性不高。体外诊断测血清sige,具有安全可靠的优点,是临床常用的ige介导的i型变态反应性疾病特异性诊断方法,具有重要的临床意义。真正意义上根据实际检测数据进行过敏原分级的检测方法只有酶联免疫荧光法和酶联免疫捕获法。酶联免疫捕获法检测总ige和sige结果符合性良好,对过敏性疾病具有良好的诊断价值,操作过程中依靠酶标仪就能得到可靠的结果,而且成本适中,性价比高,有效减轻了患者的经济负担。
过敏原体外诊断的传统方法,酶联免疫法如allergen系统,其原理如下:抗人ige包被酶标板,吸附样品中总ige和特异性ige,再与标记的过敏原形成抗体抗原复合物,最后经酶与底物显色以检测样品中ige的含量。首先,加入样品孵育使酶标板结合所有的ige抗体,然后洗涤,避免其他免疫球蛋白如igg抗体的干扰,再加入生物素标记的过敏原使其与ige特异性结合,洗涤除去未结合生物素抗原,然后加入标记了辣根过氧化酶或碱性磷酸酶的链霉亲和素标记结合物,洗涤除去未结合标记结合物,加入显色底物,通过酶与底物反应后吸光度或者发光强度的变化来检测针对特异性过敏原的ige的量。
现有试剂盒的标记抗原主要是过敏原的天然提取物或重组蛋白。由于提取物纯度不足,时有蛋白酶残留,造成提取物不稳定,其中含有的杂蛋白会造成假阳性。重组过敏原成分单一浓度高,有利于提高过敏原检测的灵敏度。重组过敏原相对于天然提取物就其生产方式而言更利于进行标准化,且在不同的生产批次之间的差别更小、更稳定。
重组过敏原的表达方式主要分为原核表达、酵母表达、昆虫细胞、哺乳细胞表达、植物细胞表达,原核表达产量高,但无法对重组蛋白进行一些必要的修饰,且表达产物若是包涵体,则蛋白难以复性。利用真核细胞表达过敏原,蛋白质能正确折叠,可提供复杂又准确的糖基化修饰功能,因此表达产物在分子结构、理化性质及生物学功能方面最接近于天然的高等蛋白质分子,但是相对来说,表达产量低,纯化困难,蛋白浓度较高才能够进行生物素化标记,标记之后还需要纯化,除去未标记物等,总的来说操作复杂、成本较高。
技术实现要素:
为克服现有技术的不足,本发明提供了一种重组融合碱性磷酸酶-过敏原蛋白及其制备方法和应用,特别是涉及重组融合碱性磷酸酶-过敏原蛋白在ige体外检测试剂盒方面的应用。本发明提供的方法能克服现有技术缺点,实现生产简单、成本低和高灵敏度。
本发明提供了一种重组融合碱性磷酸酶-过敏原蛋白的表达载体,该载体由一种含表达所述碱性磷酸酶的控制序列和表达所述过敏原蛋白的dna分子有功能地连接组成,所述表达所述碱性磷酸酶的控制序列为pcmv-signalpeptide-seap-linker-mcs-ha或pcmv-signalpeptide-mcs-linker-seap-ha,所述linker为1-250个氨基酸。
优选的,上述载体中所述过敏原蛋白包括虾原肌球蛋白、蟹原肌球蛋白、蟹精氨酸激酶、鸡蛋卵类黏蛋白gald1、鸡蛋卵清蛋白gald2、鸡蛋卵转铁蛋白gald3、鸡蛋溶菌酶gald4;牛奶β-乳球蛋白、牛奶α-乳白蛋白、牛奶酪蛋白;德国蟑螂blag1~8、屋尘螨derp1~2;粉尘螨derf1~2;猫毛feld1、狗毛canf1或豚草中的致敏蛋白,或与所述过敏原蛋白共享抗体表位的变体或片段;所述过敏原的基因通过rt-pcr扩增获得或通过人工合成获得。
优选的,上述表达载体为真核表达载体、原核表达载体、昆虫表达载体、酵母表达载体或植物表达载体。
本发明的第二个目的是提供一种重组融合碱性磷酸酶-过敏原蛋白的制备方法,包括如下步骤:
(1)将上述的表达载体转染宿主细胞进行表达;
(2)筛选并建立稳定细胞系;
(3)摇瓶发酵大量表达目的蛋白。
优选的,所述宿主细胞为哺乳细胞、昆虫表达系统、酵母表达系统、植物表达系统;
优选的,所述哺乳细胞为cho或hek293。
本发明还提供了上述方法获得的重组融合碱性磷酸酶-过敏原蛋白。
本发明的第四个目的是提供上述重组融合碱性磷酸酶-过敏原蛋白在体外诊断i型过敏症方面的应用。
本发明的第五个目的是提供一种检测过敏原特异性ige抗体的试剂盒,所述试剂盒包括:已包被抗人ige的酶标板、上述重组融合碱性磷酸酶-过敏原蛋白、样品稀释液和底物显色液;
所述样品稀释液为10mmol/lph7.4-7.6的磷酸盐缓冲液,所述磷酸盐缓冲液中含质量百分比为1%的bsa和体积百分比为0.05%的tween-20;
所述底物显色液为碱性磷酸酶化学发光液或者bcip/nbt。
本发明的第六个目的是提供一种检测过敏原特异性ige抗体的方法,包含以下步骤:
(1)捕获板的制备:将抗人ige包被于96孔板上;
(2)加入待测样品,所述待测样品与所述步骤(1)中的抗人ige进行孵育,得到抗抗体与抗体的结合物;
(3)将上述重组融合碱性磷酸酶-过敏原蛋白同所述步骤(2)中的待测样品中的目的抗体进行抗原抗体反应,形成碱性磷酸酶-抗原-抗体复合物;
(4)显色检测。
本发明的有益效果在于:
(1)本发明涉及重组融合碱性磷酸酶-过敏原蛋白在哺乳细胞中的融合表达,利用哺乳细胞获得重组过敏原,其蛋白结构、理化特征更接近天然过敏原蛋白,因此提高了检测灵敏度,减少了非特异性反应。
(2)利用分泌表达方式,表达产物碱性磷酸酶-过敏原蛋白分泌到细胞外,辅以无血清培养基,直接获得标记抗原,从而省去了传统过敏原的纯化、浓缩、标记、再纯化等过程,并且轻松实现了酶与抗原1:1的标记,绝对避免传统标记批间差,简化了标记物的生产过程。
(3)利用与碱性磷酸酶融合表达的过敏原制备试剂盒,免去了过敏原纯化、浓缩、过敏原-酶标记及标记后的纯化过程,极大地简化了关键原料的制备过程,降低了生产成本,提高了过敏原的活性,节省了试剂盒使用的操作步骤和时间。
(4)本发明的检测过敏原特异性ige抗体的方法测血清中过敏原特异性sige的捕获夹心法灵敏度高、特异性强,操作简便快捷。具体实验操作步骤,将“加入生物素标记的过敏原蛋白使其与ige特异性结合,洗涤除去未结合生物素抗原,然后加入标记了辣根过氧化酶或碱性磷酸酶的链霉亲和素标记结合物”两步反应变为“加入碱性磷酸酶-过敏原蛋白”一步反应,简化了实验操作步骤,缩短了整个试剂盒的操作时间,方便用户及早获得检测结果。
附图说明
图1为pcmv-signalpeptide-seap-linker-mcs-ha重组表达载体示意图;
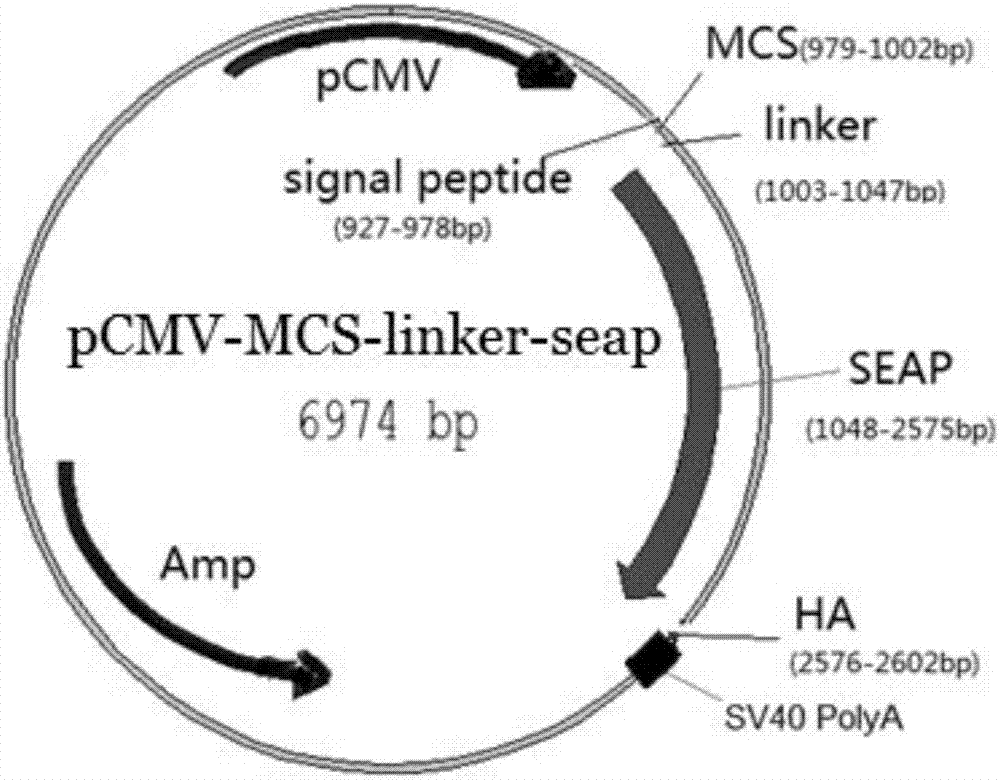
图2为pcmv-signalpeptide-mcs-linker-seap-ha重组载体示意图;
图3为pfastbaci-ecori-待插入目的基因-xbai-linker-seap-tag的示意图。
图4为ppiczalphaa-ecori-待插入目的基因-xbai-linker-seap的示意图。
具体实施方式
为使本发明更加容易理解,下面结合具体实施例,进一步阐述本发明。应理解,这些实施例仅用于说明本发明而不用于限制本发明的范围。
实施例1:碱性磷酸酶融合过敏原重组质粒的制备
1、过敏原基因制备方法:
1.1从ncbi数据库中找出甲壳类tm、蟹cak、鸡蛋gald1、鸡蛋gald2、鸡蛋gald3及蟑螂blag5、blag8的氨基酸序列,利用codehop针对每一物种分别设计兼并引物。
1.2组织制备
(1)从菜市场买回鲜活的虾,将其肌肉剪小块,-80℃保存备用。
(2)将买回的活蟹去壳,用小刀刮取其肌肉,剪小块,-80℃冻存备用。
(3)从菜市场购买刚从鸡中取出的鸡输卵管,用剪刀剪碎,-80℃冻存备用。
(4)从室内抓到的德国小蠊,饿过夜后,剪碎,-80℃冻存备用。
1.3将上述-80℃冻存的组织分别用液氮研磨,通过trizol(购自天根公司)法分别抽提rna。
1.4各物种的rna分别保存于-80℃备用,或直接做rt-pcr(购自takara)。
1.5分别rt-pcr产物回收,连接至pmd19t(购自takara)中,测序。将测序正确的重组载体保存于-20℃。
2、过敏原基因合成
除了通过rt-pcr扩增获得过敏原基因外,也可直接合成。合成的过敏原基因两端加入合适的酶切位点。
3、哺乳细胞原始表达载体pcmv-signalpeptide-seap-xhoi-ha(广东和信健康提供)改造
3.1表达载体pcmv-signalpeptide-seap-linker-mcs-ha构建
(1)在载体pcmv-signalpeptide-seap-xhoi-ha中的xhoi-ha之间插入nhei,得到载体pcmv-signalpeptide-seap-ha-1;
(2)用xhoi/nhei双酶切载体pcmv-signalpeptide-seap-xhoi-ha-1,酶切产物回收待用;
(3)合成引物sali-linker-mcs-nhei,引物退火,以退火产物做为模板进行pcr扩增,得到双链dna,双链pcr产物用sali/nhei酶切,回收待用;
(4)将(2)和(3)中得到的酶切产物按照一定比例连接,转化dh5α,挑单菌落摇过夜,抽提质粒,测序,得到构建好的表达载体pcmv-signalpeptide-seap-linker-mcs-ha,图1为pcmv-signalpeptide-seap-linker-mcs-ha重组表达载体示意图。
3.2表达载体pcmv-signalpeptide-mcs-linker-seap-ha构建
(1)将载体pcmv-signalpeptide-seap-xhoi-ha中seap-ha之间的xhoi突变掉,得到载体pcmv-signalpeptide-seap-ha-1’;
(2)在载体pcmv-signalpeptide-seap-ha-1’中signalpeptide-seap之间插入xhoi酶切位点,得到载体pcmv-signalpeptide-seap-ha-2’;
(3)合成引物xhoi-mcs-linker-sali,引物退火,以退火产物做为模板进行pcr扩增,得到双链dna,双链pcr产物用xhoi/sali酶切,回收待用;
(4)载体pcmv-signalpeptide-seap-ha-2’用xhoi单酶切,回收待用;
(5)将(3)和(4)中得到的酶切产物按照一定比例连接,转化dh5α,挑单菌落摇过夜,抽提质粒,测序,得到构建好的表达载体pcmv-signalpeptide-mcs-linker-seap-ha,图2为pcmv-signalpeptide-mcs-linker-seap-ha重组载体示意图。
4、重组质粒
分别用xhoi/nhei酶切过敏原基因及改造好的载体pcmv-signalpeptide-seap-linker-mcs-ha或者pcmv-signalpeptide-mcs-linker-seap-ha,连接转化至dh5α,做菌落pcr,挑pcr阳性单菌落摇过夜,抽提质粒酶切鉴定,选酶切阳性重组子送测序,获得过敏原重组质粒。
实施例2:过敏原表达的cho稳定细胞系构建
本实施例碱性磷酸酶-过敏原的融合表达,在哺乳细胞cho中进行,在其他实施例里,宿主细胞分别选择昆虫细胞、酵母菌和植物细胞。在本实施例中,构建过敏原表达的cho稳定细胞系。
1、细胞系构建所用培养基、试剂、器材等:
细胞:cho;
培养基:cho无血清培养基、胎牛血清、双抗;
冻存液:含10%dmso的胎牛血清或新生牛血清
抗生素:嘌呤霉素;
胰酶;
转染试剂盒lipofectamine2000;
co2培养箱;
超净工作台;
显微镜等。
2、稳定细胞系构建步骤
2.1嘌呤霉素(puromycin)浓度筛选
(1)生长良好的cho细胞,接种到24孔板,待细胞生长至70-80%;
(2)加入不同浓度梯度(3、6、9μg/ml)的嘌呤霉素进行空细胞致死最低浓度筛选;
(3)培养2天后,细胞全部死亡的最低嘌呤霉素浓度即为后续筛选浓度。
2.2cho转染(puromycin+)
(1)试剂盒抽提重组质粒,使其浓度达到300ng/μl以上;
(2)取plvx与上述重组质粒(摩尔比1:3)共转染生长密度为60%-80%的cho细胞,37℃培养;
(3)上述转染细胞培养过夜后,细胞用胰酶消化,转移至含有6μg/ml嘌呤霉素的完全培养基中,置于37℃,5%co2条件下生长2-3天。
2.3稳定细胞系构建(puromycin+)
(1)将6孔板中细胞消化,转移至2块96孔板中,100μl/孔,培养2天;
(2)若细胞过多则稍微加大嘌呤霉素的浓度,反之减少浓度,调整浓度使细胞生长稳定;
(3)96孔板中细胞培养15-20天后,取上清做elisa鉴定;
(4)选取活性高的孔,消化细胞,转移至24孔板;
(5)生长1-2天后,转12孔板;
(6)生长1-2天后,转6孔板;
(7)待细胞密度长到70-80%,取上清做elisa鉴定。
2.4单克隆筛选(puromycin-)
(1)上述6孔板中细胞计数,稀释;
(2)稀释的细胞传代至96孔板,5个cell/孔;
(3)培养约15天后,待细胞长至80%,做elisa鉴定;
(4)取(3)中鉴定活性高的孔中的细胞稀释传代,重复步骤(2)-(3)三遍;
(5)冻存稳定表达的单克隆细胞。
实施例3:碱性磷酸酶融合过敏原重组蛋白生产
过敏原重组蛋白生产所用培养基、试剂、器材等:
培养基:cho无血清培养基;
器材:摇瓶、摇床;
过敏原重组蛋白发酵步骤如下:
1、复苏筛选得到的稳定细胞系cho;
2、传代至4ml培养基中,生长3天左右,elisa鉴定;
3、消化细胞,传代至10ml培养基中,培养2-3天,加入10ml培养基,使细胞密度不超过1*106个/ml;
4、20ml细胞传代接种至250ml大瓶,继续培养,期间不断加入新鲜培养基,使细胞密度不超过1*106个/ml,使细胞液体积不超过瓶容量的1/3;
5、换500ml培养瓶,重复4.
6、收集上清。
实施例4:昆虫细胞表达载体构建
昆虫细胞表达载体以pfastbaci(购自invitrogen)为例进行改造。目的基因插入酶切位点为ecori/xbai,也可选择其他合适的酶切位点插入。
1、以改造好的pcmv-signalpeptide-seap-linker-mcs-ha和pcmv-signalpeptide-mcs-linker-seap-ha为模板,设计引物对p1、p2,通过pcr扩增seap-linker和linker-seap,引物两端带有酶切位点和起始密码子或终止密码子。
2、引物对p1扩增得到的片段为片段1:mfei-atg-seap-linker-ecori
引物对p2扩增得到的片段为片段2:xbai-linker-seap-tag-nhei
3、将pfastbaci用ecori单酶切回收,片段1用mfei/ecori双酶切回收,按一定比例连接转化dh5α,抽提质粒送测序,筛选出阳性重组质粒pfastbaci-atg-seap-linker-ecori-待插入目的基因-xbai。
4、将pfastbaci用xbai单酶切回收,片段2用xbai/nhei双酶切回收,按一定比例连接转化dh5α,抽提质粒送测序,筛选出阳性重组质粒pfastbaci-ecori-待插入目的基因-xbai-linker-seap-tag,见图3,图3为pfastbaci-ecori-待插入目的基因-xbai-linker-seap-tag的示意图。
实施例5:酵母表达载体构建
本实施例以酵母表达载体以ppiczalphaa(购自invitrogen)为例进行改造。目的基因插入酶切位点为ecori/xbai,也可选择其他合适的酶切位点插入。
1、以改造好的pcmv-signalpeptide-seap-linker-mcs-ha和pcmv-signalpeptide-mcs-linker-seap-ha为模板,设计引物p3、p4,通过pcr扩增得到seap-linker和linker-seap,引物两端带有酶切位点和起始密码子或终止密码子。
2、将ppiczalphaa用ecori单酶切回收,片段3用mfei/ecori双酶切回收,按一定比例连接转化dh5α,抽提质粒送测序,筛选出阳性重组质粒ppiczalphaa-seap-linker-ecori-待插入目的基因-xbai.
3、将ppiczalphaa用xbai单酶切回收,片段4用xbai/nhei双酶切回收,按一定比例连接转化dh5α,抽提质粒送测序,筛选出阳性重组质粒ppiczalphaa-ecori-待插入目的基因-xbai-linker-seap,见图4,图4为4.ppiczalphaa-ecori-待插入目的基因-xbai-linker-seap的示意图。
实施例6:试剂盒检测基本步骤(elisa法或clia法)
1、酶标板包被
材料
抗人ige:用ige免疫小鼠得到腹水,后用辛酸纯化得到抗人ige单抗
包被液:0.05mol/l碳酸缓冲液(ph9.6)
洗涤液:1xtbst(0.05%吐温-20)
封闭液:含0.1%bsa,ph7.00.02mpbs
样品稀释液\酶标稀释液:含10%血清,ph7.00.02mpbst
底物:碱性磷酸酯酶化学发光液
步骤
(1)用包被液将抗人ige稀释成4000ng/ml,混匀,向酶标板孔内加入100ul/孔;
(2)酶标板封膜后4度包被过夜;
(3)用洗涤液洗板1次;
(4)向酶标板孔内加入封闭液,120ul/孔;
(5)4℃封闭过夜;
(6)甩去封闭液,将酶标板干燥后,密封入带干燥剂的金属箔袋内,4℃贮存。
2、人ige标准液的制备
(1)人ige抗体
(2)步骤,用样品稀释液将ige抗体稀释成6个浓度梯度:100、50、17.5、3.5、0.70、0.35iu/ml
3、过敏原特异性ige检测试剂盒的使用方法
(1)已包被的酶标板恢复至室温后,开启包装袋
(2)样品孵育:取人ige标准液和1:10稀释后的血清样品分别加入到酶标板孔内,100ul/孔,用膜封好酶标板,37度下孵育1小时。
(3)用洗涤液洗板5次后拍干。
(4)加入一定浓度的带磷酸酯酶标记的重组过敏原,100ul/孔,封好酶标板,37度孵育30分钟,同上步骤(3)洗板。
(5)底物反应显色:每孔加入40ul碱性磷酸酯酶化学发光液,37度200rpm反应10分钟。
(6)结果计算:计算重复孔的化学发光值
以标准溶液系列的平均吸光度值的对数值对相应浓度的对数值进行线性回归,构建剂量-反应曲线;由样品重复孔的平均吸光度值大于0.165的那个过敏原孔的吸光度值从剂量-反应曲线计算出患者该过敏原的特异性ige抗体含量,并按给定的分级将该过敏原反应定级。
实施例7:试剂盒检测基本步骤(印迹法)
1、硝酸纤维素膜包被
材料
抗人ige:用ige免疫小鼠得到腹水,后用辛酸纯化得到抗人ige单抗
包被液:0.05mol/l碳酸缓冲液(ph9.6)
洗涤液:1xtbst(0.05%吐温-20)
底物:bcip/nbt
步骤
(1)用包被液将抗人ige包被于硝酸纤维素膜上;
(2)37度干燥过夜
(3)密封入带干燥剂的金属箔袋内,常温保存。
2、人ige标准液的制备
(1)人ige抗体
(2)步骤,用样品稀释液将ige抗体稀释成6个浓度梯度:100、50、17.5、3.5、0.70、0.35iu/ml
3、过敏原特异性ige检测试剂盒的使用方法
(1)将包被有抗人ige的硝酸纤维素膜条置于塑料反应槽中;
(2)用洗涤液将硝酸纤维素膜润湿,在检测膜上加入适量血清样品,在振荡器上低速混匀,室温孵育60分钟;
(3)洗涤液将膜清洗3次,每次低速震荡10分钟左右;
(4)加入碱性磷酸酯酶-过敏原融合抗原,在振荡器上低速混匀,室温孵育30分钟;
(5)洗涤同上步骤(3);
(6)加入底物bcip/nbt,在振荡器上低速混匀,室温孵育20分钟;
(7)自来水冲洗终止显色。
实施例8:本法与链霉亲和素-碱性磷酸酯酶标记物两种方法筛查过敏症比较
收集60例虾蟹及59例德国小蠊过敏样本(样本来自广州医科大学附属第一医院),样品-20℃保存备用。
样品检测步骤(以硝酸纤维素膜固相载体为例):
1、将本试剂盒包被有抗人ige的硝酸纤维素膜条及allergyscreen膜条分别置于塑料反应槽中;
2、用洗涤液将硝酸纤维素膜润湿,在检测膜上加入适量血清样品,在振荡器上低速混匀,室温孵育60分钟;
3、洗涤液将膜清洗3次,每次低速震荡10分钟左右;
4、分别加入碱性磷酸酯酶-过敏原融合抗原(本法)及链霉亲和素-碱性磷酸酯酶标记物(allergyscreen),在振荡器上低速混匀,室温孵育30分钟;
5、洗涤同上步骤3;
6、加入底物bcip/nbt,在振荡器上低速混匀,室温孵育20分钟;
7、自来水冲洗终止显色。
8、显色明显的为阳性,不显色为阴性,结果比较如下:
注:两种检测方法定性结果的一致性检验,kappa>=0.75时,两者一致性良好,测量系统较好;0.4<=kappa<=0.7时,两者一致性一般;kappa<0.4时,两者一致性较差,测量系统是不适当的。
从下列两种过敏原检测中可以看出,两种方法对虾蟹过敏原检测的一致性良好,对德国小蠊过敏原检测的一致性一般。本试剂盒检验方法较为可靠。
表1:75例虾蟹过敏原样本定性分析
kappa=0.763
表2:74例德国小蠊过敏原样本定性分析
kappa=0.684
最后所应当说明的是,以上实施例仅用以说明本发明的技术方案而非对本发明保护范围的限制,尽管参照较佳实施例对本发明作了详细说明,本领域的普通技术人员应当理解,可以对本发明的技术方案进行修改或者等同替换,而不脱离本发明技术方案的实质和范围。
- 还没有人留言评论。精彩留言会获得点赞!