跨物种特异性PSMAxCD3双特异性单链抗体的制作方法
本发明涉及双特异性单链抗体分子,其包含能够结合至人类与非黑猩猩灵长类动物cd3ε链的表位的第一结合域,其中该表位是包含在seqidno.2、seqidno.4、seqidno.6以及seqidno.8中的氨基酸序列的一部分,所述双特异性单链抗体分子还包含能够结合至前列腺-特异性膜抗原(psma)的第二结合域。本发明还提供编码所述双特异性单链抗体分子的核酸以及载体与宿主细胞和用于其生产的方法。本发明进一步涉及包含所述双特异性单链抗体分子的药物组合物以及所述双特异性单链抗体分子的医学用途。
背景技术:
t细胞识别由以克隆方式分布的αβ以及γδt细胞受体(tcr)介导,所述t细胞受体与肽mhc(pmhc)的肽负载分子相互作用(davis&bjorkman,nature334(1988),395-402)。tcr的抗原特异性链不具有信号传导结构域,而是被耦联到保守的多亚基的信号传导装置cd3(call,cell111(2002),967-979,alarcon,immunol.rev.191(2003),38-46,malissenimmunol.rev.191(2003),7-27)。将tcr连接直接传送至信号传导装置的机制在t细胞生物学中仍然是一个基本的疑问(alarcon,上述引用文;davis,cell110(2002),285-287)。看起来清楚的是:持续的t细胞反应涉及辅助受体参与、tcr寡聚合以及tcr-pmhc复合物在免疫学突触中的较高次序排列(davis&vandermerwe,curr.biol.11(2001),r289-r291,davis,nat.immunol.4(2003),217-224)。然而,非常早的tcr信号传导在没有这些事件时发生,并且可涉及cd3ε的配体诱导的构象变化(alarcon,上述引用文;davis(2002),上述引用文;gil,j.biol.chem.276(2001),11174-11179;gil,cell109(2002),901-912)。信号传导复合物的ε、γ、δ以及ζ亚基彼此缔合以形成cd3ε-γ异源二聚体、cd3ε-δ异源二聚体以及cd3ζ-ζ同源二聚体(call,上述引用文)。各种研究表明:cd3分子对于αβtcr的适当细胞表面表达和正常的t细胞发育是重要的(berkhout,j.biol.chem.263(1988),8528-8536;wang,j.exp.med.188(1998),1375-1380;kappes,curr.opin.immunol.7(1995),441-447)。小鼠cd3εγ异源二聚体的胞外域片段的溶液结构显示:εγ亚基均是c2组ig结构域,它们相互作用以形成独特的边-对-边的二聚体构型(sun,cell105(2001),913-923)。虽然富含半胱氨酸的茎似乎在驱动cd3二聚时起重要作用(su,上述引用文;borroto,j.biol.chem.273(1998),12807-12816),但是经由cd3ε和cd3γ的胞外域的相互作用足以将这些蛋白质与tcrβ组合(manolios,eur.j.immunol.24(1994),84-92;manolios&li,immunol.cellbiol.73(1995),532-536)。虽然仍存在争议,但是tcr的主要化学计量学最有可能包含一个αβtcr、一个cd3εγ异源二聚体、一个cd3εδ异源二聚体以及一个cd3ζζ同源二聚体(call,上述引用文)。鉴于人cd3εγ异源二聚体在免疫反应中的中心作用,结合至治疗性抗体okt3的这个复合物的晶体结构最近已被阐明(kjer-nielsen,pnas101,(2004),7675-7680)。
许多治疗策略通过靶向tcr信号传导来调节t细胞免疫,特别是在临床上广泛应用于免疫抑制方案的抗人cd3单克隆抗体(mab)。cd3-特异性小鼠mabokt3是第一个被许可使用于人的mab(sgro,toxicology105(1995),23-29),并且在移植(chatenoud,clin.transplant7(1993),422-430;chatenoud,nat.rev.immunol.3(2003),123-132;kumar,transplant.proc.30(1998),1351-1352)、1型糖尿病(chatenoud(2003),上述引用文)以及牛皮癣(utset,j.rheumatol.29(2002),1907-1913)中被广泛地用于临床免疫抑制剂。此外,抗-cd3mab可引起局部的t细胞信号传导以及克隆不应答(smith,j.exp.med.185(1997),1413-1422)。okt3在文献中被描述为有效的t细胞有丝分裂原(vanwauve,j.immunol.124(1980),2708-18)以及有效的t细胞杀手(wong,transplantation50(1990),683-9)。okt3以时间依赖的方式展现这些活性;随着t细胞的早期活化导致细胞因子释放,在进一步管理okt3后阻止所有已知的t细胞功能。正是由于t细胞功能的这一较晚阻断,okt3作为免疫抑制剂广泛应用在用于降低或甚至消除同种异体移植组织排斥的治疗方案中。
okt3最可能通过阻断所有t细胞的功能来改变同种异体移植的组织排斥,这在急性排斥中发挥了重要作用。okt3与人t细胞膜上的cd3复合物反应并阻断cd3复合物的功能,所述cd3复合物与t细胞的抗原识别结构(tcr)结合并且对于信号传导是必要的。哪一个tcr/cd3的亚基由okt3结合已经是许多研究的主题。虽然一些证据指出,okt3对tcr/cd3复合物的ε-亚基有特异性(tunnacliffe,int.immunol.1(1989),546-50;kjer-nielsen,pnas101,(2004),7675-7680)。进一步的证据表明,okt3结合tcr/cd3复合物需要这个复合物的其他亚基存在(salmeron,j.immunol.147(1991),3047-52)。
其他熟知的对cd3分子特异的抗体已列于tunnacliffe,int.immunol.1(1989),546-50。正如上面所指出的,这些cd3特异性抗体能够诱导各种t细胞反应,例如淋巴因子产生(vonwussow,j.immunol.127(1981),1197;palacious,j.immunol.128(1982),337)、增殖(vanwauve,j.immunol.124(1980),2708-18)以及抑制t细胞诱导(kunicka,在“lymphocytetypingii”1(1986),223中)。也就是说,取决于实验情况,cd3特异性单克隆抗体可抑制或诱导细胞毒性(leewenberg,j.immunol.134(1985),3770;phillips,j.immunol.136(1986)1579;platsoucas,proc.natl.acad.sci.usa78(1981),4500;itoh,cell.immunol.108(1987),283-96;mentzer,j.immunol.135(1985),34;landegren,j.exp.med.155(1982),1579;choi(2001),eur.j.immunol.31,94-106;xu(2000),cellimmunol.200,16-26;kimball(1995),transpl.immunol.3,212-221)。
虽然已报道了许多本领域中描述的cd3抗体识别cd3复合物的cd3ε亚基,事实上它们多数结合构象表位,并且,因此只识别在tcr原生环境中的cd3ε。构象表位的特征在于存在两个或更多个在一级序列中分开的离散的氨基酸残基,当多肽折叠成天然蛋白质/抗原时,所述残基在分子的表面上聚到一起(sela,(1969)science166,1365和laver,(1990)cell61,553-6)。在现有技术中描述的cd3ε抗体所结合的构象表位可被分成2个组。在主要的组中,所述表位由2个cd3亚基(例如,cd3ε链、cd3γ或cd3δ链的2个cd3亚基)形成。例如,在一些研究中已发现:最被广泛使用的cd3ε单克隆抗体okt3、wt31、ucht1、7d6和leu-4不结合至单独被cd3-ε链转染的细胞。然而,这些抗体染色细胞由cd3ε加上cd3γ或cd3δ的组合双重转染(tunnacliffe,上述引用文;law,int.immunol.14(2002),389-400;salmeron,j.immunol.147(1991),3047-52;coulie,eur.j.immunol.21(1991),1703-9)。在第二个较小的组中,构象表位在cd3ε亚基本身内形成。这个组的成员例如已针对变性的cd3ε出现的mabapa1/1(risueno,blood106(2005),601-8)。一并考虑,多数在本领域中被描述的cd3ε抗体识别位于两个或更多个cd3亚基上的构象表位。形成这些表位3-维结构的离散的氨基酸残基可因此坐落在cd3ε亚基本身上或在cd3ε亚基和其他cd3亚基(诸如cd3γ或cd3δ)上。
另一个关于cd3抗体的问题是许多cd3抗体已被发现具有物种特异性。抗-cd3单克隆抗体(如一般而言适用于任何其他的单克隆抗体)通过高特异性识别它们的靶分子而起作用。它们只识别在它们的靶cd3分子上的单一位点或表位。例如,一个对cd3复合物特异的最广泛使用且表征最充分的单克隆抗体是okt-3。这个抗体与黑猩猩cd3反应但不与其它灵长类动物(例如猕猴)的cd3同源物或与狗的cd3反应(sandusky等人,j.med.primatol.15(1986),441-451)。相似地,wo2005/118635或wo2007/033230描述人单克隆cd3ε抗体,其与人cd3ε但不与小鼠、大鼠、兔子或非黑猩猩灵长类动物(例如恒河猴、石蟹猕猴或狒狒)的cd3ε反应。抗-cd3单克隆抗体ucht-1也与来自黑猩猩的cd3反应但不与来自猕猴的cd3反应(自有数据)。另一方面,也有识别猕猴抗原但不识别它们的人对应物的单克隆抗体的实例。这个组的一个实例是针对来自猕猴的cd3的单克隆抗体fn-18(uda等人,j.med.primatol.30(2001),141-147)。有趣地,已发现来自大约12%石蟹猕猴的外周血淋巴细胞由于猕猴的cd3抗原的多态性而缺乏与抗-恒河猴的cd3单克隆抗体(fn-18)反应的活性。uda等人描述了相较于与fn-18抗体反应的源自动物的cd3,不与fn-18抗体反应的石蟹猕猴的cd3序列中的2个氨基酸的替换(uda等人,jmedprimatol.32(2003),105-10;uda等人,jmedprimatol.33(2004),34-7)。
识别能力(即,物种特异性,不仅cd3单克隆抗体(以及它们的片段)是固有的,对于一般的单克隆抗体也是固有的)对于所述抗体发展作为用于治疗人类疾病的治疗药物而言是一个重大的阻碍。为了获得上市许可,任何新的候选药物必须经过严格的测试。这个测试可以细分成临床前以及临床阶段,而临床阶段进一步被细分成一般已知的临床i、ii和iii期,在人类患者中进行,临床前阶段在动物中进行。临床前测试的目的是证明候选药物具有所需的活性并且最重要地是安全的。只有当动物的安全性以及该候选药物可能的有效性已在临床测试前建立时,这个候选药物将被批准由各自的监管机构用于人类临床测试。可通过下列3种方式在动物中测试候选药物的安全性:(i)在相关物种中,即在药物候选物可识别同源物抗原的物种中,(ii)在含有人抗原的转基因动物中,以及(iii)通过使用可以结合动物体内存在的同源物抗原的候选药物替代品。转基因动物的限制使得这个技术通常被限制于啮齿类动物。啮齿类动物与人在生理学上有显著的不同,安全性结果不能轻易地外推到人类。候选药物替代品的局限性在于与实际候选药物相比是不同的物质组成,并且使用的动物经常是如上面所讨论的具有局限的啮齿类动物。因此,相比于候选药物,在啮齿类动物中产生的临床前数据的预测能力有限。安全性测试方法的选择是利用相关的物种,优选低等的灵长类动物。本领域描述的适用于人的治疗性干预的单克隆抗体的现有限制是:相关物种是较高等的灵长类动物,特别是黑猩猩。黑猩猩被认为是濒危物种,并且由于它们类似人类的本质,这类动物用于药物安全性测试在欧洲已被禁止,并且在其他地方也被严格限制。为了将细胞毒性t细胞重定向至病理细胞,cd3已成功地被用作双特异性单链抗体的靶标,导致病变细胞从不同的生物体内耗竭(wo99/54440;wo04/106380)。例如,bargou等人(science321(2008):974-7)最近报道的具有临床活性的cd19xcd3双特异性抗体构建体被称为blinatumomab,其具有使人患者的所有细胞毒性t细胞用于癌细胞裂解的潜力。在非霍奇金氏淋巴瘤患者中,使用低至每天每平方米0.005毫克的剂量导致血液中的靶细胞的消除。部分以及完全的肿瘤消退最先在0.015毫克的剂量水平观察到,并且在0.06毫克的剂量水平下治疗的全部7位患者均经历了肿瘤消退。blinatumomab也导致肿瘤细胞从骨髓以及肝中清除。虽然这个研究建立了关于双特异性单链抗体形式在治疗血细胞衍生癌的治疗效力的临床证据或概念,但是仍然需要用于治疗其他类型癌症的成功概念。
在2008年,估计有186,320男性在美国将被新诊断出患有前列腺癌,并且大约28,660男性将死于这种疾病。最近一份关于癌症死亡率的报告显示:在2004年,美国人中前列腺癌的总死亡率为每100,000例有25例。在20世纪80年代末,广泛地采用前列腺-特异性抗原(psa)测试,在前列腺癌的管理上代表了一个重大的改进。这个测试测量血中psa蛋白的量,其在前列腺癌患者中往往是升高的。在1986年,美国食品药物管理局(fda)批准了使用psa测试来监测具有前列腺癌的患者,并且在1994年还批准其用作本病的筛选试验。由于psa测试在美国广泛地实施,约90%的前列腺癌在早期被诊断出,并且,因此患者在诊断后存活更久。然而,需要nci-资助的前列腺、肺、结肠直肠与卵巢(plco)筛查试验以及筛查前列腺癌的欧洲研究(erspc)这2个进行中的临床试验的结果,以确定psa筛查是否能拯救生命。在过去25年中进行着的临床试验已调查了天然与合成的化合物在预防前列腺癌的效力。例如,招收近19,000位健康男性的前列腺癌预防试验(pcpt)发现:被批准用于治疗良性前列腺增生(bph,一种非癌的前列腺肿大)的药物非那雄胺,将罹患前列腺癌的风险减少25%。另一个试验,硒与维生素e癌症预防试验(select)研究超过35,000位男性以确定每天补充硒以及维生素e是否可降低健康男性前列腺癌的发病率。其他预防前列腺癌的试验目前正在评估综合维生素、维生素c与d、大豆、绿茶以及在番茄中被发现的天然化合物番茄红素的保护潜力。一篇在2005年报导的研究显示,特定基因融合于60%至80%的被分析的前列腺肿瘤中。这个研究代表首次在前列腺癌中观察到非随机基因重排。这种基因的改变最终可用作生物标记物,以帮助本病的诊断及可能的治疗。其他研究已表明:在8号染色体特定区域内的遗传变异可以增加人罹患前列腺癌的风险。这些基因变异导致约25%的前列腺癌发生在白人男性中。它们是第一个被确认增加罹患前列腺癌风险的基因变异体,并且可帮助科学家更好地了解这个疾病的遗传原因。也有持续的研究探讨了在患者血液中循环的蛋白质如何可用于改善前列腺以及其他癌症的诊断。在2005年,科学家鉴定了由患者的免疫系统响应于前列腺肿瘤而产生的特定蛋白质。这些蛋白质是一种自身抗体,其能够以大于90%的准确度来检测血液样品中前列腺癌细胞的存在。当与psa组合使用时,这些以及其他的血液蛋白最终可用于减少仅用psa测试获得的假阳性结果的数目,并且因此降低了由于假阳性psa测试结果而每年进行的大量的不必要的前列腺活检。
除了psa之外,已经鉴定了数种用于前列腺癌的其他标记物,包括例如:前列腺的6次跨膜的上皮抗原(steap)(hubert等人,pnas96(1999),14523-14528)、前列腺干细胞抗原(psca)(reiter等人,proc.nat.acad.sci.95:1735-1740,1998)以及前列腺特异性膜抗原(psma;psm)(israeli等人,cancerres.53(1993))。psma起初由源自淋巴结前列腺腺癌(lncap)细胞系的部分纯化的膜制剂免疫的单克隆抗体(mab)7e11定义(horoszewicz等人,anticancerres.7(1987),927-35)。克隆编码psma蛋白的2.65kb的cdna片段,随后映射到染色体11p11.2(israeli等人,上述引用文;o'keefe等人,biochem.biophys.acta1443(1998),113-127)。psma的初步分析证明了在前列腺分泌上皮细胞内的广泛表达。免疫组化染色显示:psma在增生以及良性组织中缺乏至中度表达,而恶性组织以最大强度染色(horoszewicz等人,上述引用文)。后续的研究证实了这些结果,并表明psma表达为目前在几乎每个被检查的前列腺组织的普遍特征。这些报告进一步表明psma的表达与肿瘤侵袭性特别成比例地增加(burger等人,int.j.cancer100(2002),228-237;chang等人,cancerres.59(1999),3192-98;chang等人,urology57(2001),1179-83;kawakami和nakayama,cancerres.57(1997),2321-24;liu等人,cancerres.57(1997),3629-34;lopes等人,cancerres.50(1990),6423-29;silver等人,clin.cancerres.9(2003),6357-62;sweat等人,urology52(1998),637-40;troyer等人,int.j.cancer62(1995),552-558;wright等人,urology48(1996),326-334)。与在psma表达和肿瘤分期之间的相关性一致,psma的水平提高与雄性激素-非依赖性前列腺癌(pca)有关。前列腺癌患者的组织样本分析表明,在身体阉割或雄性激素耗竭的治疗后psma的水平升高。不像前列腺特异性抗原的表达即在雄性激素消除后psma减量调节,在原发性和转移性肿瘤样品中psma表达均显著地增加(kawakami等人、wright等人,上述引用文)。与在雄性激素-非依赖性的肿瘤中表达升高相一致,psma转录也被认为是由类固醇减量调节,并且睾丸激素给药介导psma蛋白以及mrna的水平显著地降低(israeli等人,cancerres.54(1994),1807-11;wright等人,上述引用文)。psma也在继发性前列腺肿瘤以及隐匿性转移疾病中高度表达。免疫组化分析显示:相较于良性前列腺组织,位于淋巴结、骨、软组织和肺的转移病灶内的psma的表达相对激烈和均一(chang等人(2001),上述引用文;murphy等人,cancer78(1996),809-818;sweat等人,上述引用文)。一些报告还显示,在包括肾脏近端小管的亚组、小肠刷状缘膜的一些细胞以及在结肠隐窝的稀少细胞的前列腺外组织中,psma表达有限(chang等人(1999)、horoszewicz等人、israeli等人(1994)、lopes等人、troyer等人,上述引用文)。然而,psma在这些组织中的水平通常是2到3个数量级,小于在前列腺中观察到的(sokoloff等人,prostate43(2000),150-157)。psma也表达于多数被检查的实体癌的肿瘤相关的血管新生系统中,但是在正常的血管内皮中是缺少的(chang等人(1999)、liu等人、silver等人,上述引用文)。虽然psma在血管内表达的显著性是未知的,但是与肿瘤相关的血管内皮细胞的特异性使psma成为用于治疗多种形式的恶性肿瘤的潜在靶标。
虽然已经把许多的精力放在鉴定用于癌症治疗方法的新靶标上,但是癌症仍是最常被诊断的疾病之一。鉴于此,仍然需要对于癌症的有效治疗。
技术实现要素:
本发明提供一种双特异性单链抗体分子,其包含能够结合至人类及非黑猩猩灵长类动物cd3ε链的表位的第一结合域,其中,该表位是包含在seqidno.2、seqidno.4、seqidno.6和seqidno.8中的氨基酸序列的一部分;和能够结合至前列腺-特异性膜抗原(psma)的第二结合域。
虽然本领域描述的t细胞-接合的双特异性单链抗体对于治疗恶性疾病有很大的治疗潜力,但是这些双特异性分子大多数被限制在:它们是物种特异性的并且只识别人类抗原,以及由于基因相似性可能识别黑猩猩对应物。本发明的优点是提供一种双特异性单链抗体,其包含对于人类以及非黑猩猩灵长类动物的cd3ε链显示具有跨物种特异性的结合域。
在本发明中令人惊奇地发现,相比于本领域描述的全部其它已知的cd3ε的表位,当cd3ε的胞外域的n-端1-27个氨基酸残基多肽片段从其在cd3复合物中的原生环境取出时(并且任选地融合到外源氨基酸序列,例如epcam或免疫球蛋白fc部分时),所述多肽片段维持其三维结构完整性。因此,本发明提供一种双特异性单链抗体分子,其包含第一结合域,所述第一结合域能够结合至人类以及至少一种非黑猩猩灵长类动物cd3ε链的cd3ε(cd3ε是,例如,从它的原生环境取出和/或由t-细胞包含的(存在于其表面))的胞外域的n-端1-27个氨基酸残基多肽片段的表位,其中,该表位是包含在seqidno.2、seqidno.4、seqidno.6和seqidno.8中的氨基酸序列的一部分;以及第二结合域,所述第二结合域能够结合至前列腺-特异性膜抗原(psma)。在此处及其它地方被提及的非黑猩猩灵长类动物是优选的。至少一种(或其中所选或全部)灵长类动物选自白鬓狨;绒顶柽柳猴、松鼠猴以及马来猴(seqid631或seqid632或者这两者)是特别优选的。恒河猕猴(也被称为恒河猴)也设想作为另一种优选的灵长类动物。因此设想:本发明的抗体结合至(能够结合至)人类以及白鬓狨、绒顶柽柳猴、松鼠猴和马来猴(seqid631或seqid632或者这两者)、以及任选地恒河猕猴的cd3ε的胞外域的n-端1-27个氨基酸残基多肽片段的环境独立表位。可根据在wo2008/119567(特别是wo2008/119567的实施例2)中的操作获得(可由此获得)或制造如本文定义的包含第一结合域的双特异性单链抗体分子。为此,设想(a)用人类和/或松鼠猴的cd3ε的胞外域的n-端1-27个氨基酸残基多肽片段免疫小鼠;(b)产生免疫鼠抗体scfv文库;(c)通过测试至少结合至seqidno.2、seqidno.4、seqidno.6和seqidno.8的能力来鉴定cd3ε特异性结合物。
本发明所提供的环境独立的cd3表位对应于cd3ε的前27个n-端氨基酸或者这27个氨基酸伸展物的功能性片段。如本文所用,关于cd3表位的用语“环境独立”意指本文描述的本发明的结合分子/抗体分子的结合不会引起构象、序列、在抗原决定簇或表位周围的结构的改变或修饰。相比之下,由常见的cd3结合分子(例如,如公开在wo99/54440或wo04/106380)识别的cd3表位位于环境独立表位的cd3ε链c-端至n-端的1-27个氨基酸上,其中若它被嵌在ε链的其余部分内并通过ε链与cd3γ或δ链的异二聚体化而被保持在正确的空间位置时,它仅采取正确的构象。
如本文提供的作为psmaxcd3双特异性单链分子的一部分的抗-cd3结合域,已在wo2008/119567有描述。针对环境独立的cd3表位产生(且指向)的这些结合域提供了关于t细胞再分布的惊人的临床改善,并且因此提供了更有利的安全性。不被理论所限制,由于cd3表位是环境独立的,形成自给自足的子域而对cd3复合物的其余部分影响不大,因此,相比于识别环境独立的cd3表位的常规cd3结合分子(如wo99/54440或wo04/106380中提供的分子),本文提供的psmaxcd3双特异性单链分子的cd3结合域在cd3构象上诱发较少的异位变化。
在用本发明所述的psmaxcd3双特异性单链抗体治疗的开始阶段期间,由本发明的psmaxcd3双特异性单链抗体的cd3结合域识别的环境独立的cd3表位与t细胞再分布(t细胞再分布等同于t细胞绝对计数的首次下降期和随后的恢复)相关较少或无相关。相比于本领域已知的识别环境依赖的cd3表位的常规cd3结合分子,这些使得本发明的psmaxcd3双特异性单链抗体具有较佳的安全性。特别地,因为t细胞再分布在用cd3结合分子治疗的开始阶段期间是不良事件的主要风险因素,如cns不良事件,所以通过识别环境独立的而不是环境依赖的cd3表位的本发明的psmaxcd3双特异性单链抗体具有很大的安全性,优于本领域已知的cd3结合分子。在用常规cd3结合分子治疗的开始阶段期间具有与t细胞再分布有关的这类cns不良事件的患者通常思维混淆和迷失方向,在一些情况下,还会出现尿失禁。混淆是一种精神状态的改变,其中患者不能够以他或她的正常的清晰水平来思考。患者通常难以集中,并且思考不仅模糊不清而且时常显著地被放慢。在用常规cd3结合分子治疗的开始阶段期间具有与t细胞再分布有关的cns不良事件的患者还可能丧失记忆。频繁地,思维混淆导致丧失识别人、地方、时间或日期的能力。迷失方向的感觉在思维混淆中是常见的,并且做出判断的能力受损。在用常规cd3结合分子治疗的开始阶段期间与t细胞再分布有关的cns不良事件还进一步包括语音模糊不清和/或选字困难。这种障碍可能会削弱语言的表达与理解,以及阅读和写作。除了尿失禁之外,在一些患者中,在用常规cd3结合分子治疗的开始阶段期间与t细胞再分布有关的cns不良事件还可能伴有眩晕以及头昏眼花。
在提及的cd3ε的n-端27个氨基酸多肽片段内的3维结构的维持可用于产生能够以相同的结合亲和力在体外结合至n-端cd3ε多肽片段以及在体内结合至在t细胞上的原生的cd3复合物(的cd3ε亚基)的结合域,优选人类的结合域。这些数据有力地表明:如本文描述的n-端片段形成类似于其体内正常存在的结构的三级构象。进行了关于cd3ε的n-端多肽片段的氨基酸1-27的结构完整的重要性的非常灵敏的测试。cd3ε的n-端多肽片段的氨基酸1-27的各个氨基酸被改变成丙氨酸(丙氨酸扫描),以测试cd3ε的n-端多肽片段的氨基酸1-27对于轻微破坏的灵敏度。cd3特异性结合域作为本发明的psmaxcd3双特异性单链抗体的一部分,用于测试与cd3ε的n-端多肽片段的氨基酸1-27的丙氨酸-突变体的结合(见wo2008/119567)。在片段的n-端处的前5个氨基酸残基以及在cd3ε的n-端多肽片段的氨基酸1-27的第23和25位的2个氨基酸的个别互换,对于这些抗体分子的结合是关键的。将包含残基q(在第1位处的谷氨酰胺)、d(在第2位处的天冬氨酸)、g(在第3位处的甘氨酸)、n(在第4位处的天冬酰胺)以及e(在第5位处的谷氨酸)的第1-5位的区域中的氨基酸残基替换成丙氨酸,消除了本发明的psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体)与所述片段的结合。然而,对于至少一些本发明的psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体),在提及的片段t(在第23位处的苏氨酸)以及i(在第25位处的异亮氨酸)的c-端处的2个氨基酸残基,降低了对本发明的psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体)的结合能。
意外地,已发现如此分离的本发明的psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体),不仅识别人cd3ε的n-端片段,而且识别各种灵长类动物的cd3ε的相应同源片段,所述灵长类动物包括新世界猴(狨猴,白鬓狨;绒顶柽柳猴;松鼠猴)和旧世界猴(马来猴,也被称为石蟹猕猴;或恒河猕猴,也被称为恒河猴)。因此,检测到本发明的psmaxcd3双特异性单链抗体的多灵长类动物特异性。下列的序列分析证实,人类和灵长类动物在cd3ε的胞外域的n-端处共有高度同源的序列延伸物。
前面所提及的cd3ε的n-端片段的氨基酸序列描述于seqidno.2(人类)、seqidno.4(白鬓狨);seqidno.6(绒顶柽柳猴);seqidno.8(松鼠猴);seqidno.631qdgneemgsitqtpyqvsisgttiltc或seqidno.632qdgneemgsitqtpyqvsisgttvilt(马来猴,也被称为石蟹猕猴)以及seqidno.633qdgneemgsitqtpyhvsisgttvilt(恒河猕猴,也被称为恒河猴)。
本发明的psmaxcd3双特异性单链抗体的第二结合域结合至前列腺-特异性膜抗原(psma)。优选地,该psmaxcd3双特异性单链抗体的第二结合域结合至人类psma或非黑猩猩灵长类动物psma;更优选地,它结合至人类psma以及非黑猩猩灵长类动物psma,并且因此是跨物种特异性的;更优选地,结合至人类psma以及猕猴psma(并且因此也是跨物种特异性的)。特别优选地,猕猴psma是石蟹猕猴psma和/或恒河猴psma。然而,不排除在本发明范围外的是:该第二结合域也可结合至其他物种的psma同源物(例如啮齿类动物的psma同源物)。
前列腺癌是男性的第二大癌症。在2008年,估计美国有186,320位男性将被新诊断出患有前列腺癌,并且大约28,660位男性将死于这种疾病。患前列腺癌的风险与年龄强相关:非常少的病例为出现在50岁以下的男性中,而四分之三的病例发生在超过65岁的男性中。最大数目的病例被诊断出现在70-74岁的男性中。目前,老年人口的增长率显著高于总人口。到2025-2030年,预测指出60岁以上人口的增长速度将是总人口的3.5倍。在下半个世纪,预测世界各地老年人的比例将超过2倍,这意味着未来诊断出的前列腺癌的发生率将进一步增加。psma的高度受限表达和它在前列腺癌的发展阶段与转移性疾病中的增量调节以及它在许多不同类型的其他实体瘤的肿瘤血管上作为新抗原的作用使得psma能够作为以抗体为基础的癌症治疗的有吸引力的靶标抗原。正如在下列实施例中所示,本发明的psmaxcd3双特异性单链抗体提供了杀死表达psma的人类癌细胞(如作为例示的人类前列腺癌细胞系lncap)的有利工具。此外,本发明的psmaxcd3双特异性单链抗体的细胞毒活性要比本领域描述的抗体的细胞毒活性高。由于优选本发明的psmaxcd3双特异性单链抗体的cd3和psma结合域均是跨物种特异性的(即,与人类以及非黑猩猩灵长类动物抗原反应),它可用于这些结合域在灵长类动物中以及以相同形式作为人用药物的安全性、活性和/或药代动力学特征的临床前评价。
有利地,本发明还提供包含结合至人类psma以及猕猴psma同源物(即,非黑猩猩灵长类动物的同源物)的第二结合域的psmaxcd3双特异性单链抗体。在一个优选的实施方案中,该双特异性单链抗体因此包含对于人类以及非黑猩猩灵长类动物psma显示具有跨物种特异性的第二结合域。在这种情况下,同样的双特异性单链抗体分子可用于这些结合域在灵长类动物中和作为人用药物的安全性、活性和/或药代动力学特征的临床前评价。换言之,相同分子可用于在临床前动物研究以及人类的临床研究。这使得结果高度可比较,并且相较于物种-特异性替代分子,动物研究的预测能力大幅增加。由于本发明的psmaxcd3双特异性单链抗体的cd3和psma结合域都是跨物种特异性的,即,与人类以及非黑猩猩灵长类动物的抗原反应,其可用于这些结合域在灵长类动物以及以同样的形式作为人用药物的安全性、活性和/或药代动力学特征的临床前评价。将会理解的是:在一个优选的实施方案中,本发明的抗体的第一和第二结合域的跨物种特异性是相同的。
在本发明中已发现,可以产生一种psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体),其中相同的分子可用于临床前动物试验以及临床研究并甚至用于人类的治疗。这是由于意想不到地发现该psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体)除了分别结合至人类cd3ε以及psma之外,(并且由于可能与黑猩猩对应物的基因相似性),还可结合至非黑猩猩灵长类动物(包括新世界猴以及旧世界猴)的所述抗原的同源物。正如下面实施例中所示,本发明的所述psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体)可用作治疗剂来对抗各种疾病,包括、但不限于癌症。该psmaxcd3双特异性单链抗体特别有利地用于治疗癌症,优选实体瘤,更优选癌以及前列腺癌。鉴于上述,需要构建psmaxcd3双特异性单链抗体的替代物,以测试远离亲源(人类)的物种的消失。结果,因为意欲将药物投入到临床试验以及后续的市场许可和治疗药物的给药,将相同的分子用于动物临床前试验。能够使用与日后施用给人类的相同分子来进行临床前动物试验,实际上消除了或者至少大大降低了在临床前动物试验中所获得的数据对人类方案的适用性有限的危险。简而言之,使用与将实际施用给人类的相同分子在动物中获得临床前安全性数据,确实会确保该数据对人类相关方案适用。相比之下,在使用替代分子的常规方法中,这种替代分子必须被以分子方式适应于用于临床前安全评价的动物试验体系。因此,用于人类治疗的分子实际上在序列上以及可能在结构上不同于用于临床前试验药代动力学参数和/或生物活性的替代分子,结果是在临床前动物试验中所获得的数据对于人类方案的适用性/可转移性有限。替代分子的使用需要构建、生产、纯化以及表征全新的构建体。这将导致需要额外的开发成本和时间来获得该分子。总之,除了用于人类治疗的实际药物,还得单独开发替代物,故需要进行2种分子的2个开发生产线。因此,显示具有跨物种特异性的本发明的psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体)的一个主要优点是,可以将相同的分子用作人类以及临床前动物试验中的治疗剂。
优选地,所述本发明的双特异性单链抗体的第一或第二结合域的至少一者是cdr-移植的、人源化的或人类的,如下文更详细所述。优选地,所述本发明的双特异性单链抗体的第一以及第二结合域均是cdr-移植的、人源化的或人类的。对于本发明的psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体),对人类患者给药时,抵抗所述结合分子的免疫反应的产生对抗被最大可能程度地排除。
本发明的psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体)的另一个主要优点是,它适用于各种灵长类动物的临床前试验。理想的候选药物在动物中的作用应该能够反应出该候选药物施用给人类后的预期作用。结果,因此从此类临床前试验获得的数据一般而言应该具有对人类方案的高度预测力。然而,从最近的tgn1412(cd28单克隆抗体)i期临床试验的不幸结果可知,候选药物在灵长类动物物种和人类中有不同作用:在石蟹猕猴的动物研究中已观察到所述抗体在临床前试验中没有或仅有有限的不良反应,但是6位人类患者在施用所述抗体后发展为多脏器功能衰竭(lancet368(2006),2206-7)。这些剧烈的、非预期负面事件的结果表明,将临床前试验限制于仅一种(非黑猩猩灵长类动物)物种未必是足够的。事实上,本发明的psmaxcd3双特异性单链抗体结合至一系列的新世界猴和旧世界猴,可有助于克服上述例子中所面临的问题。因此,本发明提供在开发和测试用于人类治疗的药物时将物种差异的影响最小化的手段和方法。
关于本发明的psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体),使测试动物适应于意欲施用给人类的候选药物,例如,建立转基因动物,也不再是必要的。根据本发明的用途和方法显示具有跨物种特异性的本发明psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体)可直接用于非黑猩猩灵长类动物的临床前试验,没有动物的任何遗传操作。正如本领域技术人员熟知的,使测试动物适应于候选药物的方法经常承受如下风险:在临床前安全性测试中获得的结果较少有代表性,并且由于该动物的修饰对于人类的预测力较低。例如,在转基因动物中,由转基因编码的蛋白质经常被高度过表达。因此,所获得的抗体对抗该蛋白质抗原的生物活性的数据可能在它们对人类(该蛋白质在人中以更低、更生理学的水平表达)的预测值方面有限。
显示具有跨物种特异性的本发明的psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体)的用途的另一个优点是以下事实:黑猩猩作为一种濒危物种,被避免用于动物试验。黑猩猩是人类最接近的亲属,并且最近根据基因组测序数据而被分入人科(wildman等人,pnas100(2003),7181)。因此,用黑猩猩获得的数据通常被认为是对于人类高度预测的。然而,由于它们的状态为濒危物种,可用于医学实验的黑猩猩的数目被高度限制。如上所述的,因此,维持黑猩猩用于动物试验是昂贵并且具有伦理问题的。本发明的psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体)的使用避免了在临床前试验期间的伦理异议以及财政负担,并且没有损害质量(即,所获得的动物试验数据的适用性)。鉴于此,本发明的优选人类的psmaxcd3双特异性单链抗体的使用在黑猩猩研究上提供了一个合理选择。
本发明的psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体)的又一优点是,当用它作为动物临床前试验(例如在动物药代动力学研究的过程中)的一部分时,提取多个血液样本的能力。非黑猩猩灵长类动物要比低等动物(例如,小鼠)更容易进行多次血液提取。多个血液样品的提取允许持续测试血液参数,以测定由本发明的psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体)诱导的生物效应。再者,多个血液样品的提取使研究者能够评估如本文定义的本发明的psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体)的药代动力学特征。此外,反映在血液参数中的可由所述本发明的psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体)诱导的可能副作用,可在施用所述抗体过程期间提取的不同血液样品中进行测量。这允许测定如本文定义的本发明的psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体)的可能毒性。
显示具有跨物种特异性的如本文定义的本发明的psmaxcd3双特异性单链抗体(优选人类的psmaxcd3双特异性单链抗体)的优点,可被简要地概述如下:
第一,用于临床前试验的如本文定义的本发明的(优选人类的)psmaxcd3双特异性单链抗体与用于人类患者的抗体是相同的。因此,不再需要开发在药代动力学性质以及生物活性上可能不同的2个独立分子。这是高度有利的,原因在于,例如相比于传统替代方法,所述药代动力学结果能够直接转移和应用于人类情况。
第二,与替代方法相比,将如本文定义的本发明的(优选人类的)psmaxcd3双特异性单链抗体用于制备人类的治疗药物时,成本和劳动密集度更低。
第三,如本文定义的本发明的(优选人类的)psmaxcd3双特异性单链抗体可用于的临床前试验不仅为在一种灵长类动物物种中,而且在一系列不同的灵长类动物物种中,从而限制了灵长类动物和人类之间潜在的物种差异的风险。
第四,如果必需,作为濒危物种的黑猩猩可避免用于动物试验。
第五,可提取多个血液样本以用于广泛的药物动力学研究。
第六,由于根据本发明的一个优选实施方案的结合分子(优选人类的结合分子)的人类来源,当给人类患者施用时,产生的对抗所述结合分子的免疫反应被降到最低。排除了诱导出对源自非人类物种的候选药物特异性的抗体的免疫反应,所述非人类物种例如导致出现对抗鼠来源的治疗分子的人类抗小鼠抗体(hama)的小鼠。
最后但并非最不重要的是,本发明的psmaxcd3双特异性单链抗体的治疗用途提供一种新颖、有创造性的用于癌症的治疗方法,所述癌症优选为实体瘤,更优选为癌和前列腺癌。如在下述实施例中所示的,本发明的psmaxcd3双特异性单链抗体为杀死表达psma的人类前列腺癌细胞提供了有利的工具。再者,本发明的psmaxcd3双特异性单链抗体的细胞毒活性要比本领域描述的抗体的活性高。
正如上述指出的,本发明提供多肽,即,双特异性单链抗体,其包含能够结合至人类及非黑猩猩灵长类动物cd3ε链的表位的第一结合域以及能够结合至psma的第二结合域。所述第二结合域优选地结合至人类psma以及非黑猩猩灵长类动物psma。双特异性单链抗体分子作为实现本发明的优选双特异性单链抗体要求的候选药物,其优点是此类分子用于临床前动物试验以及临床研究甚至用于人类的治疗。在本发明的跨物种特异性的双特异性单链抗体的一个优选实施方案中,该结合至psma的第二结合域是人类的。在根据本发明的跨物种特异性的双特异性分子中,该结合至人类与非黑猩猩灵长类动物cd3ε链的表位的结合域以vh-vl或vl-vh的次序位于该双特异性分子的n-端或c-端处。在所述第一和第二结合域中呈vh-以及vl-链不同排列的根据本发明的跨物种特异性的双特异性分子,其实例描述于所附的实施例中。
如本文所用的,“双特异性单链抗体”表示包含2个结合域的单多肽链。每个结合域包含一个来自抗体重链(“vh区域”)的可变区域,其中,所述第一结合域的vh区域特异性地结合至cd3ε分子,所述第二结合域的vh区域特异性地结合至psma。2个结合域任选地由短间隔多肽彼此相连。间隔多肽的非限制性实例是gly-gly-gly-gly-ser(ggggs)及其重复序列。每个结合域可另外包含一个来自抗体轻链(“vl区域”)的可变区域,在所述第一和第二结合域内的该vh区域和vl区域经由连接多肽彼此相连,例如在ep623679b1中所陈述和要求的类型,但在任何情况下长到足以允许所述第一结合域的vh区域和vl区域以及所述第二结合域的vh区域和vl区域彼此配对,以使得它们能够一起特异性地结合至相应的第一和第二结合域。
术语“蛋白质”是本领域熟知的,并且描述生物化合物。蛋白质包括一或多个氨基酸链(多肽),从而氨基酸经由肽键彼此结合。如本文所述的,术语“多肽”描述由超过30个氨基酸组成的分子类别。依据本发明,只要蛋白质由单链多肽组成,则多肽类别包括“蛋白质”。此外,符合该定义,术语“多肽”描述蛋白质的片段,只要这些片段由超过30个氨基酸组成即可。多肽可进一步形成多聚体,如二聚体、三聚体和较高级的寡聚物(即,由一个以上的多肽分子组成)。形成此类二聚体、三聚体等的多肽分子可以是相同的或不同的。此类多聚体的对应的高阶结构相应称为同源-或异源二聚体、同源-或异源三聚体等。异源多聚体的实例是抗体分子,其自然发生形式由2条相同的轻多肽链和2条相同的重多肽链组成。术语“多肽”和“蛋白质”也指经天然修饰的多肽/蛋白质,其中所述修饰通过如翻译后修饰(例如糖基化、乙酰化、磷酸化等等)进行。此类修饰是本领域熟知的。
根据本发明,术语“结合域”表征多肽的特异性地结合至/相互作用于给定靶标结构/抗原/表位的结构域。因此,该结合域是“抗原相互作用位点”。根据本发明,术语“抗原相互作用位点”定义多肽的基序,该基序能够特异性地与特定抗原或特异抗原组(例如,在不同物种中的相同抗原)相互作用。所述结合/相互作用也理解为定义“特异性识别”。根据本发明,术语“特异性地识别”指抗体分子能够特异性地相互作用于和/或结合至抗原的至少2个氨基酸,优选至少3个氨基酸,更优选至少4个氨基酸,例如如本文定义的人类cd3抗原。此类结合可以“锁钥原理”的特异性为例。因此,在结合域的氨基酸序列中的特定基序和抗原彼此结合,以形成它们的一级、二级或三级结构以及导致该结构的二级修饰。抗原相互作用位点与其特异性抗原的特异性相互作用也可导致所述位置与抗原的简单结合。再者,结合域/抗原相互作用位点与其特异性抗原的特异性相互作用或者导致信号的启动,例如,起因于诱到抗原构象形的改变、抗原的寡聚合等。与本发明相符的结合域的优选实施例是抗体。该结合域可以是单克隆抗体或多克隆抗体或者源自单克隆抗体或多克隆抗体。
术语“抗体”包含仍然维持结合特异性的衍生物或功能性片段。生产抗体的技术是本领域熟知的,并且描述于,例如harlow和lane“antibodies,alaboratorymanual”,coldspringharborlaboratorypress,1988;harlow和lane“usingantibodies:alaboratorymanual”coldspringharborlaboratorypress,1999以及little“recombinantantibodiesforimmunotherapy”cambridgeuniversitypress2009。术语“抗体”还包括不同类别的免疫球蛋白(ig's)(如,iga、igg、igm、igd和ige)及其亚类(如igg1、igg2等)。
术语“抗体”的定义还包括如嵌合的、单链和人源化抗体以及抗体片段(尤其像fab片段)的实施方案。抗体片段或衍生物进一步包括f(ab')2、fv、scfv片段或单结构域抗体、单可变结构域抗体或者仅包含一个可变结构域(可以是特异性地结合至抗原或表位而无关其它v区域或结构域的vh或vl)的免疫球蛋白单可变结构域;参见,例如,harlow和lane(1988)与(1999)以及little(2009),上述引用文。所述免疫球蛋白单一可变结构域不仅包括经分离的抗体单可变结构域多肽,而且包括含有抗体单可变结构域多肽序列的一个或多个单体的较大多肽。
各种操作程序是本领域已知的,并且可被用于生产此类抗体和/或片段。因此,所述(抗体)衍生物也可通过模拟肽生产。进一步地,所述生产单链抗体的技术(尤其是参见美国专利4,946,778)可适用于生产对选定多肽具有特异性的单链抗体。此外,转基因动物可被用于表达对本发明的多肽和融合蛋白质特异性的人源化或人类抗体。为了制备单克隆抗体,可以使用提供由连续细胞系培养产生的抗体的任何技术。这类技术的实例包括杂交瘤技术(
如根据本发明所用的术语“特异性相互作用”指结合域不或不显著地与多肽交叉反应,所述多肽具有与被结合域结合的那些多肽类似的结构并且由与目标多肽相同的细胞表达。可以测试处于调查中的一组结合域的交叉反应,例如通过评估在常规条件下所述组结合域的结合(参见,例如,harlow和lane(1988)与(1999)以及little(2009),上述引用文。结合域与特异性抗原的特异性相互作用的实例包括配体对其受体的特异性。所述定义特别包括配体的相互作用,在结合至其特异性受体时配体诱发信号。所述定义还特别包括的所述相互作用的实例,为抗原决定簇(表位)与抗体的结合域(抗原结合位点)的相互作用。
如本文所用的术语“跨物种特异性”或“物种间特异性”指本文描述的结合域与人类以及非黑猩猩灵长类动物中的相同靶标分子结合。因此,“跨物种特异性”或“物种间特异性”被理解为对在不同物种中表达的相同分子“x”而非对非“x”的分子的物种间反应性。例如,可通过facs分析测定识别例如人类cd3ε的单克隆抗体对非黑猩猩灵长类动物cd3ε(例如猕猴cd3ε)的跨物种特异性。所述facs分析以下述方式进行:测试各个单克隆抗体与分别表达所述人类和非黑猩猩灵长类动物cd3ε抗原的人类和非黑猩猩灵长类动物细胞(例如,猕猴细胞)的结合。适当的分析将示于下面的实施例中。上面所提及的主题物比照应用于psma抗原:例如,可通过facs分析测定识别例如人类psma的单克隆抗体对非黑猩猩灵长类动物psma(例如猕猴psma)的跨物种特异性。所述facs分析以下述方式进行:测试各个单克隆抗体与分别表达所述人类和非黑猩猩灵长类动物psma抗原的人类和非黑猩猩灵长类动物细胞(例如,猕猴细胞)的结合。
如本文所用的,cd3ε代表作为t细胞受体的一部分表达的分子并且具有如通常将其归属在现有技术中的含义。在人类,它包括个别的或独立地组合形式的所有已知的cd3亚基,例如,cd3ε、cd3δ、cd3γ、cd3ζ、cd3α和cd3β。如本文所指的非黑猩猩灵长类动物的非人类cd3抗原是例如马来猴cd3以及恒河猕猴cd3。在马来猴中,它包括cd3εfn-18阴性和cd3εfn-18阳性、cd3γ以及cd3δ。在恒河猕猴中,它包括cd3ε、cd3γ以及cd3δ。优选地,如本文所用的所述cd3是cd3ε。
人类cd3ε显示于genbank登录号nm_000733并且包含seqidno.1。人类cd3γ显示于genbank登录号nm_000073。人类cd3δ显示于genbank登录号nm_000732。
马来猴的cd3ε“fn-18阴性”(即,因为如上面所提到的多态性而不被单克隆抗体fn-18识别的cd3ε)被显示在genbank登录号ab073994。
马来猴的cd3ε“fn-18阳性”(即,被单克隆抗体fn-18识别的cd3ε)显示于genbank登录号ab073993。马来猴的cd3γ显示于genbank登录号ab073992。马来猴的cd3δ显示于genbank登录号ab073991。
恒河猕猴的各个cd3ε、γ以及δ同源物的核酸序列以及氨基酸序列可通过本领域描述的重组技术(sambrook等人molecularcloning:alaboratorymanual;coldspringharborlaboratorypress,3rdedition2001)进行鉴定和分离。这比照适用于如本文定义的其他非黑猩猩灵长类动物的cd3ε、γ和δ同源物。白鬓狨、松鼠猴以及绒顶柽柳猴的氨基酸序列的鉴定描述于所附的实施例中。白鬓狨的cd3ε胞外域的氨基酸序列显示于seqidno:3,绒顶柽柳猴的氨基酸序列显示于seqidno:5,松鼠猴的氨基酸序列显示于seqidno:7。
人类psma显示于genbank登录号'ay101595'。猕猴的psma同源物的克隆描述于下列的实施例中,对应的cdna和氨基酸序列分别显示于seqidno.223以及seqidno.224。
与上面一致地,术语“表位”定义由如本文定义的结合域特异性地结合/鉴定的抗原决定簇。该结合域可特异性地结合至/相互作用于构象或连续的表位,所述表位对于靶标结构是特有的,例如人类和非黑猩猩灵长类动物cd3ε链或者人类和非黑猩猩灵长类动物psma。多肽抗原的构象或不连续表位的特征在于:存在2个或更多个在一级序列中分开的离散的氨基酸残基,当该多肽折叠成天然蛋白质/抗原时,所述残基在该分子的表面上聚在一起(sela,(1969)science166,1365和laver,(1990)cell61,553-6)。促成所述表位的2个或更多个离散的氨基酸残基存在于一个或多个多肽链的分离部分。当所述多肽链折叠成3维结构以构成该表位时,这些残基在该分子的表面上聚在一起。相比之下,连续或线性表位由2个或更多个离散的氨基酸残基组成,该氨基酸残基存在于多肽链的单一线性区段。在本发明中,“环境依赖的”cd3表位指所述表位的构象。此位于cd3的ε链的环境依赖表位,若它被嵌在ε链的其余部分内并通过ε链与cd3γ或δ链的异二聚体化而被保持在正确位置时,仅发展它的正确构象。相比之下,如本文提供的环境独立的cd3表位指cd3ε的n-端1-27个氨基酸残基多肽或其功能性片段。在将此n-端1-27个氨基酸残基多肽或其功能性片段从其在cd3复合物的天然环境中取出时,维持其3维结构完整性和正确的构象。因此,该n-端1-27个氨基酸残基多肽或其功能性片段(作为cd3ε胞外域的一部分)的环境独立性提供了完全不同于在wo2004/106380中结合用于制备人类结合分子的方法描述的cd3ε的表位。所述方法采用单独表达的重组cd3ε。这个单独表达的重组cd3ε的构象不同于在其天然形式下采取的构象,所述天然形式即tcr/cd3复合物的cd3ε亚基作为tcr/cd3复合物的cd3δ或cd3-γ亚基的非共价复合物一部分存在的形式。当此单独表达的重组cd3ε蛋白用作用于选择来自抗体文库的抗体的抗原时,对该抗原特异性的抗体从该文库中被识别出来,虽然此文库不含具有对自身抗原/自体抗原特异性的抗体。这是由于以下事实:单独表达的重组cd3ε蛋白质不在体内存在;它不是自体抗原。因此,表达对这个蛋白质特异性的抗体的b细胞亚群在体内没有被耗竭;从该类b细胞构建的抗体文库将含有对单独表达的重组cd3ε蛋白特异性的抗体的遗传材料。
然而,由于该环境独立的n-端1-27个氨基酸残基多肽或其功能性片段是以其天然形式折叠的表位,与本发明一致的结合域不能通过基于在wo2004/106380中描述的手段的方法进行鉴定。因此,可能会在测试中证实,如公开于wo2004/106380的结合分子不能结合至cd3ε链的n-端1-27个氨基酸残基。因此,传统的抗-cd3结合分子或抗-cd3抗体分子(例如,如公开于wo99/54440中)相比于本文提供的环境独立的n-端1-27个氨基酸残基多肽或功能性片段,在更靠近c-端的位置处结合cd3ε链。现有技术中的抗体分子okt3和ucht-1也对tcr/cd3复合物的在氨基酸残基35至85的ε-亚基具有特异性,并且这些抗体的表位也更靠近c-端。此外,ucht-1结合的cd3ε链位于氨基酸残基43至77的区域(tunnacliffe,int.immunol.1(1989),546-50;kjer-nielsen,pnas101,(2004),7675-7680;salmeron,j.immunol.147(1991),3047-52)。因此,现有技术的抗-cd3分子没有结合至并且没有指向对抗本文定义的环境独立的n-端1-27个氨基酸残基表位(或其功能性片段)。特别地,本领域的技术状态无法提供特异性结合至环境独立的n-端1-27个氨基酸残基表位并且是跨物种特异性(即,结合至人类和非黑猩猩灵长类动物cd3ε)的抗-cd3分子。
为了产生包含于本发明的双特异性单链抗体分子的结合域(优选人类的结合域),可使用例如结合至人类和非黑猩猩灵长类动物cd3ε(例如猕猴cd3ε)的单克隆抗体或者结合至人类和非黑猩猩灵长类动物psma的单克隆抗体。
如本文所用的,“人类(human)”以及“人”(man)指物种智人。就本文描述的构建体的医学用途而言,人类患者用相同的分子进行治疗。
优选地,所述本发明的双特异性单链抗体的第一或第二结合域的至少一者是cdr-移植的、人源化的或人类的。优选地,所述本发明的双特异性单链抗体的第一以及第二结合域是cdr-移植的、人源化的或人类的。
如本文所用的,术语“人类”抗体被理解意为:如本文定义的双特异性单链抗体包括人类生殖细胞系抗体谱所含有的氨基酸序列。出于本文定义的目的,如果双特异性单链抗体由此类人类生殖细胞系氨基酸序列组成的话,即,若要考虑的双特异性单链抗体的氨基酸序列与表达的人类生殖细胞系氨基酸序列相同的话,所述双特异性单链抗体可因此被视为人类的。如果如本文定义的双特异性单链抗体由偏离它的(它们的)最亲近的人类生殖细胞系序列的序列不超过预期(由体细胞超突变的印记导致的)的序列组成,所述双特异性单链抗体也可被视为人类的。此外,许多非人类哺乳动物(例如啮齿类动物,如小鼠和大鼠)的抗体包含其中一个可预期也存在于表达的人类抗体谱的vhcdr3氨基酸序列。出于本发明的目的,可预期存在于表达的人类谱的人类或非人类来源的任何此类序列也被视为“人类的”。
如本文所用的,术语“人源化的”、“人源化”、“类人的”或其语法上相关的变体可在涉及双特异性单链抗体时被互换使用,该双特异性单链抗体包含:在其至少一个结合域中的至少一个互补决定区(“cdr”),所述cdr来自非人类抗体或其片段。人源化手段描述于例如wo91/09968和us6,407,213中。作为非限制性实例,该术语包括以下情况:其中至少一个结合域的可变区包括来自另一非人类动物(如啮齿类动物)的单cdr区域,例如vh的第三cdr区域(cdrh3);以及以下情况:其中可变区之一或二者在其相应的第一、第二和第三cdr中的每一个处都包含来自所述非人类动物的cdr。在该双特异性单链抗体的结合域的所有cdr已由来自例如啮齿类动物的相应等价物替换时,典型地叫法为“cdr-移植”,并且这个术语被理解为由如本文所用的术语“人源化的”或其语法上相关的变体包括。术语“人源化的”或其语法上相关的变体还包含以下情况:其中除了在该第一和/或第二结合域的vh和/或vl内替换一个或多个cdr区域之外,还在cdr之间的框架(“fr”)区域内已产生至少一个单一氨基酸残基的进一步突变(例如替换),使得在那个/那些位置处的氨基酸对应于在动物(用于替换的cdr区域源自所述动物)的那个/那些位置处的氨基酸。如本领域已知的,为了恢复用作其靶标分子的cdr-供体的非人类抗体的初始结合亲合力,经常在cdr-移植后在框架区域内进行此类个别突变。除了上述的框架区域中的氨基酸替换外,术语“人源化的”可进一步包括将来自非人类动物的cdr区域的氨基酸替换成来自人类抗体的对应的cdr区域的氨基酸的氨基酸替换。
如本文所用的,术语“同源物”或“同源”应被理解为如下:在蛋白质和dna之间的同源性经常根据序列相似性而被断定,特别是在生物信息学中。例如,一般而言,若2个或更多个基因具有高度相似的dna序列,则它们很可能是同源的。但是序列相似性可从不同的祖先产生:短序列的相似可能是偶然的,而且序列相似可能是因为这两者均被选择结合至特定蛋白质,例如转录因子。这样的序列是相似的,但没有同源性。同源的序列区域也被称为保守的。这不与氨基酸序列的保守相混淆,在后者情况下,特定位置处的氨基酸已改变但氨基酸的物理化学性质保持不变。同源的序列有2种类型:直系同源和旁系同源。如果同源的序列因为物种形成事件而被分开,则它们是直系同源的:当物种分化成2个分开的物种时,在所形成的物种中的单一基因的分化副本被称为是直系同源的。直系同源物或直系同源基因是彼此相似的不同物种中的基因,因为它们源自相同的祖先。2个相似的基因是直系同源的最有力的证据是基因谱系的系统发育分析的结果。在一个进化枝内发现的基因是直系同源物,来自相同的祖先。直系同源物经常、但非总是具有相同的功能。直系同源序列给生物体的分类学分类研究提供有用的信息。遗传分化的图谱可用于追踪生物体的关系。关系非常接近的2个生物体在2个直系同源物之间可能显示非常相似的dna序列。相比之下,在进化上从另一个生物体进一步进化的生物体可能在研究的直系同源物的序列上显示出较大分化。如果同源的序列因为基因复制事件而被分开,则它们是旁系同源的:若生物体的基因被复制到占据相同基因组的2个不同的位置,那么这2个拷贝是旁系同源的。旁系同源的一组序列称为互相为旁系同源物。旁系同源物通常具有相同或相似的功能,但有时候则不是这样:由于在重复基因的一个拷贝上缺乏初始的筛选压力,因此这个拷贝自由突变并且获得新功能。实例可见于啮齿类动物(如大鼠和小鼠)。啮齿类动物具有一对旁系同源的胰岛素基因,虽然不清楚是否已发生功能上的任何分化。旁系同源基因常常属于相同物种,但这不是必需的:例如,人类的血红蛋白基因与黑猩猩的肌红蛋白基因是旁系同源物。这在生物信息学上是一个常见的问题:当已测序不同物种的基因组并且已发现同源基因时,不能立即推断这些基因具有相同或相似的功能,因为它们可能是功能已分化的旁系同源物。
如本文所用的,“非黑猩猩灵长类动物(non-chimpanzeeprimate或non-chimpprimate)”或其语法上的变体是指不是黑猩猩的任何灵长类动物(即,非人类)(即,不是属于黑猩猩属的动物),并且包括侏儒黑猩猩(panpaniscus)以及黑猩猩(pantroglodytes)(也被称为anthropopithecustroglodytes或猩猩(simiasatyrus)物种。然而,将被理解的是:本发明的抗体还可以以它们的第一和/或第二结合域结合至所述黑猩猩的相应表位/片段等。如果需要,本发明仅避免用黑猩猩实施的动物试验。因此在另一个实施方案中,还设想的是:本发明的抗体还可以以其第一和/或第二结合域结合至黑猩猩的相应表位。“灵长类动物(primate或primates”、“灵长类动物物种(primatespecies)”或其语法上的变体代表被分成原猴类和类人猿类的2个亚目的真哺乳亚纲哺乳动物的一目,并且包括猿、猴以及狐猴。特别地,如本文使用的“灵长类动物”包括原猴亚目(非眼镜猴原猴),包括下目lemuriformes(它本身包括超科鼠狐猴以及狐猴)、下目chiromyiformes(它本身包括猴科)以及下目lorisiformes(它本身包括懒猴和婴猴科)。如本文使用的“灵长类动物”还包括简鼻亚目,包括下目tarsiiformes(它本身包括眼镜猴科)、下目simiiformes(它本身包括广鼻类或新世界猴以及狭鼻类,狭鼻类包括猴科或旧世界猴)。
在本发明的含义内,非黑猩猩灵长类动物物种可被理解为是狐猴、眼镜猴、长臂猴、一绒猿(属于卷尾猴科的新世界猴)或旧世界猴(属于猴超科)。
如本文所用的,“旧世界猴”包括任何落在猴超科内的猴子,猴超科本身被细分成:猴亚科,主要是非洲属但包括亚洲和北非的猕猴的分化属;和疣猴亚科,它包括大部分的亚洲属和非洲疣猴。
特别地,在猴亚科内,有益的非黑猩猩灵长类动物可来自tribecercopithecini,在短肢猴属(genusallenopithecus)(菲氏叶猴(allen'sswampmonkey),短肢猴(allenopithecusnigroviridis))内;在侏长尾猴属(genusmiopithecus)(侏长尾猴(angolantalapoin),侏长尾猴(miopithecustalapoin);加蓬侏长尾猴(gabontalapoin),加蓬侏长尾猴(miopithecusogouensis))内;在赤猴属(genuserythrocebus)(赤猴(patasmonkey),赤猴(erythrocebuspatas))内;在绿猴属(genuschlorocebus)(绿猴(greenmonkey),绿猴(chlorocebussabaceus);素领猴(grivet),绿猴(chlorocebusaethiops);贝尔山绿猴(balemountainsvervet),贝尔山绿猴(chlorocebusdjamdjamensis);坦塔梦斯绿猴(tantalusmonkey),坦塔梦斯绿猴(chlorocebustantalus);黑长尾猴(vervetmonkey),黑长尾猴(chlorocebuspygerythrus);布鲁克绿猴(malbrouck),布鲁克绿猴(chlorocebuscynosures)]内;或者在长尾猴属(genuscercopithecus)(德赖斯长尾猴(dryasmonkey)或salongomonkey,德赖斯长尾猴(cercopithecusdryas);黛安娜长尾(dianamonkey),黛安娜长尾(cercopithecusdiana);加纳长尾猴(rolowaymonkey),罗洛威须长尾猴(cercopithecusroloway);大白鼻猴(greaterspot-nosedmonkey),大白鼻长尾猴(cercopithecusnictitans);蓝猴(bluemonkey),青长尾猴(cercopithecusmitis);银长尾猴(silvermonkey),银长尾猴(cercopithecusdoggetti);金长尾猴(goldenmonkey),金长尾猴(cercopithecuskandti);赛克斯猴(sykes'smonkey),白喉长尾猴(cercopithecusalbogularis);摩那猴(monamonkey),白额长尾猴(cercopithecusmona);坎氏摩那猴(campbell'smonamonkey),坎氏长尾猴(cercopithecuscampbelli);洛氏摩那猴(lowe'smonamonkey),洛氏长尾猴(cercopithecuslowei);冠毛长尾猴(crestedmonamonkey),冠毛长尾猴(cercopithecuspogonias);狼莫纳猴(wolf'smonamonkey),邬氏长尾猴(cercopithecuswolfi);丹氏长尾猴(dent'smonamonkey),丹氏长尾猴(cercopithecusdenti);小白鼻长尾猴(lesserspot-nosedmonkey),小白鼻长尾猴(cercopithecuspetaurista);白领青长尾猴(white-throatedguenon),赤腹长尾猴(cercopithecuserythrogaster),斯氏长尾猴(sclater'sguenon),斯氏长尾猴(cercopithecussclateri);红耳长尾猴(red-earedguenon),红耳长尾猴(cercopithecuserythrotis);髭长尾猴(moustachedguenon),髭长尾猴(cercopithecuscephus);红尾长尾猴(red-tailedmonkey),红尾长尾猴(cercopithecusascanius);尔氏长尾猴(l'hoest'smonkey),尔氏长尾猴(cercopithecuslhoesti);高山长尾猴(preuss'smonkey),高山长尾猴(cercopithecuspreussi);阳光长尾猴(sun-tailedmonkey),阳光长尾猴(cercopithecussolatus);哈姆林氏猴(hamlyn'smonkey)或枭面猴(owl-facedmonkey),枭面长尾猴(cercopithecushamlyni);德氏长尾猴(debrazza'smonkey),白臀长尾猴(cercopithecusneglectus))内。
或者,有益的非黑猩猩灵长类动物(也在猴亚科以及在tribepapionini内)可来自在猕猴属(genusmacaca)(巴巴利猕猴(barbarymacaque),巴巴利猕猴(macacasylvanus);狮尾猴(lion-tailedmacaque),狮尾猴(macacasilenus);南方豚尾猕猴(southernpig-tailedmacaque)或豚尾猕猴(beruk),豚尾猕猴(macacanemestrina);北方豚尾猴(northernpig-tailedmacaque),北方豚尾猕猴(macacaleonina);明打威猕猴(pagaiislandmacaque)或明打威猕猴(bokkoi),明打威猕猴(macacapagensis);西比路岛猕猴(siberutmacaque),西比路岛猕猴(macacasiberu);摩尔猕猴(moormacaque),摩尔猕猴(macacamaura);穿靴猕猴(bootedmacaque),穿靴猕猴(macacaochreata);通金猕猴(tonkeanmacaque),通金猕猴(macacatonkeana);黑克猕猴(heck'smacaque),黑克猕猴(macacahecki);浅黑猕猴(gorontalomacaque),浅黑猕猴(macacanigriscens);黑冠猕猴(celebescrestedmacaque)或黑"猴"(black"ape"),黑冠猕猴(macacanigra);石蟹猕猴或食蟹猴(crab-eatingmacaque)或长尾猴(long-tailedmacaque)或猴(kera),马来猴(macacafascicularis);短尾猴(stump-tailedmacaque)或短尾猴(bearmacaque),短尾猴(macacaarctoides);恒河猕猴(rhesusmacaque),猕猴(macaque);台湾猕猴(formosanrockmacaque),台湾猕猴(macacacyclopis);日本猕猴(japanesemacaque),日本猕猴(macacafuscata);斯里兰卡猴(toquemacaque),斯里兰卡猴(macacasinica);绮帽猕猴(bonnetmacaque),冠毛猕猴(macacaradiata);巴巴利猕猴(barbarymacaque),巴巴利猕猴(macacasylvanmus);熊猴(assammacaque),熊猴(macacaassamensis);藏猕猴(tibetanmacaque)或藏猕猴(milne-edwards'macaque),藏猕猴(macacathibetana);阿鲁纳卡猕猴(arunachalmacaque)或阿鲁纳卡猕猴(munzala),阿鲁纳卡猕猴(macacamunzala))内;在白脸猴属(genuslophocebus)(白颊白眉猴(gray-cheekedmangabey),白颊白眉猴(lophocebusalbigena);lophocebusalbigenaalbigena;lophocebusalbigenaosmani;lophocebusalbigenajohnstoni;黑毛白眉猴(blackcrestedmangabey),黑毛白眉猴(lophocebusaterrimus);opdenbosch'smangabey,lophocebusopdenboschi;高地白睑猴(highlandmangabey),高地白睑猴(lophocebuskipunji))内;在狒狒属(genuspapio)(阿拉伯狒狒(hamadryasbaboon),阿拉伯狒狒(papiohamadryas);几内亚狒狒(guineababoon),几内亚狒狒(papiopapio);绿狒狒(olivebaboon),猎神狒狒(papioanubis);黄狒狒(yellowbaboon),草原狒狒(papiocynocephalus);南非大狒(chacmababoon),豚尾狒狒(papioursinus))内;在狮尾狒属(genustheropithecus)(狮尾狒(gelada),狮尾狒(theropithecusgelada))内;在白眉猴属(genuscercocebus)(乌黑白眉猴(sootymangabey),白顶白眉猴(cercocebusatys);cercocebusatysatys;cercocebusatyslunulatus;白领白眼睑猴(collaredmangabey),白领白眼睑猴(cercocebustorquatus);敏白眉猴(agilemangabey),敏白眉猴(cercocebusagilis);金腹白眉猴(golden-belliedmangabey),金腹白眉猴(cercocebuschrysogaster),塔纳河白眉猴(tanarivermangabey),冠毛白眉猴(cercocebusgaleritus);狲氏白睑猴(sanjemangabey),狲氏白睑猴(cercocebussanjei))内;或者在山魈属(genusmandrillus)(山魈(mandrill),山魈(mandrillussphinx);鬼狒(drill),鬼狒(mandrillusleucophaeus))内。
最优选的是马来猴(也被称为石蟹猕猴,因此,在实施例中被命名为“石蟹猴”)以及恒河猕猴(恒河猴,被命名为“罗猴”)。
在疣猴亚科内,有益的非黑猩猩灵长类动物可来自于非洲族群,在疣猴属(genuscolobus)(黑疣猴(blackcolobus),黑疣猴(colobussatanas);安哥拉疣猴(angolacolobus),安哥拉疣猴(colobusangolensis);黑白疣猴(kingcolobus),黑白疣猴(colobuspolykomos);花斑疣猴(ursinecolobus),花斑疣猴(colobusvellerosus);东黑白疣猴(mantledguereza),东黑白疣猴(colobusguereza))内;在红疣猴属(genuspiliocolobus)(西方红疣猴(westernredcolobus),西方红疣猴(piliocolobusbadius);piliocolobusbadiusbadius;piliocolobusbadiustemminckii;红色残余部分猿(piliocolobusbadiuswaldronae);东方红疣(pennant'scolobus),东方红疣(piliocolobuspennantii);piliocolobuspennantiipennantii;尼日尔三角洲红色疣猴(piliocolobuspennantiiepieni);piliocolobuspennantiibouvieri;普氏红疣猴(preuss'sredcolobus),普氏红疣猴(piliocolobuspreussi);thollon'sredcolobus,piliocolobustholloni;中非红疣猴(centralafricanredcolobus),中非红疣猴(piliocolobusfoai);piliocolobusfoaifoai;piliocolobusfoaiellioti;piliocolobusfoaioustaleti;piliocolobusfoaisemlikiensis;piliocolobusfoaiparmentierorum;乌干达红疣猴(ugandanredcolobus),乌干达红疣猴(piliocolobustephrosceles);uzyngwaredcolobus,piliocolobusgordonorum;桑给巴尔红疣猴(zanzibarredcolobus),桑给巴尔红疣猴(piliocolobuskirkii);塔那河红疣猴(tanariverredcolobus),塔那河红疣猴(piliocolobusrufomitratus))内;或在绿疣猴属(genusprocolobus)(绿疣猴(olivecolobus),绿疣猴(procolobusverus))内。
在疣猴亚科内,有益的非黑猩猩灵长类动物可或者来自叶猴(langur)(叶猴(leafmonkey))族群,在长尾叶猴属(genussemnopithecus)内(尼泊尔长尾叶猴(nepalgraylangur),喜山长尾叶猴(semnopithecusschistaceus);喀什米尔长尾叶猴(kashmirgraylangur),喀什米尔长尾叶猴(semnopithecusajax);达顿灰叶猴(taraigraylangur),达顿灰叶猴(semnopithecushector),哈努曼叶猴(northernplainsgraylangur),哈努曼叶猴(semnopithecusentellus);黑足长尾叶猴(black-footedgraylangur),黑足长尾叶猴(semnopithecushypoleucos);南平原灰叶猴(southernplainsgraylangur),南平原灰叶猴(semnopithecusdussumieri);(tuftedgraylangur),毛冠灰叶猴(semnopithecuspriam))内;在紫脸叶猴(t.vetulus)族群或乌叶猴属(genustrachypithecus)(紫脸叶猴(purple-facedlangur),紫脸叶猴(trachypithecusvetulus);尼尔吉里猴(nilgirilangur),黑乌叶猴(trachypithecusjohnii))内;在乌叶猴属(genustrachypithecus)的银色乌叶猴(t.cristatus)族群(爪哇叶猴(javanlutung),乌叶猴(trachypithecusauratus);银叶猴(silveryleafmonkey)或银叶猴(silverylutung),银色乌叶猴(trachypithecuscristatus);indochineselutung,trachypithecusgermaini;丹那沙林乌叶猴(tenasserimlutung),丹那沙林乌叶猴(trachypithecusbarbei))内;在乌叶猴属的郁乌叶猴(t.obscures)族群(眼镜食叶猴(duskyleafmonkey)或眼镜食叶猴(spectacledleafmonkey),郁乌叶猴(trachypithecusobscurus);菲氏叶猴(phayre'sleafmonkey),菲氏叶猴(trachypithecusphayrei))内;在乌叶猴属的戴帽叶猴(t.pileatus)族群(戴帽叶猴(cappedlangur),戴帽叶猴(trachypithecuspileatus);邵力殊叶猴(shortridge'slangur),邵力殊叶猴(trachypithecusshortridgei);金叶猴(gee'sgoldenlangur),金叶猴(trachypithecusgeei))内;在乌叶猴属的黑叶猴(t.francoisi)族群(黑叶猴(francois'langur),黑叶猴(trachypithecusfrancoisi);海亭叶猴(hatinhlangur),海亭叶猴(trachypithecushatinhensis);白头叶猴(white-headedlangur),白头叶猴(trachypithecuspoliocephalus);老挝叶猴(laotianlangur),老挝叶猴(trachypithecuslaotum);德氏黑叶猴(delacour'slangur),德氏黑叶猴(trachypithecusdelacouri);印支黑乌叶猴(indochineseblacklangur),印支黑乌叶猴(trachypithecusebenus))内;或者在莱猴属(genuspresbytis)(苏门答腊叶猴(sumatransurili),黑脊叶猴(presbytismelalophos);印尼叶猴(bandedsurili),印尼叶猴(presbytisfemoralis);沙劳越叶猴(sarawaksurili),沙劳越叶猴(presbytischrysomelas);白腿叶猴(white-thighedsurili),白腿叶猴(presbytissiamensis);白额叶猴(white-frontedsurili),白额叶猴(presbytisfrontata);爪哇叶猴(javansurili),爪哇叶猴(presbytiscomata);托马斯叶猴(thomas'slangur),托马斯叶猴(presbytisthomasi);何氏叶猴(hose'slangur),何氏叶猴(presbytishosei);红叶猴(maroonleafmonkey),红叶猴(presbytisrubicunda);孟他维岛长尾叶猴(mentawailangur)或白颊叶猴(joja),白颊叶猴(presbytispotenziani);纳土纳岛叶猴(natunaislandsurili),纳土纳岛叶猴(presbytisnatunae))内。
在疣猴亚科内,有益的非黑猩猩灵长类动物可或者来自仰鼻族群(odd-nosedgroup),在白臀叶猴属(genuspygathrix)(白臀叶猴(red-shankeddouc),白臀叶猴(pygathrixnemaeus);黑腿白臀叶猴(black-shankeddouc),黑腿白臀叶猴(pygathrixnigripes);灰腿白臀叶猴(gray-shankeddouc),灰腿白臀叶猴(pygathrixcinerea))内;在仰鼻猴属(genusrhinopithecus)(川金丝猴(goldensnub-nosedmonkey),川金丝猴(rhinopithecusroxellana);滇金丝猴(blacksnub-nosedmonkey),滇金丝猴(rhinopithecusbieti);黔金丝猴(graysnub-nosedmonkey),黔金丝猴(rhinopithecusbrelichi);越南金丝猴(tonkinsnub-nosedlangur),越南金丝猴(rhinopithecusavunculus))内;在长鼻猴属(genusnasalis)(象鼻猴(proboscismonkey),长鼻猴(nasalislarvatus))内;或者在豚尾叶猴属(genussimias)(豚尾叶猴(pig-tailedlangur),豚尾叶猴(simiasconcolor))内。
如此处所用的,术语“狨猴(marmoset)”代表任何狨属(genuscallithrix)的新世界猴,例如属于狨亚属(subgenuscallithrix)(原文如此!)(普通狨(commonmarmoset),普通狨(callithrix(callithrix)jacchus);黑耳狨猴(black-tuftedmarmoset),丛尾狨(callithrix(callithrix)penicillata);维氏狨(wied'smarmoset),维氏狨(callithrix(callithrix)kuhlii);白头狨(white-headedmarmoset),白头狨(callithrix(callithrix)geoffroyi);黄冠狨(buffy-headedmarmoset),黄冠狨(callithrix(callithrix)flaviceps),白耳狨(buffy-tuftedmarmoset),白耳狨(callithrix(callithrix)aurita))的大西洋狨猴(atlanticmarmosets);属于mico亚属(里约狨猴(rioacarimarmoset),里约狨猴(callithrix(mico)acariensis);manicoremarmoset,callithrix(mico)manicorensis;银毛猴(silverymarmoset),银毛猴(callithrix(mico)argentata);白狨(whitemarmoset),白狨(callithrix(mico)leucippe);艾氏狨(emilia'smarmoset),艾氏狨(callithrix(mico)emiliae);黑头狨(black-headedmarmoset),黑头狨(callithrix(mico)nigriceps);马氏狨(marca'smarmoset),马氏狨(callithrix(mico)marcai);黑尾狨(black-tailedmarmoset),黑尾狨(callithrix(mico)melanura);白肩狨(santaremmarmoset),白肩狨(callithrix(mico)humeralifera);mauésmarmoset,callithrix(mico)mauesi;黄肢狨(gold-and-whitemarmoset),黄肢狨(callithrix(mico)chrysoleuca);贺氏狨(hershkovitz'smarmoset),贺氏狨(callithrix(mico)intermedia);satérémarmoset,callithrix(mico)saterei)的亚马逊河狨猴(amazonianmarmosets);属于卢氏亚属(subgenuscallibella)(卢氏倭狨(callithrix(callibella))humilis))的roosmalens'dwarfmarmoset;或者属于倭狨亚属(subgenuscebuella)(倭狨(callithrix(cebuella)pygmaea)的侏儒狨猴(pygmymarmoset)。
新世界猴的其它属包括獠狨属(包括绒顶柽柳猴-族群、绢毛猴族群、黑须柽柳猴族群、长须柽柳猴族群、斑绢猴族群以及烙印柽柳猴族群)的獠狨(tamarins)以及samiri属(即,松鼠猴、红背松鼠猴、马河松鼠、亚马逊松鼠猴、黑松鼠猴)的松鼠猴。
在本发明的双特异性单链抗体分子的一个优选实施方案中,该非黑猩猩灵长类动物是旧世界猴。在该多肽的一个更优选实施方案中,该旧世界猴是狒狒属猕猴属的猴子。最优选地,猕猴属的猴子是熊猴(assamesemacaque)(熊猴(macacaassamensis))、巴巴利猕猴(barbarymacaque)(巴巴利猕猴(macacasylvanus))、绮帽猕猴(bonnetmacaque)(冠毛猕猴(macacaradiata))、穿靴(booted)或苏拉威西穿靴猕猴(sulawesi-bootedmacaque)(穿靴猕猴(macacaochreata))、苏拉威西冠猴(sulawesicrestedmacaque)(黑冠猕猴(macacanigra))、台湾猕猴(formosanrockmacaque)(台湾猕猴(macacacyclopis))、日本雪猕猴(japanesesnowmacaque)或日本猕猴(japanesemacaque)(日本猕猴(macacafuscata))、食蟹猴(cynomologusmonkey)或食蟹猴(crab-eatingmacaque)或长尾猴(long-tailedmacaque)或爪哇猕猴(javamacaque)(马来猴(macacafascicularis))、狮尾猴(lion-tailedmacaque)(狮尾猴(macacasilenus))、豚尾猕猴(pigtailedmacaque)(豚尾猕猴(macacanemestrina))、恒河猕猴(rhesusmacaque)(恒河猕猴(macacamulatta))、藏猕猴(tibetanmacaque)(藏猕猴(macacathibetana))、通金猕猴(tonkeanmacaque)(通金猕猴(macacatonkeana))、斯里兰卡猴(toquemacaque)(斯里兰卡猴(macacasinica))、短尾猴(stump-tailedmacaque)或胭脂猴(red-facedmacaque)或短尾猴(bearmacaque)(短尾猴(macacaarctoides))或者摩尔猕猴(moormacaque)(摩尔猕猴(macacamaurus))。最优选地,狒狒属的猴子是阿拉伯狒狒(hamadryasbaboon),阿拉伯狒狒(papiohamadryas);几内亚狒狒(guineababoon),几内亚狒狒(papiopapio);绿狒狒(olivebaboon),猎神狒狒(papioanubis);黄狒狒(yellowbaboon),草原狒狒(papiocynocephalus);南非大狒(chacmababoon),豚尾狒狒(papioursinus)。
在本发明的双特异性单链抗体分子的一个可选择的优选的实施方案中,该非黑猩猩灵长类动物是新世界猴。在该多肽的一个更优选地实施例中,该新世界猴是狨属(狨猿)、獠狨属或samiri属的猴子。最优选地,狨属的猴子是白鬓狨,獠狨属的猴子是绒顶柽柳猴以及samiri属的猴子是松鼠猴。
如本文所用的,术语“细胞表面抗原”是指展示在细胞表面上的分子。在大多数情况下,这个分子将位于细胞的原生质膜中或其上,使得这个分子的至少一部分以三级结构的形式保持从细胞外可接触。位于原生质膜中的细胞表面分子的非限制性的实例是其三级构象包括亲水性和疏水性区域的跨膜蛋白。这里,至少一个疏水性区域容许该细胞表面分子嵌入或者插入细胞的疏水性原生质膜中,而亲水性区域在原生质膜的任一侧分别延伸至细胞质和细胞外隙中。位于原生质膜上的细胞表面分子的非限制性实例是以下蛋白:所述蛋白在半胱氨酸残基处被修饰以带有棕榈酰基、在c-端半胱氨酸残基处被修饰以带有法尼基或者在c-端被修饰以带有糖基磷脂酰肌醇(“gpi”)锚。这些基团容许蛋白质共价连接至原生质膜的外表面,在此处它们保持可以被外细胞分子(例如抗体)识别。细胞表面抗原的实例是cd3ε和psma。如上述的,psma是细胞表面抗原,是用于治疗癌症(包括但不限于实体瘤,优选癌和前列腺癌)的靶标。
鉴于此,psma也可被表征为肿瘤抗原。如本文所用的,术语“肿瘤抗原”可理解为那些存在于肿瘤细胞上的抗原。这些抗原以胞外部分呈现在细胞表面上,所述胞外部分常与所述分子的跨膜和胞质部分相组合。这些抗原有时候仅由肿瘤细胞呈现,而从未由正常细胞呈现。肿瘤抗原可专门表达在肿瘤细胞上,或者相比于正常细胞,肿瘤抗原可呈现肿瘤特异性突变。在这种情况下,它们被称为肿瘤-特异性抗原。更常见的是由肿瘤细胞以及正常细胞呈现的抗原,它们被称为肿瘤相关抗原。相比于正常细胞,这些肿瘤相关抗原可被过度表达,或者由于肿瘤组织与正常组织相比结构较不紧密,这些肿瘤相关抗原在肿瘤细胞中可以进行抗体结合。一个符合本发明的肿瘤抗原的实例是psma。
如上面描述的,本发明的双特异性单链抗体分子以所述第一结合域结合至人类以及非黑猩猩灵长类动物cd3ε(epsilin)链的表位,其中,该表位是包含在如seqidno.2、seqidno.4、seqidno.6或seqidno.8中所示的27个氨基酸残基或者其功能性片段中的氨基酸序列的一部分。
按照本发明,对于本发明的双特异性单链抗体分子优选的是:所述表位是包含26、25、24、23、22、21、20、19、18、17、16、15、14、13、12、11、10、9、8、7、6或5个氨基酸的氨基酸序列的一部分。
更优选地,其中,所述表位至少包含氨基酸序列gln-asp-gly-asn-glu(qdgne)。
在本发明中,该n-端1-27个氨基酸残基的功能性片段是指,当从其在cd3复合物的天然环境中取出(并且被融合至异源氨基酸序列,例如epcam或免疫球蛋白fc部分,如在wo2008/119567的实施例3.1中公开的)时,所述功能性片段仍然是维持其3维结构完整性的环境独立的表位。在cd3ε的27个氨基酸n-端多肽或其功能性片段内的3维结构的维持可被用于产生结合域,所述结合域以相同结合亲和力在体外结合至n-端cd3ε多肽片段,在体内结合至t细胞上的天然cd3复合物(的cd3ε亚基)。在本发明中,该n-端1-27个氨基酸残基的功能性片段是指本文提供的cd3结合域仍然可以以环境独立的方式结合至此类功能性片段。本领域技术人员知道用于表位定位的方法,从而确定表位的哪些氨基酸残基可被此类抗-cd3的结合域识别(例如,丙氨酸扫描;参见wo2008/119567的实施例)。
在本发明的一个实施方案中,本发明的双特异性单链抗体分子包括能够结合至人类和非黑猩猩灵长类动物cd3ε链的表位的(第一)结合域和能够结合至细胞表面抗原psma的第二结合域。
在本发明中,进一步优选地:所述第二结合域结合至人类细胞表面抗原psma和/或非黑猩猩灵长类动物psma。特别优选地,所述第二结合域结合至人类psma以及非黑猩猩灵长类动物psma,优选猕猴psma。要理解的是:所述第二结合域结合至至少一种非黑猩猩灵长类动物psma,然而,它也可结合至2、3或更多种非黑猩猩灵长类动物psma同源物。例如,所述第二结合域可结合至石蟹猕猴psma和恒河猴psma。
包括本文描述的所有方法、用途、试剂盒等的本发明也与所述第二结合域本身(即,非上下文中的双特异性单链抗体)相关。“本身”进一步包括非本文描述的双特异性单链抗体的抗体形式,例如抗体片段(包含该第二结构域)、人源化抗体、包含该第二结构域的融合蛋白等。本发明的双特异性单链抗体以外的抗体形式也如上所述。
为了产生本发明的双特异性单链抗体分子的第二结合域(例如,如本文定义的双特异性单链抗体),还可以利用结合至相应的人类和/或非黑猩猩灵长类动物的细胞表面抗原(如psma)的单克隆抗体。针对如本文定义的双特异性多肽的适当的结合域例如可通过本领域描述的重组方法来自跨物种特异性的单克隆抗体。结合至人类细胞表面抗原以及黑猩猩灵长类动物的所述细胞表面抗原的同源物的单克隆抗体,可以通过如上所述的facs分析进行测定。对于本领域技术人员显而易见的是:跨物种特异性抗体也可由文献(milstein和
对于本发明的双特异性单链抗体分子特别优选的是:能够结合至人类和非黑猩猩灵长类动物cd3ε链的表位的第一结合域包含vl区域,所述vl区域含有选自下列的cdr-l1、cdr-l2以及cdr-l3:
(a)如seqidno.27中所示的cdr-l1、如seqidno.28中所示的cdr-l2以及如seqidno.29中所示的cdr-l3;
(b)如seqidno.117中所示的cdr-l1、如seqidno.118中所示的cdr-l2以及如seqidno.119中所示的cdr-l3;以及
(c)如seqidno.153中所示的cdr-l1、如seqidno.154中所示的cdr-l2以及如seqidno.155中所示的cdr-l3。
可变区域,即可变轻链(“l”或“vl”)以及可变重链(“h”或“vh”)在本领域中理解为提供抗体的结合域。这个可变区域含有互补决定区。
术语“互补决定区”(cdr)是本领域熟知的,用于指示抗体的抗原特异性。术语“cdr-l”或“lcdr”或“lcdr”是指在vl中的cdr,而术语“cdr-h”或“hcdr”或hcdr”是指在vh中的cdr。
在本发明的双特异性单链抗体分子的一个可选择的优选实施方案中,该能够结合至人类以及非黑猩猩灵长类动物cd3ε链的表位的第一结合域包含vh区域,所述vh区域含有选自下列的cdr-h1、cdr-h2以及cdr-h3:
(a)如seqidno.12中所示的cdr-h1、如seqidno.13中所示的cdr-h2以及如seqidno.14中所示的cdr-h3;
(b)如seqidno.30中所示的cdr-h1、如seqidno.31中所示的cdr-h2以及如seqidno.32中所示的cdr-h3;
(c)如seqidno.48中所示的cdr-h1、如seqidno.49中所示的cdr-h2以及如seqidno.50中所示的cdr-h3;
(d)如seqidno.66中所示的cdr-h1、如seqidno.67中所示的cdr-h2以及如seqidno.68中所示的cdr-h3;
(e)如seqidno.84中所示的cdr-h1、如seqidno.85中所示的cdr-h2以及如seqidno.86中所示的cdr-h3;
(f)如seqidno.102中所示的cdr-h1、如seqidno.103中所示的cdr-h2以及如seqidno.104中所示的cdr-h3;
(g)如seqidno.120中所示的cdr-h1、如seqidno.121中所示的cdr-h2以及如seqidno.122中所示的cdr-h3;
(h)如seqidno.138中所示的cdr-h1、如seqidno.139中所示的cdr-h2以及如seqidno.140中所示的cdr-h3;
(i)如seqidno.156中所示的cdr-h1、如seqidno.157中所示的cdr-h2以及如seqidno.158中所示的cdr-h3;以及
(j)如seqidno.174中所示的cdr-h1、如seqidno.175中所示的cdr-h2以及如seqidno.176中所示的cdr-h3。
进一步优选地,该能够结合至人类和非黑猩猩灵长类动物cd3ε链的表位的结合域包含vl区域,所述vl区域选自如seqidno.35、seqidno.39、seqidno.125、seqidno.129、seqidno.161或seqidno.165中所示的vl区域。
可选择地优选地:该能够结合至人类和非黑猩猩灵长类动物cd3ε链表位的第一结合域包含vh区域,所述vh区域选自如seqidno.15、seqidno.19、seqidno.33、seqidno.37、seqidno.51、seqidno.55、seqidno.69、seqidno.73、seqidno.87、seqidno.91、seqidno.105、seqidno.109、seqidno.123、seqidno.127、seqidno.141、seqidno.145、seqidno.159、seqidno.163、seqidno.177或seqidno.181中所示的vh区域。
更优选地,本发明的双特异性单链抗体分子的特征在于该能够结合至人类和非黑猩猩灵长类动物cd3ε链的表位的第一结合域,其包含选自下列的vl区域和vh区域:
(a)如seqidno.17或seqidno.21中所示的vl区域以及如seqidno.15或seqidno.19中所示的vh区域;
(b)如seqidno.35或seqidno.39中所示的vl区域以及如seqidno.33或seqidno.37中所示的vh区域;
(c)如seqidno.53或seqidno.57中所示的vl区域以及如seqidno.51或seqidno.55中所示的vh区域;
(d)如seqidno.71或seqidno.75中所示的vl区域以及如seqidno.69或seqidno.73中所示的vh区域;
(e)如seqidno.89或seqidno.93中所示的vl区域以及如seqidno.87或seqidno.91中所示的vh区域;
(f)如seqidno.107或seqidno.111中所示的vl区域以及如seqidno.105或seqidno.109中所示的vh区域;
(g)如seqidno.125或seqidno.129中所示的vl区域以及如seqidno.123或seqidno.127中所示的vh区域;
(h)如seqidno.143或seqidno.147中所示的vl区域以及如seqidno.141或seqidno.145中所示的vh区域;
(i)如seqidno.161或seqidno.165中所示的vl区域以及如seqidno.159或seqidno.163中所示的vh区域;以及
(j)如seqidno.179或seqidno.183中所示的vl区域以及如seqidno.177或seqidno.181中所示的vh区域。
根据本发明的双特异性单链抗体分子的一个优选实施方案,在该结合至cd3ε的第一结合域中成对的vh-区域和vl-区域呈单链抗体(scfv)的形式。该vh和vl区域以vh-vl或vl-vh的次序排列。优选地,该vh-区域的n-端被定位到一个连接序列。该vl-区域的c-端被定位到连接序列。换句话说,在本发明的双特异性单链抗体分子的cd3结合域中的结构域排列优选地是vh-vl,其中所述cd3结合域的c-端位于第二(细胞表面抗原,如psma)结合域。优选地,该vh-vl是或包含seqidno.185。
本发明的如上所述的双特异性单链抗体分子的一个优选实施方案的特征在于:该能够结合至人类和非黑猩猩灵长类动物cd3ε链的表位的第一结合域包含选自seqidno.23、seqidno.25、seqidno.41、seqidno.43、seqidno.59、seqidno.61、seqidno.77、seqidno.79、seqidno.95、seqidno.97、seqidno.113、seqidno.115、seqidno.131、seqidno.133、seqidno.149、seqidno.151、seqidno.167、seqidno.169、seqidno.185或seqidno.187的氨基酸序列。
本发明进一步涉及上述的双特异性单链抗体,其中所述第二结合域结合至该细胞表面抗原psma。
根据本发明的一个优选实施方案,上述表征的双特异性单链抗体分子包含下列序列组作为在该第二结合域中的cdrh1、cdrh2、cdrh3、cdrl1、cdrl2以及cdrl3,所述序列组选自:
a)seqidno:226-228的cdrh1-3以及seqidno:231-233的cdrl1-3;
b)seqidno:240-242的cdrh1-3以及seqidno:245-247的cdrl1-3;
c)seqidno:254-256的cdrh1-3以及seqidno:259-261的cdrl1-3;
d)seqidno:268-270的cdrh1-3以及seqidno:273-275的cdrl1-3;
e)seqidno:618-620的cdrh1-3以及seqidno:623-625的cdrl1-3;
f)seqidno:282-284的cdrh1-3以及seqidno:287-289的cdrl1-3;
g)seqidno:296-298的cdrh1-3以及seqidno:301-303的cdrl1-3;
h)seqidno:310-312的cdrh1-3以及seqidno:315-317的cdrl1-3;
i)seqidno:324-326的cdrh1-3以及seqidno:329-331的cdrl1-3;
j)seqidno:338-340的cdrh1-3以及seqidno:343-345的cdrl1-3;
k)seqidno:352-354的cdrh1-3以及seqidno:357-359的cdrl1-3;
l)seqidno:366-368的cdrh1-3以及seqidno:371-373的cdrl1-3;
m)seqidno:380-382的cdrh1-3以及seqidno:385-387的cdrl1-3;
n)seqidno:394-396的cdrh1-3以及seqidno:399-401的cdrl1-3;
o)seqidno:408-410的cdrh1-3以及seqidno:413-415的cdrl1-3;
p)seqidno:422-424的cdrh1-3以及seqidno:427-429的cdrl1-3;
q)seqidno:436-438的cdrh1-3以及seqidno:441-443的cdrl1-3;
r)seqidno:450-452的cdrh1-3以及seqidno:455-457的cdrl1-3;
s)seqidno:464-466的cdrh1-3以及seqidno:469-471的cdrl1-3;
t)seqidno:478-480的cdrh1-3以及seqidno:483-485的cdrl1-3;
u)seqidno:492-494的cdrh1-3以及seqidno:497-499的cdrl1-3;
v)seqidno:506-508的cdrh1-3以及seqidno:511-513的cdrl1-3;
w)seqidno:520-522的cdrh1-3以及seqidno:525-527的cdrl1-3;
x)seqidno:534-536的cdrh1-3以及seqidno:539-541的cdrl1-3;
y)seqidno:548-550的cdrh1-3以及seqidno:553-555的cdrl1-3;
z)seqidno:562-564的cdrh1-3以及seqidno:567-569的cdrl1-3;
aa)seqidno:576-578的cdrh1-3以及seqidno:581-583的cdrl1-3;
ab)seqidno:590-592的cdrh1-3以及seqidno:595-597的cdrl1-3;以及,
ac)seqidno:604-606的cdrh1-3以及seqidno:609-611的cdrl1-3。
双特异性单链抗体分子的优选组包括下列序列的组作为在该第二结合域中的cdrh1、cdrh2、cdrh3、cdrl1、cdrl2以及cdrl3,其选自:
a)seqidno:226-228的cdrh1-3以及seqidno:231-233的cdrl1-3;
b)seqidno:240-242的cdrh1-3以及seqidno:245-247的cdrl1-3;
c)seqidno:254-256的cdrh1-3以及seqidno:259-261的cdrl1-3;
d)seqidno:268-270的cdrh1-3以及seqidno:273-275的cdrl1-3;以及,
e)seqidno:618-620的cdrh1-3以及seqidno:623-625的cdrl1-3。
这些分子包含结合至该细胞表面抗原psma的第二结合域,所述第二结合域由源自亲本psma特异性结合分子的vh-链和源自对不同抗原(即,也被称为cd326的上皮细胞黏附分子(epcam))有特异性的结合分子的vl-链组成。令人惊讶地发现:具有此源自亲本psma特异性结合分子的vh-链和源自亲本epcam特异性结合分子的vl-链的特定组合的结合分子专门结合至psma而不是epcam。这个被包含在本发明的双特异性单链抗体分子中的psma特异性结合域组的结合特异性在所附的实施例3中描述。
双特异性单链抗体分子的另一优选组包括下列序列的组作为在该第二结合域中的cdrh1、cdrh2、cdrh3、cdrl1、cdrl2以及cdrl3,其选自:
a)seqidno:282-284的cdrh1-3以及seqidno:287-289的cdrl1-3;
b)seqidno:296-298的cdrh1-3以及seqidno:301-303的cdrl1-3;
c)seqidno:310-312的cdrh1-3以及seqidno:315-317的cdrl1-3;
d)seqidno:324-326的cdrh1-3以及seqidno:329-331的cdrl1-3;
e)seqidno:338-341的cdrh1-3以及seqidno:343-345的cdrl1-3;
f)seqidno:352-354的cdrh1-3以及seqidno:357-359的cdrl1-3;以及,
g)seqidno:366-368的cdrh1-3以及seqidno:371-373的cdrl1-3。
双特异性单链抗体分子的另一优选组包括下列序列的组作为在该第二结合域中的cdrh1、cdrh2、cdrh3、cdrl1、cdrl2以及cdrl3,其选自:
a)seqidno:380-382的cdrh1-3以及seqidno:385-387的cdrl1-3;
b)seqidno:394-396的cdrh1-3以及seqidno:399-401的cdrl1-3;
c)seqidno:408-410的cdrh1-3以及seqidno:413-415的cdrl1-3;
d)seqidno:422-424的cdrh1-3以及seqidno:427-429的cdrl1-3;
e)seqidno:436-438的cdrh1-3以及seqidno:441-443的cdrl1-3;
f)seqidno:450-452的cdrh1-3以及seqidno:455-457的cdrl1-3;
g)seqidno:464-466的cdrh1-3以及seqidno:469-471的cdrl1-3;
h)seqidno:478-480的cdrh1-3以及seqidno:483-485的cdrl1-3;
i)seqidno:492-494的cdrh1-3以及seqidno:497-499的cdrl1-3;
j)seqidno:506-508的cdrh1-3以及seqidno:511-513的cdrl1-3;
k)seqidno:520-522的cdrh1-3以及seqidno:525-527的cdrl1-3;
l)seqidno:534-536的cdrh1-3以及seqidno:539-541的cdrl1-3;
m)seqidno:548-550的cdrh1-3以及seqidno:553-555的cdrl1-3;
n)seqidno:562-564的cdrh1-3以及seqidno:567-569的cdrl1-3;
o)seqidno:576-578的cdrh1-3以及seqidno:581-583的cdrl1-3;
p)seqidno:590-592的cdrh1-3以及seqidno:595-597的cdrl1-3;以及
q)seqidno:604-606的cdrh1-3以及seqidno:609-611的cdrl1-3。
对于本发明的双特异性单链抗体分子优选的是:所述结合至该细胞表面抗原psma的第二结合域包括下列序列的组作为在该第二结合域中的vh-和vl-链,其选自:
a)seqidno:225的vh-链以及seqidno:230的vl-链;
b)seqidno:239的vh-链以及seqidno:244的vl-链;
c)seqidno:253的vh-链以及seqidno:258的vl-链;
d)seqidno:267的vh-链以及seqidno:272的vl-链;
e)seqidno:617的vh-链以及seqidno:622的vl-链;
f)seqidno:281的vh-链以及seqidno:286的vl-链;
g)seqidno:295的vh-链以及seqidno:300的vl-链;
h)seqidno:309的vh-链以及seqidno:314的vl-链;
i)seqidno:323的vh-链以及seqidno:328的vl-链;
j)seqidno:337的vh-链以及seqidno:342的vl-链;
k)seqidno:351的vh-链以及seqidno:356的vl-链;
l)seqidno:365的vh-链以及seqidno:370的vl-链;
m)seqidno:379的vh-链以及seqidno:384的vl-链;
n)seqidno:393的vh-链以及seqidno:398的vl-链;
o)seqidno:407的vh-链以及seqidno:412的vl-链;
p)seqidno:421的vh-链以及seqidno:426的vl-链;
q)seqidno:435的vh-链以及seqidno:440的vl-链;
r)seqidno:449的vh-链以及seqidno:454的vl-链;
s)seqidno:463的vh-链以及seqidno:468的vl-链;
t)seqidno:477的vh-链以及seqidno:482的vl-链;
u)seqidno:491的vh-链以及seqidno:496的vl-链;
v)seqidno:505的vh-链以及seqidno:510的vl-链;
w)seqidno:519的vh-链以及seqidno:524的vl-链;
x)seqidno:533的vh-链以及seqidno:538的vl-链;
y)seqidno:547的vh-链以及seqidno:552的vl-链;
z)seqidno:561的vh-链以及seqidno:566的vl-链;
aa)seqidno:575的vh-链以及seqidno:580的vl-链;
ab)seqidno:589的vh-链以及seqidno:594的vl-链;以及
ac)seqidno:603的vh-链以及seqidno:608的vl-链。
上述vh-和vl-链也分别示于seqidno:235、249、263、277、627、291、305、319、333、347、361、375、389、403、417、431、445、459、473、487、501、515、529、543、557、571、585、599以及613。
本发明的双特异性单链抗体分子的第二结合域对应的vl-和vh-区域以及相应的scfv的序列(氨基酸序列和核苷酸序列)示于序列表中。
在本发明的双特异性单链抗体分子中,该结合域以vl-vh-vh-vl、vl-vh-vl-vh、vh-vl-vh-vl或vh-vl-vl-vh的次序排列,如所附的实施例中所示。优选地,该结合域以vhpsma-vlpsma-vhcd3-vlcd3或vlpsma-vhpsma-vhcd3-vlcd3的次序排列。
本发明的一个特别优选的实施方案涉及上述表征的多肽,其中所述双特异性单链抗体分子包括选自下列的序列:
(a)如seqidno:237、251、265、279、629、293、307、321、335、349、363、377、391、405、419、433、447、461、475、489、503、517、531、545、559、573、587、601或615中的任一个中所示的氨基酸序列;
(b)由如seqidno:238、252、266、280、630、294、308、322、336、350、364、378、392、406、420、434、448、462、476、490、504、518、532、546、560、574、588、602或616中的任一个中所示的核酸序列编码的氨基酸序列。
本发明涉及的双特异性单链抗体分子包含如seqidno:237、251、265、279、629、293、307、321、335、349、363、377、391、405、419、433、447、461、475、489、503、517、531、545、559、573、587、601或615中的任一个中所示的氨基酸序列,以及与seqidno:237、251、265、279、629、293、307、321、335、349、363、377、391、405、419、433、447、461、475、489、503、517、531、545、559、573、587、601或615的氨基酸序列有至少96%同一性、优选97%、更优选至少98%同一性、最优选至少99%同一性的氨基酸序列。本发明还涉及如seqidno:238、252、266、280、630、294、308、322、336、350、364、378、392、406、420、434、448、462、476、490、504、518、532、546、560、574、588、602或616中的任一个所示的相对应的核酸序列,以及与示于seqidno:238、252、266、280、630、294、308、322、336、350、364、378、392、406、420、434、448、462、476、490、504、518、532、546、560、574、588、602或616的核酸序列有至少96%同一性、优选97%、更优选至少98%同一性、最优选至少99%同一性的核酸序列。应当理解的是:在整个核苷酸或氨基酸序列上确定序列同一性。关于序列比对,例如,可以使用程序gap或bestfit(needleman和wunschj.mol.biol.48(1970),443-453;smith和waterman,adv.appl.math2(1981),482-489),它被包含在gcg软体包(geneticscomputergroup,575sciencedrive,madison,wisconsin,usa53711(1991))中。对本领域的技术人员来说这是一个常规的方法,即通过使用例如上述程序之一来测定和鉴定与本发明的双特异性单链抗体的核苷酸或氨基酸序列具有例如96%(97%、98%或99%)序列同一性的核苷酸或氨基酸序列。例如,根据克里克的摆动假说,反密码子上的5'碱基不像其他2个碱基有空间限制,从而可具有非标准的碱基配对。换句话说:密码子三联体中的第三位可变化,从而在此第三位有区别的2个三联体可编码相同的氨基酸残基。所述假说是本领域技术人员熟知的(参见例如http://en.wikipedia.org/wiki/wobble_hypothesis;crick,jmolbiol19(1966):548–55)。
在本发明的psmaxcd3双特异性单链抗体构建体中优选的结构域排列在下面的实施例中示出。
在本发明的一个优选实施方案中,该双特异性单链抗体对cd3ε和由它们的第二结合域识别的人类与非黑猩猩灵长类动物细胞表面抗原psma是跨物种特异性的。
在一个备选实施方案中,本发明提供编码上述本发明的双特异性单链抗体分子的核酸序列。
本发明还涉及包含本发明的核酸分子的载体。
许多适合的载体是分子生物学领域的技术人员已知的,载体的选择取决于需要的功能,并且包括质粒、粘粒、病毒、噬菌体以及常规用于基因工程的其他载体。可以用本领域的技术人员熟知的方法构建各种质粒和载体;参见,例如,描述于sambrook等人(上述引用文)和ausubel,currentprotocolsinmolecularbiology,greenpublishingassociatesandwileyinterscience,ny(1989),(1994)的技术。可选择地,本发明的多核苷酸和载体可重构成脂质体以递送至靶细胞。如下面进一步详细讨论地,将克隆载体用于分离dna的各个序列。在需要表达特定多肽的情况下,相关序列可被转移至表达载体中。典型的克隆载体包括pbluescriptsk、pgem、puc9、pbr322以及pgbt9。典型的表达载体包括ptre、pcal-n-ek、pesp-1、pop13cat。
优选地,所述载体包含的核酸序列是可操作地连接至本文定义的上述核酸序列的调节序列。
术语“调控序列”是指dna序列,其对于实现它们连接的编码序列的表达是必要的。此类控制序列的性质随宿主生物体的不同而不同。在原核生物中,一般而言,控制序列包括启动子、核糖体结合位点和终止子。在真核生物中,一般而言,控制序列包括启动子、终止子以及在某些情况下包括增强子、反式作用因子或转录因子。术语“控制序列”意在在最低限度上包括其存在为表达必需的所有组份,并且还可包括另外的有益组份。
术语“可操作地连接”是指并列位置,其中描述的组份处于允许它们以它们所预期的方式作用的关系。“可操作地连接”至编码序列的控制序列以使得该编码序列的表达在与控制序列相容的条件下实现的方式而连接。在该控制序列是启动子的情况下,对本领域的技术人员显而易见的是优选使用双链核酸。
因此,优选地,所列举的载体是表达载体。“表达载体”是一种构建体,可被用于转化选择的宿主并且在该选择的宿主中提供编码序列的表达。表达载体可以是,例如克隆载体、双元载体或整合载体。表达包括优选地将核酸分子转录成可翻译的mrna。确保在原核和/或真核细胞中表达的调控元件是本领域技术人员所熟知的。在真核细胞的情况下,它们通常包括确保转录起始的启动子和可选的确保转录终止及转录物稳定的poly-a信号。可能容许在原核宿主细胞中表达的调控元件包括,例如,在大肠杆菌中的pl、lac、trp或tac启动子,而容许在真核宿主细胞中表达的调控元件的实例是酵母菌中的aox1或gal1启动子或者在哺乳动物或其他动物细胞中的cmv-、sv40、rsv-启动子(rous肉瘤病毒)、cmv-增强子、sv40-增强子或球蛋白内含子。
除了负责转录起始的元件以外,这些调控元件还可包括转录终止信号,如在多核苷酸下游的sv40-poly-a位点或tk-poly-a位点。此外,取决于使用的表达系统,可将能够把多肽导向至细胞隔室或将它分泌至介质内的前导序列添加至所述的核酸序列的编码序列中,并且所述前导序列是本领域熟知的;参见,例如wo2008/119567。前导序列在适合的阶段与翻译、启动和终止序列一起装配,并且优选地,前导序列能够指导翻译的蛋白质或其部分分泌至细胞周质间隙或细胞外介质内。任选地,异源序列可编码融合蛋白,该融合蛋白包括赋予所需特性(例如,被表达的重组产物的稳定性或纯化简化)的n-端识别肽;参见,如上述。在本文中,适合的表达载体是本领域已知的,例如okayama-bergcdna表达载体pcdv1(pharmacia)、pcdm8、prc/cmv、pcdna1、pcdna3(in-vitrogene)、pef-dhfr、pef-ada或pef-neo(mack等人pnas(1995)92,7021-7025以及raum等人cancerimmunolimmunother(2001)50(3),141-150)或psport1(gibcobrl)。
优选地,在能够转化或转染真核宿主细胞的载体中,所述表达控制序列是真核启动子系统,但也可以使用原核宿主的控制序列。一但该载体被导入到适当的宿主中,该宿主被维持在适合核苷酸序列高水平表达的条件下,并且如果需要,之后可进行本发明的双特异性单链抗体分子的收集和纯化;参见,例如,所附的实施例。
被用于表达细胞周期相互作用蛋白的另一种表达系统是昆虫系统。在这个系统中,苜蓿银纹夜蛾核型多角体病毒(acnpv)用作载体,以在草地贪夜蛾细胞中或在粉纹夜蛾幼虫中表达外源基因。上述的核酸分子的编码序列可被克隆至病毒的非必需区域,如,多角体蛋白基因,并且被放置在多角体蛋白启动子的控制之下。所述编码序列的成功插入将使多角体蛋白基因处于非活动状态并且产生缺乏外壳蛋白壳的重组病毒。然后用重组病毒感染草地贪夜蛾细胞或粉纹夜蛾幼虫,本发明的蛋白在其中表达(smith,j.virol.46(1983),584;engelhard,proc.nat.acad.sci.usa91(1994),3224-3227)。
附加的调控元件可包括转录以及翻译增强子。有利地,上述的本发明的载体包括可选择的和/或可评价(scorable)标记物。
可用于选择被转化细胞以及例如植物组织和植物的选择性标记物基因是本领域技术人员熟知的,所述选择性标记物基因包括,例如,以抗代谢物抗性作为选择基础的赋予对甲氨蝶呤抗性的dhfr(reiss,plantphysiol.(lifesci.adv.)13(1994),143-149);赋予对氨基糖苷类新霉素、卡那霉素以及巴龙霉素抗性的npt(herrera-estrella,emboj.2(1983),987-995)以及赋予对潮霉素抗性的hygro(marsh,gene32(1984),481-485)。还描述了另外的可选择的基因,即容许细胞利用吲哚代替色氨酸的trpb;容许细胞利用组氨醇代替组氨酸的hisd(hartman,proc.natl.acad.sci.usa85(1988),8047);容许细胞利用甘露糖(wo94/20627)的甘露糖-6-磷酸异构酶以及赋予对鸟氨酸脱羧酶抑制剂、2-(二氟甲基)-dl-鸟氨酸、dfmo抗性的odc(鸟氨酸脱羧酶)(mcconlogue,1987,in:currentcommunicationsinmolecularbiology,coldspringharborlaboratoryed.)或者赋予对杀稻瘟菌素s抗性的来自土曲霉的脱氨酶(tamura,biosci.biotechnol.biochem.59(1995),2336-2338)。
有用的可评价标记物也是本领域技术人员已知的并且有市售。有利地,所述标记物是编码萤光素酶(giacomin,pl.sci.116(1996),59-72;scikantha,j.bact.178(1996),121)、绿色萤光蛋白(gerdes,febslett.389(1996),44-47)或β-葡萄糖醛酸酶(jefferson,emboj.6(1987),3901-3907)的基因。这个实施方案对于简单以及快速筛选含有所述载体的细胞、组织以及生物体是特别有用的。
如上所述,为了达到例如纯化还有基因治疗的目的,所述核酸分子可被单独使用或作为载体的一部分使用,以在细胞中表达本发明的双特异性单链抗体分子。这些含有编码上述任一种本发明的双特异性单链抗体分子的dna序列的核酸分子或载体被导入细胞,继而所述细胞产生目标多肽。基于通过离体或体内技术将治疗基因导入细胞的基因治疗,是基因转移的最重要的应用之一。用于体外或体内基因治疗的适合的载体、方法或基因递送系统已描述于文献中并且是本领域技术人员已知的;参见,例如,giordano,naturemedicine2(1996),534-539;schaper,circ.res.79(1996),911-919;anderson,science256(1992),808-813;verma,nature389(1994),239;isner,lancet348(1996),370-374;muhlhauser,circ.res.77(1995),1077-1086;onodera,blood91(1998),30-36;verma,genether.5(1998),692-699;nabel,ann.nyacad.sci.811(1997),289-292;verzeletti,hum.genether.9(1998),2243-51;wang,naturemedicine2(1996),714-716;wo94/29469;wo97/00957,us5,580,859;us5,589,466;或者schaper,currentopinioninbiotechnology7(1996),635-640;dossantoscoura和nardivirolj.(2007),4:99。所述核酸分子和载体可被设计为直接导入或通过脂质体或病毒(例如,腺病毒、逆转录病毒)载体导入至细胞内。优选地,所述细胞是生殖细胞系细胞、胚胎细胞或卵细胞或者由其衍生的,最优选地,所述细胞是干细胞。胚胎干细胞的实例尤其可以是如描述于nagy,proc.natl.acad.sci.usa90(1993),8424-8428中的干细胞。
本发明还提供了用本发明的载体转化或转染的宿主。所述宿主可以通过将上述的本发明的载体或上述的本发明的核酸分子导入该宿主内产生。在宿主中至少一种载体或至少一种核酸分子的存在可调节编码上述的单链抗体构建体的基因的表达。
导入宿主内的本发明所述的核酸分子或载体可合并至该宿主的基因组内或它可被维持在染色体外。
宿主可以是任何原核或真核细胞。
术语“原核生物”是指包括所有可被dna或rna分子转化或转染以表达本发明的蛋白质的细菌。原核生物宿主可包括革兰氏阴性和革兰氏阳性细菌,例如大肠杆菌、鼠伤寒沙门氏菌、粘质沙雷氏菌以及枯草芽孢杆菌。术语“真核生物”是指包括酵母菌、高等植物、昆虫和优选的哺乳动物细胞。取决于在重组生产过程中采用的宿主,由本发明的多核苷酸所编码的蛋白质可被糖基化或非糖基化。特别优选的是使用含有本发明的双特异性单链抗体分子编码序列的质粒或病毒,并且该编码序列以遗传方式融合了n-端flag-标签和/或c-端his-标签。优选地,所述flag-标签的长度是大约4至8个氨基酸、最优选8个氨基酸。可以使用上述的多核苷酸,以利用本领域技术人员通常已知的技术来转化或转染宿主。再者,用于制备融合、可操作地连接的基因和例如在哺乳动物细胞和细菌中表达它们的方法是本领域技术人员熟知的(sambrook,上述引文)。
优选地,所述宿主是细菌或昆虫、真菌、植物或动物细胞。
特别设想的是:所述宿主可以是哺乳动物细胞。特别优选的宿主细胞包括cho细胞、cos细胞、骨髓瘤细胞株如sp2/0或ns/0。如在wo2008/119567的实施例中对于同类的其他分子所示例的,特别优选的是cho-细胞作为宿主。
更优选地,所述宿主细胞是人类细胞或人类细胞株,例如per.c6(kroos,biotechnol.prog.,2003,19:163-168)。
在其它实施方案中,本发明涉及一种用于生产本发明的双特异性单链抗体分子的方法,所述方法包括在允许本发明的双特异性单链抗体分子表达的条件下培养本发明的宿主并从培养物中回收产生的多肽。
被转化的宿主可在发酵罐中生长并且根据本领域已知的技术进行培养以达到最佳的细胞生长。然后可以从生长培养基、细胞裂解液、或细胞膜组分中分离本发明的双特异性单链抗体分子。例如经微生物表达的双特异性单链抗体分子的分离和纯化可以通过任何常规方法,例如,制备色谱分离和免疫学分离,正如那些涉及使用靶向例如本发明的双特异性单链抗体分子的标签的或如在所附实施例中描述的单克隆或多克隆抗体的方法。
允许表达的培养宿主的条件是本领域已知的,且取决于用于此方法的宿主系统和表达系统/载体。为了达到允许重组多肽表达的条件而被修饰的参数是本领域已知的。因此,适当的条件可由本领域技术人员无需进一步的创造性工作投入的情况下确定。
经表达后,本发明的双特异性单链抗体分子可根据本领域的标准操作程序被纯化,包括硫酸铵沉淀、亲和柱、柱层析、凝胶电泳以及类似操作;参见,scopes,"proteinpurification",springer-verlag,ny(1982)。对于药学用途,优选具有至少大约90至95%同质性的基本上纯的多肽,最优选98至99%或更高同质性。经纯化后(部分的或根据需要达到同质),本发明的双特异性单链抗体分子可被用于治疗(包括体外)或者开发和执行分析程序。此外,用于从培养物中回收本发明的双特异性单链抗体分子的方法详细描述于所附的wo2008/119567的关于同类的其他分子的实施例中。回收也可以通过分离能够结合至人类以及非黑猩猩灵长类动物cd3ε的表位的本发明的双特异性单链抗体分子的方法实现,该方法包括下列步骤:
(a)使该多肽与含有氨基酸序列gln-asp-gly-asn-glu-glu-met-gly(seqidno.211)或gln-asp-gly-asn-glu-glu-ile-gly(seqidno.212)的最多27个氨基酸的cd3ε的胞外域的n-端片段(经由它的c-端被固定至固相)接触;
(b)洗脱与所述片段结合的多肽;以及
(c)从(b)的洗脱液中分离该多肽。
优选地,本发明通过上述方法所分离的多肽具有人类来源。
这个方法或本发明的双特异性单链抗体分子的分离被理解为用于从多个候选多肽中分离一种或多种不同多肽(对其n-端包括氨基酸序列gln-asp-gly-asn-glu-glu-met-gly(seqidno.211)或gln-asp-gly-asn-glu-glu-ile-gly(seqidno.212)的cd3ε的胞外域片段具有相同的特异性)的方法以及用于从溶液中纯化多肽的方法。对于后一用于从溶液中纯化双特异性单链抗体分子的方法的非限制性实例是例如从培养悬浮液或来自此类培养物的制备物中纯化被重组表达的双特异性单链抗体分子。
如上所述的,用于这个方法的片段是灵长类动物cd3ε分子的胞外域的n-端片段。不同物种的cd3ε分子的胞外域的氨基酸序列如seqidno:1、3、5和7中所示。2种形式的n-端八聚体示于seqidno:211和212。优选地,这个n-端对于所述由本发明的方法识别的多肽的结合而言是自由获取的。术语“自由获取的”在本发明的上下文中被理解为无外加的动机,如his-标签。该his-标签与本文描述的结合分子的干扰在wo2008/119567中描述。
根据所述方法,所述片段经由它的c-端被固定至固相。本领域的技术人员依赖本发明的方法使用的实施方案将容易并且无创造性工作困扰地选择合适的固相支持物。固体支持物包括但不限于基质类珠粒(例如琼脂糖珠、琼脂糖凝胶珠、聚苯乙烯珠、葡聚糖珠)、板(培养板或多孔板)以及已知的芯片(例如来自
根据该方法,所述cd3ε的胞外域的片段可直接地或通过氨基酸的延伸物(可以是连接物或其他蛋白质/多肽部分)偶合至该固相支持物。可选择地,cd3ε的胞外域可通过一个或多个接头分子被间接耦合。
用于洗脱结合至被固定化的表位的肽或多肽的手段和方法是本领域公知的。同样适用于从洗脱液中分离被识别的多肽(们)的方法。
用于从多个候选多肽中分离一种或多种不同双特异性单链抗体分子(对于在其n-端包括氨基酸序列gln-asp-gly-asn-glu-glu-x-gly(x是met或ile)的cd3ε的胞外域的片段具有相同的特异性)的方法可包括下列用于选择抗原特异性实体的方法的一个或多个步骤:
cd3ε特异性结合域可选自抗体衍生库。噬菌体展示文库可根据如描述于“phagedisplay:alaboratorymanual”;ed.barbas,burton,scott&silverman;coldspringharborlaboratorypress,2001的标准操作程序来构建。抗体库中的抗体片段的形式可以是scfv,但一般而言也可以是fab片段或甚至单域抗体片段。为了分离抗体片段,可使用天然的抗体片段文库。对于选择潜在的低免疫原性的结合实体用于以后的治疗,人类抗体片段文库对于人类抗体片段的直接选择可以是有利的。在一些情况下,它们可形成合成抗体文库的基础(knappik等人jmol.biol.2000,296:57ff)。相对应的形式可以是fab、scfv(如下所述的)或域抗体(dabs,如在holt等人,trendsbiotechnol.2003,21:484ff中所综述的)。
在本领域中还已知的是:在许多情况下没有对抗靶标抗原的可用的免疫人类抗体源。因此用靶标抗原和分离自动物组织例如脾脏或pbmc的各自的抗体文库免疫动物。n-端片段可被生物素化或共价地连结至蛋白质,如klh或牛血清白蛋白(bsa)。根据常见的方法,啮齿类动物被用于免疫。一些非人类来源的免疫抗体库可以因为其他原因(例如源自骆驼类物种的单域抗体(vhh)的存在)而特别有利(如描述于muyldermans,jbiotechnol.74:277;degenst等人devcomoimmunol.2006,30:187ff)。因此抗体文库的对应形式可以是fab、scfv(如下所述的)或单域抗体(vhh)。
在一个可行的方法中,来自balb/cxc57black杂交的10周龄f1小鼠可以用全细胞免疫,所述全细胞例如表达n-端呈现为翻译融合成熟的cd3ε链的n-端氨基酸1至27的跨膜epcam。或者,小鼠可用1-27cd3ε-fc融合蛋白免疫(相应的方法描述于wo2008/119567的实施例中)。在加强免疫之后,取出血液样品并且对抗cd3-阳性t细胞的抗体血清效价可在如facs分析中进行测试。通常,血清效价在免疫的动物中显著地高于在非免疫的动物中。
免疫的动物可形成构建免疫抗体文库的基础。这种文库的实例包括噬菌体展示文库。这种文库一般而言可根据如在“phagedisplay:alaboratorymanual”;ed.barbas,burton,scott&silverman;coldspringharborlaboratorypress,2001中公开的标准操作程序构建。
非人类抗体也可通过噬菌体展示而被人源化,这是由于产生更多可变抗体文库,这些文库可随后在选择期间针对结合物进行富集。
在噬菌体展示方法中,呈现抗体文库的噬菌体集合中的任一个都形成用各自抗原作为靶标分子选择结合实体的基础。分离抗原特异性的抗原结合的噬菌体的中心步骤称为淘选。因为抗体片段展示在噬菌体的表面上,这个通用的方法被称为噬菌体展示。选择的一个优选方法为使用小蛋白,如经翻译融合至由噬菌体展示的scfv的n-端的丝状噬菌体n2域。本领域已知的可用来分离结合实体的另一种展示方法是核糖体展示方法(在groves&osbourn,expertopinbiolther.2005,5:125ff;lipovsek&pluckthun,jimmunolmethods2004,290:52ff中综述的)。为了证明scfv噬菌体颗粒结合至1-27cd3ε-fc融合蛋白,带有经克隆的scfv谱的噬菌体文库可通过peg(聚乙二醇)从各自的培养上清液中收获。scfv噬菌体颗粒与被固定的cd3εfc融合蛋白一起培养。该被固定的cd3εfc融合蛋白可被涂覆到固相。可以洗脱结合实体,然后洗脱液可用于感染新鲜的未被感染的细菌宿主。被编码人类scfv片段的噬菌粒拷贝成功转导的的细菌宿主可就羧苄西林抗性再次进行筛选,并且随后用例如vcms13辅助噬菌体感染,以开始第二轮的抗体展示以及体外筛选。通常总共进行4至5轮筛选。可以使用流式细胞仪,在cd3ε阳性jurkat细胞、hpball细胞、pbmc或经转染的携带融合至表面展示的epcam的n-端cd3ε序列的真核细胞上测试经分离的结合实体的结合(参见wo2008/119567)。
优选地,上述方法可以是以下方法:其中cd3ε的胞外域的片段由具有seqidno.2、seqidno.4、seqidno.6或seqidno.8中所示的任一个的氨基酸序列的多肽的一个或多个片段组成。更优选地,所述片段的长度是8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27个氨基酸残基。
这个识别双特异性单链抗体分子的方法可以是一种筛选多个包含结合至人类以及非黑猩猩灵长类动物cd3ε的表位的跨物种特异性结合域的双特异性单链抗体分子的方法。或者,识别的方法是一种纯化/分离包含结合至人类以及非黑猩猩灵长类动物cd3ε的表位的跨物种特异性结合域的双特异性单链抗体分子的方法。
此外,本发明提供一种组合物,所述组合物包含本发明的双特异性单链抗体分子或通过如上所述的方法生产的双特异性单链抗体。优选地,所述组合物是一种药物组合物。
本发明还提供如本文定义的或根据如本文定义的方法生产的双特异性单链抗体分子,其中所示双特异性单链抗体分子用于预防、治疗或改善癌症。优选地,所述癌症是实体瘤,更优选地,为癌或前列腺癌。优选地,该双特异性单链进一步包含载体、稳定剂和/或赋形剂的适合制剂。此外,优选地,所示双特异性单链抗体分子适合与另外的药物组合给药。所述药物可以是非蛋白质性的化合物或蛋白质性的化合物,并且可与如本文定义的双特异性单链抗体分子同时地或非同时地给药。
根据本发明,术语“药物组合物”涉及用于向患者给药的组合物,优选人类患者。特别优选的本发明的药物组合物包括靶向对抗环境独立的cd3表位且针对其产生的双特异性单链抗体。优选地,所述药物组合物包括载体、稳定剂和/或赋形剂的适合制剂。在一个优选实施方案中,所述药物组合物包括用于胃肠外的、透皮的、腔内的、动脉内的、鞘内和/或鼻内给药或者通过直接注射到组织内的组合物。特别设想的是所述组合物通过输注或注射向患者给药。适合的组合物的给药可有许多方式,例如,通过静脉内的、腹膜内的、皮下的、肌内的、局部的或皮内给药。特别地,本发明提供该适合的组合物不间断的给药。作为非限制性的实例,可通过患者所戴的计量治疗剂到达患者体内的量的小泵系统实现不间断的即连续给药。该包含靶向对抗本发明的环境独立的cd3表位且针对其产生的双特异性单链抗体的药物组合物可通过使用所述泵系统进行给药。所述泵系统一般而言是本领域已知的,并且通常依靠周期性地更换含有要被输注的治疗剂的药筒。当更换泵系统中的药筒时,可能接着发生到达患者体内的本应不间断流动的治疗剂的暂时中断。在这样的情况下,在药筒更换前的给药阶段和在药筒更换后的给药阶段仍会被认为是在药学手段的含义内,并且本发明的方法共同构成此类治疗剂的一个“不间断给药”。
这些靶向对抗本发明的环境独立的cd3表位且针对其产生的双特异性单链抗体的连续或不间断的给药可借助流体递送装置或小泵系统(包括用于驱使流体离开储库的流体驱动机构以及用于致动该驱动机构的致动器)进行静脉内或皮下给药。用于皮下给药的泵系统可包括用于穿透患者皮肤并将适合的组合递送到该患者体内的针或插管。无需静脉、动脉或血管,所述泵系统可直接固定或附着至患者皮肤,从而使得泵系统与患者的皮肤可直接接触。所述泵系统可被附着在患者皮肤24小时至数天。所述泵系统可以是具有小体积储库的小尺寸。作为非限制性的实例,用于给药的适合的药物组合物的储库的体积可以是0.1至50ml。
连续给药可以是借助佩戴在皮肤上并定期替换的贴片的透皮给药。本领域技术人员知道适于此目的的用于药物递送的贴片系统。注意到的是:透皮给药特别地适于不间断的给药,因为先被用完的贴片的调换可有利地同时以一片新贴片的替换而完成,例如在皮肤的表面上毗连至先用完的贴片以及立即在移除先用完的贴片之前。不会产生流体中断或动力不足的问题。
包含靶向对抗环境独立的cd3表位且针对其产生的特别的双特异性单链抗体的本发明的组合物可进一步包含药学上可接受的载体。适合的药物载体的实例是本领域熟知的,包括溶液(例如磷酸盐缓冲盐水溶液)、水、乳剂(如油/水乳剂)、各种类型的润湿剂、无菌溶液、脂质体等。包含此类载体的组合物可通过众所周知的常规方法配制。制剂可包括碳水化合物、缓冲溶液、氨基酸和/或表面活性剂。碳水化合物可以是非还原糖,优选海藻糖、蔗糖、八硫酸酯、山梨糖醇或木糖醇。这样的制剂可用于可具有和/或不具有泵系统的静脉内或皮下的连续给药。氨基酸可以是带电荷的氨基酸,优选赖氨酸、醋酸赖氨酸、精氨酸、谷氨酸和/或组氨酸。表面活性剂可以是洗涤剂(优选>1.2kd的分子量)和/或聚醚(优选>3kd的分子量)。优选地,优选洗涤剂的非限制性实例是tween20、tween40、tween60、tween80或tween85。优选聚醚的非限制性实例是peg3000、peg3350、peg4000或peg5000。本发明使用的缓冲剂系统可优选ph5-9,并且可包括柠檬酸盐、琥珀酸盐、磷酸盐、组氨酸和醋酸盐。本发明的组合物可以以适合的剂量给个体给药,所述剂量可例如通过剂量递增研究确定,所述研究通过给非黑猩猩灵长类动物(例如,猕猴)施用剂量渐增的显示具有本文描述的跨物种特异性的本发明的双特异性单链抗体分子进行。如上所述,显示具有本文描述的跨物种特异性的本发明的双特异性单链抗体分子可有利地以在非黑猩猩灵长类动物的临床前试验中以及作为人用药物的同样形式使用。这些组合物也可与其他蛋白质性的和非蛋白质性的药物组合给药。这些药物可与包含如本文定义的本发明的双特异性单链抗体分子的组合物同时给药,或者以适时地被定义的间隔和剂量在施用所述多肽之前或之后分开给药。给药方案是由主治医生和临床因素决定的。如在医学领域中熟知的,任何一位患者的剂量均取决于许多因素(包括患者的体型、体表面积、年龄、给药的特定化合物、性别、给药时间和途径、一般健康状况以及要同时给药的其他药物)。用于肠胃外给药的制剂包括无菌水性或非水性溶液、悬浮液和乳剂。非水性溶液的实例是丙二醇、聚乙二醇、植物油(如橄榄油)以及可注射的有机酯(如油酸乙酯)。水性载剂包括水、醇的/水性溶液、乳剂或悬浮液(包括生理盐水以及缓冲介质)。胃肠外的载体包括氯化钠溶液、林格氏葡萄糖、葡萄糖与氯化钠、乳酸林格氏或固定油。静脉内的载体包括流体和营养补充物、电解质补充剂(如以林格氏葡萄糖为基础的那些)以及类似物。防腐剂和其他添加剂也可存在,例如抗菌剂、抗氧化剂、螯合剂、惰性气体以及类似物。此外,本发明的组合物可包括蛋白质性的载体,例如,血清白蛋白或免疫球蛋白,优选人类来源。据设想,除了本文定义的本发明的双特异性单链抗体分子之外,本发明的组合物可进一步包括生物活性剂,这取决于组合物的预期用途。此类药剂可以是作用于胃肠系统的药物、作用如细胞抑制剂的药物、预防高尿酸血症的药物、抑制免疫反应的药物(例如皮质类固醇)、调节炎性反应的药物、作用于循环系统的药物和/或本领域已知的药剂,例如细胞因子。
本文定义的药物组合物的生物活性可例如通过细胞毒性分析测定,如在下列实例中描述:wo99/54440或schlereth等人(cancerimmunol.immunother.20(2005),1-12)。如本文所用的,“有效性”或“体内有效性“是指使用例如标准化的nci反应标准的通过用本发明的药物组合物治疗的反应。使用本发明的药物组合物治疗成功或体内有效性是指该组合物对其预定目的的有效性,即该组合物引起其所欲的效果的能力,所述效果即,病理细胞(例如癌细胞)的耗竭。体内有效性可通过对不同的疾病实体建立的标准方法进行监测,所述方法包括、但不限于白血细胞计数、分类、荧光活化细胞分选、骨髓穿刺。此外,可使用各种疾病特定临床化学参数和其他建立的标准方法。此外,可使用计算机辅助体层摄影术、x-射线、核磁共振断层扫描(例如,以美国国家癌症研究所标准为基础的反应评估[chesonbd,horningsj,coiffierb,shippma,fisherri,connorsjm,listerta,vosej,grillo-lopeza,hagenbeeka,cabanillasf,klippenstend,hiddemannw,castellinor,harrisnl,armitagejo,carterw,hopper,canellosgp。reportofaninternationalworkshoptostandardizeresponsecriteriafornon-hodgkin'slymphomas.ncisponsoredinternationalworkinggroup.jclinoncol.1999apr;17(4):1244])、正电子发射断层扫描、白血细胞计数、分类、荧光活化细胞分选、骨髓穿刺、淋巴结活检/组织学和各种癌症特定临床化学参数(例如,乳酸脱氢酶)以及其他既定的标准方法。
在开发药物(如本发明的药物组合物)中的另一项重大挑战是可预测的药代动力学性质的调整。为此,建立了候选药物的药代动力学特征,即,影响特定药物在给定病况下治疗能力的药代动力学参数的图谱。影响药物治疗某个疾病实体的能力的药物药代动力学参数包括、但不限于:半衰期、分布容量、肝首过代谢和血清结合的程度。给定药物试剂的有效性可被上面所提到的各个参数影响。
“半衰期”是指50%的施用药物通过生物处理(例如,代谢、排泄等)而被消除的时间。
所谓“肝首过代谢”是指药物在与肝脏第一次接触(即,其第一次穿过肝脏的期间)而被代谢的倾向。
“分布容量”是指遍及身体的各个隔室(例如细胞内和细胞外的空间、组织以及器官等)时药物的保留程度,以及药物在这些隔室内的分布。
“血清结合程度"是指药物与血清蛋白(如白蛋白)相互作用并结合至血清蛋白、从而导致药物生物活性降低或丧失的倾向。
药代动力学参数还包括生物利用度、滞后时间(tlag)、tmax、吸收速率、给定量的施用药物的更多发作(onset)和/或cmax。
“生物有效性”是指药物在血液隔室中的量。
“滞后时间"是指药物的给药和它在血液或血浆中的检出与可测性之间的时间延迟。
“tmax”是在此时间后药物达到最大血药浓度的时间,“cmax”是获得的给定药物的最大血药浓度。达到其生物效应所需的药物的血液或组织浓度的时间受所有参数影响。显示具有跨物种特异性的双特异性单链抗体的药代动力学参数(可在如上所列的非黑猩猩灵长类动物的临床前动物试验中测定)也在例如schlereth等人的刊物(cancerimmunol.immunother.20(2005),1-12)中提出。
如本文所用的,术语“毒性”是指在不良事件或严重的不良事件中表现的药物的毒性效用。这些不良事件可能是指在给药之后药物的整体耐受性的缺乏和/或局部耐受性的缺乏。毒性还可包括药物引起的畸形的或致癌作用。
如本文所用的,术语“安全性”、“体内安全性”或者“耐受性”定义在给药之后(局部耐受性)以及在药物应用的较长期间,药物的给药没有直接地诱发严重的不良事件。“安全性”、“体内安全性”或者“耐受性”可例如在治疗以及后续时间段期间在规律的间隔下进行评估。测试包括临床评价(例如器官表现)以及实验室异常的筛选。可进行临床评价,并且根据nci-ctc和/或meddra标准,可记录或编码偏离正常的结果。器官表现可包括以下标准,如过敏/免疫、血液/骨髓、心律失常、凝血功能以及如在常见不良事件评价标准v3.0(ctcae)中所提出的类似标准。可被测试的实验室参数包括例如血液学、临床化学、凝血情况与尿液分析以及其他体液如血清、血浆、淋巴或脊髓液、液体的检查以及类似参数。因此,安全性可例如通过体检、影像学技术即超声、x-射线、ct扫描、磁共振成像(mri)、使用技术设备(如心电图)的其他测量、生命体征,通过测量实验室参数和记录不良事件来评估。例如,根据本发明的用途和方法,非黑猩猩灵长类动物的不良事件可通过组织病理学和/或组织化学的方法检查。
如本文所用的,术语“有效且无毒的剂量”是指如本文定义的双特异性单链抗体的可耐受的剂量,该剂量高到足以耗竭病理细胞、肿瘤消除、肿瘤缩小或稳定疾病而没有或基本上没有主要毒性作用。所述有效且无毒的剂量可例如通过本领域描述的剂量递增研究测定确定并且应该低于诱发严重不良事件的剂量(即剂量限制毒性,dlt)。
上述术语也参考例如preclinicalsafetyevaluationofbiotechnology-derivedpharmaceuticalss6;ichharmonisedtripartiteguideline;ichsteeringcommitteemeetingonjuly16,1997。
此外,本发明涉及用于预防、治疗或改善癌症的包含本发明的或根据本发明的方法生产的双特异性单链抗体分子的药物组合物。优选地,所述癌症是实体瘤(优选癌或前列腺癌)。优选地,所述药物组合物进一步包含载体、稳定剂和/或赋形剂的适合制剂。
另一方面,本发明涉及如上述定义的或根据上述定义的方法生产的双特异性单链抗体分子/多肽用于制备预防、治疗或改善疾病的药物组合物的用途。优选地,所述疾病是癌症。更优选地,所述癌症是实体瘤(优选癌或前列腺癌)。
在本发明的双特异性单链抗体分子的用途的另一个优选实施方案中,所述药物组合物适于与另外的药物(即,作为共同疗法的一部分)组合给药。在所述共同疗法中,活性剂可任选地与本发明的双特异性单链抗体分子包含在相同的药物组合物中或者可被包含在分开的药物组合物中。在后一种情况下,所述分开的药物组合物适于在所述包含本发明的双特异性单链抗体分子的药物组合物的给药前、同时或之后给药。所述另外的药物或药物组合物可以是非蛋白质性的化合物或蛋白质性的化合物。在所述另外的药物是蛋白质性的化合物的情况下,有利的是该蛋白质性的化合物能够提供用于免疫效应细胞的激活信号。
优选地,所述蛋白质性的化合物或非蛋白质性的化合物可与本发明的双特异性单链抗体分子、如上文定义的核酸分子、如上文定义的载体或如上文定义的宿主同时地或非同时地给药。
本发明的另一方面涉及一种用于在有此需要的受试者中预防、治疗或改善疾病的方法,所述方法包括施用有效量的本发明的药物组合物的步骤。优选地,所述疾病是癌症。优选地,所述癌症是实体瘤(优选癌或前列腺癌)。
在本发明的方法的另一个优选实施方案中,所述药物组合物适于与另外的药物(即,作为共同疗法的一部分)组合给药。在所述共同疗法中,活性剂可任选地与本发明的双特异性单链抗体分子包含在相同的药物组合物中或者可被包含在分开的药物组合物中。在后面的这种情况下,所述分开的药物组合物适合于在所述包含本发明的双特异性单链抗体分子的药物组合物的给药前、同时或之后给药。所述另外的药物或药物组合物可以是非蛋白质性的化合物或蛋白质性的化合物。在所述另外的药物是蛋白质性的化合物的情况下,有利的是该蛋白质性的化合物能够提供用于免疫效应细胞的激活信号。
优选地,所述蛋白质性的化合物或非蛋白质性的化合物可与本发明的双特异性单链抗体分子、如上文定义的核酸分子、如上文定义的载体或如上文定义的宿主同时地或非同时地给药。
对上述的本发明的方法优选地,所述受试者是人。
又一方面,本发明涉及含有本发明的双特异性单链抗体分子、本发明的核酸分子、本发明的载体或本发明的宿主的试剂盒。
这些以及其他实施例公开并包含于本发明的描述部分和实施例中。免疫学的重组技术和方法描述于例如sambrook等人molecularcloning:alaboratorymanual;coldspringharborlaboratorypress,3rdedition2001;lefkovits;immunologymethodsmanual;thecomprehensivesourcebookoftechniques;academicpress,1997;golemis;protein-proteininteractions:amolecularcloningmanual;coldspringlaboratorypress,2002。关于根据本发明采用的抗体、方法、用途和化合物中任一个的其它文献可利用例如电子设备在公共图书馆查询系统和数据库检索。例如,可利用在互联网可获得的公共数据库“medline”,例如在http://www.ncbi.nlm.nih.gov/pubmed/medline.html上。其他的数据库和网址,例如http://www.ncbi.nlm.nih.gov/或者在http://www.embl.de/services/index.html上所列的embl服务首页,均是本领域技术人员已知的,并且,还可使用例如http://www.google.com获得。
附图说明
图1
指定的跨物种特异性的双特异性单链构建体与用人类psma转染的cho细胞、人类cd3+t细胞株hpb-all、用猕猴psma转染的cho细胞以及猕猴t细胞株4119lnpx的facs结合分析。如实施例2.1中所述进行facs染色。粗线代表随后用抗-his抗体和pe标记的检测抗体孵育的用细胞培养上清液孵育的细胞。细直方图线反映阴性对照:仅用抗-his抗体和检测抗体孵育的细胞。
其中,
图1(1):psma76-b10衍生物;
图1(2):psma76-b10衍生物;
图1(3):psma34c7衍生物;
图1(4):psma29-g1衍生物;
图2:
由重新定向到靶细胞株的指定的跨物种特异性的双特异性单链构建体诱导的细胞毒活性。经刺激的cd4-/cd56-人类t细胞用作效应细胞,用人类psma转染的cho细胞用作靶细胞。分析如实施例2.2所述进行。
其中,
图2(1):psma特异性
a)效应细胞:经刺激的人类t细胞;靶细胞:用人类psma转染的cho细胞;b)效应细胞:经刺激的人类t细胞;靶细胞:用人类psma转染的cho细胞;
图2(2):psma特异性
c)效应细胞:经刺激的人类t细胞;靶细胞:用人类psma转染的cho细胞;d)效应细胞:经刺激的人类t细胞;靶细胞:用人类psma转染的cho细胞;
图2(3):psma特异性
e)效应细胞:经刺激的人类t细胞;靶细胞:用人类psma转染的cho细胞;
本发明此外借助以下示例性的非限制性实施例进行描述,所述实施例使得更好地理解本发明及其多个优点。
具体实施方式
实施例1psma和cd3跨物种特异性的双特异性单链抗体分子的产生与表征
1.1人类psma抗原在cho细胞上的克隆与表达
通过根据标准操作程序的基因合成,将人类psma抗原的序列(‘ay101595’,智人前列腺特异膜抗原mrna,完整的cds,美国国家生物技术信息中心,http://www.ncbi.nlm.nih.gov/entrez)用于获得合成分子。基因合成片段还被设计为含有用于构建体真核表达的kozak位点和在dna的起始以及末端的限制性位点。被导入的限制性位点(在5'端的xbai和在3'端的sali)在克隆步骤期间被用在mack等人(mackm等人,procnatlacadsciusa1995;92:7021-5以及raum等人cancerimmunolimmunother(2001)50(3))描述的称为pefdhfr的表达质粒内。在序列确认之后,使用质粒转染cho/dhfr-细胞,如下所述。经序列确认的质粒被用来转染cho/dhfr-细胞(atccno.crl9096;培养于具有获自biochromagberlin,germany的稳定的谷氨酰胺的rpmi1640中,所述rpmi1640补充有10%fcs、1%青霉素/链霉素(全部获自biochromagberlin,germany)和来自细胞培养级试剂的母液的核苷(获自sigma-aldrichchemiegmbh,taufkirchen,germany)(终浓度为10μg/ml腺苷、10μg/ml脱氧腺苷以及10μg/ml胸苷),在培养箱(37℃、95%湿度以及7%co2)中)。根据制造商的操作规程,使用polyfect转染试剂(qiagengmbh,hilden,germany)和5μg的质粒dna进行转染。在24小时的培养后,用pbs洗涤细胞一次,并且在上述培养基中再次培养(该培养基没有补充核苷和经透析的fcs(获自biochromagberlin,germany获得))。因此该细胞培养基不含核苷并由此在经转染的细胞上进行筛选。在转染之后大约14天,观察到抗性细胞生长。在另外7至14天后,转染子经由facs测试呈阳性即表达构建体。在dhfr缺乏的cho细胞中真核生物的蛋白质表达如由kaufmannrj(1990)methodsenzymol.185,537-566描述地进行。构建体的基因扩增通过增加甲氨喋呤(mtx)的浓度至高达20nmmtx的最终浓度而被诱导。
1.2猕猴psma抗原在cho细胞上的克隆与表达
猕猴psma(石蟹猕猴)的cdna序列通过在来自根据标准规程制备的猕猴前列腺的cdna上的一组5次pcr获得。下列的反应条件:1个循环的在94℃下历时2分钟,然后以94℃历时1分钟、52℃历时1分钟以及72℃历时1.5分钟进行40个循环,接着进行72℃历时3分钟的最终循环,并且使用以下引物:
·正向引物:5'-cactgtggcccaggttcgagg-3'(seqidno.213)
反向引物:5'-gacataccacacaaattcaatacgg-3'(seqidno.214)
·正向引物:5'-gctctgctcgcgccgagatgtgg-3'(seqidno.215)
反向引物:5'-acgctggacaccacctccagg-3'(seqidno.216)
·正向引物:5'-ggttctactgagtgggcagagg-3'(seqidno.217)
反向引物:5'-acttgttgtggctgcttggagc-3'(seqidno.218)
·正向引物:5'-gggtgaagtcctatccagatgg-3'(seqidno.219)
反向引物:5'-gtgctctgcctgaagcaattcc-3'(seqidno.220)
·正向引物:5'-ctcggcttcctcttcgggtgg-3'(seqidno.221)
反向引物:5'-gcatattcatttgctgggtaacctgg-3'(seqidno.222)
这些pcr产生5个重叠片段,所述片段根据标准规程使用pcr引物进行分离和测序,并且由此提供编码猕猴psma的成熟蛋白质的自第3个密码子至最后一个密码子的cdna序列的一部分。为了产生用于表达猕猴psma的构建体,提供根据标准规程的基因合成获得cdna片段(构建体的cdna和氨基酸序列列于seqidno.223和224)。在这个构建体中,其后为终止密码子的成熟psma蛋白质的第3个氨基酸至最后一个氨基酸的猕猴psma的编码序列在框架内被融合至人类psma蛋白质的前2个氨基酸的编码序列。该基因合成片段还被设计为含有用于构建体的真核表达的kozak位点和在该含有cdna的片段的起始以及末端的限制性位点。被导入的限制性位点(在5'端的xbai以及在3'端的sali)在下列克隆操作程序中使用。基因合成片段根据标准规程经xbai和sali被克隆至称为pef-dhfr的质粒内。上述的操作程序根据标准的规程(sambrook,molecularcloning;alaboratorymanual,3rdedition,coldspringharbourlaboratorypress,coldspringharbour,newyork(2001))进行。具有经序列确认的核苷酸序列的克隆被转染至dhfr缺乏的cho细胞内,以用于构建体的真核表达。在dhfr缺乏的cho细胞中真核生物的蛋白质表达如由kaufmannrj(1990)methodsenzymol.185,537-566描述地进行。构建体的基因扩增通过增加甲氨喋呤(mtx)的浓度至高达20nmmtx的最终浓度而被诱导。
实施例2
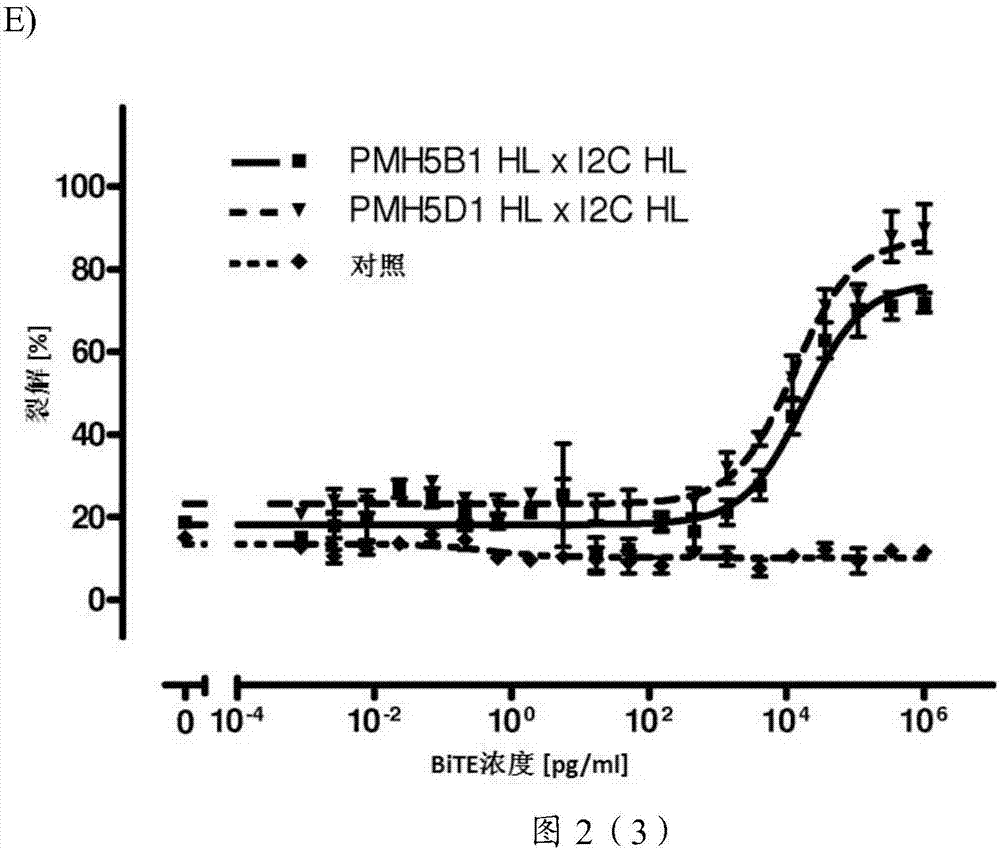
2.1psma和cd3跨物种特异性的双特异性抗体的流式细胞术结合分析
为了测试跨物种特异性的双特异性抗体构建体关于对人类和猕猴psma以及对人类和猕猴cd3的结合能力的功能,进行了facs分析。为此,将用人类psma转染的cho细胞和人类cd3阳性t细胞白血病细胞系hpb-all(dsmz,braunschweig,acc483)用于检查对人类抗原的结合。通过使用产生的猕猴psma转染子和猕猴t细胞系4119lnpx(由fickenscher教授(hygieneinstitute,virology,erlangen-nuernbe)友情提供;公开于knappea等人,和fickenscherh.,blood2000,95,3256-61中]测试对猕猴抗原的结合反应性。
流式细胞术分析进行如下:
不同细胞株的200.000个细胞与50μl的经纯化的跨物种特异性的双特异性抗体构建体的蛋白质(2μg/ml)或表达跨物种特异性的双特异性抗体构建体的转染细胞的培养上清液在冰上孵育,历时30分钟。这些细胞在具有2%fcs的pbs中洗涤2次,然后用鼠抗-his抗体(pentahis抗体;qiagen;在50μl具有2%fcs的pbs中以1:20稀释)检测该构建体的结合。洗涤之后,用被缀合至藻红素的fcγ-特异性抗体(dianova)(在具有2%fcs的pbs中以1:100稀释)检测结合的抗-his抗体。未被转染的cho细胞的上清液用作结合至t细胞系的阴性对照。具有不相关的靶标特异性的单链构建体用作结合至经psma转染的cho细胞的阴性对照。
在facs-calibur设备上进行流式细胞术;cellquest软件用于获得和分析数据(bectondickinsonbiosciences,heidelberg)。facs染色和测量荧光强度如在currentprotocolsinimmunology(coligan,kruisbeek,margulies,shevachandstrober,wiley-interscience,2002)中描述地进行。
可清楚地检测到对psma跨物种特异性的和对人类与猕猴cd3跨物种特异性的单链分子的双特异性结合,如图1所示。在facs分析中,相比于各自的阴性对照,所有构建体均显示结合至cd3和psma。证明了双特异性抗体对人类和猕猴cd3以及对人类和猕猴psma抗原的跨物种特异性。
2.2psma和cd3跨物种特异性的双特异性单链抗体的生物活性
使用人类psma阳性细胞系cho细胞,在体外细胞毒性分析中通过铬51(51cr)释放,分析产生的双特异性单链抗体的生物活性。作为效应细胞,使用经刺激的人类cd4/cd56耗竭的pbmc。
经刺激的cd4/cd56耗竭的pbmc的产生如下进行:
在37℃下,用终浓度为1μg/ml的市售抗-cd3特异性抗体(例如okt3,othoclone)涂布皮氏培养皿(145mm直径,greinerbio-onegmbh,frickenhausen)1小时。未结合的蛋白质通过pbs的一个洗涤步骤而被去除。根据标准规程通过ficoll密度梯度离心从外周血(30-50ml人类血液)分离新鲜的pbmc。将3-5x107pbmc添加至已涂布的皮氏培养皿中的120ml的rpmi1640中,所述rpmi1640具有稳定的谷氨酰胺/10%fcs/il-220u/ml(proleukin,chiron),并且刺激2天。在第3天,收集这些细胞并且用rpmi1640洗涤1次。将il-2添加至20u/ml的最终浓度,并且这些细胞在与上述相同的细胞培养基中再次培养1天。通过根据标准规程耗竭cd4+t细胞和cd56+nk细胞,富集了cd8+细胞毒性t淋巴细胞(ctl)。
用pbs清洗靶细胞2次并且在37℃下,在最终体积为100μl的具有50%fcs的rpmi中用11.1mbq51cr进行标记,历时45分钟。随后用5mlrpmi洗涤这些被标记的靶细胞3次并且接着用于细胞毒性分析中。该分析在96孔板中、在250μl总体积的具有e:t比10:1的补充的rpmi(如上面)中进行。应用1μg/ml的跨物种特异性的双特异性单链抗体分子和其20份3倍稀释液。分析时间是18小时,并且以在上清液中被释放的铬相对于最大裂解(添加triton-x)与自发裂解(无效应细胞)的差值的相对值,测量细胞毒性。所有测量重复四次。上清液中的铬活性的测量用wizard3”γ计数器(perkinelmerlifesciencesgmbh,
如图2所示,所有产生的跨物种特异性的双特异性单链抗体构建体证明了生物毒活性对抗由经刺激的人类cd4/cd56耗竭的pbmc所引发的psma阳性靶细胞。
实施例3scfv对抗各种细胞系的结合分析
3.1.在大肠杆菌中单链抗体构建体的表达
scfv分子epcam4-7(wo99/25818)、pm74-g3、pm52-h3、pm52-c3、pm75-a10以及pm91-b6通过使用质粒pcomb3h5bflag/his表达,其中表达构建体(例如scfv)包括flag-标签(dykddddk)和his6-标签。各个scfv分子的质粒dna被转化到100μl热休克感受态大肠杆菌tg1中,然后在羧苄西林lb-琼脂上进行涂板。在用1mmiptg诱导后,用含有vl-和vh-区段的pcomb3h5bflag/his转化的大肠杆菌产生足够数量的可溶性scfv。由于适合的信号序列,该scfv-链被输出至细胞周质内,在该处它折叠成功能性构象。
挑取来自转化板的单个大肠杆菌tg1细菌菌落,用于细胞周质的小规模制备并且在补充了20mmmgcl2和羧苄西林50μg/ml的sb-培养基(例如10ml)中生长,以及在收集后再溶解于pbs(例如1ml)。通过4轮的在-70℃下冷冻、在37℃下解冻,细菌的外膜因为温度冲击被破坏,并且可溶性的细胞周质蛋白(包括scfv)被释放至上清液中。在通过离心去除完整的细胞和细胞碎片之后,收集含有抗-psmascfv的上清液并且用于结合至不同细胞系的测定。
3.2单链抗体构建体对各种细胞系的流式细胞术结合分析
产生scfv分子epcam4-7、pm74-g3、pm52-h3、pm52-c3、pm75-a10以及pm91-b6的大肠杆菌克隆的细胞周质制备物被用来检验对人类psma或人类epcam转染的细胞株的特异性结合。未被转染的cho细胞用作阴性对照。
关于流式细胞术,用50μl的scfv细胞周质制备物孵育2,5x105个细胞。scfv对这些细胞的结合用在50μl具有2%fcs的pbs中的2μg/ml抗-his抗体(penta-his抗体,没有bsa,qiagengmbh,hilden,frg)检测。作为第二步骤试剂,使用r-藻红素-缀和的亲和纯化的f(ab')2片段、山羊抗小鼠igg(特定的fc-γ片段)(在50μl具有2%fcs的pbs(dianova,hamburg,frg)中以1:100稀释)。样品在facsscan(bdbiosciences,heidelberg,frg)上测量。
流式细胞术在facs-calibur设备上进行;cellquest软件用于获得和分析数据(bectondickinsonbiosciences,heidelberg)。facs染色和测量荧光强度如在currentprotocolsinimmunology(coligan,kruisbeek,margulies,shevachandstrober,wiley-interscience,2002)中描述地进行。
scfvepcam4-7显示出对人类epcam转染的cho细胞的强结合,但对人类psma转染的或未被转染的细胞无显著的结合。相比之下,scfvpm74-g3、pm52-h3、pm52-c3、pm75-a10以及pm91-b6显示出对人类psma转染的cho细胞的强结合,但没有对人类epcam转染的cho细胞或未被转染的cho细胞的强结合(不同的scfv的细胞周质细胞提取物在被转染的不同细胞系上的结合结果见表1)。
表1.facs分析的结果:'+'表示结合信号,'–'表示无显著的结合信号
- 还没有人留言评论。精彩留言会获得点赞!
- 用于治疗HBV感染和相关病症的三特异性结合分子的制造方法与工艺
- 对磺胺类药物具有特异性的固相微萃取纤维的制备方法与流程
- 一种检测猪初乳中抗PEDV特异性IgA抗体的夹心ELISA试剂盒的制造方法与工艺
- 用于施用CD19xCD3双特异性抗体的给药方案的制造方法与工艺
- 双特异性抗‑VEGF/抗‑ANG‑2抗体及其在治疗眼血管疾病中的应用的制造方法与工艺
- 特异性人EWS‑FLI1融合蛋白抗体的制备方法及在制备尤文肉瘤诊断抗体试剂的应用与制造工艺
- 双特异性四价抗体及其制造和使用方法与制造工艺
- 利用特异性挥发物筛选昆虫相关嗅觉蛋白的方法与制造工艺
- 使用与表面偶联的单特异性四聚抗体复合物组合物从样品中分离出靶实体的方法与制造工艺
- 抗-WT1/HLA双特异性抗体的制造方法与工艺
- 与CD56分子特异性结合的多肽及其应用的制造方法与工艺
- 用于治疗HBV感染和相关病症的三特异性结合分子的制造方法与工艺
- 对磺胺类药物具有特异性的固相微萃取纤维的制备方法与流程
- 避免信号干扰的肌原纤维蛋白丝氨酸磷酸化水平的测定方法与流程
- 一种检测猪初乳中抗PEDV特异性IgA抗体的夹心ELISA试剂盒的制造方法与工艺
- 用于施用CD19xCD3双特异性抗体的给药方案的制造方法与工艺
- 双特异性抗‑VEGF/抗‑ANG‑2抗体及其在治疗眼血管疾病中的应用的制造方法与工艺
- 特异性人EWS‑FLI1融合蛋白抗体的制备方法及在制备尤文肉瘤诊断抗体试剂的应用与制造工艺
- 活化人CD40的抗体和针对人PD‑L1的抗体的组合疗法的制造方法与工艺
- 双特异性四价抗体及其制造和使用方法与制造工艺