能显著提高棉花抗病性的GhLMM基因及其应用的制作方法
本发明涉及一种棉花的GhLMM基因,其编码5-氨基乙酰丙酸脱水酶(ALAD)。利用首次在陆地棉中发现的类病变突变体Ghlmm,通过图位克隆技术得到该基因。在Ghlmm突变体中,由于位于D亚组的GhLMM基因,其一个单核苷酸的SNP突变造成编码区提前终止,功能丧失。抗病性检测表明该突变体抗黄萎病性显著提高。进一步通过PCR技术在陆地棉遗传标准系TM-1A亚组和D亚组中获得该基因的基因组序列,全长ORF序列及编码的氨基酸序列。在四倍体棉花中,该基因含四个拷贝,利用生物技术调控该基因在四倍体棉花中表达剂量,可以提高棉花抗病性,属于生物技术应用领域。
背景技术:
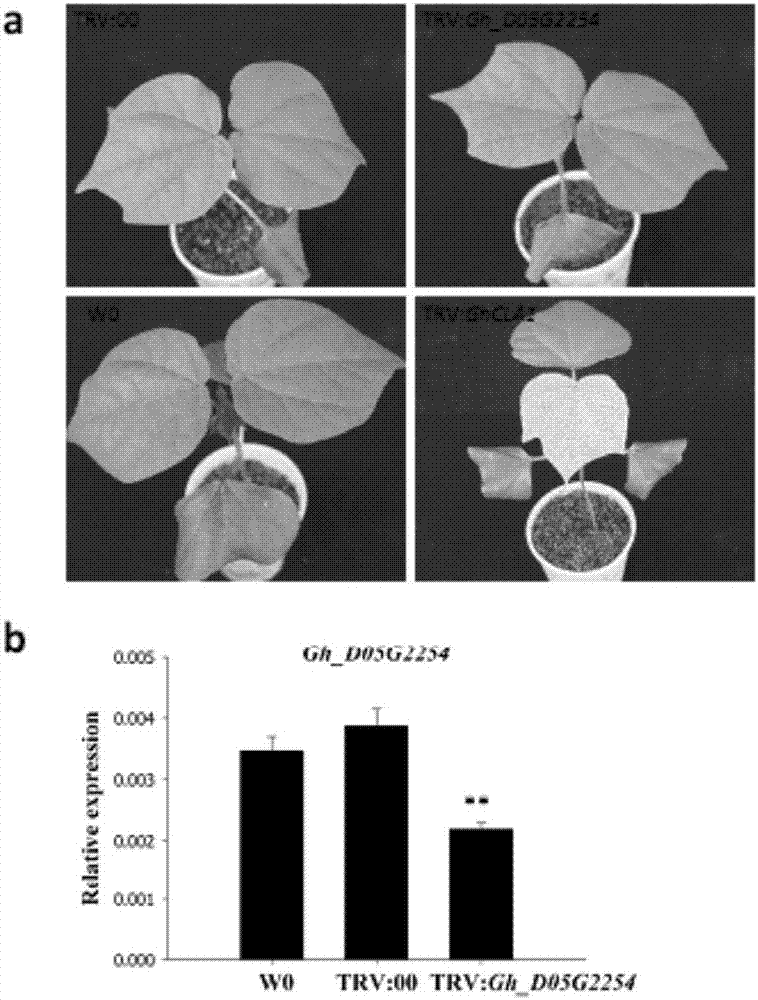
:细胞程序性死亡(programmedcelldeath,PCD)是多细胞生物在信号分子的作用下由基因调控的细胞主动有序的死亡过程,是有机体生长发育和抵制外界胁迫的重要方式之一。根据诱导产生PCD的因素,可划分为生长发育过程中的PCD和环境诱导的PCD。生长发育过程中的PCD是植物正常生长发育所必须的,其参与植物组织器官的分化和形态建成、保证功能性孢子和配子体的发育、生殖过程的完成。而环境诱导的PCD主要指盐,干旱等非生物胁迫以及病原菌侵染等生物胁迫产生的超敏反应,通过受侵染部位的快速细胞死亡,起到隔离作用,达到保护临近健康组织目的。在遇到病原菌等生物胁迫中,植物为了应对病菌的攻击,进化出了复杂的信号和防卫机制来保护自己。概括分为自身的基本抗性和病菌诱导的抗性两大类。当病原菌侵染时,其表面的病原相关分子特征PAMP(pathogen-associatedmolecularpatterns)被植物特异的病原识别受体PRR(pathogenesis-relatedproteins)所识别,激发PAMP激活的免疫反应(PTI)。目前已鉴定出来的病菌相关分子包括,革兰氏阴性菌的鞭毛蛋白,脂多糖真菌和卵菌的几丁质和葡聚糖。PTI表现在植物利用自身固有的介质来抵抗外界病原物的入侵,如:细胞壁的蜡质,角质,小分子抗病物质(酚类化合物、富半胱氨酸抗微生物多肽以及相关的过氧化物酶)来破坏病原菌渗调蛋白和核糖体失活蛋白。PTI是抵御病菌侵染的第一道防线。当病菌突破植物的第一道防线之后,分泌效应蛋白进入植物体内,被植物的R蛋白识别后,触发效应蛋白激活的免疫反应(ETI)。该防御体系又可分为局部抗性和系统获得性抗性。其中,局部抗性是指超敏反应(HR)。当植物受到病菌的侵染后,在侵染部位或者附近迅速局部死亡,形成病斑。阻断病菌营养供给,同时诱导周围细胞产生抑制病原菌生长的物质,以达到限制病菌进一步增殖。在HR过程中会导致细胞程序性死亡,其过程在相关基因精准调控下完成。与局部抗性相对应的是系统获得性抗性(SAR)。在病菌侵染后的几天到一周后,植株整体水平产生新的抗性,对病菌的再一次侵染会产生抵抗力。SAR的标志事件就是植株SA含量升高,PR基因表达量升高。研究表明,植物的抗病信号通路十分复杂,许多植物激素参与其中,如:SA,JA,乙烯,一氧化氮,ROS等。各种激素信号之间相互联系,相互作用,在正常野生型中研究十分困难,利用突变体可加速抗病机制研究进程。类病变突变体是在没有受到生物和非生物的胁迫下,植物自身产生类似病菌侵染的表型的突变体。类病变突变体上病斑的产生,类似于植物超敏反应的细胞死亡过程。因此,类病变突变体不仅是揭示细胞程序性死亡的有力工具,还是研究植物抗病信号通路和抗病分子机制的优异材料。目前,至少60余种类病变突变体在玉米,水稻,大麦,拟南芥中发现。遗传分析表明,大多数的类病变突变体都是隐性突变。通过正向遗传学,导致突变体的一些基因已经被克隆。其突变基因主要涉及:1、叶绿素的合成和降解过程。2、鞘脂类和氧化脂类等脂肪酸的代谢过程。3、植物的防卫免疫通路。4、胞内运输过程。5、核内信号的整合及调控基因表达过程(Bruggemanetal.,2015)。研究表明,虽然有少数类病变突变体对病菌更加敏感,大多数的类病变突变体表现为增强的病菌抗性。例如,叶绿素的合成和降解过程均会导致植株类病变表型。在植物四吡咯代谢通路中的基因突变会导致类病变突变体的形成,且植株抗病性增强。叶绿素a、b,亚铁血红素,血红素都属于四吡咯化合物,其正常的合成和代谢对光合作用,呼吸作用,氮硫同化等过程,意义重大。拟南芥rug1突变体,是由于胆色素原脱氨酶(PBGD)功能缺失,造成植物体内积累胆色素原(PBG)。表现为叶片形状变化,伴有坏死斑产生。其开花期延迟,株高变矮,生长发育受到抑制。在rug1突变体中,体内ROS和SA含量增加,抗病相关基因表达升高(Quesadaetal.,2013)。拟南芥的LIN1编码粪卟啉原氧化酶(CPO),其lin1突变体的叶片和长角果上具有病斑。LIN1基因突变造成粪卟啉原的积累和ROS含量升高,组成型表达抗病相关基因,增强对病菌的抗性。进一步研究发现,该突变体与转nahG拟南芥(不能积累SA的转基因材料)杂交其病斑并没有消失,但PR1基因的表达水平降低,说明lin1病斑的产生不依赖SA,但lin1中PR基因的高表达依赖SA(Sunetal.,2011)。FHY3和FAR1是拟南芥转座酶派生的转录因子。拟南芥fhy3far1双突变体叶片上具有自发形成的坏死斑,属于类病变突变体。突变体植株矮小,生长发育受到影响。研究表明,fhy3far1突变导致其下游HEMB基因下调表达,引起ROS的增加。并且SA信号通路激活,PR基因上调表达,表现为抗性增强(Wangetal.2015)。玉米LES22编码l尿卟啉原脱羧酶(UROD),在les22突变体中,积累尿卟啉原,造成玉米叶片产生病斑,并且病斑的产生与光照和植物生长年龄相关(Huetal.,1998)。叶绿素代谢的缺陷同样导致细胞死亡。当叶绿素降解过程中的酶被破坏之后,也会产生自发的细胞死亡的类病变表型。如:acd1突变体是因为破坏了脱镁叶绿酸加氧酶(PAO)基因,造成了光敏的脱镁叶绿酸a的积累,引起细胞死亡(Tanakaetal.,2003)。综上所述,对类病变突变体的突变基因研究有利于揭示PCD与抗病作用机理,阐明抗病信号通路间相互关系,为植物抗病分子育种提供理论指导。棉花是一种重要的经济作物,不仅提供天然纤维,也是重要的油料作物。在棉花的生长过程中会遭受很多病害的威胁,其中黄萎病是目前对棉花威胁最大的病害之一。感染黄萎病后,棉花叶片会黄化、萎蔫、脱落造成棉花产量品质下降,甚至绝收。目前我国黄萎病感染面积达到250万公顷,造成几十亿的经济损失。我国发现的致病的黄萎病为大丽轮枝菌(Verticilliumdahliae),它是一种土传真菌。其发病过程由下向上,首先是叶片边缘失水萎蔫,之后形成西瓜皮状的叶上斑点,最后叶片焦枯脱落,变成光杆。针对黄萎病防治主要采用化学控制和培育高抗品种方法。但是由于黄萎病是通过植物的维管系统进行运输,寄生在植物体内,外施农药不能有效的缓解黄萎病病害。植物死亡之后,黄萎病菌可以产生微菌核,它可以在土壤内存活十年之久。当植物根部受到损伤,产生渗出液,微菌核便可复苏,通过伤口进入植物体内。目前只有熏蒸的方法可以杀死土壤中的黄萎病菌,但是土壤熏蒸对环境有极大的影响,并且耗费大量财力。所以培育抗黄萎病的棉花品种就显得尤为重要。陆地棉是主要的栽培品种,全球90%以上种植的棉花都是陆地棉。但是陆地棉的黄萎病抗性差,并且陆地棉中缺少免疫或者高抗的抗源,这是一直困扰棉花育种家的一大难题。到目前为止,棉花中还没有类病变突变体的报道。我们以异源四倍体陆地棉材料W0为受体,在组织培养的后代中发现了一个类病变突变体Ghlmm。该突变体表现为在植株的叶片局部自发形成黑色病斑,但对棉花的纤维产量和品质没有显著影响。遗传分析表明,Ghlmm是由单基因控制的隐性性状,定位到棉花的D5染色体上。精细定位和图位克隆发现,Ghlmm是由于Gh_D05G2237编码区单核苷酸突变,引起翻译的多肽提前终止,造成功能缺失。Gh_D05G2237编码5-氨基乙酰丙酸脱水酶(ALAD),其功能是将2分子的5-氨基乙酰丙酸(ALA)转换成1分子胆色素原(PBG)。在异源四倍体陆地棉中共有2个编码ALAD基因,是部分同源基因。一个为定位在D亚基因组的Gh_D05G2237,命名为GhLMMD,另外一个为定位在A亚基因组,其部分同源基因Gh_A05G3720,命名为GhLMMA。在异源四倍体棉花中,每个基因含2个拷贝。通过接菌黄萎病抗性鉴定,结果表明,我们发现的Ghlmm突变体具有显著提高的黄萎病抗性。棉花黄萎病危害巨大,治理困难,而占全球种植面积90%以上的陆地棉抗源匮乏。鉴于此,陆地棉突变体Ghlmm的发现及抗病分子机理研究,可以拓宽对棉花黄萎病抗性的认识,通过调控目标基因的表达,实现精确调控棉花生长和抗病之间的平衡,培育抗性显著提高的新种质并在生产上应用,为棉花黄萎病抗性育种提供重要的指导作用。技术实现要素:本发明的目的是提供一种棉花GhLMM基因在提高棉花抗病性及培育棉花新种质中的应用。利用GhLMMA或GhLMMD基因功能缺失突变体与常规种植的棉花品种杂交配置杂交种提高棉花的黄萎病抗性。以此基因为靶基因,通过反义RNA技术等基因工程方法,在一定范围内下调GhLMM基因的表达,培育生长发育正常,且抗性显著提高的棉花新种质并在生产上应用。本发明的另一目的在于提供一种提高棉花抗黄萎病的方法。本发明的又一目的在于提供一种棉花GhLMM基因,并提供该基因在陆地棉遗传标准系TM-1A亚组(GhLMMA)和D亚组(GhLMMD)的全长cDNAORF和基因组序列。另外还提供了上述棉花的GhLMM基因所编码的蛋白。该基因在陆地棉遗传标准系TM-1A亚组和D亚组中编码的氨基酸序列。本发明的目的通过以下技术方案实现:如SEQIDNO.1或SEQIDNO.2所示的GhLMM基因在广谱性提高棉花抗病性及培育棉花新种质中的应用。上述的应用,其在于:利用所述GhLMM基因的功能缺失突变体与常规种植的棉花品种杂交配置杂交种提高棉花的抗病性。上述的应用,其在于:以所述的GhLMM基因为靶基因,通过反义RNA技术等基因工程方法,在一定范围内下调GhLMM基因的表达,培育生长发育正常,且抗性显著提高的新种质并在生产上应用。一种提高棉花抗黄萎病的方法,在棉花中调控核苷酸序列如SEQIDNO.1或SEQIDNO.2所示的GhLMM基因的表达剂量。一个能显著提高棉花抗病性的GhLMM基因,该基因为(1)~(2)中的任意一种:(1)该基因在陆地棉遗传标准系TM-1的基因组A亚组(GhLMMA)中核苷酸序列如SEQIDNO.1所示;(2)该基因在陆地棉遗传标准系TM-1的基因组D亚组(GhLMMD)中核苷酸序列如SEQIDNO.2所示。如(1)所述的基因GhLMMA,该基因具有如SEQIDNO.3所示的ORF序列;如(2)所述的基因GhLMMD,该基因具有如SEQIDNO.4所示的ORF序列。上述的GhLMM基因编码的蛋白,其中基因GhLMMA编码的蛋白具有如SEQIDNO.5所示的氨基酸序列;基因GhLMMD编码的蛋白具有如SEQIDNO.6所示的氨基酸序列。含有上述GhLMM基因的重组载体、表达盒、转基因细胞系或重组菌。上述的重组载体、表达盒、转基因细胞系或重组菌在提高棉花广谱抗病性和培育棉花新种质中的应用。所述的抗病性为黄萎病抗病性。本发明的优点表现在:(1)首次在棉花中发现类病变突变体Ghlmm并克隆其关键基因。到目前为止,棉花中还没有类病变突变体的报道。我们以异源四倍体陆地棉材料W0为受体,在组织培养的后代中发现了一个类病变突变体Ghlmm。遗传分析表明,该突变性状是由单基因控制的隐性性状。图位克隆表明该突变性状是由于异源四倍体棉花D亚组的GhLMMD基因编码区发生SNP突变,导致翻译提前终止,造成基因功能丧失,而A亚组的GhLMMA功能正常。Ghlmm突变体可以完成生长周期,与野生型相比,其纤维品质和产量无显著差异。GhLMM编码5‐氨基乙酰丙酸脱水酶(ALAD),其功能是将2分子的5‐氨基乙酰丙酸(ALA)转换成1分子胆色素原(PBG)。在异源四倍体陆地棉中共有2个编码ALAD基因,是部分同源基因。每个基因含2个拷贝,因此,异源四倍体棉花中编码ALAD的基因共有四个拷贝。表达分析显示,GhLMMA和GhLMMD在不同组织器官中表达水平一致,表明该基因行使功能的剂量效应。由于GhLMMA和GhLMMD核苷酸序列相似为98%,氨基酸序列相似性达99%。使用基因工程基因编辑技术很难创造单个亚组基因功能缺失的突变体。(2)类病变突变体Ghlmm是研究棉花抗病信号通路及分子机制的优异材料,调控GhLMM表达可增强棉花抗病性。黄萎病抗性鉴定发现,类病变突变体Ghlmm具有增强的黄萎病抗性。与正常材料W0相比,突变体中5‐氨基乙酰丙酸(ALA)积累,ROS和SA合成显著增加,PR基因表达显著提高。因此,研究Ghlmm突变体中差异表达基因,可以揭示其中的抗病信号通路和抗病的分子机制。调控GhLMM表达可增强棉花抗病性,为棉花抗病分子育种提供新思路和新基因。(3)研究GhLMM基因的剂量效应,可以实现植物生长和抗性之间的最佳平衡,达到精确调控育种。陆地棉野生型W0具有4个拷贝的GhLMM基因(AtAtDtDt,A亚组和D亚组中各含2个拷贝),突变体具有两个拷贝的GhLMM(AtAtdtdt)基因,用W0和Ghlmm突变体进行杂交可以获得3个拷贝的F1(AtAtDtdt)。在多倍体中可以研究GhLMM的剂量效应,实现植物生长和抗性之间的最佳平衡。通过调控GhLMM基因的表达,控制GhLMM的剂量效应,可以为棉花的抗病育种提供新思路和新基因。以此基因为靶基因,通过反义RNA技术等基因工程方法,以农杆菌侵染的手段得到转基因纯合材料,在一定范围内下调GhLMM基因的表达,实现精确调控棉花生长和抗病之间的平衡,培育抗性显著提高的棉花新种质并在生产上应用。附图说明图1Ghlmm突变体的精细定位和图位克隆a:生长3个月大的突变体和野生型植株表型。Ghlmm突变体在棉花叶片自发形成病斑。b:Ghlmm突变体突变基因的精细定位。c:叶片中表达的12个候选基因在W0和Ghlmm中进一步定量验证。d:Ghlmm突变体中GhLMMD在127bp处核苷酸由G变为T的单突变,导致翻译提前终止。图2在野生型W0中沉默Gh_D05G2254不能产生Ghlmm突变体表型a:在W0中沉默Gh_D05G2254后的表型。b:VIGS沉默后Gh_D05G2254表达的定量验证。图3在W0中沉默GhLMM产生Ghlmm突变体表型a:在W0中沉默GhLMM后的表型b:VIGS沉默GhLMM后表达的定量验证,该基因在A、D亚组的部分同源基因其表达均下调c:VIGS沉默后ALA、过氧化氢、MDA的含量。图4GhLMM基因的组织器官表达、亚细胞定位及其同源基因的系统进化分析a:GhLMM的组织器官表达特征。该基因在A、D亚组的部分同源基因其表达基本一致b:不同物种来源的GhLMM同源基因(ALAD基因)系统进化分析c:GhLMM基因的亚细胞定位。图5GhLMMD基因的功能缺失导致突变体ALA含量升高,ROS含量增加。a:GhLMMA和GhLMMD基因在W0和Ghlmm之间的表达。b-d:W0和Ghlmm的ALAD酶活,ALA、PBG含量比较。e-f:W0和Ghlmm的过氧化氢和MDA含量比较。g:ROS染色。h:过氧化氢染色。图6喷施20mM的LA导致W0表现出Ghlmm突变体表型。a:喷施20mM的LA(ALAD抑制剂)导致W0表现出Ghlmm突变体表型。b:喷施20mM的LA后会造成ALA积累。c:喷施20mM的LA后过氧化氢含量升高。图7Ghlmm突变体的黄萎病抗性鉴定。a:Ghlmm黄萎病抗性鉴定。突变体受体陆地棉材料W0、感病陆地棉品种军棉1号、抗病海岛棉品种H7124作为对照。b:病级调查c:接病16天后第一片真叶的黄萎病相对含量。图8接病16天后劈杆和病菌复苏实验。a:接病16天后子叶结向上1cm茎秆劈杆实验和徒手切片实验。b:接病16天后,第一片真叶和子叶结向上1cm处茎段,土豆培养基培养4天后病菌复苏实验。c:对照不处理茎秆劈杆实验和徒手切片实验。d:对照不处理叶片和茎段病菌复苏实验。图9Ghlmm突变体SA含量升高,PR基因表达升高。a:Ghlmm和W0的SA含量b:PR基因在W0和Ghlmm之间的表达。c:转录组表达差异基因GO富集分析。图10ROS和SA具有循环放大效应ROS诱导SA信号通路基因高表达导致SA含量增加。SA抑制过氧化氢酶酶活,导致ROS含量升高。a:SA信号通路基因EDS1、PAD4、PAL基因受过氧化氢诱导高表达。b:SA信号通路基因EDS1、PAD4、PAL基因在Ghlmm突变体中高表达。c:Ghlmm中CAT酶活受到抑制。d:用2-氨基茚满-2磷酸(AIP)抑制Ghlmm的SA合成后,Ghlmm表型消失,ROS含量减少。e:AIP处理后Ghlmm和W0的SA含量减少。图11抑制Ghlmm突变体中SA合成,PR基因表达降低抑制Ghlmm突变体SA合成后,与未抑制相比,PR基因表达降低。图12PR基因受SA诱导表达PR基因受SA诱导后,表达量显著升高。图13利用不同浓度LA处理,模拟GhLMM基因在细胞死亡和诱导抗病基因表达的剂量效应a:0、1、5、10、20、30mMLA处理W0后叶片表型。b:不同浓度LA处理后ALA、过氧化氢、SA含量。c:不同浓度LA处理后PR基因的表达。图14GhLMM基因在抗黄萎病中的剂量效应配置4拷贝材料W0(AtAtDtDt),2拷贝材料Ghlmm突变体(AtAtdtdt),以及3拷贝材料(W0×lmm)F1进行黄萎病抗性鉴定。a:W0、lmm、(W0×lmm)F1的黄萎病抗性鉴定结果。b:病级调查。图15GhLMM基因剂量效应影响ALA、ROS、SA含量和PR基因的表达a:GhLMM基因在W0、(W0×Ghlmm)F1、Ghlmm中的表达模式。b:ALAD酶活,ALA、PBG的含量。c:过氧化氢、SA含量和CAT酶活。d:抗病基因PR基因的表达。图16Ghlmm突变体用于棉花抗黄萎病育种利用Ghlmm突变体分别与TM-1,军棉1号杂交后,其F1黄萎病抗性显著提高。a:TM-1、(TM-1×Ghlmm)F1、军棉1号、(军棉1号×Ghlmm)F1的黄萎病抗性鉴定。b:病级调查。图17调控GhLMM基因的剂量效应,实现植物生长和抗病的高效平衡a:4拷贝GhLMM基因W0(AtAtDtDt)植株生长正常,但是没有ALA积累,抗病信号通路没有被激活。b:3拷贝GhLMM基因(W0×Ghlmm)F1,ALA积累,抗病信号通路激活,植株表现正常生长和抗病性提高。c:2拷贝ALAD基因Ghlmm突变体,ALA大量积累,但表现叶片细胞死亡,同时ROS、SA含量显著增加,抗病信号通路激活,抗病性显著提高。d:利用VIGS沉默棉花GhLMM基因,植物的四吡咯合成受阻,缺少能量不能正常生长发育,进而植株死亡。具体实施方式(一)棉花类病变突变体Ghlmm的发现,精细定位和图位克隆我们在陆地棉品种W0的组织培养再生苗中,发现一个类病变突变体(Lesionmimicmutant)。在没有任何生物和非生物胁迫条件下,该突变体在叶片上自发形成病斑。我们将其命名为陆地棉类病变突变体Ghlmm。Ghlmm突变体可以完成正常的生长周期,与对照W0相比,其纤维品质和种子产量基本不受影响(表1)。将陆地棉类病变突变体Ghlmm与遗传标准系TM-1杂交,F1植株叶片无病斑,表型为野生型。F1自交产生含763株的F2分离群体。在F2群体的苗期、蕾期、花铃期三个生长发育时期进行叶片表型性状调查,确定单株表现型。结果表明,在F2群体中209株表型为Ghlmm表型,554株为野生型。卡方值为2.202,符合叶片正常和突变3:1分离比,说明Ghlmm突变体是隐性单基因控制的质量性状。通过本实验室构建的高密度遗传连锁图谱,将GhLMM基因定位到D5染色体上标记NAU7928和S2393之间,两个标记的物理距离为371kb。通过TM-1基因组测序结果发现此区间内共有25个开放阅读框,其中,叶片中FPKM值大于1的有12个。通过定量PCR检测发现,只有Gh_D05G2237和Gh_D05G2254在突变体中表达量显著下降(图1)。将两个基因在TM-1,W0和Ghlmm中克隆发现,Gh_D05G2254没有差异。利用VIGS沉默技术将Gh_D05G2254在W0中沉默,并不能产生Ghlmm表型(图2)。而Gh_D05G2237在Ghlmm中127bp处有一单核苷酸突变,由G突变为T,Glu(GGA)突变为终止密码子(TGA),造成了基因的提前终止。进一步利用突变位点设计T能产生扩增产物,而G不能产生扩增产物的SNP引物。用763株F2群体进行扩增,发现所有的209株Ghlmm表型植株都具有扩增产物。利用VIGS沉默技术将Gh_D05G2237在W0中沉默,注射10天之后表现出Ghlmm表型,继续培养植株不能产生新的幼叶继而死亡,qPCR表明该基因在陆地棉A、D亚组的拷贝均被沉默(图3)。综上所述,我们确定Ghlmm突变体其突变表型是由于Gh_D05G2237中的单核苷酸突变导致该基因提前终止,造成功能失活。在陆地棉正常材料中该基因被定名为GhLMM,含2个部分同源基因。一个定位在A亚基因组,命名为GhLMMA(该基因的基因组序列如SEQIDNO.1所示,其cDNAORF序列如SEQIDNO.3所示,该基因编码的蛋白质序列如SEQIDNO.5所示)。一个定位在D亚基因组,命名为GhLMMD(该基因的基因组序列如SEQIDNO.2所示,其cDNAORF序列如SEQIDNO.4所示,其cDNAORF序列如SEQIDNO.6所示)。GhLMM编码5-氨基乙酰丙酸脱水酶(ALAD),其功能是将2分子的5-氨基乙酰丙酸(ALA)转换成1分子胆色素原(PBG)。研究所用引物列于表2。表1:受体W0和类病变突变体Ghlmm的纤维品质和产量比较表2:精细定位、克隆、功能验证目标基因所用引物引物名称引物序列(5'-3')用途S2470FTAAGTATCGTGGCCCATGGA精细定位S2470RTGTAACCCCATTGTGTCGCA精细定位S3183FTCGTGGCCCATGGATTAAACT精细定位S3183RACTGGACGAAGCCTTGACAT精细定位NAU7928FGGGTAAACAAACATTGCATC精细定位NAU7928RTCCATTCATTAGACTGCTACTTCTG精细定位S2393FAATGTGATGTAGCTTAGACCAACTTCCAACCTAC精细定位S2393RGCAAATATCCTGACATGAGAACATATGAATCCA精细定位S3184FACCAACCATGCAATAGTCCC精细定位S3184RGCAGAAGGGTTCGAGAAGCA精细定位S2450FGTTTAAGGTTCAATTCTGGTAGGACTTCAACCTTAT突变位点SNP引物S2450RTTGTCACAAATTCCTATCACATTACCCCCTATTT突变位点SNP引物S2657FTTTAAGGAACGAAAAAGAAATAGAGhLMM的cDNA序列克隆S2657RTCATAGAAGGAAAATCAGACCAAGGhLMM的cDNA序列克隆S2346FTTCATTGTGTACCCATTTCTTGTGhLMM基因组分段克隆-1S2346RCATTGTCATTGTAGGCTTCATCCGhLMM基因组分段克隆-1S2347FCAGGATGTTATAGGCTTGGATGGGhLMM基因组分段克隆-2S2347RGATTTGATAGGCGAAAAGGGATTGhLMM基因组分段克隆-2S2348FAGGTTTTACTTTCTTTTCACTTCCGhLMM基因组分段克隆-3S2348RCATTCTAACAACTAGCACTCTCGAGhLMM基因组分段克隆-3S2454FCAATGACAATGGTTTAGTGCCTGhLMMVIGS载体构建S2454RTTCTCTGTAATTTGCTGGGTTCGhLMMVIGS载体构建S4349FGTCGAGAGGGAATAACTGGGAGh_05D2254的cDNA序列克隆S4349RAGTCCAACCACCGTTGATGTTGh_05D2254的cDNA序列克隆S3649FCGGCCACTTAAATCCCTCCAGh_05D2254VIGS载体构建S3649RCTGAACCAAGCACTCAACCGGh_05D2254VIGS载体构建(2)GhLMM基因的特征及进化分析基于已释放的棉花四倍体TM-1基因组序列数据,以及测序过的物种序列信息。利用PFAM网站http://pfam.janelia.org,下载种子文件(Pfam00490)。我们调查了藻类:衣藻、团藻,蕨类:卷柏,苔藓:小立碗苔藓,裸子植物:挪威云杉,单子叶植物:水稻、谷子、高粱、二穗短柄草双子叶植物:拟南芥、大豆、葡萄、可可、杨树以及三个棉种雷蒙德氏棉、亚洲棉和陆地棉中GhLMM的同源基因(ALAD基因)。利用MEGA软件,进行系统进化树构建。在调查的所有物种中,ALAD基因数量最多的只有2条,不同来源的ALAD基因结构上十分相似。说明ALAD基因进化上十分保守,功能非常重要。通过设计GhLMM亚组专化的定量引物,以TM-1的根、茎、叶以及不同时期的胚珠、纤维的cDNA为模板,用定量PCR技术,对GhLMM基因在不同组织器官表达特征进行研究。GhLMM基因在供试的不同组织器官种均表达,在叶片和胚珠发育早期和纤维中表达量相对较高。设计异源四倍体陆地棉亚组专化引物(表3),进行亚组基因表达分析。GhLMMA和GhLMMD在不同组织器官中表达水平一致,无明显差异。构建GFP4亚细胞定位载体,利用农杆菌注射烟草叶片的方法,利用激光共聚焦显微镜观察GhLMM基因的亚细胞定位,GhLMMA和GhLMMD均定位到叶绿体上(图4)。表3:扩增所用引物引物名称引物序列(5'-3')用途S2622FTGGTGAGAAGAGGTGAACTTGGTCCGGhLMM-Dt定量PCRS2622RAGGCTTATGAAGATGCGGACTACGAACAGGhLMM-Dt定量PCRS2706FTTATCCTTACATATTTTGCTCTACAAGCTGCAAGAGGhLMM-At定量PCRS2706RCTCCTCATCGTTTTAACAACTAGCACTCTCGAGhLMM-At定量PCRS2979FGAACGATAGGGTACCCCCGGGATGGCTTCCACAATTGTAAAGhLMM亚细胞定位S2979RGCCCTTGCTCACCATGGATCCCCTCTTCTCACCACATAAAGGhLMM亚细胞定位(3)类病变突变体的细胞程序性死亡机理的研究GhLMM基因编码5-氨基乙酰丙酸脱水酶(ALAD),是四吡咯化合物合成中的关键酶,其功能是将ALA转换成PBG。我们推测Ghlmm突变体中编码ALAD基因的GhLMM突变会造成ALA的积累。ALA积累过量后,会烯醇化产生ROS。ROS的累积会对植物细胞膜的脂类分子产生氧化胁迫造成植物细胞死亡。以W0和Ghlmm为研究材料,在棉花生长到两叶一心时期取样。检测GhLMM基因的表达,发现W0和Ghlmm中GhLMMA表达并没有差异,但是GhLMMD基因提前终止造成基因功能丧失,其表达量也极显著下降。同时我们检测了ALAD酶活,发现Ghlmm中具有ALAD酶活,但与野生型相比其酶活水平极显著下降。说明GhLMMA具有ALAD酶活活性,在野生型材料中GhLMMA与GhLMMD转录水平相当,GhLMMA不能补偿GhLMMD基因功能缺失。在Ghlmm中,ALAD底物ALA积累,产物PBG含量降低。ALA大量积累造成ROS含量升高,对植物细胞膜造成氧化损伤,MDA含量升高(图5)。乙酰丙酸(LA)是ALAD的抑制剂,我们利用体外喷施LA抑制ALAD活性来模拟Ghlmm突变体GhLMM基因失活。喷施20mM的LA之后W0叶片中产生细胞死亡现象,并且ALA、过氧化氢含量升高。VIGS沉默GhLMM之后ALA、过氧化氢和MDA含量升高(图6)。通过以上实验表明植物体内ALA积累会造成ROS含量的增加,导致植物细胞死亡。综上所述,引起Ghlmm突变体叶片细胞死亡表型的原因是由于ALA积累,ROS爆发对植物细胞膜的氧化损伤导致。(4)类病变突变体Ghlmm的抗病鉴定、抗病机理的研究用突变体Ghlmm和受体W0以及抗病材料H7124和感病材料军棉1号作研究材料,对Ghlmm突变体的黄萎病抗性进行了鉴定。棉花种植16-18天后,植株处于2叶一心时期。用培养的黄萎病菌V991,20mL1×107的悬浮孢子液,撕底伤根的方式进行接病。接病后调查病级。在接病16天后W0和军棉1号叶片表现出黄化萎蔫的表型,而Ghlmm和H7124没有感病表型。提取棉花第一片真叶DNA,用黄萎病菌内标基因引物进行定量验证,棉花的组蛋白3基因作为内标,检测棉花叶片中黄萎病的相对含量,发现Ghlmm和H7124接病16天后,第一片真叶中基本检测不到黄萎病,而W0和军棉一号已有较高含量的黄萎病菌(图7)。茎秆切片实验发现W0和军棉1号维管束变为棕黑色。取植株同一部位的茎段和第一片真叶,置于土豆培养基中培养四天发现,W0和军棉1号茎中和第一片真叶黄萎病菌菌斑明显。而Ghlmm和H7124茎秆切片实验和病菌复苏实验表明,在接病16天后,没有明显病菌侵染表型(图8)。为了进一步揭示Ghlmm突变体抗病的分子机理,我们以W0和Ghlmm为研究材料,在棉花生长到两叶一心时期取样开展研究。SA含量升高是植物系统获得性抗性(SAR)的必要条件,通过检测植株SA含量,发现与对照相比,Ghlmm中SA含量显著升高。SAR的标志是PR基因的高表达,且PR基因是植物抵抗病菌侵染的直接效应基因。在两叶一心时期,取W0和突变体Ghlmm根,茎,叶检测PR基因的表达特征。结果表明,Ghlmm叶片中PR基因表达量极显著升高,与对照相比,PR1和PR5表达量差异百倍以上。在茎和根上,也检测到PR基因的显著高表达。为了在全基因组水平上揭示Ghlmm的抗病机理。我们提取W0和Ghlmm叶片RNA,进行转录组差异表达分析。根据测序结果,筛选表达差异基因,并进行GO富集。结果显示Ghlmm中与胁迫,防卫反应,植物免疫等相关基因被显著富集(图9)。为研究突变体中ROS爆发与SA信号的产生之间的关系,我们设计基因专化引物(表4)检测了W0和Ghlmm的SA信号通路基因表达特征。发现EDS1、PAD4、PAL在Ghlmm中表达量显著升高,并且这些基因受到ROS的诱导,说明ROS可以诱导SA的合成。过氧化氢酶CAT是SA的结合蛋白,SA可以抑制CAT的酶活,导致过氧化氢含量的升高。在Ghlmm中CAT酶活显著下降。在突变体中ROS和SA形成了一个循环放大的效应。AIP可以抑制PAL的活性,抑制SA的合成。为了进一步研究突变体中ROS和SA的关系,我们用2-氨基茚满-2磷酸(AIP)喷施W0和Ghlmm。喷施1mM的AIP之后,W0和Ghlmm的SA含量显著下降,Ghlmm突变体叶片细胞死亡表型消失,并且与喷施前相比,ROS含量显著下降(图10)。同时我们检测了AIP处理后PR基因的表达,发现PR基因表达也显著下降(图11)。由于PR基因受到SA诱导高表达(图12),研究表明Ghlmm中PR基因的表达是由于SA信号通路的激活引起的,且ROS与SA之间的循环放大效应导致细胞死亡和抗病基因的表达。表4:扩增所用引物(5)GhLMM基因在调控植物生长和抗病之间的剂量效应研究Ghlmm突变体叶片细胞死亡表型和抗病性的增加是由于受体W0的GhLMMD基因突变所导致。GhLMMA不能补偿GhLMMD基因功能的丧失,说明二者之间的表型差异和抗病性差异是由于GhLMM基因的拷贝数不同,因此,GhLMM的功能特征具有剂量效应。为了研究GhLMM的剂量效应,我们设计了2个实验。一、外施不同浓度LA抑制ALAD酶活,模拟不同程度的GhLMM表达抑制。在W0生长到两叶一心时期,采用体外喷施的方法,喷施0、1、5、10、20、30mM的LA。我们发现喷施24小时后,20mM和30mM叶片表现出Ghlmm突变体表型,即叶片局部细胞褐化死亡,形成坏死斑。而10mM具有不明显的细胞死亡现象。0、1、5mMLA喷施后,棉花叶片不形成坏死斑。通过对ALA、过氧化氢、SA含量的检测发现,随着LA喷施浓度的增加,ALA、过氧化氢、SA的含量也随着增加。并且PR基因的表达也随着LA浓度的增加,表达量升高(图13)。二、利用遗传学方法,创制不同拷贝GhLMM基因的材料,调查其生长表型和抗病性。用四个GhLMM基因拷贝的W0(AtAtDtDt)和两个GhLMM基因拷贝的Ghlmm(AtAtdtdt),杂交产生三个GhLMM拷贝的(W0×Ghlmm)F1(AtAtDtdt)。对三个材料的黄萎病抗性进行了鉴定。结果显示,(W0×Ghlmm)F1的黄萎病抗性处于W0和Ghlmm之间,接病28天之后其病级达60%,而W0病级超过95%,Ghlmm约为40%。接病28天后的劈杆实验和病菌培养实验同样验证了这一结果(图14)。棉花生长到2叶一心时期,检测三个材料中GhLMM基因的表达,(W0×Ghlmm)F1基因GhLMMD的表达处于三个材料中间。而GhLMMA的表达并没有显著差异。ALAD酶活检测同样得到了类似结果。随着拷贝数的减少,ALA积累增多,PBG含量减少。过氧化氢的含量和SA含量也随着GhLMM的拷贝数的减少而增多。(W0×Ghlmm)F1叶片中PR基因的表达也显著高于W0,低于Ghlmm,处于中间水平,这与其抗病表现一致(图15)。通过外施LA模拟实验和不同拷贝GhLMM基因材料的生长和抗病性鉴定,表明GhLMM在调控植物生长和植物免疫中具有剂量效应。(6)Ghlmm突变体在棉花抗病育种上的应用陆地棉黄萎病抗性育种是困扰棉花育种家的一大难题,主要是因为陆地棉缺少黄萎病抗源。陆地棉突变体Ghlmm具有显著升高的黄萎病抗性,并且其纤维品质和种子产量基本不受影响。与亲本W0杂交产生的(W0×Ghlmm)F1也表现出了黄萎病抗病性的显著提高。所以我们设想利用Ghlmm突变体进行杂交育种,改善已有的黄萎病抗性差的陆地棉品种。我们选择了两个感病品种:陆地棉标准系TM-1和军棉1号,分别与Ghlmm进行杂交产生F1代,检测其黄萎病抗性。通过鉴定发现,与感病亲本相比,(TM-1×Ghlmm)F1、(军棉1号×Ghlmm)F1都表现出了显著提高的黄萎病抗性(图17)。因此,Ghlmm突变体可有效用于黄萎病抗性育种。另外,以此基因为靶基因,通过反义RNA技术等基因工程方法,在一定范围内下调GhLMM基因的表达,实现精确调控棉花生长和抗病之间的平衡,培育抗性显著提高的新种质并在生产上应用。SEQUENCELISTING<110>南京农业大学<120>能显著提高棉花抗病性的GhLMM基因及其应用<130>2017<160>6<170>PatentInversion3.3<210>1<211>4642<212>DNA<213>陆地棉<221>GhLMM在陆地棉遗传标准系TM-1的基因组A亚组中核苷酸序列<400>1tttcattgtgtacccatttcttgttttagctgatttgtgctgttattttagttagaattt60catcttttacgattgcagagatcgtgagaattttgtgtcatggcttccacaattgtaaat120gctccctgcacggttccttctgttaaaggttttgagactcaaaactacgttggtcttaga180ccaatttctagtttaaggttcaattctggtaggacttcaacctcaggacgttctcggggc240ctttttgtggtgagggcatgcgaaaggcatgatggacatgtgaaaaagattgaaatgagt300attgaagaatgtgaggctgctgttgtagctgggaatgcgcctgaagctcctccagttccc360cctaagccagctgctcctgttgggactcctgtgattaaaccttatgtgagtgtttgaaat420gaaatgataatattttatcttttgcccatattttacggtgcttgcttatttatgttcata480tgcaaatagggggtaatgtgataggaatttgtgacaaattgattgtgtttgagtggtttt540agaatgtctccgataagtgtttggtctattgaatggtgatttgttttagacttttatatg600tgtttgaaaattcacattcacaggtttaattttcttgttcgctgacatgaatttgattta660atcattaattgtttgttcatctcatctcatcaattcatgacattgaagtaaaatagtttt720aagttataatagaggttttgaggtcaattggtggacctctcctttctcctttgctcactc780ggtgtttgtttgtattagcaacttaccaggcgtcctcgtcgtaaccgtaagtcccctgca840ttgagagcatcattccaggaaacaagtatatctcctgcaaattttgtatatcctcttttt900attcatgaaggttggtattcatggatcttagtcctttctttttggtgaaataagttagat960attaaaagtggagcaattttaccaaagatcatgataagcgccctcaaatttgttacatga1020atattatattgttgcagttttcttttcttttccttttttgttgcgacttttgtgatactt1080ttctgtttactctctttcttaccttccagtacgtgcttacatctacttggtactgttccc1140ttgacctgcaggtcaggaggacacacccattggagctatgccaggatgttataggcttgg1200atggagacatggacttgttgaagaggtgatcttttgtattatgcatcctttttcataaac1260aatttgcatttgtccctcaaaactagttcccgggtgctgtatgtgttgttttaatgactt1320atctattttatgtaactgtatgatgataaacattctatttgatatcaaagttataaacat1380atacaagcatgttatatacttatttagtgtgtcaactgtgaagttgttttgaagtgtgca1440ttatctattgcaggtggcaaaggcctgggatgttggagttaatagtattgtgctcttccc1500caaagttccagatgctctgaaggttttgatctcccgctataatgctatttgcttcatatg1560ttgatgtatataattggttttttgcactttagtctcccacaggagatgaagcctacaatg1620acaatggtttagtgcctcgaacaattcgtctgcttaaggacaagttccctgaccttgtaa1680gtcaatgaagttcattcattgatttttatattgtttagtgcctccagccttatgaaatca1740tggtgatatcttatcggttttttaggttatctatacggatgttgctttagatccatactc1800ctcagatgggcatgatggtattgtcagagaagatggtaagacaagatatgcttcacacac1860ttttgcccgctaaaagaagagtatcttatttagaagacatgttttcaccttgttgaactg1920ttttttggtttgccacattgaaatacaagttaacttagaagaatgatacatgagctcttt1980cctaaaaaatgtcgtgtgcatactaattggtcaaaataaataatgttcttataaagtcat2040agaaatactaattgcttctttgtttttttaccctgtaggagtcataatgaatgatgagac2100agtgcatcaattatgtaagcaagcggtgtcccaggtaatttttgtagtttttttttttta2160aatatactcttaggcagaagtttactgtcttcataagagttctaaatgccataatggtga2220tgacttactgatgtttgttgtggtcactggtgtctcataaaaaaaaatcagtgaccttgt2280tcgcttttgttaaattcttctctcccctccccttgcttgtcttgatacatatccatggca2340taacttttaataaatactttgaggaactagtgtttcaaatgcttaaagcaagcaaaaact2400tgctgttacagctttaaatggctagtcagtgtgatgcttggagttttctaatcatttgaa2460tggatattggaggtctcctatgctcgtttgtgtcctattcttttttttgctagatttaac2520ttgattggacatgttttaatttttttgggtggtgtattcttaaaggctcgagctggagct2580gatgttattagtcccagtgatatgatggatggacgtgttggagctattcgtgctgccctt2640gattctgaaggctttcaccatgtttctatcatgtcctatacagcaaagtaatttcactat2700ctgcccccttgcattcctatagttgtttgtttgtaggtctaccagtttctaaaagtaggt2760ttgaagcatggtaacagatatatgattatgttaaaccaccgcaaaggttttactttcttt2820tcacttcctgaagaacttttgagaaagattattaagaagtcatgaagagttcaaagagaa2880attgagatgaaaacttcttactaatctgaattttacagccttattttccagctttatata2940cagaaaaaaatttaactataataagaaagaaaattttactattaaaactaaaatctctcc3000tatgaataaagaataaaatagctgtacctagtccactaattttgcagtattccgtccttc3060aaattagtctatcaaatcccttttcgcctatcaaatcttgagaatcttggaataagagtc3120ccaggtttccacacttgcccactatgcaagtagaatggtattattcagttgccttgctcg3180aaggcaatagtctttcttcatgttgttgtctaattgcataagacattatcatttttctta3240tcagttggtcactgaacactttagaaaaataaactatgttgacagaagttattctgagtt3300attcttggcggctgggtgaaatttattgacacagtttgtcattctctttttcttgggata3360acagatatgcaagttcattttatggccccttccgagaagcactggactctaacccacgct3420ttggagataaaaaaacgtatgaattcacttgcctgtactgtattttttgggctttcatgc3480gctctttttggtgcttacatttgattgttaatcttgtttgttaaaaacaggtatcagatg3540aacccagcaaattacagagaagcattagttgaggctcatgaagatgaggctgaaggtgct3600gacattctattggtcagtatgacgcttttaaggaattttgagttgtttttgttaatggct3660aagctcccatcatgctgaagtctctctaactggatcacaggtcaagcctggtctccctta3720tttggatatcattagacttctcagggatcaatctcctttgcctattgctgcctaccaggt3780attctcttaactttccgtgtcctgcattaactaatattcttggcattgttttcattgcaa3840cttgaaacctttcgataataattccccatggtccctggaaatgaaaatgaaaatgaaaat3900ttgaggcaacattcatatatattagtaattgttttgtcagttccagcactaggtccttag3960attcccccttttggaattgatgcccttagcaaaactattcttgacacgaaattatttata4020ttacatttttgttctttttagcgaaatcaggggattggtatttttaactacattgtctga4080ttcattcttgtagagtgtagttcattcttaataggctgaaggatgcaggaagtaaggatg4140gcacgagaatagtagaaaactgagaagggggggaaaatataagtgggttgcttttgcata4200tttcatgaaccaatcattcttgtacgagatactcaagattacgtctttattactactaac4260tattctataattagtggtgtaaaaaaagaaaatatactatttctcccccccccccccgaa4320aaaaaaatgcagactttcactatcctaccgctatcaaaatttgattttttttccaggttt4380caggtgaatactccatgatcaaagccggtagtgttctcaaaatgattgatgaagaaaggg4440tcatgatggaatcattgatgtgcctccgccgggctggtgccgacattatccttacatatt4500ttgctctacaagctgcaagggctttatgtggtgagaagaggtgaacttggtctaattttc4560cttctatgaagctttggttgattctgcaaaaaaggcttgagacactaaatcttccattcg4620agagtgctagttgttagaatga4642<210>2<211>4699<212>DNA<213>陆地棉<221>GhLMM在陆地棉遗传标准系TM-1的基因组D亚组中核苷酸序列<400>2tttcattgtgtacccatttcttgttttagctgatttgtgctgctattttagttagaattt60catcttttacgattgcagagatcgtgagaattttatgtcatggcttccacaattgtaaat120gccccctgcacggttccttctgttaaaggttttgagactcaaaactacgttggtcttaga180ccaatttctagtttaaggttcaattctggtaggacttcaacctcaggacgttctcggggc240ctttttgtggtgagggcatgcgaaaggcatgatggacatgtgaaaaagattgaaatgagt300attgaagaatgtgaggctgctgttgtagctgggaatgcccccgaagctcctccagttccc360cccaagccagctgctcctgttgggactcctgtgattaaaccttatgtgagtgtttgaaat420gaaatgataatattttatcttctgcccatattttacagtgcttgcttattaatgttcata480tgcaaatagggggtaatgtgataggaatttgtgacaaattgattgtgtttgagtggtttt540agaatgtctccggcaagtgtttggtctattgaatggtgatttgttttagacttttatatg600tgtttgaaaattcacattcacaggtttaattttcttgttcgctgacatgaatttgattta660atcagtaattgtttgttcacctcatctcatcaattcatgacattggagtaaaatagtttt720aagttataattgaggttttgaggccaattggtggacctctcctttctcctttgctcactc780ggtgtttgtttgtattagcaacttaccaggcgtcctcgtcgtaaccgtaagtcccctgca840ttgagagcatcattccaggaaacaagtatatctcctgcaaattttgtatatcctcttttt900attcatgaaggttggtattcatggatcttagtcctttctttttggtgaaataagttagat960attaaaggtggagcaattttaccaaagatcatgataagcgccctcaaatttgtaaaatga1020atattatattgttgcagttttcttttcttttccttttttgttgcgacttttgtgatactt1080ttctgtttactctctttcttaccttccagtatgtgcttacatctactttggtactgttcc1140ctcgacctgcaggtcaggaggacacacccattggagctatgccaggatgttataggcttg1200gatggagacatggacttgttgaagaggtgaccttttgcattatgcatcctttttcataaa1260caatttgcatttttccccattatgcatcctttttcataaacaatttgcatttttccctca1320aaactagttcccaggtgctgtatatgttgttttaatgacttatctatttcatgtaactgt1380aaggtgataaacattctatttgatatcgaagttataaacatatacaagcatgttatatac1440ttatttagtgtgtcaactgtgaagttgttttgaagtgtgcattatctattgcaggtggca1500aaggcccgggatgttggagttaatagtattgtgctcttccccaaagttccagatgctctg1560aaggttttgatctcccgctataatgctatttgcttcatatgttgatgtatataattggtt1620ttttgcactttagtctcccacaggggatgaagcctacaatgacaatggtttagtgcctcg1680aacaattcgtctgcttaaggacaaattccctgaccttgtaagtcaatgaagttcattcat1740tgatttttatattgtttagtgcctccagccttatgaaatcatgttgatatcttatcggtt1800ttttaggttatctatacggatgttgctttagatccatactcctcagatgggcatgatggt1860attgtcagagaagatggtaagacaagatatgcttcacacacttttggctgctaaaagaag1920agtatcttatttacaagacatgttttcaccttgttgaactgttttttggtttgccacatt1980gacatacaagttaacttagaagaatgatacatgagctctttcctaaaaaatgtcgtgtgc2040ttactaattggtcaaaataaatagtgttcttataaagtcatagaaatactaattgctact2100ttgtttttttcccctgtaggagtcataatgaatgatgagacagtgcatcaattatgtaag2160caagctgtgtcccaggtaatttttgtagggtttttttttttttaaaatatactctttggc2220agaagtctactgtcttcataagagttctaaatgccataatggtgatgacttactgatgtt2280tgttgtggtcactggtgtctcataaaaaaaatcagtgaccttgtttgcttttgttatatt2340cttctctccccctcccctcgcttgtcttgatacatatccatggcataacttttaataaat2400actttgaggaactagtgtttcaaatgcttaaagcaagcaaaaacttgctgttacagcttt2460aaatggctagtcagtgtgatacttggagttctctaatcatttgaatggatattggaggtc2520tcctatgctcatttgtgtcctattcttttttttgctagctttaacttgattggacatgtt2580ttaaattttttgggtggtgtattcttaaaggcccgagctggagctgatgttgttagcccc2640agtgatatgatggatggacgtgttggagctattcgtgctgcccttgattctgaaggcttt2700caccatgtttctatcatgtcctatacagcaaagtaatttgactatctgcccccttgcatt2760cctatagctgtttgtttgtaggtctaccagtttctaaaagtaggtttgaagcatggtaac2820agatatatgattatgtaaaccatcgcaaaggttttactttcttttcacttcctgaagaac2880ttttgagaaagattattaagaagtcttgaagagttcaaagagaaattgagatgaaaactt2940cttactaatctgaattttacagccttattttccagctttatatacacaaaaaaatttaac3000tataataagaaagaaaatcttactattaaaactaaaatctctcccatgaataaagaataa3060aatagctgtacctaatccactaatttagcagtattccgtccttcaaattagtctatcaaa3120tcccttttcgcctatcaaatctggagaatcttggaataagagccccaggtttccacactt3180gcccactatgcaagtagaatggtattattctgttgccttgttcgaaggcaatagtccttt3240tcatgttgttgtctaattgcataagacattatcgtttttcttattagttggtcactgaac3300aatttagaaaagtaaactatgttgacagaagttattctgagttattcttggcggctgggt3360gagatttattgacacagtttgtcattctctttttcttgggataacagatatgcgagttca3420ttttatggccccttccgagaagcactggactctaacccacgctttggagataaaaaaacg3480tatgaattcacttgcctgtactgtattttatgggctttcatgggctctttttggtgctta3540catttgattgttaatcttgtttgttaaaaacaggtatcagatgaacccagcaaattacag3600agaagcattagttgaggctcatgaagatgaggctgaaggtgctgacattctattggtcag3660tatgacgcttttaaggaattttgagttctttttgttaatggctaagctcccatcatgatg3720aagtctctctaactggatcacaggtcaagcctggtctcccttatttggatatcataagac3780ttctcagggatcaatctcctttgcctattgctgcctaccaggtattctcttaactttttc3840gtgtcctgcattaactaatattcttgtcattgttttcattgcaacttgaaacctttcttt3900gataagaattccctatggtccctggaaatgaaaatgaaaatgaaaatttgaggcaacatt3960catatataaccagtaattgttttgtcagttcctgcactaggtccttggattgcccctttt4020ggaattaatgcccttagcaaaacttttcgtgacacgaaattatttatattacatttttgt4080tctttttagcaaaatcaggggattggtatttttaactacattgtctgatattgtctgatt4140cattcttgtagagtgtagttcatacttaataggttgaaggatgcaggaagtaaggatggc4200atgagaatagtagaaaactaagaaggggtgggggaaatataagcgggttgcttttgcata4260tttcatgaaccaatcattcttgtacgagatactcaagattacgtctttattactactaac4320tgttctataattagtggtggaaaaaaagaaaatatactatttctcccccccccccaaaaa4380aaatgcagactttcactttcctacccctatcaaaatttgatttttttttccaggtttcag4440gtgaatactccatgatcaaagctggtggtgttctcaaaatgattgatgaagaaagggtca4500tgatggaatccttgatgtgtctccgccgagctggtgccgacattatccttacatattttg4560ctctacaagctgcaaggtctttatgtggtgagaagaggtgaacttggtctgattttcctc4620ctatgaagctttggttgattctgcaaaaaaggcttgagacactaaatcttccattcgaga4680gtgctagttgttagaatga4699<210>3<211>1293<212>DNA<213>人工序列<221>GhLMM在陆地棉遗传标准系TM-1的基因组A亚组中cDNAORF序列<400>3atggcttccacaattgtaaatgctccctgcacggttccttctgttaaaggttttgagact60caaaactacgttggtcttagaccaatttctagtttaaggttcaattctggtaggacttca120acctcaggacgttctcggggcctttttgtggtgagggcatgcgaaaggcatgatggacat180gtgaaaaagattgaaatgagtattgaagaatgtgaggctgctgttgtagctgggaatgcg240cctgaagctcctccagttccccctaagccagctgctcctgttgggactcctgtgattaaa300ccttatcaacttaccaggcgtcctcgtcgtaaccgtaagtcccctgcattgagagcatca360ttccaggaaacaagtatatctcctgcaaattttgtatatcctctttttattcatgaaggt420caggaggacacacccattggagctatgccaggatgttataggcttggatggagacatgga480cttgttgaagaggtggcaaaggcctgggatgttggagttaatagtattgtgctcttcccc540aaagttccagatgctctgaagtctcccacaggagatgaagcctacaatgacaatggttta600gtgcctcgaacaattcgtctgcttaaggacaagttccctgaccttgttatctatacggat660gttgctttagatccatactcctcagatgggcatgatggtattgtcagagaagatggagtc720ataatgaatgatgagacagtgcatcaattatgtaagcaagcggtgtcccaggctcgagct780ggagctgatgttattagtcccagtgatatgatggatggacgtgttggagctattcgtgct840gcccttgattctgaaggctttcaccatgtttctatcatgtcctatacagcaaaatatgca900agttcattttatggccccttccgagaagcactggactctaacccacgctttggagataaa960aaaacgtatcagatgaacccagcaaattacagagaagcattagttgaggctcatgaagat1020gaggctgaaggtgctgacattctattggtcaagcctggtctcccttatttggatatcatt1080agacttctcagggatcaatctcctttgcctattgctgcctaccaggtttcaggtgaatac1140tccatgatcaaagccggtagtgttctcaaaatgattgatgaagaaagggtcatgatggaa1200tcattgatgtgcctccgccgggctggtgccgacattatccttacatattttgctctacaa1260gctgcaagggctttatgtggtgagaagaggtga1293<210>4<211>1293<212>DNA<213>人工序列<221>GhLMM在陆地棉遗传标准系TM-1的基因组D亚组中cDNAORF序列<400>4atggcttccacaattgtaaatgccccctgcacggttccttctgttaaaggttttgagact60caaaactacgttggtcttagaccaatttctagtttaaggttcaattctggtaggacttca120acctcaggacgttctcggggcctttttgtggtgagggcatgcgaaaggcatgatggacat180gtgaaaaagattgaaatgagtattgaagaatgtgaggctgctgttgtagctgggaatgcc240cccgaagctcctccagttccccccaagccagctgctcctgttgggactcctgtgattaaa300ccttatcaacttaccaggcgtcctcgtcgtaaccgtaagtcccctgcattgagagcatca360ttccaggaaacaagtatatctcctgcaaattttgtatatcctctttttattcatgaaggt420caggaggacacacccattggagctatgccaggatgttataggcttggatggagacatgga480cttgttgaagaggtggcaaaggcccgggatgttggagttaatagtattgtgctcttcccc540aaagttccagatgctctgaagtctcccacaggggatgaagcctacaatgacaatggttta600gtgcctcgaacaattcgtctgcttaaggacaaattccctgaccttgttatctatacggat660gttgctttagatccatactcctcagatgggcatgatggtattgtcagagaagatggagtc720ataatgaatgatgagacagtgcatcaattatgtaagcaagctgtgtcccaggcccgagct780ggagctgatgttgttagccccagtgatatgatggatggacgtgttggagctattcgtgct840gcccttgattctgaaggctttcaccatgtttctatcatgtcctatacagcaaaatatgcg900agttcattttatggccccttccgagaagcactggactctaacccacgctttggagataaa960aaaacgtatcagatgaacccagcaaattacagagaagcattagttgaggctcatgaagat1020gaggctgaaggtgctgacattctattggtcaagcctggtctcccttatttggatatcata1080agacttctcagggatcaatctcctttgcctattgctgcctaccaggtttcaggtgaatac1140tccatgatcaaagctggtggtgttctcaaaatgattgatgaagaaagggtcatgatggaa1200tccttgatgtgtctccgccgagctggtgccgacattatccttacatattttgctctacaa1260gctgcaaggtctttatgtggtgagaagaggtga1293<210>5<211>430<212>PRT<213>人工序列<221>GhLMM在陆地棉遗传标准系TM-1的基因组A亚组中氨基酸序列<400>5MetAlaSerThrIleValAsnAlaProCysThrValProSerValLys151015GlyPheGluThrGlnAsnTyrValGlyLeuArgProIleSerSerLeu202530ArgPheAsnSerGlyArgThrSerThrSerGlyArgSerArgGlyLeu354045PheValValArgAlaCysGluArgHisAspGlyHisValLysLysIle505560GluMetSerIleGluGluCysGluAlaAlaValValAlaGlyAsnAla65707580ProGluAlaProProValProProLysProAlaAlaProValGlyThr859095ProValIleLysProTyrGlnLeuThrArgArgProArgArgAsnArg100105110LysSerProAlaLeuArgAlaSerPheGlnGluThrSerIleSerPro115120125AlaAsnPheValTyrProLeuPheIleHisGluGlyGlnGluAspThr130135140ProIleGlyAlaMetProGlyCysTyrArgLeuGlyTrpArgHisGly145150155160LeuValGluGluValAlaLysAlaTrpAspValGlyValAsnSerIle165170175ValLeuPheProLysValProAspAlaLeuLysSerProThrGlyAsp180185190GluAlaTyrAsnAspAsnGlyLeuValProArgThrIleArgLeuLeu195200205LysAspLysPheProAspLeuValIleTyrThrAspValAlaLeuAsp210215220ProTyrSerSerAspGlyHisAspGlyIleValArgGluAspGlyVal225230235240IleMetAsnAspGluThrValHisGlnLeuCysLysGlnAlaValSer245250255GlnAlaArgAlaGlyAlaAspValIleSerProSerAspMetMetAsp260265270GlyArgValGlyAlaIleArgAlaAlaLeuAspSerGluGlyPheHis275280285HisValSerIleMetSerTyrThrAlaLysTyrAlaSerSerPheTyr290295300GlyProPheArgGluAlaLeuAspSerAsnProArgPheGlyAspLys305310315320LysThrTyrGlnMetAsnProAlaAsnTyrArgGluAlaLeuValGlu325330335AlaHisGluAspGluAlaGluGlyAlaAspIleLeuLeuValLysPro340345350GlyLeuProTyrLeuAspIleIleArgLeuLeuArgAspGlnSerPro355360365LeuProIleAlaAlaTyrGlnValSerGlyGluTyrSerMetIleLys370375380AlaGlySerValLeuLysMetIleAspGluGluArgValMetMetGlu385390395400SerLeuMetCysLeuArgArgAlaGlyAlaAspIleIleLeuThrTyr405410415PheAlaLeuGlnAlaAlaArgAlaLeuCysGlyGluLysArg420425430<210>6<211>430<212>PRT<213>人工序列<221>GhLMM在陆地棉遗传标准系TM-1的基因组D亚组中氨基酸序列<400>6MetAlaSerThrIleValAsnAlaProCysThrValProSerValLys151015GlyPheGluThrGlnAsnTyrValGlyLeuArgProIleSerSerLeu202530ArgPheAsnSerGlyArgThrSerThrSerGlyArgSerArgGlyLeu354045PheValValArgAlaCysGluArgHisAspGlyHisValLysLysIle505560GluMetSerIleGluGluCysGluAlaAlaValValAlaGlyAsnAla65707580ProGluAlaProProValProProLysProAlaAlaProValGlyThr859095ProValIleLysProTyrGlnLeuThrArgArgProArgArgAsnArg100105110LysSerProAlaLeuArgAlaSerPheGlnGluThrSerIleSerPro115120125AlaAsnPheValTyrProLeuPheIleHisGluGlyGlnGluAspThr130135140ProIleGlyAlaMetProGlyCysTyrArgLeuGlyTrpArgHisGly145150155160LeuValGluGluValAlaLysAlaArgAspValGlyValAsnSerIle165170175ValLeuPheProLysValProAspAlaLeuLysSerProThrGlyAsp180185190GluAlaTyrAsnAspAsnGlyLeuValProArgThrIleArgLeuLeu195200205LysAspLysPheProAspLeuValIleTyrThrAspValAlaLeuAsp210215220ProTyrSerSerAspGlyHisAspGlyIleValArgGluAspGlyVal225230235240IleMetAsnAspGluThrValHisGlnLeuCysLysGlnAlaValSer245250255GlnAlaArgAlaGlyAlaAspValValSerProSerAspMetMetAsp260265270GlyArgValGlyAlaIleArgAlaAlaLeuAspSerGluGlyPheHis275280285HisValSerIleMetSerTyrThrAlaLysTyrAlaSerSerPheTyr290295300GlyProPheArgGluAlaLeuAspSerAsnProArgPheGlyAspLys305310315320LysThrTyrGlnMetAsnProAlaAsnTyrArgGluAlaLeuValGlu325330335AlaHisGluAspGluAlaGluGlyAlaAspIleLeuLeuValLysPro340345350GlyLeuProTyrLeuAspIleIleArgLeuLeuArgAspGlnSerPro355360365LeuProIleAlaAlaTyrGlnValSerGlyGluTyrSerMetIleLys370375380AlaGlyGlyValLeuLysMetIleAspGluGluArgValMetMetGlu385390395400SerLeuMetCysLeuArgArgAlaGlyAlaAspIleIleLeuThrTyr405410415PheAlaLeuGlnAlaAlaArgSerLeuCysGlyGluLysArg420425430当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1