一种双靶向嵌合抗原受体及其用途的制作方法
本发明属于嵌合抗原受体领域,具体涉及双靶向cd19和cd20的嵌合抗原受体及其用途。
背景技术:
嵌合抗原受体(chimericantigenreceptor-tcell,car-t)t细胞是指经基因修饰后,能以mhc非限制性方式识别特定目的抗原,并且持续活化扩增的t细胞。2012年国际细胞治疗协会年会中指出生物免疫细胞治疗已经成为手术、放疗、化疗外的第四种治疗肿瘤的手段,并将成为未来肿瘤治疗必选手段。car-t细胞回输治疗是当前肿瘤治疗中最明确有效的免疫治疗形式。大量研究表明,car-t细胞可以有效的识别肿瘤抗原,引起特异性的抗肿瘤免疫应答,显著改善患者的生存状况。
嵌合抗原受体(car)是car-t的核心部件,赋予t细胞hla非依赖的方式识别肿瘤抗原的能力,这使得经过car改造的t细胞相较于天然t细胞表面受体tcr能够识别更广泛的目标。car的基础设计中包括一个肿瘤相关抗原(tumor-associatedantigen,taa)结合区(通常来源于单克隆抗体抗原结合区域的scfv段),一个胞外铰链区,一个跨膜区和一个胞内信号区。目标抗原的选择对于car的特异性、有效性以及基因改造t细胞自身的安全性来讲都是关键的决定因素。
cd19是一种b细胞表面的95kda的糖蛋白,从b细胞发育的早期即开始表达,直至其分化为浆细胞。cd19是免疫球蛋白(ig)超家族的成员之一,作为b细胞表面信号转导复合物的组成元素之一,参与调控了b细胞受体的信号转导过程。在cd19缺陷的小鼠模型中,外周淋巴组织中b细胞的数量会出现明显的减少,对疫苗和丝裂原的应答也会下降,同时伴有血清ig水平的减低。通常认为,cd19的表达只限于b细胞系(b-celllineage),而不表达于多能造血干细胞表面。cd19还表达于大多数b细胞淋巴瘤、套细胞淋巴瘤、alls、clls、多毛细胞白血病,和一部分急性髓性白血病细胞的表面。因此,在对白血病/淋巴瘤的治疗中,cd19是一种非常有价值的免疫治疗靶点。重要的是,cd19不会表达于除b细胞外的大多数正常细胞表面,包括多能造血干细胞,这一特征使cd19可以作为一种安全的治疗靶点,可将患者发生自身免疫性疾病或不可逆性骨髓毒性损伤的风险降至最低。当前,已经研制出了抗cd19的抗体或scfv片段,并且在小鼠模型和人类/灵长类动物中证明了其应用的前景。
近年来,cd19cart细胞领域竞争激烈,一些大的制药公司也和研究机构建立了合作关系。接受表达cd28或4-1bb的cd19cart细胞治疗后,小儿和成人复发或难治性急性b细胞淋巴瘤具有大约90%的完全缓解率。最近,cd19cart细胞治疗在弥漫性大b细胞淋巴瘤,滤泡淋巴瘤或慢性淋巴瘤中具有50%-100%的总体缓解率。cd19cart细胞治疗多发性骨髓瘤病人中具有临床优势,由于终末分化的浆细胞不表达cd19,恶性b细胞前体持续产生恶性浆细胞。
b淋巴细胞抗原cd20是人类b淋巴细胞表面上所特有的标识,主要由297个氨基酸构成。根据不同的磷酸化水平,cd20的分子量为33-37kd,属于非糖基化磷蛋白。cd20分子是一种有四个跨膜区的跨膜蛋白,它的n端和c端都位于细胞质内侧。cd20分子的主要抗原表位位于第三、四跨膜区之间,是一个由43个氨基酸组成的环状区域。从前体b细胞到后来的分化过程,cd20广泛表达于b细胞的表面,但是不表达于晚期分化的浆细胞中。cd20表达于b细胞发展的除了最初和最后以外的所有阶段。cd20在b细胞淋巴瘤、毛细胞白血病、b细胞慢性淋巴细胞白血病和黑色素瘤肿瘤干细胞中均被发现。cd20分子位于b细胞膜表面,抗体接近的空间位阻较小,因此cd20分子成为治疗b细胞淋巴瘤的理想靶点。虽然cd20的功能尚未完全清楚,但是它在ca2+跨膜传递,维持细胞内ca2+聚集和b细胞激活方面发挥着重要作用。cd20是一种在成熟b细胞表面高表达的蛋白,同时也在95%以上的b细胞性淋巴瘤细胞表面高表达,这个特性使之成为靶向单克隆治疗的重要靶点。事实上,抗cd20抗体已经被广泛用来治疗b细胞性淋巴瘤。抗体与cd20结合以后,通过引起细胞凋亡以及抗体依赖的细胞介导的细胞毒作用(antibodydependentcellmediatedcytotoxicity,adcc)、补体依赖的细胞毒作用(complementdependentcytotoxicity,cdc)发挥杀伤作用。但是,抗cd20抗体常因温和的杀伤能力在治疗cd20阳性b细胞淋巴瘤上存在缺陷:只对部分病人有治疗效果,对一些cd20阳性病人起初就无效或者很快使其产生耐药;治愈率低;躲避抗体杀死的肿瘤细胞很快造成病情复发。非霍奇金淋巴瘤(nhl)是最常见的淋巴系统恶性肿瘤,好发于青壮年,其中绝大多数为b细胞来源,约占85%。cd20分子在95%以上的b细胞性nhl中均有表达,且抗原分子在膜上比较暴露,容易接近,与单抗结合后无显著内化和脱落,也不会因与抗体的结合而发生抗原调变,因此成为治疗b细胞淋巴瘤的理想靶点。目前已有人鼠嵌合抗cd20抗体上市(rituximab-c2b8)。尽管rituximab在临床治疗中已显示出较好的疗效,仍有部分的患者对rituximab的治疗不产生反应,且单独使用该药治愈率仅为10%。
美国国家癌症研究所(nci)在用cd19car-t细胞治疗20例复发难治性all虽取得显著成果,完全缓解(cr)率达70%,但仍有2例患者分别在3个月和5个月之后复发,并且cd19转为阴性。maude等报道30例儿童及成人all采用cd19car-t治疗后,27例患者获得cr,其中7例在cd19car-t治疗后6周至8.5个月期间复发:4例cd19阳性,3例cd19阴性。grupp等报道2例all患者在经过cd19car-t输注治疗后均在1个月内获得cr。其中1例患儿持续缓解,另1例患儿2个月后复发,且复发后肿瘤细胞cd19为阴性。在2013美国血液学会(ash)年会上,费城儿童医院(chop)报道17例患者cd19car-t治疗后,14例(82%)在1个月内实现cr,其中cr患者中有3个复发:2例cd19阳性,1例cd19阴性。目前研究表明,cd19car-t细胞治疗复发难治性b-all后复发存在两种模式:(1)流式细胞技术仍可检测到b系标志物cd19,即cd19阳性白血病复发;(2)流式细胞技术不能检测到b系标志物cd19,即cd19阴性白血病复发。复发的原因可能包括car-t细胞在部分患者体内的持续时间短,以及肿瘤抗原表达逃逸变异等。临床研究发现患者外周血的car-t细胞减少或者消失,随即出现白血病的复发,往往研究者为防止这部分患者复发,再次追加注射cd19car-t,但维持白血病缓解的效果不一。
为了防止cd19car-t治疗后cd19阴性复发,双特异性抗原靶点的car-t也许是一个治疗选择。同时采用2个b细胞特异性抗原的car,或者一个car具有2个特异性的靶点,如cd19/cd20双特异性的car。
car-t细胞的一大优点是它们是活性药物,一旦输入,生理机制会调控t细胞的平衡、记忆形成和抗原驱动的扩增。然而,这种治疗尚未完善,t细胞会脱靶而攻击其他的组织,或扩增量过高,超出治疗所需。鉴于car-t细胞已被纳入标准治疗范围,设计病人或药物可控的启动或关闭机制来调控car-t细胞的存在是非常有用的。由于技术原因,关闭机制更易应用于t细胞。作为其中之一,icas9系统正在临床研究当中。细胞在表达icas9时,使用小分子化合物可诱导icas9前体分子形成二聚体,激活凋亡途径,从而实现清除细胞的目的。在移植物抗宿主病中,小分子ap1903已被用于诱导icas9二聚体和清除t细胞,表明了这种方法的可行性(clincancerres.2016apr15;22(8):1875-84.)。
另外,还可利用临床上已经使用的清除性抗体,使car-t细胞同时表达这些抗体针对的蛋白,如tegfr,在治疗相关的毒性反应产生或是治疗已经完成后,通过给予抗体药物清除相应的car-t细胞(scitranslmed.2015;7:275ra22.)。基于安全性的考虑我们的car-t细胞引入了安全开关即tegfr,构建好的cd19-tegfr可实时安全的控制其在体内的表达。我们专利是以cd19的scfv的重链和轻链作为car的结构,同时也引入了tegfr结构。tegfr缺乏细胞外的n末端配体结合结构域和胞内受体酪氨酸激酶活性,但保留了天然氨基酸序列,属于i型跨膜细胞表面定位,其空间构象可与药物级别的抗egfr单克隆抗体西妥昔单抗紧密结合(blood.2011aug4;118(5):1255-63.)。tegfr的主要功能:可以作为细胞表面的标记,同时也适合t细胞的体内追踪可通过流式和免疫组化检测;也可以在体内被妥西单抗(cetuximab)清除。
本发明专利采用双靶向cd19和cd20的car元件同时引入了tegfr结构既可以在使car-t细胞体内进行很好的被示踪,更重要的是此结构可以作为car-t细胞的安全开关:即不想其发挥作用时可加入妥西单抗,安全有效的控制输注的针对cd19和cd20靶点的car-t细胞在体内发挥作用。为临床实验和临床治疗奠定良好的基础。
技术实现要素:
本发明第一方面提供一种多核苷酸序列,所述多核苷酸序列选自:
(1)含有依次连接的抗cd20和抗cd19单链抗体的编码序列、人igg4铰链区的编码序列、人cd28跨膜区的编码序列、人41bb胞内区的编码序列、人cd3ζ胞内区的编码序列和任选的egfr的含胞外结构域iii和胞外结构域iv的片段的编码序列的多核苷酸序列;和
(2)(1)所述多核苷酸序列的互补序列。
在一个或多个实施方案中,所述多核苷酸序列在所述抗cd20单链抗体的编码序列前还含有信号肽的编码序列。在一个或多个实施方案中,所述信号肽的氨基酸序列如seqidno:2第1-20位氨基酸所示。在一个或多个实施方案中,所述抗cd20单链抗体的轻链可变区的氨基酸序列如seqidno:2第21-126位氨基酸所示。在一个或多个实施方案中,所述抗cd20单链抗体的重链可变区的氨基酸序列如seqidno:2第144-263位氨基酸所示。在一个或多个实施方案中,所述抗cd19单链抗体的重链可变区的氨基酸序列如seqidno:2第287-406位氨基酸所示。在一个或多个实施方案中,所述抗cd19轻链抗体的重链可变区的氨基酸序列如seqidno:2第425-531位氨基酸所示。在一个或多个实施方案中,所述人igg4铰链区的氨基酸序列如seqidno:2第532-543位氨基酸所示。在一个或多个实施方案中,所述人cd28跨膜区的氨基酸序列如seqidno:2第545-571位氨基酸所示。在一个或多个实施方案中,所述人41bb胞内区的氨基酸序列如seqidno:2第572-613位氨基酸所示。在一个或多个实施方案中,所述人cd3ζ胞内区的氨基酸序列如seqidno:2第614-725位氨基酸所示。在一个或多个实施方案中,所述egfr的片段含有egfr的胞外结构域iii、胞外结构域iv以及跨膜区,或由egfr的胞外结构域iii、胞外结构域iv以及跨膜区组成。在一个或多个实施方案中,所述egfr的片段含有人egfr的第310-646位氨基酸序列,或由人egfr的第310-646位氨基酸序列组成。如seqidno:2第774-1108位氨基酸所示。
在一个或多个实施方案中,在所述抗cd20单链抗体的编码序列前的所述信号肽的编码序列如seqidno:1第1-60位核苷酸序列所示。在一个或多个实施方案中,所述抗cd20单链抗体的轻链可变区的编码序列如seqidno:1第61-378位核苷酸序列所示。在一个或多个实施方案中,所述抗cd20单链抗体的重链可变区的编码序列如seqidno:1第433-798位核苷酸序列所示。在一个或多个实施方案中,所述抗cd19单链抗体的重链可变区的编码序列如seqidno:1第859-1218位核苷酸序列所示。在一个或多个实施方案中,所述抗cd19单链抗体的轻链可变区的编码序列如seqidno:1第1273-1593位核苷酸序列所示。在一个或多个实施方案中,所述人igg4铰链区的编码序列如seqidno:1第1594-1629位核苷酸序列所示。在一个或多个实施方案中,所述人cd28跨膜区的编码序列如seqidno:1第1633-1713位核苷酸序列所示。在一个或多个实施方案中,所述人41bb胞内区的编码序列如seqidno:1第1714-1839位核苷酸序列所示。在一个或多个实施方案中,所述人cd3ζ胞内区的编码序列如seqidno:1第1840-2175位核苷酸序列所示。在一个或多个实施方案中,所述egfr的片段的编码序列如seqidno:1第2320-3324位核苷酸序列所示。
本发明第二方面提供一种融合蛋白,所述融合蛋白选自:
(1)含有依次连接的抗cd20单链抗体和抗cd19单链抗体、人igg4铰链区、人cd28跨膜区、人41bb胞内区和人cd3ζ胞内区的融合蛋白和任选的egfr的含胞外结构域iii和胞外结构域iv的片段的编码序列;和
(2)在(1)限定的氨基酸序列中经过取代、缺失或添加一个或几个氨基酸且保留活化t细胞活性的由(1)衍生的融合蛋白;
优选地,所述抗cd19单链抗体为抗cd19单克隆抗体fmc63;
优选地,所述抗cd20单链抗体为抗cd20单克隆抗体leu-16。
本发明第三方面提供一种核酸构建物,所述核酸构建物含有本文所述的多核苷酸序列。
在一个或多个实施方案中,所述核酸构建物为载体。在一个或多个实施方案中,所述核酸构建物为逆转录病毒载体,含有复制起始位点,3’ltr,5’ltr,本文所述的多核苷酸序列,以及任选的可选择的标记。
本发明第四方面提供一种逆转录病毒,所述逆转录病毒含有本文所述的核酸构建物,优选含有所述载体,更优选含有所述逆转录病毒载体。
本发明第五方面提供一种基因修饰的t细胞,所述细胞含有本文所述的多核苷酸序列,或含有本文所述的核酸构建物,或感染了本文所述的逆转录病毒,或稳定表达本文所述的融合蛋白和任选的egfr的含胞外结构域iii、胞外结构域iv和任选的跨膜区的片段。
本发明第六方面提供一种含本文所述的基因修饰的t细胞的药物组合物。
本发明第七方面提供本文所述的多核苷酸序列、融合蛋白、核酸构建物或逆转录病毒在制备活化的t细胞中的应用。
本发明第八方面提供本文所述的多核苷酸序列、融合蛋白、核酸构建物、逆转录病毒、或基因修饰的t细胞或其药物组合物在制备治疗cd19和cd20介导的疾病的药物中的用途。
在一个或多个实施方案中,cd19和cd20介导的疾病为白血病、淋巴瘤。
附图说明
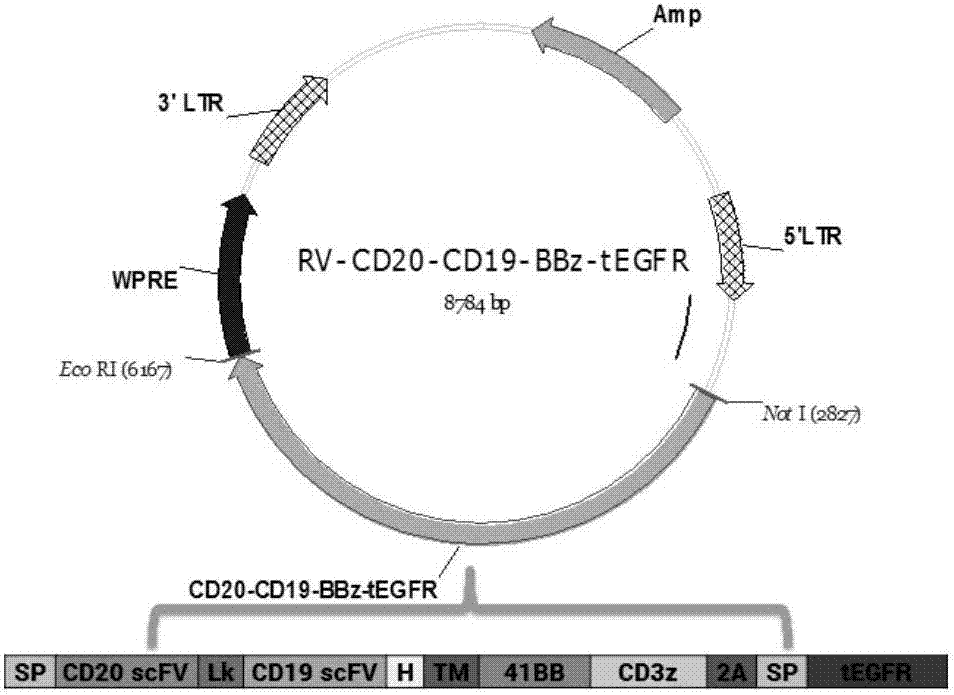
图1为rv-cd20-cd19-bbz-tegfr逆转录病毒表达载体示意图。sp:信号肽;vl:轻链可变区;lk:接头(g4s)3;vh:重链可变区;h:igg4铰链区;tm:cd28跨膜区。
图2为rv-cd20-cd19-bbz-tegfr逆转录病毒表达质粒的部分测序结果峰值图。
图3为流式细胞仪检测逆转录病毒感染t细胞72小时cd20-cd19-bbz-tegfrcart表达效率。。
图4为流式细胞仪检测各靶细胞表面cd19和cd20表达。
图5为制备5天的cd20-cd19-tegfrcart细胞与各靶细胞共培养5小时cd107a表达。
图6为制备5天的cd20-cd19-tegfrcart细胞与各靶细胞共培养5小时inf-γ的分泌。
图7为制备5天的cd20-cd19-tegfrcart细胞与各靶细胞共培养5小时后对肿瘤细胞的杀伤作用。
具体实施方式
本发明提供一种双靶向cd19和cd20的嵌合抗原受体(car)。该car含有依次连接的抗cd20单链抗体和抗cd19单链抗体、人igg4铰链区、人cd28跨膜区、人41bb胞内区、人cd3ζ胞内区和任选的egfr的含胞外结构域iii和胞外结构域iv的片段。
适用于本发明的抗cd20单链抗体可衍生自本领域周知的各种抗cd20单克隆抗体。
适用于本发明的抗cd19单链抗体可衍生自本领域周知的各种抗cd19单克隆抗体。
任选地,所述轻链可变区和重链可变区可通过接头序列连接在一起。可举例的这类单链cd19抗体包括但不限于fmc63,sj25c1。在某些实施方案中,所述单克隆抗体是克隆号为fmc63的单克隆抗体。在某些实施方案中,所述抗cd20单链抗体的轻链可变区的氨基酸序列如seqidno:2的第21-126位氨基酸残基所示。在其它实施方案中,所述抗cd20单链抗体的重链可变区的氨基酸序列如seqidno:2的第144-263位氨基酸残基所示。在某些实施方案中,所述抗cd19单链抗体的重链可变区的氨基酸序列如seqidno:2的第287-406位氨基酸残基所示。在其它实施方案中,所述抗cd19单链抗体的轻链可变区的氨基酸序列如seqidno:2的第425-531位氨基酸残基所示。
适用于本发明的人igg4铰链区的氨基酸序列可如seqidno:2第532-543位氨基酸所示。
适用于本发明的人cd28跨膜区可以是本领域常用于car的各种人cd28跨膜区序列。在某些实施方案中,所述人cd28跨膜区的氨基酸序列如seqidno:2第545-571位氨基酸所示。
适用于本发明的41bb可以是本领域已知的各种用于car的41bb。作为示范性例子,本发明使用seqidno:2第572-613位氨基酸序列所示的41bb。
适用于本发明的人cd3ζ胞内区可以是本领域常规用于car的各种人cd3ζ胞内区。在某些实施方案中,所述人cd3ζ胞内区的氨基酸序列如seqidno:2第614-725位氨基酸所示。
形成本发明的融合蛋白的上述各部分,如抗cd19和cd20单链抗体的轻链可变区和重链可变区、人igg4铰链区、人cd28跨膜区、41bb和人cd3ζ胞内区等,相互之间可直接连接,或者可通过接头序列连接。接头序列可以是本领域周知的适用于抗体的接头序列,例如含g和s的接头序列。通常,接头含有一个或多个前后重复的基序。例如,该基序可以是gggs、ggggs、ssssg、gsgsa和ggsgg。优选地,该基序在接头序列中是相邻的,在重复之间没有插入氨基酸残基。接头序列可以包含1、2、3、4或5个重复基序组成。接头的长度可以是3~25个氨基酸残基,例如3~15、5~15、10~20个氨基酸残基。在某些实施方案中,接头序列是多甘氨酸接头序列。接头序列中甘氨酸的数量无特别限制,通常为2~20个,例如2~15、2~10、2~8个。除甘氨酸和丝氨酸来,接头中还可含有其它已知的氨基酸残基,例如丙氨酸(a)、亮氨酸(l)、苏氨酸(t)、谷氨酸(e)、苯丙氨酸(f)、精氨酸(r)、谷氨酰胺(q)等。作为例子,接头可由seqidno:7-18中任一氨基酸序列组成。在某些实施方案中,本发明抗cd19单链抗体的轻链可变区和重链可变区之间由(ggggs)n连接,其中n为1~5的整数。
在某些实施方案中,本发明的car的氨基酸系列中还包含如下文所述的egfr的含胞外结构域iii和胞外结构域iv的片段,其信号肽以及接头序列。
应理解,在基因克隆操作中,常常需要设计合适的酶切位点,这势必在所表达的氨基酸序列末端引入了一个或多个不相干的残基,而这并不影响目的序列的活性。为了构建融合蛋白、促进重组蛋白的表达、获得自动分泌到宿主细胞外的重组蛋白、或利于重组蛋白的纯化,常常需要将一些氨基酸添加至重组蛋白的n-末端、c-末端或该蛋白内的其它合适区域内,例如,包括但不限于,适合的接头肽、信号肽、前导肽、末端延伸等。因此,本发明的融合蛋白(即所述car)的氨基端或羧基端还可含有一个或多个多肽片段,作为蛋白标签。任何合适的标签都可以用于本文。例如,所述的标签可以是flag,ha,ha1,c-myc,poly-his,poly-arg,strep-tagii,au1,ee,t7,4a6,ε,b,ge以及ty1。这些标签可用于对蛋白进行纯化。
本发明也包括seqidno:2第21-613位氨基酸序列所示的car、seqidno:2第21-613位氨基酸序列所示的car、seqidno:2第1-613位氨基酸序列所示的car或seqidno:2所示的car的突变体。这些突变体包括:与该car具有至少80%,优选至少85%,优选至少90%,优选至少95%,优选至少97%的序列相同性并保留该car的生物学活性(如活化t细胞)的氨基酸序列。可采用例如ncbi的blastp计算两条比对的序列之间的序列相同性。
突变体还包括:在seqidno:2第21-613位所示的氨基酸序列、seqidno:2第21-613位所示的氨基酸序列、seqidno:2第1-613位所示的氨基酸序列或seqidno:2所示的氨基酸序列中具有一个或数个突变(插入、缺失或取代)、同时仍保留该car的生物学活性的氨基酸序列。所述数个突变通常指1-10个以内,例如1-8个、1-5个或1-3个。取代优选是保守性取代。例如,在本领域中,用性能相近或相似的氨基酸进行保守性取代时,通常不会改变蛋白质或多肽的功能。“性能相近或相似的氨基酸”包括例如,具有相似侧链的氨基酸残基的家族,这些家族包括具有碱性侧链的氨基酸(例如赖氨酸、精氨酸、组氨酸)、具有酸性侧链的氨基酸(例如天冬氨酸、谷氨酸)、具有不带电荷的极性侧链的氨基酸(例如甘氨酸、天冬酰胺、谷氨酰胺、丝氨酸、苏氨酸、酪氨酸、半胱氨酸)、具有非极性侧链的氨基酸(例如丙氨酸、缬氨酸、亮氨酸、异亮氨酸脯氨酸、苯丙氨酸、甲硫氨酸、色氨酸)、具有β-分支侧链的氨基酸(例如苏氨酸、缬氨酸、异亮氨酸)和具有芳香侧链的氨基酸(例如酪氨酸、苯丙氨酸、色氨酸、组氨酸)。因此,在本发明多肽中用来自同一侧链类的另一氨基酸残基替换一个或几个位点,将不会在实质上影响其活性。
本发明包括编码本发明融合蛋白的多核苷酸序列。本发明的多核苷酸序列可以是dna形式或rna形式。dna形式包括cdna、基因组dna或人工合成的dna。dna可以是单链的或是双链的。dna可以是编码链或非编码链。本发明也包括编码融合蛋白的多核苷酸序列的简并变异体,即编码相同的氨基酸序列但核苷酸序列有所不同的核苷酸序列。
本文所述的多核苷酸序列通常可以用pcr扩增法获得。具体而言,可根据本文所公开的核苷酸序列,尤其是开放阅读框序列来设计引物,并用市售的cdna库或按本领域技术人员已知的常规方法所制备的cdna库作为模板,扩增而得有关序列。当序列较长时,常常需要进行两次或多次pcr扩增,然后再将各次扩增出的片段按正确次序拼接在一起。例如,在某些实施方案中,编码本文所述融合蛋白的多核苷酸序列如seqidno:1第60-1218位核苷酸所示,或如seqidno:1第1-1218位核苷酸所示。
在某些实施方案中,本发明的多核苷酸序列还包含编码egfr的片段的核苷酸序列。
适用于本发明的egfr可以是本领域周知的egfr,例如来自人的egfr。egfr含有n末端胞外结构域i和ii,胞外结构域iii,胞外结构域iv,跨膜区,近膜区结构域以及酪氨酸激酶结构域。本发明优选使用截短的egfr(“tegfr”,即本文所述的egfr的片段),尤其是不包括其胞内区域(近膜区结构域及酪氨酸激酶结构域)的截短的egfr。在某些实施方案中,还可以进一步将不包括胞内区域的egfr进一步截短成不包括胞外结构域i和ii。因此,在某些实施方案中,本发明使用的tegfr含有egfr的胞外结构域iii、胞外结构域iv以及跨膜区,或由egfr的胞外结构域iii、胞外结构域iv以及跨膜区组成。在某些实施方案中,所述tegfr含有人egfr的第310-646位氨基酸序列,或由人egfr的第310-646位氨基酸序列组成,其中第310-480位氨基酸序列为人egfr的胞外结构域iii,第481-620为人egfr的胞外结构域iv,第621-646位氨基酸为人egfr的跨膜区。在某些实施例中,所述tegfr的氨基酸序列的胞外结构域iii和iv如seqidno:2第774-1108位氨基酸所示的氨基酸序列。
本发明也涉及核酸构建物,该核酸构建物含有本文所述的多核苷酸序列,以及与这些序列操作性连接的一个或多个调控序列。本发明所述的多核苷酸序列可以多种方式被操作以保证所述融合蛋白(car和/或tegfr)的表达。在将核酸构建物插入载体之前可根据表达载体的不同或要求而对核酸构建物进行操作。利用重组dna方法来改变多核苷酸序列的技术是本领域已知的。
调控序列可以是合适的启动子序列。启动子序列通常与待表达蛋白的编码序列操作性连接。启动子可以是在所选择的宿主细胞中显示转录活性的任何核苷酸序列,包括突变的、截短的和杂合启动子,并且可以从编码与该宿主细胞同源或异源的胞外或胞内多肽的基因获得。调控序列也可以是合适的转录终止子序列,由宿主细胞识别以终止转录的序列。终止子序列与编码该多肽的核苷酸序列的3’末端操作性连接。在选择的宿主细胞中有功能的任何终止子都可用于本发明。调控序列也可以是合适的前导序列,对宿主细胞翻译重要的mrna的非翻译区。前导序列与编码该多肽的核苷酸序列的5′末端可操作连接。在选择的宿主细胞中有功能的任何终止子都可用于本发明。
在某些实施方案中,所述核酸构建物是载体。通常通过可操作地连接本发明的多核苷酸序列至启动子,并将构建体并入表达载体,实现本发明多核苷酸序列的表达。该载体对于复制和整合真核细胞可为合适的。典型的克隆载体包含可用于调节期望核酸序列表达的转录和翻译终止子、起始序列和启动子。
本发明的多核苷酸序列可被克隆入许多类型的载体。例如,可被克隆入质粒、噬菌粒、噬菌体衍生物、动物病毒和粘粒。进一步地,载体是表达载体。表达载体可以以病毒载体形式提供给细胞。病毒载体技术在本领域中是公知的并在例如sambrook等(2001,molecularcloning:alaboratorymanual,coldspringharborlaboratory,newyork)和其他病毒学和分子生物学手册中进行了描述。可用作载体的病毒包括但不限于逆转录病毒、腺病毒、腺伴随病毒、疱疹病毒和慢病毒。
通常,合适的载体包含在至少一种有机体中起作用的复制起点、启动子序列、方便的限制酶位点和一个或多个可选择的标记(例如,wo01/96584;wo01/29058;和美国专利号6,326,193)。
例如,在某些实施方案中,本发明使用逆转录病毒载体,该逆转录病毒载体含有复制起始位点,3’ltr,5’ltr,本文所述的多核苷酸序列,以及任选的可选择的标记。
合适的启动子的一个例子为即时早期巨细胞病毒(cmv)启动子序列。该启动子序列是能够驱动可操作地连接至其上的任何多核苷酸序列高水平表达的强组成型启动子序列。合适的启动子的另一个例子为延伸生长因子-1α(ef-1α)。然而,也可使用其他组成型启动子序列,包括但不限于类人猿病毒40(sv40)早期启动子、小鼠乳癌病毒(mmtv)、人免疫缺陷病毒(hiv)长末端重复(ltr)启动子、momulv启动子、鸟类白血病病毒启动子、eb病毒即时早期启动子、鲁斯氏肉瘤病毒启动子、以及人基因启动子,诸如但不限于肌动蛋白启动子、肌球蛋白启动子、血红素启动子和肌酸激酶启动子。进一步地,也可考虑使用诱导型启动子。诱导型启动子的使用提供了分子开关,其能够在期限表达时打开可操作地连接诱导型启动子的多核苷酸序列的表达,而在当表达是不期望的时关闭表达。诱导型启动子的例子包括但不限于金属硫蛋白启动子、糖皮质激素启动子、孕酮启动子和四环素启动子。
为了评估car多肽或其部分的表达,被引入细胞的表达载体也可包含可选择的标记基因或报道基因中的任一个或两者,以便于从通过病毒载体寻求被转染或感染的细胞群中鉴定和选择表达细胞。在其他方面,可选择的标记可被携带在单独一段dna上并用于共转染程序。可选择的标记和报道基因两者的侧翼都可具有适当的调节序列,以便能够在宿主细胞中表达。有用的可选择标记包括例如抗生素抗性基因,诸如neo等等。
报道基因用于鉴定潜在转染的细胞并用于评价调节序列的功能性。在dna已经被引入受体细胞后,报道基因的表达在合适的时间下进行测定。合适的报道基因可包括编码荧光素酶、β-半乳糖苷酶、氯霉素乙酰转移酶、分泌型碱性磷酸酶或绿色萤光蛋白基因的基因。合适的表达系统是公知的并可利用已知技术制备或从商业上获得。
将基因引入细胞和将基因表达入细胞的方法在本领域中是已知的。载体可通过在本领域中的任何方法容易地引入宿主细胞,例如,哺乳动物、细菌、酵母或昆虫细胞。例如,表达载体可通过物理、化学或生物学手段转移入宿主细胞。
将多核苷酸引入宿主细胞的物理方法包括磷酸钙沉淀、脂质转染法、粒子轰击、微注射、电穿孔等等。将感兴趣的多核苷酸引入宿主细胞的生物学方法包括使用dna和rna载体。将多核苷酸引入宿主细胞的化学手段包括胶体分散系统,诸如大分子复合物、纳米胶囊、微球、珠;和基于脂质的系统,包括水包油乳剂、胶束、混合胶束和脂质体。
将多核苷酸引入宿主细胞的生物学方法包括使用病毒载体,特别是逆转录病毒载体,这已经成为最广泛使用的将基因插入哺乳动物例如人细胞的方法。其他病毒载体可源自慢病毒、痘病毒、单纯疱疹病毒i、腺病毒和腺伴随病毒等等。已经开发了许多基于病毒的系统,用于将基因转移入哺乳动物细胞。例如,逆转录病毒提供了用于基因传递系统的方便的平台。可利用本领域中已知的技术将选择的基因插入载体并包装入逆转录病毒颗粒。该重组病毒可随后被分离和传递至体内或离体的对象细胞。许多反转录病毒系统在本领域中是已知的。在一些实施方案中,使用腺病毒载体。许多腺病毒载体在本领域中是已知的。在一个实施方案中,使用慢病毒载体。
因此,在某些实施方案中,本发明还提供用于活化t细胞的逆转录病毒,该病毒含有本文所述的逆转录病毒载体以及相应的包装基因,如gag、pol和vsvg。
适用于本发明的t细胞可以是各种来源的各种类型的t细胞。例如,t细胞可来源于b细胞恶性肿瘤患者的pbmc。
在某些实施方案中,获得t细胞后,可先用适量的(例如30~80ng/ml,如50ng/ml)的cd3抗体刺激活化,然后在含有适量的(例如30~80iu/ml,如50iu/ml)的il2培养基进行培养备用。
因此,在某些实施方案中,本发明提供一种基因修饰的t细胞,该基因修饰的t细胞含有本文所述的多核苷酸序列,或含有本文所述的逆转录病毒载体,或感染了本文所述的逆转录病毒,或采用本文所述的方法制备得到,或稳定表达本文所述的融合蛋白和任选的tegfr。
本发明的car-t细胞可经历稳固的体内t细胞扩展并在血液和骨髓中以高水平持续延长的时间量,并形成特异性记忆t细胞。不希望被任何具体的理论所束缚,在遇到并随后消除表达替代抗原的靶细胞后,本发明的car-t细胞可体内分化成中心记忆样状态。
本发明还包括一类细胞疗法,其中t细胞被基因修饰以表达本文所述的car和任选的tegfr,和car-t细胞被注入需要其的接受者中。注入的细胞能够杀死接受者的肿瘤细胞。不像抗体疗法,car-t细胞能够体内复制,产生可导致持续肿瘤控制的长期持久性。
由car-t细胞引起的抗肿瘤免疫应答可为主动或被动免疫应答。另外,car介导的免疫应答可为过继免疫疗法步骤的一部分,其中car-t细胞诱导对car中的抗原结合部分特异性的免疫应答。
因此,可采用本发明的car、其编码序列、核酸构建物、表达载体、病毒以及car-t细胞治疗的疾病优选为cd19和cd20介导的疾病。
本发明的car-修饰的t细胞可被单独施用或作为药物组合物与稀释剂和/或与其他组分诸如相关的细胞因子或细胞群结合施用。简单地说,本发明的药物组合物可包括如本文所述的car-t细胞,与一种或多种药学或生理学上可接受载体、稀释剂或赋形剂结合。这样的组合物可包括缓冲液诸如中性缓冲盐水、硫酸盐缓冲盐水等等;碳水化合物诸如葡萄糖、甘露糖、蔗糖或葡聚糖、甘露醇;蛋白质;多肽或氨基酸诸如甘氨酸;抗氧化剂;螯合剂诸如edta或谷胱甘肽;佐剂(例如,氢氧化铝);和防腐剂。
本发明的药物组合物可以以适于待治疗(或预防)的疾病的方式施用。施用的数量和频率将由这样的因素确定,如患者的病症、和患者疾病的类型和严重度。
当指出“免疫学上有效量”、“抗肿瘤有效量”、“肿瘤-抑制有效量”或“治疗量”时,待施用的本发明组合物的精确量可由医师确定,其考虑患者(对象)的年龄、重量、肿瘤大小、感染或转移程度和病症的个体差异。可通常指出:包括本文描述的t细胞的药物组合物可以以104至109个细胞/kg体重的剂量,优选105至106个细胞/kg体重的剂量。t细胞组合物也可以以这些剂量多次施用。细胞可通过使用免疫疗法中公知的注入技术(见例如rosenberg等,neweng.j.ofmed.319:1676,1988)施用。对于具体患者的最佳剂量和治疗方案可通过监测患者的疾病迹象并因此调节治疗由医学领域技术人员容易地确定。
对象组合物的施用可以以任何方便的方式进行,包括通过喷雾法、注射、吞咽、输液、植入或移植。本文描述的组合物可被皮下、皮内、瘤内、结内、脊髓内、肌肉内、通过静脉内注射或腹膜内施用给患者。在一个实施方案中,本发明的t细胞组合物通过皮内或皮下注射被施用给患者。在另一个实施方案中,本发明的t细胞组合物优选通过静脉注射施用。t细胞的组合物可被直接注入肿瘤、淋巴结或感染位置。
在本发明的一些实施方案中,本发明的car-t细胞或其组合物可与本领域已知的其它疗法结合。所述疗法包括但不限于化疗、放疗和免疫抑制剂。例如,可结合本领域周知的治疗cd19和cd20介导的疾病的放疗或化疗制剂进行治疗。
本文中,“抗肿瘤效应”指一种生物学效应,其可由肿瘤体积的减少、肿瘤细胞数的减少、转移数的减少、预期寿命的增加或与癌相关的各种生理症状的改善表示。
“患者”、“对象”、“个体”等等在本文中可交换使用,指可引起免疫应答的活有机体,如哺乳动物。例子包括但不限于人、狗、猫、小鼠、大鼠和其转基因物种。
本发明的有益效果是:本发明采用cd20scfv+cd19scfv基因序列,并从ncbigenbank数据库中搜索到人igg4铰链区、人cd28跨膜区、人41bb胞内区、以及人cd3ζ胞内区和人的truncatedegfr(tegfr)基因序列信息,全基因合成嵌合抗原受体cd20scfv-cd19scfv-41bb-cd3ζ-tegfr的基因片段,插入到逆转录病毒载体rv,其可以用于重组引入目的核酸序列,即编码car的核酸序列。重组质粒在293t细胞中包装病毒,感染t细胞,使t细胞表达该嵌合抗原受体。在本发明的一个实施方案中,实现嵌合抗原受体基因修饰的t淋巴细胞的转化方法是基于逆转录病毒转化方法。该方法具有转化效率高,外源基因能够稳定表达,且可以缩短体外培养t淋巴细胞到达临床级数量的时间等优点。在该转基因t淋巴细胞表面,转化的核酸通过转录、翻译表达在其上。本发明所获得的表达car的逆转录病毒通过retronectin法制备car-t细胞,制备3天后的car-t细胞用流式检测car的感染效率,制备5天后car-t细胞在体外与cd19或cd20阳性的肿瘤细胞(k562-cd19、k562-cd20、raji、nalm6)共培养5小时检测cd107a表达和ifnγ的分泌,制备5天后car-t细胞在体外与cd19或cd20阳性的肿瘤细胞(k562-cd19、k562-cd20、raji、nalm6)共培养5小时法检测car-t细胞对肿瘤细胞的特异性杀伤作用(细胞毒性)。因此本发明所述cd20-cd19-bbz-tegfrcart可在b细胞急性/慢性淋巴细胞白血病和淋巴瘤治疗中得到应用。
本发明通过参考以下实验实施例进一步详细地进行描述。这些实施例仅出于说明性的目的提供,并不意欲为限制性的,除非另有规定。因此,本发明决不应被解释为限于以下实施例,而是应被解释为包括由于本文提供的教导变得显而易见的任何和全部的变化。实施例中所用的方法和试剂,除非另有说明,否则为本领域常规的方法和试剂。
实施例1:cd20scfv-cd19scfv-41bb-cd3ζ-tegfr基因序列的确定
1.1从ncbi网站数据库搜索到人igg4铰链区、人cd28跨膜区、人41bb胞内区、以及人cd3ζ胞内区和人的truncatedegfr(tegfr)基因序列,抗cd20单链抗体克隆号为leu-16,抗cd19单链抗体克隆号为fmc63,这些序列在网站http://sg.idtdna.com/site上进行密码子优化,保证在编码氨基酸序列不变的情况下更适合人类细胞表达。
各氨基酸和基因序列信息见seqencelisting(sequnceidno.1-2)。
将上述序列依次进行连接,在各序列连接处引入不同酶切位点,形成完整的cd20-cd19-bbz-tegfr基因序列信息。
1.2重组质粒测序
将重组质粒送上海生工生物技术有限公司进行测序,将测序结果与拟合成的cd20-cd19-bbz-tegfr序列比对来验证序列是否正确。测序引物为:
正义序列:agcatcgttctgtgttgtctc(sequnceidno.3)
反义序列:tgtttgtcttgtggcaatacac(sequnceidno.4)
本实施例所构建得到的质粒图谱如图1所示。图2显示该逆转录病毒表达质粒的部分测序结果峰值图。
实施例2:包含car分子的核酸序列的病毒载体的构建
将实施例1中制备的car分子的核苷酸序列经noti(neb)和ecori(neb)双酶切、经t4连接酶(neb)连接插入逆转录病毒rv载体的noti-ecori位点,转化到感受态e.coli(dh5α),经测序正确后,使用qiagen公司的质粒纯化试剂盒提取并纯化质粒,纯化质粒的质粒磷酸钙法转染293t细胞进行逆转录病毒包装实验。
实施例3:逆转录病毒包装
1.第1天293t细胞应是小于20代,不过分长满的。以0.6*10^6cells/ml铺板,10cm皿添加10ml的dmem培养基,充分混匀细胞,37度培养过夜;
2.第2天293t细胞融合度达到90%左右进行转染(通常是铺板14-18h左右);准备质粒复合物,各种质粒的量为rv-cd20-cd19-bbz-tegfr为12.5ug,gag-pol为10ug,vsvg为6.25ug,cacl2250ul,h2o为1ml总体积为1.25ml;在另一个管里添加跟质粒复合物等体积的hbs,边加质粒复合物边涡旋震荡20s。温柔地将混合物沿着边加入到293t皿中,37度培养4h,去除培养基,pbs洗一遍,重新加入预热的新鲜培养基;
3.第4天:转染48h后收集上清并用0.45um滤器过滤后分装保存于-80度,继续添加预热的新鲜dmem培养基。
实施例4:逆转录病毒感染人的t细胞
1.用ficcol分离液(天津灏洋)分离获得较纯的cd3+t细胞,用含5%ab血清x-vivo(lonza)培养基调整细胞密度为1×106/ml。将细胞以1ml/孔接种到预先用抗人50ng/mlcd3抗体(北京同立海元)和50ng/ml41bb抗体(北京同立海元),再加入100iu/ml的白细胞介素2(北京双鹭),刺激培养48小时后病毒感染。
2.t细胞活化培养后隔天,pbs稀释至终浓度为15μg/ml的retronectin(takara)包被non-tissuetreated(corning)培养板,24孔板每孔250μl。避光,4℃过夜备用。3.t细胞活化培养两天后,取出2块包被好的24孔板,吸弃包被液,加入含2%bsa的hbss室温封闭30min。封闭液体积为每孔500μl,吸弃封闭液,用含2.5%hepes的hbss洗板两次。
4.病毒液加入孔内,每孔加2ml病毒液,32℃,2000g,离心2h。
5.弃去上清液,24孔板每孔加入活化后的t细胞1×106个,体积1ml,培养基为t细胞培养基中添加il-2200iu/ml。30℃,1000g,离心10min。
6.离心完毕后,将培养板置于37℃,5%co2培养箱中培养。
7.感染后24h,将细胞悬液吸出,1200rpm,4℃,离心7min。
8.细胞感染后,每天观察细胞的密度,适时补加含il-2100iu/ml的t细胞培养液,使t细胞的密度维持在5×105/ml左右,使细胞扩增。
实施例5:流式细胞仪检测感染后t淋巴细胞表面car蛋白的表达
分别离心收集感染后72小时的cd20-cd19-bbz-tegfr细胞,pbs洗涤1次后弃上清,加入相应的抗体避光30min后pbs洗涤,重悬,最后流式细胞仪检测。car+由anti-mouseiggf(ab')antibody(jacksonimmunoresearch)和apc-antiegfr(biolegend)抗体检测。
本实施例结果如图3所示,cd20-cd19-tegfrcart阳性率在10%以上。
实施例6:检测各靶细胞cd19和cd22表达
k562-cd19、k562-cd20、raji、nalm6、k562同时染色anti-humancd19和anti-humancd20,流式上机检测cd19和cd20表达。
本实施例结果如图4所示,k562-cd19细胞和nalm6细胞只表达cd19,k562-cd20细胞只表达cd20,raji细胞同时表达cd19和cd20。
实施例7:car-t细胞与靶细胞共培养后cd107a表达检测
1.取一块v底96孔板,每孔加cart/nt细胞2*105个和靶细胞(k562-cd19、k562-cd20、raji、nalm6)/对照细胞(k562)2*105个,重悬为200ul不含il-2的x-vivo完全培养基,加入bdgolgistop(含monesin,每1ml培养基中加入1μlbdgolgistop),每孔加入2ulcd107a抗体(1:50),37℃孵育4小时,收集细胞。
2.将样品离心去除培养基,pbs洗细胞一次,400g,4℃离心5分钟。弃上清,每管加入适量特异性表面抗体car、cd3,重悬体积100ul,冰上避光孵育30分钟。
3.每管用3ml的pbs清洗细胞1次,400g离心5分钟。仔细吸去上清。
适量pbs重悬,流式细胞仪检测car、cd3、cd107a。
本实施例结果如图5所示,cd20-cd19-tegfrcart细胞与靶细胞(k562-cd19、k562-cd20)共培养cd107a表达在70%左右;与靶细胞(raji、nalm6)共培养cd107a表达在80%左右;与对照细胞(k562)则几乎没有cd107a表达。
实施例8:car-t细胞与靶细胞共培养后ifnγ分泌检测
1.取制备好的car-t细胞,重悬与lonza培养基中,调整细胞浓度为1×106/ml。
2.实验组每孔含靶细胞(k562-cd19、k562-cd20、raji、nalm6)或阴性对照细胞(k562)2×105个,car-t/nt细胞2×105个,200μl不含il-2的lonza培养基。充分混匀后加入96孔板中。加入bdgolgistop(含monesin,每1ml培养基中加入1μlbdgolgistop),充分混匀后,37℃孵育5小时。收集细胞,作为实验组。
3.每管用1ml的pbs清洗细胞1次,300g离心5分钟。弃上清,每管加入适量特异性表面抗体car、cd3,重悬体积100ul,冰上避光孵育30分钟。
4.pbs洗细胞后,加入250μl/ep管fixation/permeabilizationsolution,4℃孵育20分钟以固定细胞及破膜。用1×bdperm/washtmbuffer清洗细胞2次,1ml/次。
5.进行胞内因子染色,取适量ifn-γ细胞因子荧光抗体或阴性对照,用bdperm/washtmbuffer稀释至50μl。用此抗体稀释液充分重悬已固定破膜的细胞,4℃避光孵育30min,1×bdperm/washtmbuffer1ml/次清洗细胞2次,然后用pbs重悬。
6.流式细胞仪检测car、cd3、ifn-γ。
本实施例结果如图6所示,cd20-cd19-tegfrcart细胞与靶细胞(k562-cd19、k562-cd20)共培养inf-γ分泌在40%左右;与靶细胞(raji、nalm6)共培养inf-γ分泌在60%左右;与对照细胞(k562)则几乎没有inf-γ分泌。
实施例9:car-t细胞与靶细胞共培养后检测肿瘤特异性细胞杀伤作用
1.k562细胞(不含cd19或cd20,为阴性对照细胞)重悬在无血清培养基(1640)中,调整细胞浓度为1×106/ml,加入荧光染料bmqc(2,3,6,7-tetrahydro-9-bromomethyl-1h,5hquinolizino(9,1-gh)coumarin)至终浓度为5μm。
2.混匀,37℃孵育30min。
3.室温,1500rpm离心5min,弃上清,并重悬细胞于细胞毒性培养基(无酚红1640+5%ab血清)中,37℃孵育60min。
4.新鲜细胞毒性培养基清洗细胞两遍,并重悬在新鲜细胞毒性培养基中,密度1×106/ml。
5.k562-cd19、k562-cd20、raji、nalm6细胞(含cd19或cd20,为靶细胞)悬浮在含有0.1%bsa的pbs中,调整浓度为1×106/ml。
6.加入荧光染料cfse(carboxyfluoresceindiacetatesuccinimidylester)至终浓度为1μm。
7.混匀,37℃孵育10min。
8.孵育结束后,加入与细胞悬液等体积的fbs,室温孵育2min以终止标记反应。
9.清洗细胞并重悬在新鲜细胞毒性培养基中,密度1×106/ml。
10.清洗效应t细胞并悬浮在细胞毒性培养基中,调整浓度为5×106/ml。
11.在所有的实验中,感染了cd20-cd19-bbz-tegfrcar的效应t细胞(car-tcell)的细胞毒性和未感染的阴性对照效应t细胞(ntcell)的细胞毒性做比较,并且这些效应t细胞来自同一个病人。
12.cd20-cd19-bbz-tegfrcar-t和阴性对照效应t细胞,按照t细胞:靶细胞=5:1,1:1,的比例,于5ml无菌试验管(bdbiosciences)进行培养。每一个共培养组中,靶细胞为raji细胞100,000个(50μl),阴性对照细胞为100,000个k562细胞(50μl)。同时设置一组只包含raji靶细胞和k562阴性对照细胞。
13.将共培养细胞置于37℃孵育5h。
14.孵育完成后,pbs清洗细胞,然后立即按照说明书推荐的浓度快速加入7-aad(7-aminoactinomycind),冰上孵育30min。
15.不需清洗,直接进行流式上机检测,数据用flowjo进行分析。
16.分析使用7aad阴性的活细胞设门,测定t细胞和靶细胞共培养后活的靶细胞和活的阴性对照细胞的比例。
细胞毒性杀伤细胞%=(1-(含效应细胞时靶细胞活细胞数/含效应细胞时k562活细胞数)/(无效应细胞时靶细胞活细胞数/无效应细胞时k562活细胞数))*100%。
本实施例结果如图7所示,效靶比e:t=5:1时,cd20-cd19-tegfrcart对靶细胞(k562-cd19、k562-cd20)杀伤率都大于30%;对靶细胞(raji、nalm6)杀伤率都在60%左右。表明cd20-cd19-tegfrcart对表达cd19或cd20的靶细胞都有明显功能。
序列表
<110>上海恒润达生生物科技有限公司
<120>一种双靶向嵌合抗原受体及其用途
<160>1
<170>patentinversion3.3
<210>1
<211>3324
<212>dna
<213>人工序列
<400>1
atggagacagacacactcctgctatgggtgctgctgctctgggttccaggttccacaggt60
gacattgtgctgacccaatctccagctatcctgtctgcatctccaggggagaaggtcaca120
atgacttgcagggccagctcaagtgtaaattacatggactggtaccagaagaagccagga180
tcctcccccaaaccctggatttatgccacatccaacctggcttctggagtccctgctcgc240
ttcagtggcagtgggtctgggacctcttactctctcacaatcagcagagtggaggctgaa300
gatgctgccacttattactgccagcagtggagttttaatccacccacgttcggagggggg360
accaagctggaaataaaaggcagtactagcggtggtggctccgggggcggttccggtggg420
ggcggcagcagcgaggtgcagctgcagcagtctggggctgagctggtgaagcctggggcc480
tcagtgaagatgtcctgcaaggcttctggctacacatttaccagttacaatatgcactgg540
gtaaagcagacacctggacagggcctggaatggattggagctatttatccaggaaatggt600
gatacttcctacaatcagaagttcaaaggcaaggccacattgactgcagacaaatcctcc660
agcacagcctacatgcagctcagcagcctgacatctgaggactctgcggactattactgt720
gcaagatctaattattacggtagtagctactggttcttcgatgtctggggcgcagggacc780
acggtcacagtaagtagcggtggaggcggcagtggcggaggtgggagcggagggggcggt840
tccggtggcgggggatctgaggtgaagctgcaggaaagcggccctggcctggtggccccc900
agccagagcctgagcgtgacctgcaccgtgagcggcgtgagcctgcccgactacggcgtg960
agctggatccggcagccccccaggaagggcctggaatggctgggcgtgatctggggcagc1020
gagaccacctactacaacagcgccctgaagagccggctgaccatcatcaaggacaacagc1080
aagagccaggtgttcctgaagatgaacagcctgcagaccgacgacaccgccatctactac1140
tgcgccaagcactactactacggcggcagctacgccatggactactggggccagggcacc1200
agcgtgaccgtgagcagcggcagcacctccggcagcggcaagcctggcagcggcgagggc1260
agcaccaagggcgacatccagatgacccagaccacctccagcctgagcgccagcctgggc1320
gaccgggtgaccatcagctgccgggccagccaggacatcagcaagtacctgaactggtat1380
cagcagaagcccgacggcaccgtcaagctgctgatctaccacaccagccggctgcacagc1440
ggcgtgcccagccggtttagcggcagcggctccggcaccgactacagcctgaccatctcc1500
aacctggaacaggaagatatcgccacctacttttgccagcagggcaacacactgccctac1560
acctttggcggcggaacaaagctggaaatcaccgagagcaagtacggaccgccctgcccc1620
ccttgccctatgttctgggtgctggtggtggtcggaggcgtgctggcctgctacagcctg1680
ctggtcaccgtggccttcatcatcttttgggtgaaacggggcagaaagaaactcctgtat1740
atattcaaacaaccatttatgagaccagtacaaactactcaagaggaagatggctgtagc1800
tgccgatttccagaagaagaagaaggaggatgtgaactgcgggtgaagttcagcagaagc1860
gccgacgcccctgcctaccagcagggccagaatcagctgtacaacgagctgaacctgggc1920
agaagggaagagtacgacgtcctggataagcggagaggccgggaccctgagatgggcggc1980
aagcctcggcggaagaacccccaggaaggcctgtataacgaactgcagaaagacaagatg2040
gccgaggcctacagcgagatcggcatgaagggcgagcggaggcggggcaagggccacgac2100
ggcctgtatcagggcctgtccaccgccaccaaggatacctacgacgccctgcacatgcag2160
gccctgcccccaaggcgagctaaacgaggctcaggcgcgacgaactttagtttgctgaag2220
caagctggggatgtagaggaaaatccgggtcccatgttgctccttgtgacgagcctcctg2280
ctctgcgagctgccccatccagccttcctcctcatcccgcggaaggtgtgcaatggcata2340
ggcattggcgagtttaaagattctctgagcataaatgctacgaatattaagcatttcaag2400
aattgtacttctattagtggcgacctccatattcttccggttgccttcaggggtgactct2460
ttcacccacacacctccattggatccacaagaacttgacatcctgaagacggttaaagag2520
attacaggcttcctccttatccaagcgtggcccgagaacagaacggacttgcacgccttt2580
gagaacctcgaaataatacggggtcggacgaagcaacacggccaatttagccttgcggtt2640
gttagtctgaacattacttctctcggccttcgctctttgaaagaaatcagcgacggagat2700
gtcatcattagtggaaacaagaacctgtgctacgcgaacacaatcaactggaagaagctc2760
ttcggtacttcaggccaaaagacaaagattattagtaacagaggagagaatagctgtaag2820
gctaccggacaagtttgtcacgccttgtgtagtccagagggttgctggggaccggaacca2880
agggattgcgtcagttgccggaacgtgagtcgcggacgcgagtgtgtggataagtgcaat2940
cttctggaaggggaaccgcgagagtttgtagaaaattccgaatgtatacagtgtcatccc3000
gagtgtcttccacaagcaatgaatatcacatgtacagggaggggtcctgataactgtatc3060
caatgtgcacactacatagatggtcctcactgtgtaaagacgtgccccgccggagtaatg3120
ggtgaaaacaacaccctcgtgtggaagtacgccgatgccgggcatgtctgtcatttgtgt3180
catcccaactgcacatatggctgtaccggtcctggattggagggctgtccaacaaacggg3240
ccgaaaataccgagtatcgcaacaggcatggtgggagcacttttgcttctcctcgttgtc3300
gccctgggcatcggcttgttcatg3324
<160>1
<170>patentinversion3.3
<210>2
<211>1108
<212>prt
<213>人工序列
<400>2
metgluthraspthrleuleuleutrpvalleuleuleutrpvalpro
151015
glyserthrglyaspilevalleuthrglnserproalaileleuser
202530
alaserproglyglulysvalthrmetthrcysargalaserserser
354045
valasntyrmetasptrptyrglnlyslysproglyserserprolys
505560
protrpiletyralathrserasnleualaserglyvalproalaarg
65707580
pheserglyserglyserglythrsertyrserleuthrileserarg
859095
valglualagluaspalaalathrtyrtyrcysglnglntrpserphe
100105110
asnproprothrpheglyglyglythrlysleugluilelysglyser
115120125
thrserglyglyglyserglyglyglyserglyglyglyglyserser
130135140
gluvalglnleuglnglnserglyalagluleuvallysproglyala
145150155160
servallysmetsercyslysalaserglytyrthrphethrsertyr
165170175
asnmethistrpvallysglnthrproglyglnglyleuglutrpile
180185190
glyalailetyrproglyasnglyaspthrsertyrasnglnlysphe
195200205
lysglylysalathrleuthralaasplysserserserthralatyr
210215220
metglnleuserserleuthrsergluaspseralaasptyrtyrcys
225230235240
alaargserasntyrtyrglysersertyrtrpphepheaspvaltrp
245250255
glyalaglythrthrvalthrvalserserglyglyglyglysergly
260265270
glyglyglyserglyglyglyglyserglyglyglyglysergluval
275280285
lysleuglngluserglyproglyleuvalalaproserglnserleu
290295300
servalthrcysthrvalserglyvalserleuproasptyrglyval
305310315320
sertrpileargglnproproarglysglyleuglutrpleuglyval
325330335
iletrpglysergluthrthrtyrtyrasnseralaleulysserarg
340345350
leuthrileilelysaspasnserlysserglnvalpheleulysmet
355360365
asnserleuglnthraspaspthralailetyrtyrcysalalyshis
370375380
tyrtyrtyrglyglysertyralametasptyrtrpglyglnglythr
385390395400
servalthrvalserserglyserthrserglyserglylysprogly
405410415
serglygluglyserthrlysglyaspileglnmetthrglnthrthr
420425430
serserleuseralaserleuglyaspargvalthrilesercysarg
435440445
alaserglnaspileserlystyrleuasntrptyrglnglnlyspro
450455460
aspglythrvallysleuleuiletyrhisthrserargleuhisser
465470475480
glyvalproserargpheserglyserglyserglythrasptyrser
485490495
leuthrileserasnleugluglngluaspilealathrtyrphecys
500505510
glnglnglyasnthrleuprotyrthrpheglyglyglythrlysleu
515520525
gluilethrgluserlystyrglyproprocysproprocyspromet
530535540
phetrpvalleuvalvalvalglyglyvalleualacystyrserleu
545550555560
leuvalthrvalalapheileilephetrpvallysargglyarglys
565570575
lysleuleutyrilephelysglnprophemetargprovalglnthr
580585590
thrglnglugluaspglycyssercysargpheprogluglugluglu
595600605
glyglycysgluleuargvallyspheserargseralaaspalapro
610615620
alatyrglnglnglyglnasnglnleutyrasngluleuasnleugly
625630635640
argarggluglutyraspvalleuasplysargargglyargasppro
645650655
glumetglyglylysproargarglysasnproglngluglyleutyr
660665670
asngluleuglnlysasplysmetalaglualatyrsergluilegly
675680685
metlysglygluargargargglylysglyhisaspglyleutyrgln
690695700
glyleuserthralathrlysaspthrtyraspalaleuhismetgln
705710715720
alaleuproproargargalalysargglyserglyalathrasnphe
725730735
serleuleulysglnalaglyaspvalglugluasnproglypromet
740745750
leuleuleuvalthrserleuleuleucysgluleuprohisproala
755760765
pheleuleuileproarglysvalcysasnglyileglyileglyglu
770775780
phelysaspserleuserileasnalathrasnilelyshisphelys
785790795800
asncysthrserileserglyaspleuhisileleuprovalalaphe
805810815
argglyaspserphethrhisthrproproleuaspproglngluleu
820825830
aspileleulysthrvallysgluilethrglypheleuleuilegln
835840845
alatrpprogluasnargthraspleuhisalaphegluasnleuglu
850855860
ileileargglyargthrlysglnhisglyglnpheserleualaval
865870875880
valserleuasnilethrserleuglyleuargserleulysgluile
885890895
seraspglyaspvalileileserglyasnlysasnleucystyrala
900905910
asnthrileasntrplyslysleupheglythrserglyglnlysthr
915920925
lysileileserasnargglygluasnsercyslysalathrglygln
930935940
valcyshisalaleucysserprogluglycystrpglyproglupro
945950955960
argaspcysvalsercysargasnvalserargglyargglucysval
965970975
asplyscysasnleuleugluglygluproarggluphevalgluasn
980985990
serglucysileglncyshisproglucysleuproglnalametasn
99510001005
ilethrcysthrglyargglyproaspasncysileglncysala
101010151020
histyrileaspglyprohiscysvallysthrcysproalagly
102510301035
valmetglygluasnasnthrleuvaltrplystyralaaspala
104010451050
glyhisvalcyshisleucyshisproasncysthrtyrglycys
105510601065
thrglyproglyleugluglycysprothrasnglyprolysile
107010751080
proserilealathrglymetvalglyalaleuleuleuleuleu
108510901095
valvalalaleuglyileglyleuphemet
11001105
<210>3
<211>21
<212>dna
<213>人工序列
<223>引物
<400>3
agcatcgttctgtgttgtctc21
<210>4
<211>22
<212>dna
<213>人工序列
<223>引物
tgtttgtcttgtggcaatacac22
- 还没有人留言评论。精彩留言会获得点赞!