一株多抑制物胁迫耐受性酿酒酵母及制备方法、应用与流程
本发明属于微生物生物技术领域,特别涉及一株多抑制物胁迫耐受性酿酒酵母及制备方法、应用。
背景技术:
燃料乙醇作为一种清洁的可再生能源是重要的石油替代品,具有良好的发展前景。以来源丰富且廉价的纤维素生物质农林废弃物为原料生产燃料乙醇,是国内外生物能源发展的重要方向。但是,木质纤维素原料在预处理过程中可产生对细胞生长和代谢具有毒性的副产物,包括弱酸类、醛类和酚类等(omics:ajournalofintegrativebiology,2011,15:647-653)。这些副产物通过抑制酵母菌有氧呼吸、增加细胞膜透性、破坏胞内酶活性等途径抑制微生物菌体生长和乙醇发酵(journalofindustrialmicrobiology&biotechnology,2004,31:345-352),导致较低的纤维素乙醇生成效率。
yol032w是磷脂合成蛋白基因,在胆碱存在时该基因的表达明显被抑制,表明yol032w与磷脂酰胆合成相关(genetics,2006,173:621-634)。有研究者将单倍体酿酒酵母by4741的yol032w基因敲除后其在含山梨酸平板上的生长状态明显下降,表明该基因与弱酸耐性相关(molecularbiologyofthecell,2004,15:706-720)。而该基因过表达与乙酸、糠醛和香草醛耐受性关系尚有待研究。
技术实现要素:
为弥补现有技术的不足,本发明提供一株多抑制物胁迫耐受性酿酒酵母及制备方法、应用,能在分别含有高浓度乙酸、糠醛和香草醛等环境胁迫条件下良好生长和具有较高的乙醇发酵性能。
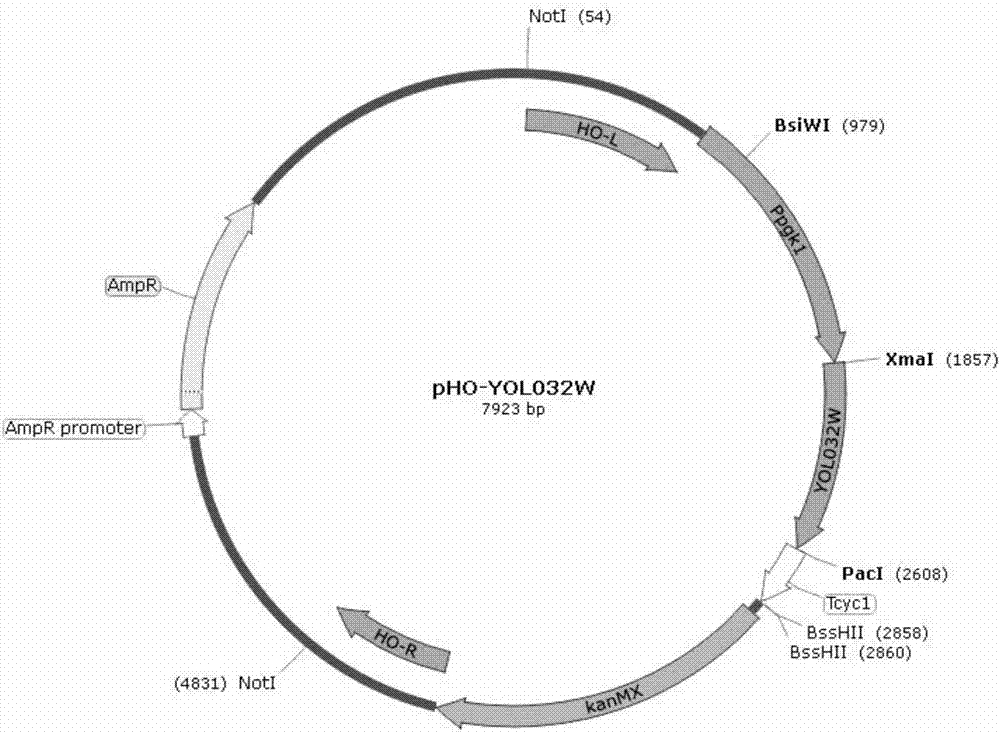
本发明的技术方案如下:一株多抑制物胁迫耐受性酿酒酵母s288c-yol032w(saccharomycescerevisiae)及制备方法、应用,该菌株的保藏编号为cgmccno.13926,保藏于中国微生物菌种保藏管理委员会普通微生物中心,保藏日期为2017年03月24日,保藏地址为北京市朝阳区北辰西路1号院3号;该菌株携带有yol032w基因,该基因genbank登录号为nm_001183286.1。该酿酒酵母菌株为单细胞真菌,一般呈卵圆形、圆形。菌落形态与细菌相似,但较大较厚,呈乳白色,表面湿润、粘稠。营养方式为异养兼性厌氧型。在培养基中生长时为游离态,乳白色。
pgk1启动子序列genbank登录号为fj415226.1,cyc1终止子序列genbank登录号为ef210198.1。通过基因工程手段在酵母的pho整合载体(ncbi:#af324728,美国utah大学davidj.stillman惠赠;nucleicacidsresearch,2001,29:e59)的基础上连接pgk1强启动子和cyc1终止子构成pho组成型表达载体。然后把pcr扩增获得的yol032w基因连接在pgk1启动子和cyc1终止子之间,线性化后电转导入酿酒酵母s288c中,进行过表达。导入的目的片段在酿酒酵母基因组的ho基因位点(yeast,1997,13:1563-1573)进行整合表达。
本发明采用酿酒酵母的pho组成型表达载体进行重组菌株的整合表达,pho组成型表达载体可以通过同源重组的方式在酿酒酵母的ho基因位点进行整合表达,该载体通过基因工程手段在质粒的多克隆位点插入pgk1启动子和cyc1终止子,获得组成型整合表达,根据遗传霉素g418抗性进行筛选。
自絮凝酵母spsc01具有絮凝能力好、絮凝性状稳定及乙醇发酵性能优良等特点,已在燃料乙醇生产中得到应用。酵母细胞絮凝后,对包括乙醇抑制在内的诸多环境胁迫的耐受性得到提高(journalofbiotechnology,2007,131:270-275)。鉴于絮凝酵母spsc01在乙醇发酵中的诸多优点,且与模式菌株s288c相比,其基因组有大量突变。因此,本发明中以絮凝酵母spsc01为模板获取yol032w基因,在酿酒酵母s288c中进行过表达,获得重组酿酒酵母菌株s288c-yol032w。目前的研究大多只关注菌株对一种或者一类抑制物的耐性的提高,而本发明中重组酵母菌株s288c-yol032w分别对木质纤维素预处理过程中产生的多种抑制物的耐性均有提高,如乙酸、糠醛和香草醛。在高浓度抑制物添加下s288c-yol032w与对照菌株s288c-ho相比,不仅生长状态良好且乙醇发酵性能提高。
本发明同时请求保护所述的重组酿酒酵母菌s288c-yol032w在抑制物胁迫条件下的应用,比如在高浓度乙酸、糠醛和香草醛的条件下生长良好,并且有较高的乙醇发酵效率。
本发明的有益效果如下:(1)本发明提供了一种新型菌株;(2)本发明的重组酿酒酵母s288c-yol032w能在分别含有高浓度乙酸、糠醛和香草醛环境胁迫条件下良好生长和具有较高的乙醇发酵效率。
附图说明
图1yol032w基因片段的pcr扩增。
图2pho-yol032w载体。
图3阳性转化子pcr鉴定。
图4重组空载酵母s288c-ho对照菌株和重组酿酒酵母s288c-yol032w在含有不同胁迫抑制物平板中生长比较;
其中:(a)不添加抑制物情况下过表达菌株和对照菌株生长情况;(b)乙酸10g/l添加下过表达菌株和对照菌株生长情况;(c)香草醛1g/l添加下过表达菌株和对照菌株生长情况;(d)糠醛1.5g/l添加下过表达菌株和对照菌株生长情况。
图5重组空载酵母s288c-ho对照菌株和重组酿酒酵母s288c-yol032w在10g/l乙酸添加下乙醇发酵情况;
其中:(a)乙酸10g/l添加下过表达菌株和对照菌株生长随时间变化情况;(b)乙酸10g/l添加下发酵液中乙醇浓度随时间的变化情况;(c)乙酸10g/l添加下发酵液中残余葡萄糖随时间变化情况。
图6重组空载酵母s288c-ho对照菌株和重组酿酒酵母s288c-yol032w在2g/l香草醛添加下乙醇发酵情况;
其中:(a)2g/l香草醛添加下过表达菌株和对照菌株生长随时间变化情况;(b)2g/l香草醛添加下发酵液中乙醇浓度随时间的变化情况;(c)2g/l香草醛添加下发酵液中残余葡萄糖随时间变化情况。
图7重组空载酵母s288c-ho对照菌株和重组酿酒酵母s288c-yol032w在3g/l糠醛添加下乙醇发酵情况;
其中:(a)3g/l糠醛添加下过表达菌株和对照菌株生长随时间变化情况;(b)3g/l糠醛添加下发酵液中乙醇浓度随时间的变化情况;(c)3g/l糠醛添加下发酵液中残余葡萄糖随时间变化情况。
具体实施方式
实施例1:磷脂合成蛋白基因yol032w的重组酿酒酵母的构建和转化方法
本发明中磷脂合成蛋白基因yol032w序列来自于ncbi公共数据库,yol032w基因的genbank登录号为nm_001183286.1。基因的启动子用pgk1,用cyc1作为终止子,以酶切连接的方式在pgk1启动子和cyc1终止子之间分别插入yol032w基因。然后通过限制性内切酶noti把构建的质粒酶切成线性,转化到酿酒酵母s288c中进行整合表达。
1.1酿酒酵母基因组dna提取
(1)从新培养的ypd平板上挑取酵母单菌落,接入100ml液体ypd培养基中,在30℃,150rpm的条件下过夜培养。
(2)取4ml菌液,10000rpm,离心1min,收集酵母细胞,用去离子水洗涤2次,将培养基等细胞表面杂质去除。
(3)在上述酵母细胞中加入500μl裂解液(2%tritonx-100,1%sds,10mmtris·hclph8.0,1mmedta),重悬酵母细胞,转入1.5ml离心管。
(4)加入大约500μl直径为0.5mm的酸洗玻璃珠,与25ul5mm的nacl溶液混合,高速漩涡振荡5min,破碎细胞。
(5)将上述细胞破碎液在12000rpm离心5min,将上清转移至干净的1.5ml离心管中。
(6)在(5)离心后获得的上清中加入等体积的苯酚/氯仿/异戊醇(25:24:1),振荡混匀,12000rpm离心5min后,把上清转移至干净的离心管中。
(7)在(6)离心后获得的上清中加入2倍体积预冷的无水乙醇,-80℃静置30min,沉淀基因组dna,4℃,12000rpm离心10min。
(8)弃上清,沉淀用预冷的70%乙醇洗涤2次,室温放置5min使乙醇挥发,用50μlte缓冲液溶解dna沉淀。
(9)加入1μl10mg/mlrnasea,在37℃温育30min,消化rna后,-20℃保存。
1.2pcr扩增目标基因条带
以絮凝酵母spsc01基因组dna作为模板扩增基因yol032w。
引物序列如下:
pcr反应体系如下(50μl):
pcr反应程序设置:
pcr结束后,产物用琼脂糖凝胶电泳检测片断大小,,与ncbi数据库中序列长度一致(图1)。
1.3目标片段酶切
将pcr产物用dna纯化试剂盒进行纯化,然后酶切。酶切反应体系如下:
37℃温育反应3h。
凝胶回收试剂盒回收目的dna片段,经检测后于-20℃冻存备用。
1.4目的基因与载体连接(25μl)
将制备好的目的片段与酶切回收后的载体片段进行连接。
反应体系如下:
在连接仪中反应时间10h,反应温度为16℃(构建的载体见图2)。
1.5连接产物转化大肠杆菌dh5α
1.5.1大肠杆菌感受态细胞制备
(1)取-80℃保存的大肠杆菌dh5α200μl接种于10mllb液体培养基中,37℃,200rpm培养过夜;
(2)按1:100的比例转接过夜培养的菌液至50ml新鲜lb液体培养基中,37℃,200rpm培养3-4h,至od600约为0.5;
(3)将菌液转入在冰上预冷的50ml离心管中,冰上放置30min,4℃,4000rpm离心5min;
(4)弃上清,用预冷的0.1mcacl2溶液15ml悬浮细胞,冰上放置30min,4℃,4000rpm离心5min,弃去上清;
(5)重复上述步骤;
(6)加入预冷的0.1mol/lcacl2溶液2ml重悬细胞,加入2ml预冷的30%甘油,轻轻混匀,分装成200μl的小份,-80℃冻存。
1.5.2连接产物的转化
(1)从-80℃冰箱中取200μl感受态细胞,冰上融化;
(2)将10μl连接产物加入感受态细胞中,用移液枪轻轻混匀,冰上放置30min;
(3)将感受态细胞置于42℃水浴锅中热激90s,然后迅速置于冰浴中冷却2min;
(4)向管中加入800μllb液体培养基(不含抗生素),混匀后37℃,200rpm振荡培养1h;
(5)将震荡培养后的菌体4000rpm离心5min,弃部分上清,将剩余溶液混匀后涂布于含相应抗生素的筛选平板上(氨苄青霉素终浓度100μg/ml),正面向上放置半小时,待菌液完全晾干后倒置培养皿,37℃培养12-16h;
(6)挑阳性菌落进行鉴定。
1.5.3转化子质粒提取
使用omeg质粒提取试剂盒提取转化子质粒(e.z.n.aplasmiddnaminikitiprotocol-spinprotocol)。
1.5.4pcr鉴定阳性转化子
将提取的质粒作为模板进行pcr验证,以验证转化子。按照1.2中方法扩增目的基因,进行pcr验证。
pcr反应结束后,产物用琼脂糖凝胶电泳检测基因大小是否正确。
1.6重组质粒的线性化
因为重组质粒需要线性化后才能转入酵母中,进行整合表达,采用noti限制性内切酶对质粒进行酶切。酶切反应体系如下:
37℃温育反应10h。
凝胶回收试剂盒回收目的dna片段,检测无误后于-20℃冻存备用。
实施例2:磷脂合成蛋白基因yol032w的酿酒酵母电转化方法
2.1酿酒酵母电转化感受态细胞的制备
(1)酵母接种ypd培养基,30℃,150rpm培养12-14h,然后转接入新的ypd培养基(1%接种)培养6h;
(2)将培养瓶放在冰上至少15min,让菌停止生长。将50ml离心管,超纯水,1m的山梨醇溶液放在冰上预冷,整个实验过程中保持低温状态;
(3)将离心后收集到的菌体用等体积的超纯水将菌体轻轻混匀(上下颠倒晃动,不要用移液枪吹打),3000rpm,5min,4℃离心收集菌体,弃上清,重复此步骤2次;
(4)用20ml预冷的1m山梨醇溶液洗涤菌体沉淀2次,最后用0.5ml,1m山梨醇溶液重悬酵母细胞,即为酵母感受态细胞。
2.2电转法获得重组酵母转化子
(1)将感受态细胞分装至1.5ml离心管中,加入10μl线性化目的片段,用移液枪轻轻混匀,放置冰上;
(2)将100μl感受态细胞和线性化目的片段混合液加入0.2cm的电转杯中,冰浴5-10min;
(3)设置酵母参数“fungi”,“sc02”,点击pulse;
(4)取出电转杯,迅速加入1ml冰浴的山梨醇,用枪轻轻吹打,再转移至1.5ml灭菌离心管中,30℃静置,孵育5h;
(5)3000rpm离心5min,去部分上清,将剩余溶液混匀后涂布于含有300μg/mlg418的ypd平板上。
(6)将平板于30℃培养箱培养,至酵母转化子克隆长出。
(7)随机挑取生长状态较好的单菌落接入ypd液体培养基中(含100μg/ml的g418抗生素),获得一定量的菌体进行基因组dna提取(方法与1.1相同),以获得的基因组为模板扩增验证片段(引物如下),验证(方法与1.2相同)目的基因的整合情况,验证片段包含部分目的基因yol032w和部分抗性基因g418,全长1000bp。并进一步通过琼脂糖凝胶电泳验证(图3)。
验证片段引物如下:
实施例3:重组酵母s288c-yol032w总rna提取及目的基因实时定量分析重组菌株基因的表达情况
(1)将重组空载酵母s288c-ho对照菌株和重组酿酒酵母s288c-yol032w接种到含有100ml种子培养基(30g/l葡萄糖,3g/l蛋白胨,4g/l酵母浸粉)的250ml摇瓶中,30℃,150rpm,培养24h;
(2)分别取菌液,测其在620nm处吸光值,然后用种子培养基将菌液od调节至1.5;
(3)将调好od的菌液以10%(v/v)的接种量接入发酵培养基中(100g/l葡萄糖,3g/l蛋白胨,4g/l酵母浸粉,ph4.5),在30℃,150rpm条件下进行培养;
(4)菌株发酵到12h(对数生长期)时,分别取4ml菌液离心弃上清液,细胞用无菌水洗涤2次,菌体迅速用液氮冷冻,-80℃保存,提取rna备用;
(5)采用tiangenrnasimple总rna提取试剂盒提取重组菌s288c-yol032w和对照菌s288c-ho对数生长期的总rna;
(6)参照takaraprimescripttmrtreagentkitwithgdnaeraser试剂盒方法将提取的rna反转录为cdna;real-timepcr参照
结果:在不添加任何抑制物情况下重组菌株s288c-yol032w基因yol032w的表达量是对照菌株表达量的1.6倍。表明基因yol032w在重组菌中实现了过表达。
实施例4:重组空载酵母s288c-ho对照菌株和重组酿酒酵母s288c-yol032w在含有不同抑制物胁迫因素下的平板生长比较
(1)将重组空载酵母s288c-ho对照菌株和重组酿酒酵母s288c-yol032w接入含有100ml种子培养基(30g/l葡萄糖,3g/l蛋白胨,4g/l酵母浸粉)的250ml摇瓶中,30℃,150rpm,过夜培养;
(2)取过夜培养的菌液,以10%(v/v)的接种量分别转接到种子培养基中(同步骤1),培养12h;
(3)取上述菌液,测其在620nm处吸光值,用灭菌后的种子培养基调节od值至1.0;
(4)将上述菌液分别进行5倍梯度稀释;
(5)取2μl梯度稀释过的菌液依次点样于分别含10g/l乙酸,1.5g/l糠醛,1g/l香草醛和不添加抑制物的ypd平板上(20g/l葡萄糖,20g/l蛋白胨,10g/l酵母浸粉,15g/l琼脂粉);
(6)于30℃培养箱中静置培养,待长出清晰菌落后进行观察分析并拍照。
结果:从图4中看出,在正常培养条件下(培养36h)两株菌种s288c-ho和s288c-yol032w的生长状态一致,说明过表达磷脂合成蛋白基因yol032w对菌株生长没有影响。
当培养基中添加10g/l的乙酸(培养了36h),其它条件不变时,菌株s288c-yol032w在5-5稀释梯度仍有菌落生长,而对照菌株s288c-ho在5-2稀释梯度基本不能生长,说明yol032w基因过表达后提高了菌株的乙酸耐受性。同样,s288c-yol032w菌株在含有1.5g/l糠醛及1g/l香草醛的固体培养基中生长均明显优于对照菌株s288c-ho,说明yol032w基因过表达提高了菌株对糠醛和香草醛的耐受性。
实施例5:重组空载的对照酵母s288c-ho和重组酿酒酵母s288c-yol032w在不同抑制物胁迫下的乙醇发酵情况比较
5.1重组空载的对照酵母s288c-ho和重组酿酒酵母s288c-yol032w在含10g/l乙酸的发酵培养基中的生长和乙醇发酵情况
为了验证重组酿酒酵母的发酵性能,考察了重组菌株在含有高浓度乙酸发酵培养基中的生长和乙醇发酵性能。
(1)分别将对照酵母菌株s288c-ho和重组酿酒酵母s288c-yol032w接入含有100ml种子培养基(30g/l葡萄糖,3g/l蛋白胨,4g/l酵母浸粉)的250ml摇瓶中,30℃,150rpm,培养24h;
(2)分别取菌液,测其在620nm处吸光值,然后用种子培养基调节菌液od至1.5;
(3)将上述菌液以10%(v/v)的接种量接入发酵培养基中(100g/l葡萄糖,3g/l蛋白胨,4g/l酵母浸粉,添加10g/l乙酸,ph4.5),在30℃,150rpm进行乙醇发酵实验。
在含空载对照酵母s288c-ho和重组酿酒酵母s288c-yol032w乙醇发酵过程中定时取样,分别取一定量菌液用酶标仪测定吸光值来确定菌体密度(图5a);剩余的菌液离心后取上清液通过气相色谱仪检测乙醇浓度(图5b),dns法检测发酵液中剩余葡萄糖浓度(图5c),当发酵液中葡萄糖的浓度低于1g/l时定义为乙醇发酵结束。
结果:从图5a看出,随着乙醇发酵的进行,从12h开始,过表达菌株s288c-yol032w的od值快速增加,在18h时od值达到1.8左右,是对照组s288c-ho的3倍,相比于对照菌株s288c-ho,表明过表达菌株s288c-yol032w在含有10g/l乙酸的发酵培养基中适应性较强,能够快速进入对数期的。
根据图5b、5c所示,过表达菌株s288c-yol032w的葡萄糖消耗速度及乙醇浓度均高于对照菌株s288c-ho。在发酵36h时,s288c-yol032w已消耗完所有的葡萄糖,比对照菌株s288c-ho提前了12h。在24h时,重组菌株乙醇浓度达到31.5g/l,是对照菌株的3.7倍。表明过表达磷脂合成蛋白基因yol032w的菌株s288c-yol032w在乙醇发酵过程中提高了对乙酸的耐受性。
5.2重组空载的酵母s288c-ho对照菌株和重组酿酒酵母s288c-yol032w在含2g/l香草醛发酵培养基中的乙醇发酵情况
为了验证重组酿酒酵母的发酵性能,考察了重组菌株在含有高浓度香草醛发酵培养基的发酵性能。
(1)分别将对照酵母菌株s288c-ho和重组酿酒酵母s288c-yol032w接入含有100ml种子培养基(30g/l葡萄糖,3g/l蛋白胨,4g/l酵母浸粉)的250ml摇瓶中,30℃,150rpm,培养24h;
(2)分别取菌液,测其在620nm处吸光值od,然后用种子培养基调节菌液od至1.5;
(3)以10%(v/v)的接种量上述菌液接入发酵培养基中(100g/l葡萄糖,3g/l蛋白胨,4g/l酵母浸粉,添加2g/l香草醛,ph4.5),在30℃,150rpm进行乙醇发酵。
在空载酵母s288c-ho和重组酿酒酵母s288c-yol032w乙醇发酵过程中定时取样,分别取一定量菌液用酶标仪测定吸光值来确定菌体密度(图6a);剩余的菌液离心后取上清液通过气相色谱仪检测乙醇浓度(图6b),dns法检测发酵液剩余葡萄糖浓度(图6c),当发酵液中葡萄糖的浓度低于1g/l时定义为发酵结束。
结果:从图6a看出,在乙醇发酵过程中,从12h开始,过表达菌株s288c-yol032w的od值快速增加,至36h时od值达到最大,比对照菌株提前了30h。表明过表达菌株s288c-yol032w在含有2g/l香草醛的发酵培养基中菌体生长明显优于对照菌株s288c-ho。
根据图6b、6c所示,过表达菌株s288c-yol032w的葡萄糖消耗速度及乙醇产率均高于对照菌株s288c-ho。在发酵48h时,s288c-yol032w消耗完所有的葡萄糖,比对照菌株s288c-ho提前了32h。重组菌株s288c-yol032w的乙醇最大浓度比对照菌株s288c-ho提高了76%,发酵时间缩短了44h。表明过表达磷脂合成蛋白基因yol032w的重组酿酒酵母s288c-yol032w在乙醇发酵过程中提高了对香草醛的耐受性。
5.3重组空载的对照酵母s288c-ho和重组酿酒酵母s288c-yol032w在含3g/l糠醛的发酵培养基中的乙醇发酵情况
为了验证重组酿酒酵母的乙醇发酵性能,考察了重组菌株在含有高浓度糠醛发酵培养基的发酵性能。
(1)分别将对照酵母菌株s288c-ho和重组酿酒酵母s288c-yol032w接入含有100ml种子培养基(30g/l葡萄糖,3g/l蛋白胨,4g/l酵母浸粉)的250ml摇瓶中,30℃,150rpm,培养24h;
(2)分别取一定量菌液,测其在620nm处吸光值od,并用种子培养基调节菌液od至1.5;
(3)以10%(v/v)的接种量将上诉菌液接入发酵培养基中(100g/l葡萄糖,3g/l蛋白胨,4g/l酵母浸粉,添加3g/l糠醛,ph4.5),在30℃,150rpm进行乙醇发酵实验。
在空载酵母s288c-ho和重组酿酒酵母s288c-yol032w乙醇发酵过程中定时取样,分别取部分菌液用酶标仪测定吸光值来确定菌体的浓度(图7a);剩余的菌液离心后取上清液通过气相色谱仪检测乙醇的浓度(图7b),dns法检测发酵液中剩余葡萄糖浓度(图7c),当发酵液中葡萄糖的浓度低于1g/l时定义为发酵结束。
结果:从图7a看出,在乙醇发酵过程中,从12h开始,过表达菌株s288c-yol032w的od值快速增加,在36h时od值达到最大,比对照菌株提高了42.9%,生长时间提前了24h。表明过表达菌株s288c-yol032w在含有3g/l糠醛的发酵培养基中菌体生长优于对照菌株s288c-ho。
根据图7b、7c所示,过表达菌株s288c-yol032w的葡萄糖消耗速度及乙醇产率均高于对照菌株s288c-ho。在乙醇发酵过程中,重组菌株s288c-yol032w达到最大乙醇浓度的时间为48h,发酵时间比对照菌株缩短了24h,最高乙醇浓度比对照菌株提高了20%,葡萄糖消耗时间提前了44h。表明过表达磷脂合成蛋白基因yol032w的重组酿酒酵母s288c-yol032w在乙醇发酵过程中明显提高了对糠醛的耐受性。
<110>大连理工大学
<120>一株多种抑制物胁迫耐受性酿酒酵母及制备方法、应用
<160>1
<210>1
<211>741
<212>dna
<400>1
atgtttgctgcaattgcctcaggaaatccattgcaactatcggtagaagtccccaattcaaatgggttac
agcatacaattgtcctttctcgaacgaaaccgaaactatactcgcatataacactgtttatattacctaa
tgttacctttcctcaggattatatagctacagtctatttcaaacttagccctcaggaagaatttaaacta
tttgggtacctaagcagtgagaaacccagtgccatattcaaagtgcaaataccgagctcgaagaaagacg
ctggagatacaagtgatggattaggtgagatagacatggatgtagatgatggttcgggggccgctgatcc
gtttactgacaccaatggaagtagcagtaataacatatcagaattgattattggtatttccattgagcct
cgagaacaaggaatgatgaaattagaagagtggaaggccagcatgaatgccgaagcacaaaagaacaact
cattgatcttgtcaagacctaacttaggaataattcgaaatattaccacggcagggcagttggcacaagt
ttacccttcattgacacaagaattggccgccaaaatcgttcagcatgcatataactatttatcagggttc
ctagatgctcagggaaacgtgcccatcaagaggtttgatacatggtgggacaaatttagaaatagactgg
ccaacgacgggacgtttttggatgaagtaactaaaaattag
<110>大连理工大学
<120>一株多种抑制物胁迫耐受性酿酒酵母及制备方法、应用
<160>2
<210>1
<211>246
<212>氨基酸
<400>1
mfaaiasgnplqlsaevpnsnglqhtivlsrtkpklyshitlfilpnvtfpqdfiatvyfklspqeefkl
fgylsgekpsaifkvqipsskketgdvsdglgeidmdvddgsgttdtfndnngnsnnniseliigisiep
reqgmlkleewkasmnaeaqknnslvlsrpnlgiirnittagqlaqvyptltqelaakivqhaynylsgf
ldaqgnvpikkfdtwwekfrnrlandgtfldevtkn
- 还没有人留言评论。精彩留言会获得点赞!