基因LOC_Os05g38680在增加水稻有效分蘖数中的应用的制作方法
本发明属于植物分子遗传领域,具体涉及基因在增加水稻有效分蘖数中的应用。
背景技术:
水稻的产量性状主要由单位面积有效穗数、每穗总粒数以及千粒重等基本因素组成,直接决定水稻单位面积的产量。水稻的有效穗数与分蘖数直接相关。分蘖是水稻等禾本科植物在地面以下或接近地面处所发生的分枝,能抽穗结实的分蘖称为有效分蘖,不能抽穗或抽穗而不结实的称为无效分蘖,水稻的有效分蘖数直接决定了有效穗数。
目前,研究表明水稻的分蘖受多个基因调控,其中转录因子moc1及其互作蛋白tad1是调控水稻分蘖的主控因子,调控分蘖芽的起始、生长与发育等过程。研究发现,水稻moc1突变体只产生单个分蘖,而突变体tad1则分蘖数显著性增多;tad1作用于moc1的上游,与moc1形成蛋白复合体,通过泛素化26s蛋白酶体途径介导moc1的降解,调控水稻分蘖数。
201711469003x的发明《水稻高光合效率的基因及其应用》告知了该基因能用于促进水稻的光合作用,具体为促进水稻叶片中叶绿素a(chla)与叶绿素b(chlb)增加。
技术实现要素:
本发明要解决的技术问题是提供一种基因在增加水稻有效分蘖数中的应用。
为了解决上述技术问题,本发明提供了基因在增加水稻有效分蘖数中的应用,该基因的核苷酸序列如seqidno:1所示。
在水稻中过表达该基因,能显著性增加水稻有效分蘖数。
本发明的技术方案具体如下:
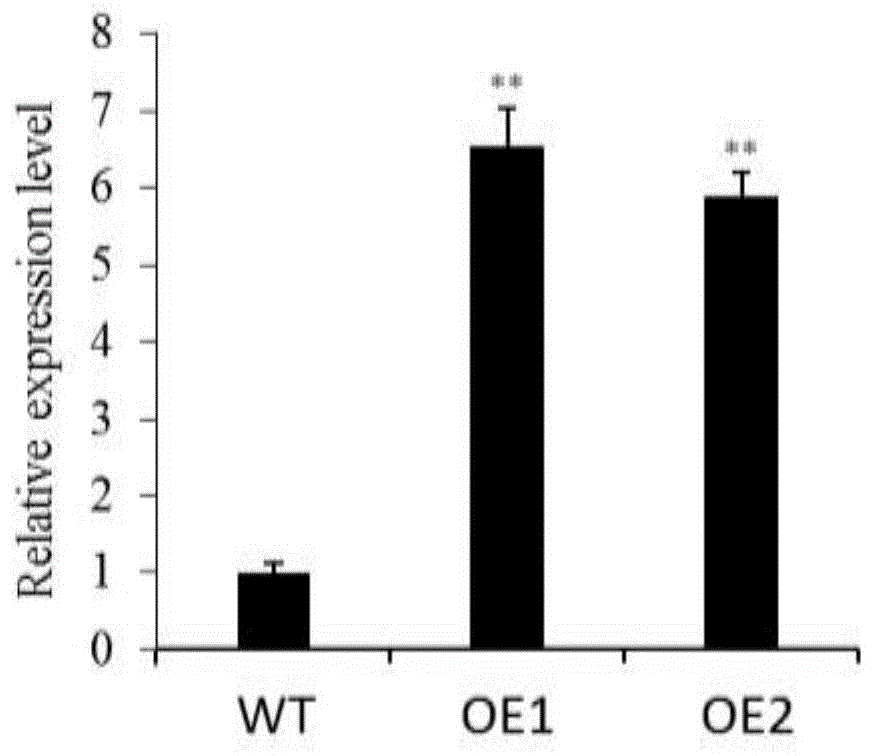
本发明从水稻cdna中pcr扩增了loc_os05g38680基因(seqidno:1),构建该基因的过表达载体,并利用农杆菌介导法,将该载体遗传转化到野生型水稻品种日本晴中,鉴定出两个阳性的过表达株系oe1与oe2,利用real-timepcr检测这两个株系中loc_os05g38680基因的表达量,结果其表达两量均显著高于野生型对照日本晴(图1)。在水稻成熟期,这两个过表达株系的有效分蘖数均显著高于野生型对照日本晴(图2,图3),说明在水稻中增加loc_os05g38680基因的表达水平,可有效促进水稻的有效分蘖数。
虽然,201711469003x的发明《水稻高光合效率的基因及其应用》告知了该基因能用于促进水稻的光合作用,但是按照本行业的常识,促进水稻的光合作用带来的性能优势一般为净光合速率的提高,从而提高水稻产量,其与增加水稻有效分蘖数并没有必然的关系。
现有的参与水稻光合作用过程的基因pgl11、coi1b对植株分蘖数目无明显影响,且参与调控水稻分蘖发育相关基因如moc1、moc2、tad1、lax1、lax2、d53与d14等基因对水稻叶片的光合效率也无明显影响。因此,201711469003x的发明《水稻高光合效率的基因及其应用》并不能提供本发明以技术启示。
综上所述,本发明在水稻中新鉴定了一个与moc1互作的蛋白,由基因loc_os05g38680编码;在水稻中过量表达基因loc_os05g38680能显著性增加水稻的有效分蘖数,在水稻高产育种中具有潜在的应用价值。
附图说明
下面结合附图对本发明的具体实施方式作进一步详细说明。
图1为水稻loc_os05g38680基因过表达株系中loc_os05g38680基因的表达量分析。
图2为水稻loc_os05g38680基因过表达株系的植株表型。
图3为水稻loc_os05g38680基因过表达株系的有效分蘖数统计。
图中wt为野生型对照日本晴;oe1与oe2分别为loc_os05g38680基因两个过表达株系。*表示t检验存在显著性(p<0.05)差异;**表示t检验存在极显著性(p<0.01)差异。
具体实施方式
步骤1、水稻总rna提取与cdna的合成
采用rneasyplantminikit(qiagen,德国),按产品说明提取水稻野生型品种日本晴叶片的总rna;并用primescripttm1ststrandcdnasynthesiskit(takara,日本)进行反转录成cdna,方法按照产品说明操作。
步骤2、loc_os05g38680基因的pcr扩增
采用
步骤3、loc_os05g38680基因的pcr产物的酶切与纯合
首先,用axypreppcr清洁试剂盒(axygen,美国)对pcr产物进行纯化,方法按照产品说明操作;其次,采用takara公司(日本)的限制性内切酶kpni和psti进行双酶切。酶切反应体系为:kpni与psti各1μl、3μlbufferm(takara公司限制性内切酶bufferm)、10μlpcr产物、ddh2o补足至20μl,37℃酶切4h;最后,再axypreppcr清洁试剂盒(axygen,美国)对切酶产物进行纯化,方法按照产品说明操作。
步骤4、pcambia-1300-2×35s载体的酶切与纯合
按照步骤3的方法,对pcambia-1300-2×35s载体质粒进行双酶切和纯化。
步骤5、loc_os05g38680基因过表达载体的构建
①,采用promega公司(美国)的t4连接酶试剂盒,将步骤3和步骤4获得的纯化产物进行连接,连接体系为:1μl酶切后的载体质粒、2μl目的基因酶切片段、0.5μlt4连接酶、1μlbuffer、ddh2o补足至10μl,4℃过夜(12小时)。
②,取步骤①所得的10μl连接产物与jm109感受态细胞冰上共孵育30min,42℃热激90sec,立即冰浴10min;加入800μl无抗生素的lb培养基后37℃恒温振荡培养1h后,将菌液均匀涂布于含kan(25mg/l)抗性的固体平板培养基上,37℃过夜。
③,挑取平板上长出的单克隆菌落,于4ml含kan(25mg/l)抗性的lb液体培养基中,置于试管中,37℃200rpm摇床培养过夜。用2×taqpcr预混试剂(天根,北京)对菌液进行pcr验证,反应体系:1μl菌液、10μl2×taqpcrmastermixⅱ、1μf1+r1引物(10μm),ddh2o补足至20μl。pcr扩增程序为:94℃预变性5min;94℃变性30sec,55℃退火30sec,72℃延伸30sec,35个循环;72℃延伸5min。反应结束后取5μl反应产物,琼脂糖凝胶电泳检测。
④、将pcr鉴定的阳性克隆菌落寄到生物技术公司,用pcambia-1300的通用引物p1:5’-ccaggctttacactttatgc-3’和p2:5’-gcgattaagttgggtaacgc-3’测序,将插入基因序列全部测通;从而获得loc_os05g38680基因过表达载体。
即,该loc_os05g38680基因过表达载体中含有seqidno:1所示的核苷酸序列。
步骤6、水稻遗传转化
将以上构建好的过表达载体,用nishimura等的方法(nishimuraetal,natprotoc,2006)转化水稻野生型品种日本晴,获得转基因水稻植株。
步骤7、转基因水稻的鉴定
按照步骤1的方法提取转基因植株叶片的总rna并合成cdna。用loc_os05g38680基因的pcr引物f2:5’-ggagcttgctgatccagtagtc-3’,r2:5’-tagctagagagctcatgtgaagag-3’,对loc_os05g38680基因的表达进行real-timepcr分析。使用takara公司的
通过real-timepcr分析,在转基因植株中鉴定出两个loc_os05g38680基因高表达株系oe1和oe2,其表达量均显著高于对照日本晴(图1)。
步骤8、转基因水稻的有效分蘖数统计
在金华水稻大田中,5月初播种,6月初插秧,进行常规的水稻。在水稻成熟期(同年的10月),分别随机选取20株oe1转基因株系、oe2转基因株系及其野生型对照日本晴,进行有效分蘖数统计,统计结果采用t-test方法分析转基因植株与野生型对照之间的显著性差异。
所得结果为oe1和oe2转基因株系的有效分蘖数均显著多于野生型对照植株(图2,图3)。
最后,还需要注意的是,以上列举的仅是本发明的若干个具体实施例。显然,本发明不限于以上实施例,还可以有许多变形。本领域的普通技术人员能从本发明公开的内容直接导出或联想到的所有变形,均应认为是本发明的保护范围。
序列表
<110>浙江师范大学
<120>基因loc_os05g38680在增加水稻有效分蘖数中的应用
<160>1
<170>siposequencelisting1.0
<210>1
<211>309
<212>dna
<213>水稻(oryzasativa)
<400>1
atgggagactcatcatcctcagcttcgtacatcagaatggttcaccacctgatagagaag60
tgcatttgcttcaacctgaacaaggaggagtgcatggaggccctggagaagcatgccaac120
atcaaccctgtcgtcacttccacagtatggaaggagctggagaaggagaacaaggagttc180
ttcgagacctacaacaaggacagggcggagcgcaacatcgaggcggagacgatgcagcgg240
atccagaagatgctctccgacgccgcggcatccaagggctccgacgacgacgacgacgac300
gagagctag309
- 还没有人留言评论。精彩留言会获得点赞!