环状RNAhsa_circ_0004872在胃癌诊断中的应用的制作方法
本发明属于分子生物学领域,具体涉及环状rnahsa_circ_0004872在胃癌诊断中的应用。
背景技术:
胃癌是一种常见的消化系统恶性肿瘤,属于全球第五大常见癌症之一,其死亡率高居全球癌症死亡率第三位,仅2018年新增胃癌患者就超过100万,估测死于胃癌的患者超过78.3万。传统的胃癌治疗方式是手术治疗,同时以放疗或化疗进行辅助治疗。但由于胃癌发生发展涉及多个基因的异常表达,缺乏有效的早期诊断特征,胃癌患者被诊断时多为晚期,使得胃癌患者五年生存率差,胃癌转移患者中位生存期不足一年。因此探寻胃癌发生发展的分子机制,寻找有效的分子诊断标志物对于提高胃癌患者的生存率是极为重要的。
环状rna(circularrna,circrna)是一种在真核细胞中大量表达的非编码rna,不具有5’端帽子结构和3’端poly(a)尾巴结构,并且5’端和3’端以共价键首尾相接而形成的环形结构rna。环状rna在基因表达及调控中发挥着非常重要作用。研究发现,circrna大量存在于自然界生物中,具有丰富性、保守性及结构稳定性等特征。circrna的以上特性,使其具有作为肿瘤诊断标志物和应用治疗的潜能。
目前研究较多的circrna功能是作为“mirna海绵”。mirna是一类长度约为22-23个核苷酸的单链微小rna。mirna可以通过结合到靶基因mrna的3’utr区,抑制靶基因的翻译或者介导其降解过程,从而发挥负向调控其表达的功能。mirna可调控人类约30%的基因。一个基因可能受多个mirna的调控,同时一个mirna可以调控多个靶基因的表达,从而形成复杂的mirna调控网络。在癌症的发生发展中,circrna作为“mirna海绵”结合mirna,使得功能性mirna减少,抑制了mirna负向调控其靶基因的功能,引起基因的异常表达,最终导致或改变生理病理过程。
技术实现要素:
为解决现有技术上环状rna利用的不足,本发明提供了一种环状rnahsa_circ_0004872在胃癌诊断中的应用。
本发明是通过以下技术方案实现的:
我们通过环状rna二代测序技术寻找候选circrna分子,检测其在胃癌组织及癌旁组织中的表达,确定目的circrna分子hsa_circ_0004872,并确定hsa_circ_0004872可以作为mir-224-5p的“分子海绵”来抑制细胞中mir-224-5p的水平,从而影响胃癌细胞的增殖、侵袭和转移能力,并初步探讨调控hsa_circ_0004872表达的分子机制,为胃癌的早期诊断提供新的分子标志物。
本发明提供了一种环状rna,所述环状rnahsa_circ_0004872的基因的核苷酸序列如seqidno.1所示,所述环状rnahsa_circ_0004872来自22号染色体mapk1基因的第2、3、4外显子反向剪切,mapk1pre-mrna第4外显子3’端ag碱基与第2外显子5’端ct碱基相连,形成的共价闭合环状rna。
本发明提供了所述环状rna及其基因相关产物在胃癌诊断中的应用。
所述环状rna可以作为胃癌诊断标志物在制备胃癌诊断试剂中应用。
所述环状rna基因片段或基因的剪切体可以作为胃癌诊断标志物在制备胃癌诊断试剂中应用。
所述环状rna基因编码的蛋白质或多肽片段可以作为胃癌诊断标志物在制备胃癌诊断试剂中应用。
所述环状rna基因编码的蛋白抗体可以作为胃癌诊断标志物在制备胃癌诊断试剂中应用。
所述环状rna的类似物或环状rna的抑制物可以作为胃癌诊断标志物在制备胃癌诊断试剂中应用。
所述环状rna作为“mirna海绵”所吸附的mirnas作为胃癌诊断标志物在制备胃癌诊断试剂中的应用。
本发明的有益效果是:hsa_circ_0004872在胃癌组织中的表达水平比癌旁正常组织显著降低,hsa_circ_0004872的低表达可显著促进胃癌细胞的增殖、侵袭转移能力。hsa_circ_0004872的这种作用是通过作为mir-224-5p“分子海绵”抑制mir-224-5p实现的,mir-224-5p可靶向smad4和p21的3’utr,抑制二者表达,从而调节胃癌细胞的增殖、侵袭和转移。胃癌细胞中,hsa_circ_0004872表达的下调是受腺苷脱氨酶adar1的抑制。而smad4又可作为转录因子直接结合在adar1启动子区抑制adar1的表达,从而形成一个负反馈调控环路,使hsa_circ_0004872介导的调控信号不断放大。因此hsa_circ_0004872可作为胃癌诊断的潜在标志物。
附图说明
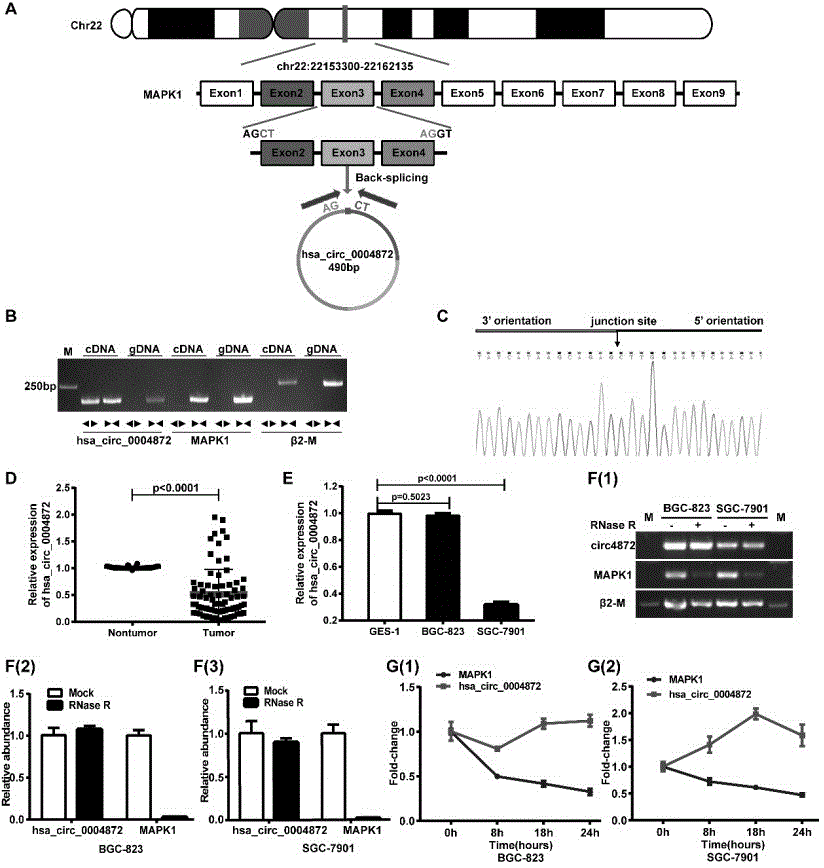
图1是hsa_circ_0004872的形成和结构鉴定。其中,(a)模式图显示hsa_circ_0004872由基因mapk1的第2、3、4外显子反向剪接形成。(b)扩散引物(divergentprimers)和收敛引物(convergentprimers)分别以胃癌细胞bgc-823的gdna和cdna为模板,pcr扩增β2-m、mapk1、hsa_circ_0004872。(c)hsa_circ_0004872剪切位点sanger测序鉴定(d)hsa_circ_0004872在胃癌组织(tumor)和癌旁组织(nontumor)的表达(下降比例66/77=85.7%)。(e)hsa_circ_0004872在胃永生上皮细胞及胃癌不同细胞株中的表达。(f)(1)为bgc-823、sgc-7901中rnaser消化后,一般pcr后琼脂糖凝胶电泳结果。(2)、(3)为qrt-pcr后量化分析结果。(g)(1)、(2)分别是bgc-823、sgc-7901中actd处理,qrt-pcr检测细胞中hsa_circ_0004872的含量。
图2是过表达hsa_circ_0004872对胃癌细胞增殖、侵袭和转移的影响。其中,(a)hsa_circ_0004872的过表达载体plcdh-circ-4872转染胃癌细胞bgc-823、sgc-7901后的过表达效率的验证。(b)plcdh-circ-4872分别转染bgc-823、sgc-7901,edu实验检测细胞增殖能力。(c)edu实验量化分析图。(d)plcdh-circ-4872分别转染bgc-823、sgc-7901,cck8实验检测细胞增殖能力。(e)plcdh-circ-4872分别转染bgc-823、sgc-7901,划痕实验检测细胞转移能力。(f)划痕实验量化分析图。(g)plcdh-circ-4872分别转染bgc-823、sgc-7901,transwell实验检测细胞的转移(migration)和侵袭(invasion)能力。(h)transwell实验量化分析图。
图3是干扰hsa_circ_0004872对胃癌细胞增殖、侵袭和转移的影响。其中,(a)hsa_circ_0004872sirna位点示意图。(b)在bgc-823中转染hsa_circ_0004872sirna后,qrt-pcr检测hsa_circ_0004872与mapk1的表达水平。结果证实了hsa_circ_0004872sirna的特异性。(c)向bgc-823细胞中转染hsa_circ_0004872sirna,edu实验检测细胞的增殖能力。(d)edu实验量化分析图。(e)向bgc-823细胞中转染hsa_circ_0004872sirna,划痕实验检测细胞的转移能力。(f)划痕实验量化分析图。(g)bgc-823中转染hsa_circ_0004872sirna,transwell实验检测细胞的转移(migration)和侵袭(invasion)能力。(h)transwell实验量化分析图。
图4是hsa_circ_0004872作为mir-224-5p的“海绵作用”。其中,(a)fish实验显示hsa_circ_0004872主要存在于细胞质中。(b)rip实验显示ago2抗体免疫沉淀物可以扩增出hsa_circ_0004872。(c)不同数据库预测与hsa_circ_0004872存有潜在结合位点的mirnas。(d)向bgc-823、sgc-7901细胞中分别转染hsa_circ_0004872过表达载体,qrt-pcr检测网站预测存有潜在结合位点的mirnas的表达水平。(e)向bgc-823细胞中转染hsa_circ_0004872sirna,qrt-pcr检测网站预测存有潜在结合位点的mirnas的表达水平。(f)fish实验显示hsa_circ_0004872与mir-224-5p可发生共定位。(g)双荧光素酶载体pmirglo-circ4872-wt/mut构建示意图(wt为野生型,mut为突变型)。(h)bgc-823、sgc-7901共转染mir-224-5pmimics与pmirglo-circ4872-wt/mut,双荧光素酶相对活性检测结果。(i)hsa_circ_0004872在胃癌组织(tumor)和癌旁组织(nontumor)的表达(上升比例29/39=74.36%)。(j)胃癌组织统计学分析,mir-224-5p与hsa_circ_0004872存在负相关关系(n=32)。
图5是mir-224-5pmimics对胃癌细胞增殖、侵袭和转移的影响。其中,(a)向bgc-823、sgc-7901细胞中转染mir-224-5pmimics,mir-224-5p的过表达效率验证。(b)向bgc-823、sgc-7901细胞中转染mir-224-5pmimics,edu实验检测细胞的增殖能力。(c)edu实验量化分析图(d)bgc-823、sgc-7901细胞中转染mir-224-5pmimics,cck8实验检测细胞的增殖能力。(e)bgc-823、sgc-7901中转染mir-224-5pmimics,划痕实验检测细胞的转移能力。(f)划痕实验量化分析图。(g)bgc-823、sgc-7901中转染mir-224-5pmimics,transwell实验检测细胞的转移(migration)和侵袭(invasion)能力。(h)transwell实验量化分析图。
图6是mir-224-5pinhibitor对胃癌细胞增殖、侵袭和转移的影响。其中,(a)向bgc-823、sgc-7901中转染mir-224-5pinhibitor,mir-224-5p的干扰效率。(b)bgc-823、sgc-7901中转染mir-224-5pinhibitor,edu实验检测细胞的增殖能力。(c)edu实验量化分析图。(d)bgc-823、sgc-7901中转染mir-224-5pinhibitor,cck8实验检测细胞的增殖能力。(e)bgc-823、sgc-7901中转染mir-224-5pinhibitor,划痕实验检测细胞的转移能力。(f)划痕实验量化分析图。(g)bgc-823、sgc-7901中转染mir-224-5pinhibitor,transwell实验检测细胞的转移(migration)和侵袭(invasion)能力。(h)transwell实验量化分析图。
图7是mir-224-5p调控胃癌细胞增殖、侵袭和转移的机制。其中,(a)bgc-823、sgc-7901中分别转染mir-224-5pmimics与inhibitor,72h后westernblot检测pten、hoxd10、p21和smad4的表达。(b)sgc-7901中分别转染mir-224-5pmimics与inhibitor,72h后qrt-pcr检测p21和smad4的表达。(c)双荧光素酶载体pmir-smad4-3’utr-wt/mut构建示意图(wt为野生型,mut1为突变潜在结合位点a,mut2为突变潜在结合位点b,mut为突变潜在结合位点ab)。(d)mir-224-5pmimics与pmir-smad4-3’utr-wt/mut及prl-tk质粒共转染bgc-823、sgc-7901细胞,双荧光素酶相对活性检测结果。(e)双荧光素酶载体pmir-p21-3’utr-wt/mut构建示意图(wt为野生型,mut为突变型)。(f)向bgc-823、sgc-7901中共转染mir-224-5pmimics与pmir-p21-3’utr-wt/mut及prl-tk质粒,双荧光素酶相对活性检测结果。
图8是hsa_circ_0004872/mir-224-5p/smad4(p21)调控胃癌细胞增殖、侵袭和转移,其中,(a)向sgc-7901细胞中共转染hsa_circ_0004872与mir-224-5pmimics,edu实验检测细胞的增殖能力。(b)edu实验量化分析图。(c)向bgc-823、sgc-7901细胞中共转染hsa_circ_0004872与mir-224-5pmimics,cck8实验检测细胞的增殖能力。(d)bgc-823、sgc-7901中共转染hsa_circ_0004872与mir-224-5pmimics,划痕实验检测细胞的转移能力,(e)划痕实验量化分析图。(f)bgc-823、sgc-7901中共转染hsa_circ_0004872与mir-224-5pmimics,transwell实验检测细胞的转移(migration)和侵袭(invasion)能力。(g)transwell实验量化分析图。(h)bgc-823、sgc-7901中共转染hsa_circ_0004872与mir-224-5pmimics,72h后westernblot实验检测smad4和p21的表达。
图9是hsa_circ_0004872抑制胃癌细胞在裸鼠体内增殖、侵袭和转移。其中,(a)裸鼠皮下注射稳转细胞bgc-82326天后,颈部脱臼法处死裸鼠,裸鼠皮下成瘤情况。(b)皮下肿瘤he染色照片。(c)裸鼠皮下瘤体积增长曲线。(d)裸鼠皮下瘤质量对应图。(e)qrt-pcr检测裸鼠皮下瘤hsa_circ_0004872的表达含量。(f)qrt-pcr检测裸鼠皮下瘤mir-224-5p的表达含量。(g)裸鼠尾静脉注射稳转细胞bgc-823一个月后,颈部脱臼法处死裸鼠,裸鼠肺照片及he染色。(h)裸鼠尾静脉注射稳转细胞bgc-823,肺部重量统计图。(i)裸鼠尾静脉注射稳转细胞bgc-823,肺部肿瘤转移灶统计图。
图10是胃癌细胞中adar1调控hsa_circ_0004872的表达,其中,(a)bgc-823、sgc-7901中过表达adar1-p110、adar1-p150后,westernblot检测adar1的表达情况。(b)bgc-823、sgc-7901中过表达adar1-p110、adar1-p150后,qrt-pcr检测adar1的表达情况。(c)bgc-823、sgc-7901中过表达adar1-p110、adar1-p150后,qrt-pcr检测hsa_circ_0004872的表达情况。(d)bgc-823、sgc-7901中转染adar1小干扰后,westernblot检测adar1的表达情况。(e)bgc-823、sgc-7901中转染adar1小干扰后,qrt-pcr检测adar1的表达情况。(f)bgc-823、sgc-7901中转染adar1小干扰后,qrt-pcr检测hsa_circ_0004872的表达情况。(g)ncbigeo数据库(gse27342),adar1在胃癌组织(tumor)和癌旁组织(nontumor)的表达(上升比例59/80=73.75%)。(h)kaplan-meierplotter数据库(201786_s_at)分析胃癌病人adar1的表达对生存期的影响。(i)kaplan-meierplotter数据库(201786_s_at)分析有淋巴结转移的胃癌病人adar1的表达对生存期的影响。
图11是胃癌细胞中smad4调控adar1的表达。其中,(a)bgc-823、sgc-7901过表达smad4,westernblot检测adar1的表达情况。(b)bgc-823、sgc-7901过表达smad4,qrt-pcr检测adar1的表达情况。(c)bgc-823中过表达smad4,smad4标签蛋白flag抗体进行免疫沉淀,一般pcr扩增预测结合位点片段。(d)双荧光素酶载体pgl3-adar1-promoter-wt/mut构建示意图(wt为野生型,mut为突变潜在结合位点)。(e)bgc-823中共转染smad4与pgl3-adar1-promoter-wt/mut,双荧光素酶相对活性检测结果。(f)ncbigeo数据库(gse27342),smad4在胃癌组织(tumor)和癌旁组织(nontumor)的表达(下降比例42/75=56%)。(g)ncbigeo数据库(gse27342)胃癌组织统计学分析,adar1与smad4存在负相关关系(n=63)。(h)kaplan-meierplotter数据库(202527_s_at)分析胃癌病人smad4的表达对生存期的影响。(i)kaplan-meierplotter数据库(202527_s_at)分析有淋巴结转移的胃癌病人smad4的表达对生存期的影响。
图12是胃癌细胞smad4/adar1/hsa_circ_0004872/mir-224-5p/smad4(p21)环路调控机制图。
具体实施方式
实施例1hsa_circ_0004872在胃癌中的表达
1.hsa_circ_0004872是mapk1第2、3、4外显子反向剪接形成的环状rna
我们利用环状rna二代测序技术筛选了在胃癌及相应癌旁组织中表达差异的环状rna,选择其中差异表达明显的hsa_circ_0004872作为我们的候选研究分子。通过对新鲜的胃癌组织及对应癌旁组织进行qrt-pcr检测,发现hsa_circ_0004872在胃癌组织中的表达显著下调(85.7%p<0.0001),因此,我们选择hsa_circ_0004872(序列见表1,序列编号seqidno.1)作为我们的研究对象。
表1hsa_circ_0004872序列表
hsa_circ_0004872(hsa_circ_27008)来自22号染色体mapk1基因的第2、3、4外显子反向剪切,mapk1pre-mrna第4外显子3’端ag碱基与第2外显子5’端ct碱基相连,形成的共价闭合环状rna(图1a)。
2.hsa_circ_0004872在胃癌组织中表达显著降低
由于hsa_circ_0004872是mapk1的部分外显子首尾相接而形成的闭合环状结构,其剪切位点处的碱基序列与mapk1碱基序列不同,因此,为验证我们设计的pcr引物仅用于扩增hsa_circ_0004872而非线性的mapk1,我们设计了包含剪切位点的扩散引物(divergentprimers)和不包括剪接位点的收敛引物(convergentprimers)。通过收敛引物和扩散引物对胃癌细胞cdna和gdna进行pcr和琼脂糖凝胶电泳,发现hsa_circ_0004872扩散引物只有以cdna为模板时可以扩增出条带,而以gdna为模板时扩增不出条带,(cdna中既有环状结构又有线性结构,gdna中只有线性结构),而收敛引物以cdna和gdna为模板均可以扩增出条带。线性内参基因β2-m及亲本基因mapk1以cdna和gdna为模板均仅收敛引物可以扩增出条带(图1b)。此外,为进一步证实我们的扩散引物扩增出的片段为环状rna,我们对扩散引物的pcr产物进行了sanger测序,发现mapk1第4外显子3’端ag与第2外显子5’端ct碱基确实相连(图1c),以上结果表明,hsa_circ_0004872是mapk1转录后通过第2、3、4外显子反向剪切形成的环状rna,且扩散型引物可被用来特异检测hsa_circ_0004872。
然后我们用qrt-pcr检测了hsa_circ_0004872在胃癌及相应的癌旁组织中的表达,发现其在胃癌组织中的表达水平显著下降(66/77,85.7%,p<0.0001)(图1d)。
我们进一步通过qrt-pcr检测了hsa_circ_0004872在胃上皮永生化细胞ges-1和胃癌细胞bgc-823、sgc-7901中的表达,结果发现:与ges-1相比,hsa_circ_0004872在sgc-7901中表达含量较低(图1e)。因此,后续研究中hsa_circ_0004872高表达实验在bgc-823及sgc-7901中进行,而干扰实验在bgc-823细胞中进行。
3.hsa_circ_0004872的稳定性检测
为检测hsa_circ_0004872是否较线性rna分子具有更高的稳定性,我们用rnaser处理rna,rnaser是一类mg2+依赖的3’→5’外切核糖核酸酶,可以消化线性rna分子,但不消化具有套索结构和环状结构的rna。用rnaser消化bgc-823和sgc-7901细胞rna后,用一般pcr及qrt-pcr进行扩增,我们发现hsa_circ_0004872可以耐受rnaser的消化能力,而线性分子β2-m及mapk1则对rnaser不耐受(图1f)。
放线菌素d(actd)可以抑制rna的合成。通过对bgc-823及sgc-7901细胞施加一系列时间梯度的actd,我们发现mapk1mrna的形成明显受到抑制,而hsa_circ_0004872的形成则不受抑制(图1g)。以上两个实验均说明hsa_circ_0004872比其亲本基因mapk1具有更高的稳定性。
实施例2hsa_circ_0004872对胃癌细胞增殖、侵袭和转移的影响,可显著抑制胃癌细胞增殖
1.过表达hsa_circ_0004872对胃癌细胞增殖、侵袭和转移的影响
为研究hsa_circ_0004872对胃癌细胞生物学效应的影响,我们构建了hsa_circ_0004872过表达载体plcdh-circ-4872。将其与对照载体分别转染bgc-823及sgc-7901,qrt-pcr检测结果表明,转染过表达质粒后,胃癌细胞中hsa_circ_0004872的表达水平明显提高(图2a)。
转染hsa_circ_0004872过表达质粒后,通过edu实验及cck8实验检测细胞增殖效率,发现过表达hsa_circ_0004872,胃癌细胞的增殖能力明显受到抑制(图2b-d)。通过划痕实验及transwell实验(有无基质胶),我们发现胃癌细胞的侵袭转移能力受到抑制(图2e-h)。
2.hsa_circ_0004872干扰对胃癌细胞增殖、侵袭和转移的影响
为研究hsa_circ_0004872低表达对胃癌细胞生物学效应的影响,我们设计了两条跨剪切位点的小干扰rna(序列见表2)。在bgc-823中转染si-circ4872后,我们发现hsa_circ_0004872的表达水平明显降低,而亲本基因mapk1的表达水平则没有明显变化(图3a-b)。
表2本研究所涉及干扰序列
通过edu实验检测细胞增殖效率,发现干扰hsa_circ_0004872的表达,细胞的增殖明显得到促进(图3c-d)。划痕实验及transwell实验(有无基质胶)结果表明细胞的侵袭转移受到促进(图3e-h)。
实施例3hsa_circ_0004872抑制胃癌细胞增殖、侵袭转移的分子机制
1.hsa_circ_0004872在胃癌细胞中的亚细胞定位
由于环状rna的功能通常与其在细胞中的不同定位有关,目前研究显示,由外显子形成的环状rna通常存在于细胞质中。为检测hsa_circ_0004872在胃癌细胞中的定位,我们进行了荧光原位杂交(fish)实验。我们设计带有cy3红色荧光标记的hsa_circ_0004872探针(序列见表3)。待胃癌细胞生长于细胞爬片上后,固定并通透细胞,cy3红色荧光标记探针与细胞爬片杂交过夜,dapi染色,封片后于激光共聚焦显微镜下拍照。实验结果表明:hsa_circ_0004872在胃癌细胞中主要存在于细胞质(图4a),这与目前大多数的文献报道相吻合。
表3本研究所涉及探针序列
据文献报道,细胞质中的环状rna通常通过结合mirnas或者结合蛋白质来行使功能。我们猜想hsa_circ_0004872可能通过结合mirna或者结合蛋白质从而影响其功能。我们首先探寻hsa_circ_0004872是否具有结合mirna的功能。argonaute2(ago2)蛋白可以与成熟的mirna结合,是形成rna诱导沉默复合体(risc)的重要组成部分。通过rna结合蛋白免疫沉淀实验(rip),发现ago2抗体进行免疫沉淀可以扩增出hsa_circ_0004872条带。说明hsa_circ_0004872可能具有结合mirna的功能(图4b)。
2.软件预测hsa_circ_0004872可能结合的mirnas
通过targetscan、circmir和starbase三个生物学软件预测hsa_circ_0004872可能存在结合位点的mirnas,将该三个数据库取交集(图4c),发现存在4个mirnas,分别是hsa-mir-224-5p、hsa-mir-654-3p、hsa-mir-513a-5p、hsa-mir-599;targetscan和circmir两个数据库取交集,发现存在10种mirnas;circmir和starbase两个数据库取交集,发现存在30种mirnas,具体mirnas见附表4。我们查阅文献寻找交集中已报道的具有调控细胞增殖、侵袭转移等生物学功能的mirnas,最终选择hsa-mir-224-5p、hsa-mir-421、hsa-mir-487a-5p、hsa-mir-616-3p作为我们候选研究的mirnas。通过在bgc-823及sgc-7901细胞中过表达hsa_circ_0004872,发现只有hsa-mir-224-5p(以下简称为mir-224-5p)明显受hsa_circ_0004872负调控,其他mirnas的表达水平并没有显著影响(图4d)。为进一步探究hsa_circ_0004872对mir-224-5p是否具有调控作用,我们在hsa_circ_0004872表达含量高的胃癌细胞株bgc-823中干扰hsa_circ_0004872的表达,发现hsa_circ_0004872可以上调mir-224-5p的表达(图4e)。这提示hsa_circ_0004872可能通过“mirna海绵作用”调控mir-224-5p的表达。
表4生物学信息学软件预测has_circ_0004872可能具有结合位点的mirnas
3.hsa_circ_0004872作为mir-224-5p的“分子海绵”来发挥调控作用
为了检测hsa_circ_0004872能否与mir-224-5p发生共定位,我们首先利用荧光原位杂交(fish)实验对mir-224-5p在胃癌细胞中的分布进行了检测,我们设计带有fitc绿色荧光标记的mir-224-5p探针及cy3红色荧光标记的hsa_circ_0004872探针(序列见表3)。待胃癌细胞生长于细胞爬片上,固定并通透细胞,探针与细胞爬片杂交过夜,dapi染色,封片后于激光共聚焦显微镜下拍照。发现mir-224-5p与hsa_circ_0004872共同定位在细胞质中(图4f)。
利用targetscan软件寻找hsa_circ_0004872碱基序列上mir-224-5p潜在的结合位点,构建了包含hsa_circ_0004872部分碱基序列的野生型荧光素酶载体(pmirglo-circ4872-wt)及突变型荧光素酶载体(pmirglo-circ4872-mut)。通过在bgc-823中同时转染mir-224-5pmimics与hsa_circ_0004872野生型荧光素酶载体,发现双荧光素酶相对活性显著下调,而同时转染mir-224-5pmimics与hsa_circ_0004872突变型荧光素酶载体,双荧光素酶相对活性则基本不发生变化(图4g-h)。以上实验说明hsa_circ_0004872能与mir-224-5p的结合,hsa_circ_0004872通过“mirna海绵作用”调控mir-224-5p的表达。
标本的相关性分析:我们在39对新鲜胃癌组织及对应癌旁组织中利用qrt-pcr检测了mir-224-5p的表达,发现其在胃癌组织中是明显上调的(29/39,74.36%,p=0.0098)。统计学分析显示,mir-224-5p与hsa_circ_0004872在此32对组织中呈负相关关系(p=0.0183)(图4i-j)。
实施例4mir-224-5p的生物学功能及调控机制
1.mir-224-5p对胃癌细胞增殖、侵袭和转移的影响
基于以上实验结论,我们接下来对mir-224-5p在胃癌细胞中的生物学功能进行了检测。通过在bgc-823及sgc-7901细胞中转染mir-224-5p的化学模拟物mimics及抑制剂inhibitor,qrt-pcr验证其表达效率(图5a,图6a),edu实验及cck8实验发现,上调mir-224-5p的表达可以促进胃癌细胞的增殖,降低mir-224-5p的表达可以抑制胃癌细胞的增殖(图5b-d,图6b-d);划痕实验及transwell实验(有无基质胶)发现上调mir-224-5p的表达可以促进胃癌细胞的侵袭转移能力,降低mir-224-5p的表达可以抑制胃癌细胞的侵袭转移能力(图5e-h,图6e-h)。
2.mir-224-5p调控胃癌细胞增殖、侵袭和转移的机制
为研究mir-224-5p调控胃癌细胞增殖、侵袭和转移的机制,我们通过生物学网站及文献检索发现mir-224-5p可与smad4、p21、hoxd10、pten等多种蛋白存在潜在相互作用。因此,我们探寻mir-224-5p在胃癌细胞中是否可以通过调控以上蛋白的表达而发挥作用。
通过在bgc-823及sgc-7901细胞中转染mir-224-5p的化学模拟物mimics及inhibitor,westernblot发现mir-224-5p在胃癌细胞中可以显著下调smad4及p21的蛋白表达,而对hoxd10、pten没有明显的调控作用(图7a)。qrt-pcr结果证明sgc-7901中转染mir-224-5pmimics后可以显著降低smad4及p21的mrna水平;sgc-7901中转染mir-224-5pinhibitor后可以显著升高smad4及p21的mrna(图7b)。
为进一步探寻mir-224-5p在胃癌细胞中是否可以通过直接靶向smad4的3’utr区域。我们构建包含两个潜在结合位点的smad43’utr区域靶向序列的野生型荧光素酶报告质粒(pmir-smad4-3’utr-wt)以及突变1个潜在结合位点的荧光素酶报告质粒(pmir-smad4-3’utr-mut1、pmir-smad4-3’utr-mut2)及两个潜在结合位点共同突变的荧光素酶报告质粒(pmir-smad4-3’utr-mut)。在bgc-823、sgc-7901中共转染mir-224-5pmimics以及smad4野生型或者突变型荧光素酶报告质粒结果显示:过表达mir-224-5p可以显著降低smad4野生型荧光素酶报告质粒的荧光素酶活性,smad4一个潜在结合位点突变型的荧光素酶活性轻微降低,而两个潜在结合位点突变型的荧光素酶活性完全不受影响(图7c-d)。
同时我们还构建包含p213’utr区域靶向序列的野生型荧光素酶报告质粒载体(pmir-p21-3’utr-wt)以及突变型荧光素酶报告质粒载体(pmir-p21-3’utr-mut)。在bgc-823、sgc-7901中共转染mir-224-5pmimics以及p21野生型或者突变型荧光素酶报告质粒结果显示:过表达mir-224-5p可以显著降低p21野生型荧光素酶报告质粒的荧光素酶活性,p21突变型的荧光素酶活性则不受影响(图7e-f)。以上双荧光素酶活性实验结果显示:在胃癌细胞中,mir-224-5p可以直接靶向smad4、p21的3’utr区域。
实施例5.hsa_circ_0004872/mir-224-5p/smad4(p21)调控胃癌细胞增殖、侵袭转移
为进一步检测hsa_circ_0004872是否通过mir-224-5p发挥功能,我们进行了回复实验,在胃癌细胞bgc-823、sgc-7901中共转染hsa_circ_0004872过表达质粒及mir-224-5pmimics,通过edu实验、cck8实验、划痕实验及transwell实验(有无基质胶)发现过表达mir-224-5p可以部分回复hsa_circ_0004872对细胞增殖、侵袭和转移的抑制作用(图8a-f)。westernblot实验同样显示,高表达hsa_circ_0004872引起的smad4及p21的表达升高可以通过过表达mir-224-5p得到部分回复(图8g)。以上实验显示hsa_circ_0004872通过hsa_circ_0004872/mir-224-5p/smad4(p21)通路调控胃癌细胞增殖、侵袭和转移。
实施例6hsa_circ_0004872抑制胃癌细胞在裸鼠体内增殖、侵袭和转移
为进一步检测hsa_circ_0004872对体内胃癌细胞增殖、侵袭和转移的影响,我们分别进行了裸鼠皮下注射实验和裸鼠尾静脉注射实验。
1.hsa_circ_0004872抑制胃癌细胞在裸鼠体内增殖
我们将稳转hsa_circ_0004872的bgc-823细胞及对照细胞分别注射到裸鼠的背部两侧(左侧为对照组,右侧为过表达组)。接种一周后测量肿瘤的直径计算肿瘤的体积,绘制生长曲线发现,过表达hsa_circ_0004872组肿瘤生长速度明显慢于对照组(图9c)。荷瘤二十天后,处死裸鼠,取出肿瘤,发现过表达hsa_circ_0004872组肿瘤体积和质量明显小于对照组(图9a,d)。取部分肿瘤组织进行he染色,发现形成的肿瘤均为实体瘤(图9b),通过对肿瘤组织进行qrt-pcr,发现过表达hsa_circ_0004872组其hsa_circ_0004872的表达明显高于对照组,而mir-224-5p的表达明显低于对照组(图9e-f)。以上实验进一步说明hsa_circ_0004872过表达后,胃癌细胞在裸鼠皮下的成瘤能力下降。
2.hsa_circ_0004872抑制胃癌细胞在裸鼠体内侵袭转移
我们将稳转hsa_circ_0004872的bgc-823细胞、对照细胞及pbs分别注射到裸鼠尾静脉中。待其体重减轻,精神萎靡时,处死裸鼠,记录裸鼠肺部质量及转移灶的数目,结果显示过表达hsa_circ_0004872组裸鼠的肺比对照细胞组体积小,重量轻,转移灶少,he染色也验证了这一现象(图9g-i)。以上实验进一步说明在体内hsa_circ_0004872高表达后胃癌细胞在肺的定植能力受到显著抑制。
实施例7hsa_circ_0004872在胃癌细胞中表达下调的分子机制
经典的环状rna形成模型理论-内含子驱动的环化理论认为外显子环化依赖于其相近的内含子序列,内含子包含的大量反向重复序列(如alu序列)可以通过碱基之间的反向互补配对形成套索结构,从而拉近剪切位点、切除内含子序列,外显子得以成环。而88%的环状rna侧翼的内含子中含有大量的alu重复序列。两个反向的alu序列之间可以形成双链dna,此双链结构可以作为腺苷脱氨酶的反应位置。腺苷脱氨酶1(adenosinedeaminaseactingonrna,adar1)是一类可将腺苷a经脱氨水解转化为肌苷i的rna编辑酶,使得外显子首尾之间无法闭合为环状rna。
我们在bgc-823、sgc-7901中分别转染adar1过表达质粒(pmgfp-adar1-p110及pmgfp-adar1-p150)后进行qrt-pcr,发现hsa_circ_0004872的表达明显降低,同时在bgc-823、sgc-7901中转染adar1的小干扰rna,发现hsa_circ_0004872的表达显著上调(图10a-f)。以上实验说明adar1在胃癌细胞中可以调控hsa_circ_0004872的表达。据文献报道,几乎所有的胃癌组织中都会出现adar1异常调控引起的rna错误编辑表型。因此我们利用ncbigeo数据库(gse27342)分析adar1在胃癌组织与癌旁组织中的表达情况(图10g),结果发现adar1在胃癌组织中的表达明显高于对应癌旁组织(59/80,73.75%),这与文献报道的结果一致。利用kaplan-meierplotter数据库(201786_s_at)分析胃癌病人adar1的表达对其生存期的影响发现,adar1高表达的病人,具有较低的生存期(图10h)。对有淋巴结转移的胃癌病人进行生存曲线分析发现,adar1高表达的病人,同样具有较低的生存期(图10i)。
通过数据库检索发现adar1启动子区存在smad4的结合位点,而hsa_circ_0004872可以通过mir-224-5p调控smad4的表达,因此我们猜想smad4在胃癌细胞中是否可以通过作为adar1的转录因子调控其表达进而形成smad4/adar1/hsa_circ_0004872/smad4的调控环路。为了验证我们的猜想,我们首先在bgc-823、sgc-7901中分别转染smad4过表达质粒(p-3×flag-smad4)及对照质粒(p-3×flag-cmv-14),发现过表达smad4后adar1的蛋白水平及mrna水平表达均明显降低(图11a-b)。说明smad4可以调控adar1的表达。
通过生物学网站jaspar发现,adar1的启动子区存在五处smad4潜在的结合位点,分别是-1993到-1981、-1959到-1947、-1590到-1578、-1280到-1268及-760到-748。我们采用染色质免疫共沉淀(chip)实验探究smad4能否结合在adar1启动子区。通过在bgc-823细胞中转染smad4过表达质粒及其对照质粒,用smad4标签蛋白flag抗体进行免疫沉淀,利用这五处潜在结合位点的引物进行扩增,结果发现只有转染smad4过表达载体的bgc-823细胞加入flag抗体后其-1280到-1268处可以扩增出条带(图11c)。该实验说明smad4可以结合在adar1启动子区的-1280到-1268处。
为研究这种结合是否是功能性结合,我们构建了包含该处结合位点的adar1启动子区的野生型双荧光素酶载体(pgl3-adar1-promoter-wt)以及该结合位点突变的荧光素酶报告质粒(pgl3-adar1-promoter-mut)(图11d)。通过在bgc-823共转染smad4过表达载体以及adar1启动子区野生型或突变型荧光素酶报告质粒结果显示:过表达smad4可以显著降低adar1启动子区野生型荧光素酶报告质粒的荧光素酶活性,adar1启动子区潜在结合位点突变型的荧光素酶活性不变(图11e)。
我们利用公共数据库ncbigeo(gse27342)分析了smad4的表达情况,发现smad4在胃癌组织中的表达是下调的(42/75,56%,p=0.0249)(图11f),统计学分析显示,adar1与smad4具有负相关关系(p=0.0119)(图11g)
通过kaplan-meierplotter数据库(202527_s_at)分析胃癌病人smad4的表达对生存期的影响,发现smad4低表达的胃癌患者生存期低(图11h)。对有淋巴结转移的胃癌病人进行生存曲线分析,同样验证了以上结论(图11i)。
以上所述仅是本专利的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本专利技术原理的前提下,还可以做出若干改进和替换,这些改进和替换也应视为本专利的保护范围。
序列表
<110>山东大学
<120>环状rnahsa_circ_0004872在胃癌诊断中的应用
<160>1
<170>siposequencelisting1.0
<210>1
<211>490
<212>dna
<213>人类(homosapiens)
<400>1
ctctgcttatgataatgtcaacaaagttcgagtagctatcaagaaaatcagcccctttga60
gcaccagacctactgccagagaaccctgagggagataaaaatcttactgcgcttcagaca120
tgagaacatcattggaatcaatgacattattcgagcaccaaccatcgagcaaatgaaaga180
tgtatatatagtacaggacctcatggaaacagatctttacaagctcttgaagacacaaca240
cctcagcaatgaccatatctgctattttctctaccagatcctcagagggttaaaatatat300
ccattcagctaacgttctgcaccgtgacctcaagccttccaacctgctgctcaacaccac360
ctgtgatctcaagatctgtgactttggcctggcccgtgttgcagatccagaccatgatca420
cacagggttcctgacagaatatgtggccacacgttggtacagggctccagaaattatgtt480
gaattccaag490
- 还没有人留言评论。精彩留言会获得点赞!