一种增强型合成Notch受体及其应用的制作方法
本发明涉及生物技术领域,具体为一种增强型合成notch受体、其编码的核苷酸、重组表达载体和宿主细胞,以及应用。
背景技术:
notch信号通路在生物体中普遍存在,是在进化过程中高度保守的信号通道,能够介导细胞之间的相互作用,从而调节组织和器官的发育及分化,调控机体细胞的增殖、分化和凋亡。
notch受体是一类i型跨膜蛋白,由notch细胞外结构域(notchextracellulardomain,necd)、notch核心调节结构域(notchcoreregulatoryregion,nc)、notch细胞内结构域(notchintracellulardomain,nicd)组成。necd的结构十分庞大,由数十余个表皮生长因子样重复序列(epidermalgrowthfactor-likerepeats,egfs)组成。nicd包含rbpj-associatedmodule(ram)、锚蛋白重复序列(ankyrinrepeats,anks)、c末端反式激活结构域(c-terminaltrans-activationdomain,tad)和pest结构域(pestdomain)。
nc横跨细胞膜,其由n端胞外段较长的负调节结构域(negativeregulatoryregion,nrr)和跨膜结构域(transmembranedomain,tmd)组成。nrr部分又可以进一步细分为3个lin12/notchrepeats(lnrs)和一个近膜部分的异二聚体结构域(heterodimerizationdomain,hd)。hd区域在notch受体成熟过程中被furin蛋白酶在s1位点处水解,将hd分为n端(hd-n')和c端(hd-c'),后再被转运到细胞膜表面。notch激活所涉及的s2位点位于hd-c'并受到lnr-a/b保护,s3位于细胞膜内部。新的核磁研究表明紧邻tmd的ram结构域能够在细胞膜内表面形成折返结构(deatherage,c.l.,etal.structuralandbiochemicaldifferencesbetweenthenotchandtheamyloidprecursorproteintransmembranedomains.scienceadvances3,e1602794(2017).)。
notch受体的激活以一种直接而简单的机制进行。notch受体的necd在和配体(如delta)结合后通过机械力变构效应,lnrs被展开,暴露s2位点,使其能够被整联蛋白和金属蛋白酶(adisintegrinandmetalloproteinase,adam)切割,进而导致γ分泌酶(γ-secretase)进一步切割s3位点后释放nicd入核调控下游信号。
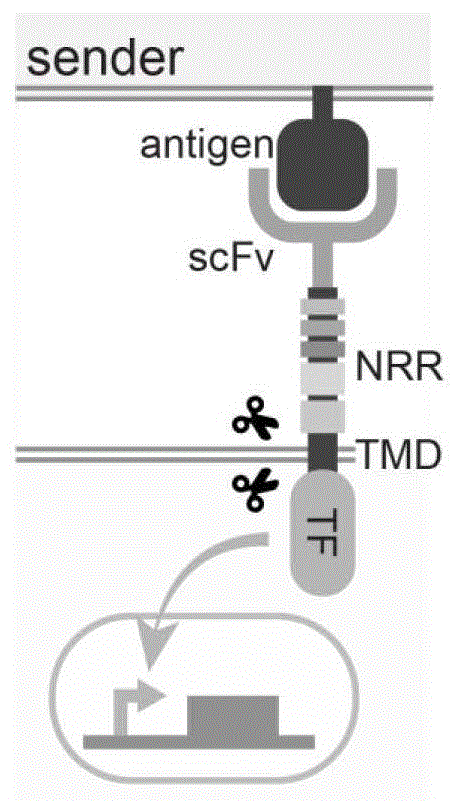
由于notch非常直接和简单的信号转导机制,gordon(gordon,w.r.,etal.mechanicalallostery:evidenceforaforcerequirementintheproteolyticactivationofnotch.devcell33,729-736(2015).)和morsut(morsut,l.,etal.engineeringcustomizedcellsensingandresponsebehaviorsusingsyntheticnotchreceptors.cell164,780-791(2016).)等人已经分别证明,通过使用单链抗体(singlechainvariablefirmament,scfv)和转录因子(transcriptionfactor,tf)分别替换天然notch的necd和nicd,而保留nc,可以模块化地构建syntheticnotchreceptor(synnotch)。synnotch是高度模块化的合成膜受体,由细胞外识别结构域(如scfv),notch核心调节结构域(notchcoreregulatoryregion,nc)和转录因子串联而成(transcriptionfactor,tf)(图1)。在原始synnotch构建时,ram结构域的功能尚未被解析,因此原始synnotch构建中并没有包含ram结构域。
尽管目前尚不完全清楚,但synnotch受体可能和天然notch受体一样,能够以一种直接而简单的机制被激活。当scfv识别并结合靶细胞的膜表达抗原(antigen,ag),在机械力变构效应的作用下,nc中的nrr的lnrs区域被展开,暴露s2位点,使其能够被adam切割,导致nrr区域脱落。接下来,γ分泌酶将进一步切割位于tmd内部的s3位点,后释放细胞内tf入核调控下游信号。
synnotch已经被证明具有利用于细胞免疫疗法治疗癌症(roybal,k.t.,etal.precisiontumorrecognitionbytcellswithcombinatorialantigen-sensingcircuits.cell164,770-779(2016).,roybal,k.t.,etal.engineeringtcellswithcustomizedtherapeuticresponseprogramsusingsyntheticnotchreceptors.cell167,419-432e416(2016).)的潜力。然而,在我们的测试中发现,现有synnotch存在着高水平的配体非依赖性激活(ligand-independentactivation,lia),导致其在没有ag激活的情况下也能够被错误地激活而产生显著的错误信号。这种配体非依赖性激活或错误激活将严重限制synnotch被稳健而安全地应用于基础和临床研究。因此,开发一种具有低lia或错误激活的新型synnotch十分重要。
我们的研究发现synnotch的lia或错误激活产生的关键原因是由于ram结构域的缺失所致。通过添加最小ram序列,我们构建了几乎完全消除了lia或错误激活的新型(增强型)synnotch。
技术实现要素:
本发明旨在提供一种降低或者消除配体非依赖性激活或错误激活的合成notch受体。
提供了一种降低或消除合成notch受体配体非依赖性激活或错误激活的方法。
本发明涉及一种增强型合成notch受体(enhancedsyntheticnotchreceptor,esnotch),从n末端到c末端依次包括:
a.特异性识别抗原相关结构的细胞外识别结构域;
b.包含蛋白水解切割位点的notch核心调节结构域(notchcoreregulatoryregion,nc);
c.ram结构域;
d.细胞内转录调节结构域;
所述的ram结构域为源自天然ram结构域的、长度为5~100个氨基酸的片段,或者,所述的ram结构域为长度为5~100个氨基酸的人工序列片段;优选的,所述的天然ram结构域为人或小鼠来源的ram结构域;
所述的ram结构域含有至少一个或多个由疏水性氨基酸任意组合的二氨基酸对;所述的疏水性氨基酸选自色氨酸(trp,w)、苯丙氨酸(phe,f)、亮氨酸(leu,l)、异亮氨酸(ile,i)、缬氨酸(val,v)、甲硫氨酸(met,m)、半胱氨酸(cys,c)或酪氨酸(tyr,y);
优选的,所述的ram结构域长度为5~40个氨基酸,更优选为5~10个氨基酸;
优选的,所述ram结构域为含有三个连续的疏水性氨基酸的片段。
优选的,所述的ram结构域含有以下氨基酸序列之一(如seqidno.1~10所示):
seqidno.1qhgqlwf
seqidno.2qhgqlwfp
seqidno.3khgslwl
seqidno.4khgflwl
seqidno.5ehstlwf
seqidno.6ehgalwl
seqidno.7qhgqlfw
seqidno.8qhgqlww
seqidno.9qhgqlff
seqidno.10qhgqlll
进一步,所述的ram结构域序列如上述氨基酸序列seqidno.1~10所示;或者,ram结构域是n端为上述序列、c末端带有任意氨基酸、长度为8~40的氨基酸片段,优选的,长度为8~10个氨基酸。
notch核心调节结构域的蛋白水解切割位点包括一个或多个选自s1、s2或s3蛋白酶裂解位点中的配体诱导型蛋白酶裂解位点。其中,s1蛋白酶裂解位点为弗林蛋白酶型蛋白酶裂解位点;s2蛋白酶裂解位点为adam型蛋白酶裂解位点;s3蛋白酶裂解位点为γ-分泌酶型蛋白酶裂解位点。
notch核心调节结构域来源于人或小鼠的天然notch核心调节结构域,可为具有至少75%~100%氨基酸序列同一性的来自人notch1-4、小鼠notch1-4的notch核心调节结构域的氨基酸序列;优选的可为具有至少75%~100%氨基酸序列同一性的来自人notch1、小鼠notch1的notch核心调节结构域的氨基酸序列。
notch核心调节结构域中包含负调节结构域(negativeregulatoryregion,nrr)和跨膜结构域(transmembranedomain,tmd)。
所述的细胞外识别结构域所识别的对应特异性识别抗原相关结构域可为:抗体特异性结合的肿瘤特异性抗原、肿瘤相关抗原、自身免疫疾病相关抗原、病原体相关抗原、疾病相关抗原或细胞外基质组分。
所述的细胞内转录调节结构域包含以下结构域的至少一种:dna结合结构域、转录调节结构域。
所述的dna结合结构域可选自:天然转录因子的dna结合结构域,或基于锌指蛋白、转录激活因子样效应物、特异性核酸酶构建的合成dna结合结构域。
所述的转录调节结构域可为:转录激活结构域或转录阻遏结构域。
所述的细胞内转录调节结构域中可以选择性插入一个或多个核定位信号肽。
一种核苷酸序列,用于编码上述增强型合成notch受体。
一种重组表达载体,含有上述核酸序列。
一种宿主细胞,含有上述核酸或者重组载体。所述的宿主细胞为真核细胞。进一步,所述的宿主细胞为哺乳动物细胞;进一步,所述宿主细胞为免疫细胞、上皮细胞、内皮细胞、神经细胞或干细胞。其中所述免疫细胞为t细胞、细胞毒性t细胞、辅助t细胞、调节t细胞、b细胞、自然杀伤细胞、单核细胞、树突细胞或巨噬细胞。
上述增强型合成notch受体、核苷酸序列、重组载体或宿主细胞可用于制备或者筛选治疗肿瘤、感染、自身免疫疾病或其他疾病的药物。
一种降低合成notch受体的配体非依赖性激活或者错误激活的方法,在合成notch受体的notch核心调节结构域与细胞内转录调节结构域之间插入ram结构域;所述的ram结构域含有至少一个或多个由疏水性氨基酸任意组合的二氨基酸对;所述的疏水性氨基酸选自色氨酸、苯丙氨酸、亮氨酸、异亮氨酸、缬氨酸、甲硫氨酸、半胱氨酸或酪氨酸。
优选的,所述ram结构域为含有三个连续的疏水性氨基酸、长度5~40个氨基酸的片段。
所述的ram结构域含有以下氨基酸片段之一:
seqidno.1qhgqlwf
seqidno.2qhgqlwfp
seqidno.3khgslwl
seqidno.4khgflwl
seqidno.5ehstlwf
seqidno.6ehgalwl
seqidno.7qhgqlfw
seqidno.8qhgqlww
seqidno.9qhgqlff
seqidno.10qhgqlll。
进一步,所述的ram结构域序列如上述氨基酸序列seqidno.1~10所示;或者,ram结构域是n端为上述序列、c末端带有任意氨基酸、长度为8~40的氨基酸片段,优选的,长度为8~10个氨基酸。
上述增强型合成notch受体、核苷酸序列、重组载体或宿主细胞在应用时具有显著降低的配体非依赖性激活或错误激活,并且不影响抗原结合以及正常的notch通路。
经过我们实验证明,现有的synnotch产生配体非依赖性激活或错误激活的原因是由于错误缺失了ram结构域;然而,保留过长的ram结构域将影响正常的notch通路。本发明通过对天然ram序列进行筛选和改造,确定了能够用于降低synnotch的配体非依赖性激活或错误激活的ram结构域的最短长度和序列氨基酸特征。通过添加ram序列,我们构建了几乎完全消除配体非依赖性激活或错误激活的增强型合成notch受体。
本发明的有益效果在于:
1.通过在合成notch受体的notch核心调节结构域与细胞内转录调节结构域之间插入筛选和改造的ram结构域,既不影响notch通路,又能最大限度地降低或消除配体非依赖性激活或错误激活。
2.本发明的增强型合成notch受体具有低配体非依赖性激活或错误激活,而具备更好的临床应用的安全性。
3.本发明的增强型合成notch受体可以高保真地将细胞外信号转导进入细胞内,自定义地调节各种细胞过程,能够被安全而稳健地应用于多种临床或基础研究情景;这扩展了原始合成notch受体的应用范围,为使用嵌合受体改造宿主细胞提供了新的应用前景,具有巨大的经济价值和意义。
4.本发明的增强型合成notch受体可以使用人来源的序列作为notch核心调节结构域或ram结构域因而具有更低的潜在免疫原性,进一步提升了临床应用的安全性。
附图说明
图1为抗原诱导的synnotch信号转导的示意图。
图2为用于瞬时共转染测定的质粒,用于测量synnotch的配体非依赖性激活(lia)。
图3为实施例2通过mcherry荧光测量瞬时共转染效率。
图4为实施例2lag16-mn1c-ttaa的lia实验结果;图4a为synnotch(lag16-mn1c-ttaa)的结构示意图;图4b为d2egfp的表达;图4c为每个共转染组的中值d2egfp荧光强度,其中条高为平均值,误差条为标准偏差。虚线对应于共转染0ngsynnotch时的d2egfp荧光强度的平均值。
图5为实施例2不同转染量下synnotch(lag16-mn1c-ttaa)的膜表达量。
图6为实施例2稳定表达synnotch(lag16-mn1c-ttaa)的细胞被固定后染色,mcherry和细胞外myc标签的荧光信号散点图。
图7为实施例2表达gfp的发送细胞(靶细胞)和转染了不同剂量synnotch(lag16-mn1c-ttaa)的细胞共培养实验结果;图7a为实验原理图;图7b为不同转染量下,每个synnotch共转染组在和不表达gfp或表达gfp的发送细胞(靶细胞)共培养条件下的synnotch细胞的中值d2egfp荧光强度。
图8为实施例3在synnotch(lag16-mn1c-ttaa)中插入egf重复的实验结果;图8a为插入了egf重复的synnotch(lag16-egf-mn1c-ttaa)的结构示意图;图8b为每个共转染组的中值d2egfp荧光强度,egf(-)代表图4a中的synnotch,而egf(+)代表图8a中的synnotch。
图9为实施例4将具有s1、s2或s3蛋白水解切割位点突变的synnotch共转染实验结果;图9a为具有蛋白水解切割位点突变的synnotch的结构示意图;
图9b为不同转染量下,每个共转染组的中值d2egfp荧光强度。
图10为实施例4用特异性蛋白酶抑制剂处理的结果;图10a为实验原理图;图10b和图10c分别为使用不同浓度的adam特异性抑制剂bb94和γ分泌酶特异性抑制剂化合物e孵育转染了synnotch的细胞的中值荧光强度图。
图11为实施例5具有ram结构域的esnotch(lag16-mn1c-hn1ram8-ttaa)的实验结果;图11a为该esnotch的结构示意图;图11b为共转染lag16-mn1c-ttaa或lag16-mn1c-hn1ram8-ttaa的中值d2egfp荧光强度。
图12为实施例5共转染synnotch或esnotch的细胞固定后用抗合成notch上myc标签的抗体染色的myc标签的荧光信号。
图13为实施例6具有源自人notch1不同长度ram序列的esnotch共转染后的中值d2egfp荧光强度。
图14为实施例6具有源自人notch1不同长度短ram序列的esnotch共转染后的中值d2egfp荧光强度。
图15为人和小鼠notch受体tmd附近的氨基酸序列的比对。
图16为实施例7具有不同来源ram序列的esnotch共转染细胞的中值d2egfp荧光强度。
图17为实施例8具有不同氨基酸突变的ram序列的esnotch共转染细胞的中值d2egfp荧光强度。
图18为实施例9具有或不具有nrr和/或ram的esnotch结构缺失体共转染细胞的中值d2egfp荧光强度。
图19为notch信号中的nand逻辑模块的示意图
图20为实施例10具有不同细胞外识别结构域、notch核心调节结构域和转录调节结构域的esnotch的实验结果。图20a-c为具有不同细胞外识别结构域、notch核心调节结构域和转录调节结构域的具有ram序列的esnotch的结构示意图;图20a”-c”分别对应其lia结果;图20a’-c’分别对应其抗原诱导的激活倍率。
具体实施方式
以下结合具体的实施例来对本发明的技术方案加以说明。以下实施例是对本发明的进一步说明,而不是对本发明的限制。
实施例1构建合成notch受体及其变体、及合成notch受体及其变体的重组表达载体
使用clonexpress(vazyme#c115)无缝连接dna部分,并在pgk启动子后克隆到p2z质粒骨架中。通过sanger测序(生工)验证所有构建体。所有合成notch受体及其变体的结构总结于表1中。
简而言之,通过融合scfv(lag16或αcd19),notch核心调节结构域(mn1c,egf-mn1c,hn1c或notch核心调节结构域与ram序列)和转录因子(ttaa或gv2)构建合成notch受体及其变体。所有合成notch受体及其变体含有用于膜靶向的n末端cd8α信号肽(malpvtalllplalllhaarp)和用于测定膜表面表达的myc标签(eqkliseedl)。
表1.本研究中使用的合成notch受体及其变体
缩写:mn1c,小鼠notch1核心调节结构域。hn1c,人notch1核心调节结构域。nrr,负调节结构域。tmd,跨膜结构域(相当于没有nrr的notch核心调节结构域)。
小鼠notch1核心调节结构域(mousenotch1coreregulatoryregion,mn1c):小鼠notch1核心(mn1c)根据(morsut,l.etal.engineeringcustomizedcellsensingandresponsebehaviorsusingsyntheticnotchreceptors.cell164,780-791,doi:10.1016/j.cell.2016.01.012(2016).)设计。将小鼠notch1(np_032740)的necd上的最后一次egf重复后的ile1427用作mn1c的n末端。跨膜结构域c末端的5个连续碱性氨基酸(rkrrr)的最后一个arg1752用作mn1c的c末端。
ildysftggagrdipppqieeacelpecqvdagnkvcnlqcnnhacgwdggdcslnfndpwknctqslqcwkyfsdghcdsqcnsagclfdgfdcqltegqcnplydqyckdhfsdghcdqgcnsaecewdgldcaehvperlaagtlvlvvllppdqlrnnsfhflrelshvlhtnvvfkrdaqgqqmifpyygheeelrkhpikrstvgwatssllpgtsggrqrreldpmdirgsivyleidnrqcvqsssqcfqsatdvaaflgalaslgslnipykieavksepvepplpsqlhlmyvaaaafvllffvgcgvllsrkrrr
underline,thecleavagesitesofs1,s2,s3.第225-228、284-285、315-318分别为s1,s2,s3位点。
带有蛋白水解切割位点突变的mn1c突变体:mn1c[s1],mn1c[s2],mn1c[s3]分别具有已报告的失活突变:s1,rqrr|→aaaa;s2,a|v→ed;s3,gcg|v→llff。
logeat,f.etal.thenotch1receptoriscleavedconstitutivelybyafurin-likeconvertase.procnatlacadsciusa95,8108-8112,doi:10.1073/pnas.95.14.8108(1998).
brou,c.etal.anovelproteolyticcleavageinvolvedinnotchsignaling:theroleofthedisintegrin-metalloproteasetace.molcell5,207-216,doi:doi10.1016/s1097-2765(00)80417-7(2000).
egf-mn1c:具有额外的egf重复的延长mn1c(np_032740,pro1390至arg1752)。参考(morsut,l.etal.engineeringcustomizedcellsensingandresponsebehaviorsusingsyntheticnotchreceptors.cell164,780-791,doi:10.1016/j.cell.2016.01.012(2016).)。
passpcvgsnpcynqgtceptsenpfyrclcpakfngllchildysftggagrdipppqieeacelpecqvdagnkvcnlqcnnhacgwdggdcslnfndpwknctqslqcwkyfsdghcdsqcnsagclfdgfdcqltegqcnplydqyckdhfsdghcdqgcnsaecewdgldcaehvperlaagtlvlvvllppdqlrnnsfhflrelshvlhtnvvfkrdaqgqqmifpyygheeelrkhpikrstvgwatssllpgtsggrqrreldpmdirgsivyleidnrqcvqsssqcfqsatdvaaflgalaslgslnipykieavksepvepplpsqlhlmyvaaaafvllffvgcgvllsrkrrr。
第1-41位氨基酸为ecf重复的序列。
人notch1核心调节结构域(humannotch1coreregulatoryregion,hn1c):将人notch1(np_060087)的necd上的最后一次egf重复后的ile1427用作hn1c的n末端。跨膜结构域c末端的5个连续碱性氨基酸(rkrrr)的最后一个arg1762用作hn1c的c末端。hn1c和mn1c的序列高度相似。
ildysfgggagrdippplieeacelpecqedagnkvcslqcnnhacgwdggdcslnfndpwknctqslqcwkyfsdghcdsqcnsagclfdgfdcqraegqcnplydqyckdhfsdghcdqgcnsaecewdgldcaehvperlaagtlvvvvlmppeqlrnssfhflrelsrvlhtnvvfkrdahgqqmifpyygreeelrkhpikraaegwaapdallgqvkasllpggseggrrrreldpmdvrgsivyleidnrqcvqassqcfqsatdvaaflgalaslgslnipykieavqsetveppppaqlhfmyvaaaafvllffvgcgvllsrkrrr
第1-294位为hn1c的nrr,295-336为tmd。
人notch2核心调节结构域(humannotch2coreregulatoryregion,hn2c):将人notch2(np_077719)的necd上的最后一次egf重复后的leu1413用作hn2c的n末端。跨膜结构域c末端的4个连续碱性氨基酸(krkr)的最后一个arg1704用作hn2c的c末端。
lytappstppatclsqycadkardgvcdeacnshacqwdggdcsltmenpwancssplpcwdyinnqcdelcntveclfdnfecqgnsktckydkycadhfkdnhcdqgcnseecgwdgldcaadqpenlaegtlvivvlmppeqllqdarsflralgtllhtnlrikrdsqgelmvypyygeksaamkkqrmtrrslpgeqeqevagskvfleidnrqcvqdsdhcfkntdaaaallashaiqgtlsyplvsvvsesltpertqllyllavavviilfiillgvimakrkr
小鼠notch2核心调节结构域(mousenotch2coreregulatoryregion,mn2c):将小鼠notch2(np_035058)的necd上的最后一次egf重复后的leu1413用作mn2c的n末端。跨膜结构域c末端的4个连续碱性氨基酸(krkr)的最后一个arg1706用作mn2c的c末端。
lytaptstppatcqsqycadkardgicdeacnshacqwdggdcsltmedpwanctstlrcweyinnqcdeqcntaeclfdnfecqrnsktckydkycadhfkdnhcdqgcnseecgwdgldcasdqpenlaegtliivvllppeqllqdsrsflralgtllhtnlrikqdsqgalmvypyfgeksaamkkqkmtrrslpeeqeqeqevigskifleidnrqcvqdsdqcfkntdaaaallashaiqgtlsyplvsvfselesprnaqllyllavavviilffillgvimakrkr
人notch3核心调节结构域(humannotch3coreregulatoryregion,hn3c):将人notch3(np_000426)的necd上的最后一次egf重复后的pro1375用作hn3c的n末端。跨膜结构域c末端的4个连续碱性氨基酸(rrkr)的最后一个arg1669用作hn3c的c末端。
paaapevseeprcpraacqakrgdqrcdrecnspgcgwdggdcslsvgdpwrqcealqcwrlfnnsrcdpacsspaclydnfdchaggrertcnpvyekycadhfadgrcdqgcnteecgwdgldcasevpallargvlvltvllppeellrssadflqrlsailrtslrfrldahgqamvfpyhrpspgseprarrelapevigsvvmleidnrlclqspendhcfpdaqsaadylgalsaverldfpyplrdvrgepleppepsvpllpllvagavlllvilvlgvmvarrkr
小鼠notch3核心调节结构域(mousenotch3coreregulatoryregion,mn3c):将小鼠notch3(np_032742)的necd上的最后一次egf重复后的pro1376用作mn3c的n末端。跨膜结构域c末端的4个连续碱性氨基酸(rrkr)的最后一个arg1670用作mn3c的c末端。
psaapevpeeprcpraacqakrgdqncdrecntpgcgwdggdcslnvddpwrqcealqcwrlfnnsrcdpacsspaclydnfdcysggrdrtcnpvyekycadhfadgrcdqgcnteecgwdgldcasevpallargvlvltvllppeellrssadflqrlsailrtslrfrldargqamvfpyhrpspgsesrvrrelgpevigsvvmleidnrlclqsaendhcfpdaqsaadylgalsaverldfpyplrdvrgepleapeqsvpllpllvagavflliifilgvmvarrkr
人notch4核心调节结构域(humannotch4coreregulatoryregion,hn3c):将人notch4(np_004548)的necd上的最后一次egf重复后的pro1162用作hn4c的n末端。跨膜结构域c末端的5个连续碱性氨基酸(rrrrr)的最后一个arg1476用作hn4c的c末端。
phsspgprcqkpgakgcegrsgdgacdagcsgpggnwdggdcslgvpdpwkgcpshsrcwllfrdgqchpqcdseeclfdgydcetppactpaydqychdhfhnghcekgcntaecgwdggdcrpedgdpewgpslallvvlsppaldqqlfalarvlsltlrvglwvrkdrdgrdmvypypgaraeeklggtrdptyqeraapqtqplgketdslsagfvvvmgvdlsrcgpdhpasrcpwdpglllrflaamaavgalepllpgpllavhphagtappanqlpwpvlcspvagvillalgallvlqlirrrrr
小鼠notch4核心调节结构域(mousenotch4coreregulatoryregion,hn3c):将小鼠notch4(np_035059)的necd上的最后一次egf重复后的pro1158用作mn4c的n末端。跨膜结构域c末端的5个连续碱性氨基酸(rrrrr)的最后一个arg1472用作mn4c的c末端。
ppdspgprcqrpgasgcegrggdgtcdagcsgpggdwdggdcslgvpdpwkgcpphsqcwllfrdgrchpqcdseeclfdgydceipltcipaydqycrdhfhnghcekgcnnaecgwdggdcrpegedsegrpslallvvlrppaldqqllalarvlsltlrvglwvrkdsegrnmvfpypgtrakeelsgardssswerqapptqplgketeslgagfvvvmgvdlsrcgpehpasrcpwdsglllrflaamaavgalepllpgpllaahpqagtrpsanqlpwpilcspvvgvlllalgallvlqlirrrrr
hn1ram:来自人notch1的具有前100个氨基酸的ram结构域(np_060087,gln1763至pro1862)。
qhgqlwfpegfkvseaskkkrreplgedsvglkplknasdgalmddnqnewgdedletkkfrfeepvvlpdlsdqtdhrqwtqqhldaadlrmsamaptp。
第1-7位为hn1ram7。第1-8位为hn1ram8。第1-10位为hn1ram10。第1-20位为hn1ram20。第1-30位为hn1ram30。第1-40位为hn1ram40。
mn1ram:来自小鼠notch1的具有前100个氨基酸的ram结构域(np_032740,gln1753tophe1852)。mn1ram7和hn1ram7的顺序相同。
qhgqlwfpegfkvseaskkkrreplgedsvglkplknasdgalmddnqnewgdedletkkfrfeepvvlpdlsdqtdhrqwtqqhldaadlrmsamaptp
第1-7位为mn1ram7。
hn2ram:来自人notch2的具有前100个氨基酸的ram结构域(np_077719,lys1705toser1804)。
khgslwlpegftlrrdasnhkrrepvgqdavglknlsvqvseanligtgtsehwvddegpqpkkvkaedeallseeddpidrrpwtqqhleaadirrtps
第1-7位为hn2ram7。
mn2ram:来自小鼠notch2的具有前100个氨基酸的ram结构域(np_035058,lys1707toleu1806)。
khgflwlpegftlrrdssnhkrrepvgqdavglknlsvqvseanligsgtsehwvddegpqpkkakaedeallseddpidrrpwtqqhleaadirhtpsl
第1-7位为mn2ram7。
hn3ram:来自人notch3的具有前100个氨基酸的ram结构域(np_000426,glu1670tothr1769)。
ehstlwfpegfslhkdvasghkgrrepvgqdalgmknmakgeslmgevatdwmdtecpeakrlkveepgmgaeeavdcrqwtqhhlvaadirvapamalt
第1-7位为hn3ram7。
mn3ram:来自小鼠notch3的具有前100个氨基酸的ram结构域(np_032742,glu1671tophe1677)。mn3ram7和hn3ram7的顺序相同。
ehstlwfpegfalhkdiaaghkgrrepvgqdalgmknmakgeslmgevvtdlndsecpeakrlkveepgmgaeepedcrqwtqhhlvaadirvapatalt
第1-7位为mn3ram7。
hn4ram:来自人notch4的具有前100个氨基酸的ram结构域(np_004548,glu1477toglu1576)。
ehgalwlppgftrrprtqsaphrrrpplgedsiglkalkpkaevdedgvvmcsgpeegeevgqaeetgppstcqlwslsggcgalpqaamltppqeseme
第1-7位为hn4ram7。
mn4ram:来自小鼠notch4的具有前100个氨基酸的ram结构域(np_035059,glu1473toval1572)。mn4ram7和hn7ram7的顺序相同。
ehgalwlppgfirrpqtqqaphrrrpplgedniglkalkpeaevdedgvamcsgpeegeaeetasasrcqlwplnsgcgelpqaamltppqecesevldv
第1-7位为mn4ram7。
lag16:对gfp具有高亲和力的纳米抗体。
maqvqlvesggrlvqagdslrlscaasgrtfstsamawfrqapgrerefvaaitwtvgntilgdsvkgrftisrdrakntvdlqmdnlepedtavyycsarsrgyvlsvlrsvdsydywgqgtqvtvs
anti-cd19scfv(αcd19):对cd19具有高亲和力的单链抗体。
diqmtqttsslsaslgdrvtiscrasqdiskylnwyqqkpdgtvklliyhtsrlhsgvpsrfsgsgsgtdysltisnleqediatyfcqqgntlpytfgggtkleitggggsggggsggggsevklqesgpglvapsqslsvtctvsgvslpdygvswirqpprkglewlgviwgsettyynsalksrltiikdnsksqvflkmnslqtddtaiyycakhyyyggsyamdywgqgtsvtvss
tta-advanced(ttaa):转录因子,其相应的启动子是tre3g,具有7个拷贝的四环素响应元件(tccctatcagtgatagaga)。
msrldkskvinsalellnevgieglttrklaqklgveqptlywhvknkralldalaiemldrhhthfcplegeswqdflrnnaksfrcallshrdgakvhlgtrptekqyetlenqlaflcqqgfslenalyalsavghftlgcvledqehqvakeeretpttdsmppllrqaielfdhqgaepaflfgleliicglekqlkcesggpadalddfdldmlpadalddfdldmlpadalddldldmlpg
gal4-vp64(gv2):转录因子,其相应的启动子是具有5个拷贝的gal4靶序列(ggagcactgtcctccgaacg)的puas。
mkllssieqacdicrlkklkcskekpkcakclknnwecryspktkrspltrahltevesrlerleqlfllifpredldmilkmdslqdikalltglfvqdnvnkdavtdrlasvetdmpltlrqhrisatssseessnkgqrqltvaaaggsggsggsdalddfdldmlgsdalddfdldmlgsdalddfdldmlgsdalddfdldmlgs
mousenotch1nrr
ildysftggagrdipppqieeacelpecqvdagnkvcnlqcnnhacgwdggdcslnfndpwknctqslqcwkyfsdghcdsqcnsagclfdgfdcqltegqcnplydqyckdhfsdghcdqgcnsaecewdgldcaehvperlaagtlvlvvllppdqlrnnsfhflrelshvlhtnvvfkrdaqgqqmifpyygheeelrkhpikrstvgwatssllpgtsggrqrreldpmdirgsivyleidnrqcvqsssqcfqsatdvaaflgalaslgslnipykieavksepvepplp
mousenotch1tmd
sqlhlmyvaaaafvllffvgcgvllsrkrrr
加粗:s3蛋白水解位点
humannotch1nrr
ildysfgggagrdippplieeacelpecqedagnkvcslqcnnhacgwdggdcslnfndpwknctqslqcwkyfsdghcdsqcnsagclfdgfdcqraegqcnplydqyckdhfsdghcdqgcnsaecewdgldcaehvperlaagtlvvvvlmppeqlrnssfhflrelsrvlhtnvvfkrdahgqqmifpyygreeelrkhpikraaegwaapdallgqvkasllpggseggrrrreldpmdvrgsivyleidnrqcvqassqcfqsatdvaaflgalaslgslnipykieavqsetvepppp
第235-238位:s1蛋白水解位点;第294-295位:s2蛋白水解位点。
humannotch1tmd
aqlhfmyvaaaafvllffvgcgvllsrkrrr
第20-23位:s3蛋白水解位点
实施例2测定合成notch受体的配体非依赖性激活强度
抗原(antigen,ag)诱导的synnotch信号转导的示意图如图1所示。当表达synnotch的细胞识别发送细胞(sender,或称为靶细胞)上呈递的特异性抗原(antigen)时,发生顺序蛋白水解切割,以释放转录因子(transcriptionfactor,tf)进入细胞核并诱导其对应启动子下游的转录。
我们根据实施例1中所述方法,构建了一系列合成notch受体即其变体。现有的synnotch结构由scfv、nrr、tmd和tf依次连接形成,并且不包含ram结构域。用于瞬时共转染测定的质粒如图2,用于测量synnotch的配体非依赖性激活或错误激活。使用pgk启动子驱动表达synnotch,表达synnotch的细胞总是同时表达pgk驱动的mcherry,短寿命版本的egfp(d2egfp)受tf对应的启动子(promoter,pro)驱动表达。
我们将携带有上述元件的质粒共转染入hek293t细胞,每次转染使用总量为450ngpgk驱动的质粒,包括恒定量的200ngpgk驱动的mcherry和各种量的pgk驱动的synnotch(10、50、250ng)。表达synnotch的细胞通过检测mcherry的荧光以测量共转染效率。在不同synnotch转染剂量(10、50、250ng)下,我们都能够获得40-60%的转染效率(图3),这说明我们的共转染实验体系是稳定的。
接下来我们选用一个代表性的synnotch构建体(lag16-mn1c-ttaa)来测试synnotch的配体非依赖性激活。synnotch即lag16-mn1c-ttaa结构如图4a所示,其细胞外抗原识别结构域为能够特异性识别gfp的高亲和力纳米抗体lag16;notch核心调节结构域为来源于小鼠notch1的核心调节结构域;细胞内转录调节结构域为ttaa。ttaa可以诱导其对应启动子tre3g下游的d2egfp的表达。不同synnotch转染量下其下游表达d2egfp荧光强度如图4b的堆积山峰图所示;对于0ngsynnotch的共转染组,使用仅含有pgk启动子且不含synnotch的质粒250ng。通过计算图4b中每个共转染组的中值d2egfp荧光强度并以柱状图表示,如图4c;虚线对应于共转染0ngsynnotch时的平均值。我们发现该切除了野生型ram结构域的synnotch,即使在没有抗原激活的情况下,也能够观测到下游的d2egfp的错误的激活信号。并且synnotch转染剂量越高,产生的错误激活信号也越高(图4b、4c)。
由于在上述测定条件下,我们并没有使用抗原表达的细胞以通过配体依赖途径激活synnotch,因而我们所检测到的错误信号由配体非依赖性激活途径触发。上述实验表明,现有synnotch具有显著的配体非依赖性激活(ligand-independentactivation,lia)或错误的激活。
接下来,我们通过进一步的实验以了解synnotch膜表达量和lia间的关系。向细胞转染入如图4a所示的不同剂量的synnotch,固定细胞后用抗myc标签的抗体染色(不透化)细胞(我们在信号肽和scfv之间插入了myc标签,以便于检测细胞膜上的synnotch表达)。我们发现synnotch的膜表达量与转染的dna的量正相关(图5)。图5显示了不同转染条件下细胞mcherry和细胞外myc标签的荧光信号;抗myc标签的抗体的荧光强度指征不同转染剂量下synnotch的膜表达量,mcherry荧光用于标记被成功转染synnotch的细胞;每个等高线图的右上角的数字是表示高表达synnotch的细胞占所有细胞的的百分比。我们构建了稳定膜表达不同量synnotch的细胞。稳定表达synnotch的细胞被固定后染色,不同单细胞的细胞外myc标签的荧光信号(用于指征synnotch的膜表达量)和d2egfp的荧光强度(用于指征对应细胞的lia强度)如图6散点图所示。具有高synnotch稳定表达的细胞的lia也更高。这再次说明,细胞的lia是伴随synnotch的表达而产生。
synnotch作为转导细胞膜外信号进入细胞内的工具,错误的信号的产生可能导致强烈的底噪因而影响信号的分辨率。因此,我们接下来测试了不同转染剂量下synnotch细胞转导抗原依赖性激活信号的信噪比。我们用gfp作为模拟抗原。将不表达或表达gfp的发送细胞(sender)和转染了不同剂量synnotch的细胞共培养(如图7a)。计算每个共培养组的中值d2egfp荧光强度并以柱状图(平均值±sd)表示;比较与不表达gfp的发送细胞/表达gfp的发送细胞共培养的synnotch细胞的d2egfp荧光强度平均值计算激活倍率,如图7b。结果显示只有在低或中等synnotch转染剂量下,表达synnotch的细胞才能够表现出可分辨的抗原依赖性激活信噪比。具有高synnotch表达的细胞,由于具有高lia,因而在没有或有发送细胞的情况下,均产生相似水平的d2egfp荧光(图7b)。上述实验说明,synnotch转导信号的信噪比随synnotch的lia的增加而降低。
实施例3插入egf不能够降低synnotch的lia
先前的一项研究表明,在notch核心调节结构域的n末端插入一个egf重复序列可以减少lia。
我们在lag16-mn1c-ttaa的基础上,在lag16和nrr之间插入了一个egf重复,以构建lag16-egf-mn1c-ttaa。使用瞬时共转染测试两种synnotch的lia。如图8b所示,计算每次共转染的中值d2egfp荧光强度并以柱状图(平均值±sd)表示;egf(-)代表图4a中的synnotch,而egf(+)代表图8a中的synnotch。结果显示,lag16-mn1c-ttaa和lag16-egf-mn1c-ttaa的lia均随着synnotch转染剂量的上升而显著上升(图8b)。该实验不支持先前研究的结论,我们认为egf结构域的缺失不是synnotch的lia或错误激活产生的原因。
实施例4synnotch的lia由于配体非依赖性的s3切割导致
synnotch的激活依赖于s1,s2和s3的顺序切割。为了研究lia的产生原因,我们产生了在s1,s2或s3蛋白水解位点具有突变的synnotch突变体(s1突变体:lag16-mn1c[s1]-ttaa;s2突变体:lag16-mn1c[s2]-ttaa;s3突变体:lag16-mn1c[s3]-ttaa),如图9a。将这些synnotch变体分别转染入细胞,计算每次共转染的中值d2egfp荧光强度,如图9b。我们发现具有s1或s2突变的synnotch变体的lia仍然很高,而具有s3突变的synnotch变体则具有显著降低的lia(降低9.5倍)(图9b)。
接下来,我们通过用特异性蛋白酶抑制剂处理表达有不含有水解位点突变的synnotch(lag16-mn1c-ttaa)的细胞来进一步证实前述结论。如图10a,在表达synnotch的细胞中加入不同浓度的蛋白酶抑制剂一起温育,计算每次共转染的中值d2egfp荧光强度,并以柱状图(平均值±sd)表示,通过双尾t检验计算统计p值,结果如图10b和图10c。使用adam(能够特异性切割s2位点)特异性抑制剂bb94(sigma-aldrichcas#130370-60-4)并未在统计上改变lia(图10b)。而使用γ分泌酶(能够特异性切割s3位点)特异性抑制剂化合物e(sigma-aldrichcas#209986-17-4),可以以剂量依赖得方式有效地降低lia(图10c)。上述实验表明,γ-分泌酶受到抑制可以降低synnotch的lia。总之,我们在实验中检测了不同测试条件下synnotch的lia,发现过量产生的synnotch的抗原非依赖性s3切割可能是lia产生的原因。
实施例5具有ram结构域的增强型合成notch受体(esnotch)具有低lia
基于最近对γ-分泌酶的结构研究,当天然notch被γ-分泌酶识别时,跨膜结构域(tmd)的c末端必须精确定位以与蛋白酶形成杂合β-折叠(yang,g.,etal.structuralbasisofnotchrecognitionbyhumangammasecretase.nature565,192-197(2019).)。另一项结构研究表明,tmdc末端后面的ram序列可以形成膜折返结构,这可能有助于tmd的定位(deatherage,c.l.,etal.structuralandbiochemicaldifferencesbetweenthenotchandtheamyloidprecursorproteintransmembranedomains.scienceadvances3,e1602794(2017).)。目前研究没有任何证据表明ram序列可能对synnotch正确激活或lia产生有直接关系。但是我们同时注意到,ram序列未包含在任何已发布的synnotch中,因而我们认为应该检查它对synnotch的lia的影响。
我们构建了在lag16-mn1c-ttaa的mn1c和ttaa间的插入了ram序列(q1763hgqlwfp1770,来自人notch1ram结构域,该序列命名为hn1ram8)的synnotch变体(lag16-mn1c-hn1ram8-ttaa),如图11a所示。使用瞬时共转染测试lag16-mn1c-ttaa和lag16-mn1c-hn1ram8-ttaa在不同转染剂量下的lia。计算每次共转染的中值d2egfp荧光强度,并以柱状图(平均值±sd)表示,基于来自不含或含有hn1ram8的250ng共转染的synnotch组的荧光平均值计算倍数变化,结果如图11b所示。结果显示,后者相对于前者,由于添加了ram序列,因而lia显著降低,降低倍率高达14.6倍(图11b)。上述结果证明hn1ram8对于减少lia是有效的。
为了了解添加ram序列是否会干扰合成notch受体的膜靶向。我们用lag16-mn1c-ttaa和lag16-mn1c-hn1ram8-ttaa分别转染细胞。用抗myc标签的抗体在不透化条件下对细胞染色。如图12的堆积山峰图显示,具有ram序列的synnotch变体,相对于无ram的原始synnotch,其表达量和表达分布均未改变。综合上述结果,说明ram序列的插入,能在不干扰合成notch受体的膜靶向的条件下降低synnotch的lia。上述实验首次发现了ram序列对于降低synnotch的lia的重要功能,并首次证明了可以使用插入ram序列以降低synnotch的lia的方法的有效性。我们将这种特别携带有ram序列的合成notch受体命名为增强型合成notch受体(enhancedsyntheticnotchreceptor,esnotch)。
实施例6用于构建esnotch所需的最短ram序列的确认
在构建合成蛋白时,我们总是希望能够使用最精简的元件来作为构建的素材,以最大限度的减少元件冗余功能对合成蛋白可能带来的潜在干扰。天然ram结构域在天然notch-csl转录复合物组装过程中形式重要功能(choi,sungh.,etal.conformationallockinguponcooperativeassemblyofnotchtranscriptioncomplexes.structure20,340-349(2012).),因而采用未经优化的天然ram序列具有干扰天然notch下游通路的风险。因而,为了防止这样不利因素的产生,一种优选的方法就是确定ram序列的最简化结构,以在保证所插入的ram序列能够有效降低合成notch受体的lia同时,又尽可能的对天然notch下游通路产生干扰。
为了确定能够降低synnotch的lia或错误激活的ram序列的最小结构。我们构建了具有不同ram序列(ram7、ram8、ram10、ram20、ram30和ram40分别为来自人notch1ram结构域n末端的前7、8、10、20、30和40个氨基酸序列)插入的esnotch:lag16-hn1c-hn1ram7-ttaa、lag16-hn1c-hn1ram8-ttaa、lag16-hn1c-hn1ram10-ttaa、lag16-hn1c-hn1ram20-ttaa、lag16-hn1c-hn1ram30-ttaa、lag16-hn1c-hn1ram40-ttaa。如图13,初步实验显示7-40个氨基酸长度的ram序列都具有能够降低synnotch的lia或错误激活的能力。同时注意到,在这些序列长度中,具有ram7、ram8插入esnotch的lia或错误激活要相对低于具有ram10、ram20、ram30、ram40插入esnotch的lia。
这暗示我们,具有功能的最简化的ram序列长度可能更短。我们接下来进行了进一步的实验以对此进行验证。
因此,我们进一步构建了一系列具有不同长度ram序列(ram1、ram2、ram3、ram4、ram5、ram6、ram7和ram8分别为来自人notch1ram结构域n末端的前1、2、3、4、5、6、7和8个氨基酸序列)的esnotch变体:lag16-mn1c-hn1ram1-ttaa、lag16-mn1c-hn1ram2-ttaa、lag16-mn1c-hn1ram3-ttaa、lag16-mn1c-hn1ram4-ttaa、lag16-mn1c-hn1ram5-ttaa、lag16-mn1c-hn1ram6-ttaa、lag16-mn1c-hn1ram7-ttaa、lag16-mn1c-hn1ram8-ttaa。并通过瞬时共转染测试不同变体的lia,计算每次共转染的中值d2egfp荧光强度,并以柱状图(平均值±sd)表示,统计p值由双尾t检验计算,如图14。我们发现仅携带有ram7和ram8的esnotch和原始synnotch相比具有显著降低的lia。而具有更短长度的ram序列的变体的lia仍然维持在较高水平。因而,我们确定了减少lia所需的最小ram序列为具有7个氨基酸长度的hn1ram7。hn1ram7能够有效降低synnotch的lia,同时其相对于hn1ram8,又由于缺失最后一个pro而不具有完整的φwφp基序(φwφp基序是notch-csl复合物组装过程的关键位点(choi,sungh.,etal.conformationallockinguponcooperativeassemblyofnotchtranscriptioncomplexes.structure20,340-349(2012).)),因而能够最小化对于天然notch通路的干扰。综上ram7是优选的能够被应用于降低synnotch的高lia的最简化结构域。
实施例7检验具有其他天然ram序列的esnotch的lia
为了研究ram减少lia的序列要求,我们比对了来自人和小鼠的不同notch同种型。
我们注意到不同来源的ram7序列都含有高度保守的疏水性三肽φwφ(φ=疏水性残基)(图15)。因而我们猜想这些具有差异的ram序列应该都能够被应用于降低synnotch的lia。
我们构建了一系列具有不同ram7序列插入的esnotch:lag16-hn1c-hn1ram7-ttaa、lag16-hn1c-hn2ram7-ttaa、lag16-hn1c-mn2ram7-ttaa、lag16-hn1c-hn3ram7-ttaa、lag16-hn1c-hn4ram7-ttaa。使用瞬时共转染法测试不同esnotch的lia,计算每次共转染的中值d2egfp荧光强度,结果如图16所示;组(-)是如图4a中的synnotch。结果显示,具有不同来源的ram7序列插入的esnotch的lia或错误激活,相比于原始synnotch均显著降低,降低倍率达10倍以上。
实施例8检验具有人工合成ram序列的esnotch的lia
接下来,我们将具有不同氨基酸突变的人工合成ram序列插入synnotch的tmd和ttaa之间,更进一步构建了具有不同氨基酸序列的人工合成ram序列的esnotch:lag16-hn1c-hn1ram7[fw]-ttaa、lag16-hn1c-hn1ram7[ww]-ttaa、lag16-hn1c-hn1ram7[ff]-ttaa、lag16-hn1c-hn1ram7[ll]-ttaa、lag16-hn1c-hn1ram7[aa]-ttaa、lag16-hn1c-hn1ram7[gg]-ttaa、lag16-hn1c-hn1ram7[pp]-ttaa、lag16-hn1c-hn1ram7[ss]-ttaa、lag16-hn1c-hn1ram7[ee]-ttaa。
使用瞬时共转染法测试不同esnotch的lia,计算每次共转染的中值d2egfp荧光强度,并以柱状图(平均值±sd)表示,如图17;组(-)是如图4a中的synnotch。结果显示,当所采用的人工ram序列中含有二疏水氨基酸对时(如wf、fw、ww、ff、ll),esnotch具有低lia或错误激活。而那些使用亲水性氨基酸对突变(如aa、gg、pp、ss、ee)而不具有任意二疏水氨基酸对的人工合成ram序列的esnotch的lia或错误激活仍然维持高水平。因而,我们得到以下结论:在使用人工合成ram序列以构建具有低lia或错误激活的esnotch时,合成ram序列中需要具有任意一种疏水性二氨基酸对。
实施例9
本实施例分析本发明的增强型合成notch受体(esnotch)具有低lia的原因。
由于使用天然来源或人工合成的ram序列都能够的构建具有低lia或错误激活的esnotch。ram序列的变异性使得其不大可能是通过被某种潜在蛋白识别而发挥功能。同时由于能够发挥降低lia功能的最短ram序列仅为7个氨基酸,因而其也不大可能是独立发挥功能以降低lia。因此,我们假定ram是和notch蛋白的其他结构配合下而导致的低lia。为了验证这一猜想,我们构建了缺失局部结构的esnotch构建体(hn1c-hn1ram7-ttaa、hn1c-ttaa、hn1tmd-hn1ram7-ttaa、hn1tmd-ttaa)。结果显示,hn1ram7必须和nrr同时存在才能维持esnotch的低lia(图18)。ram,nrr,notchsignaling的关系满足nand逻辑,可表示为notchsignaling=not(nrrandram)或notchsignaling=nrrnandram(图19)。该发现和γ分泌酶对于notch的切割的基本要求一致,但首次揭示ram结构域在该过程中所起到的关键所用。
实施例10构建具有不同细胞外识别结构域、notch核心调节结构域和转录调节结构域的esnotch,并测试其lia和抗原诱导的激活倍率
为了说明本研究所提供的降低合成notch受体的lia或错误激活的方法具有通用性。我们构建了一系列具有不同细胞外识别结构域、notch核心调节结构域和转录调节结构域的具有ram序列的esnotch:
lag16-mn1c-hn1ram7-gal4-vp64(结构如图20a所示,类似于图7a,但是在tmd和gv2之间插入了hn1ram7。因为tf从ttaa变为gv2,所以使用puas驱动d2egfp)、lag16-hn1c-hn1ram7-ttaa(结构如图20b所示,类似于图7a,但是在tmd和ttaa之间插入hn1ram7,以及用人notch1核心调节结构域替换小鼠notch1核心调节结构域)、cd19-hn1c-hn1ram7-ttaa(结构如图20c所示,类似于图20b,但使用αcd19作为scfv)。值得注意的是,lag16-hn1c-hn1ram7-ttaa、cd19-hn1c-hn1ram7-ttaa两种esnotch的设计采用了人来源的notch核心调节结构域因而被应用于临床研究时能够降低潜在的免疫原性,因而具有更好的安全性。
将转染esnotch的细胞与发送细胞一起共培养,以测试esnotch的抗原诱导激活倍率。图20a-c所对应的esnotch的实验结果分别如图20a’-c’所示;计算每个共转染组的中值d2egfp荧光强度并以柱状图表示,其中条高为平均值,误差条为标准偏差;基于与具有或不具有抗原的发送器细胞以1:4的比例共培养的synnotch细胞的平均值计算倍数激活;标记为(-)的组的共转染没有ram序列synnotch。
转染esnotch的细胞不与发送细胞一起共培养,以测试esnotch的lia。图20a-c所对应的esnotch的实验结果分别如图20a”-c”所示;计算每个共转染的中值d2egfp荧光强度并以柱状图(平均值±sd)表示,基于来自没有hn1ram7的原始synnotch或具有hn1ram7的esnotch的250ng共转染组的荧光平均值计算倍数变化。
我们发现,我们所设计的具有ram序列的esnotch(lag16-mn1c-hn1ram7-gal4-vp64、lag16-hn1c-hn1ram7-ttaa、αcd19-hn1c-hn1ram7-ttaa)相对于没有ram序列的原始synnotch(lag16-mn1c-gal4-vp64、lag16-hn1c-ttaa、αcd19-hn1c-ttaa),都具有显著降低的lia或错误激活(图20a”-c”),并且能够被抗原正确激活,而具有显著的激活倍率(图20a’-c’)。
综上,基于我们所提供的降低synnotch的lia或错误方法而构建具有ram序列的esnotch,由于具有显著降低的lia而具有优异的信号转导性能,因而能够更有效地自定义调节各种细胞过程,而能够被安全而稳健地应用于多种临床或基础研究情景;这扩展了原始synnotch的应用范围,为使用嵌合受体细胞改造宿主细胞提供了新的应用前景,具有巨大的经济价值和意义。
- 还没有人留言评论。精彩留言会获得点赞!