包含重组毒素的针对艰难梭菌的疫苗的制作方法
本发明涉及使用选自来自艰难梭菌(Clostridium difficile)的毒素A (TcdA)、毒素B (TcdB)和二元毒素(CDTa和CDTb)蛋白的至少一种重组毒素蛋白以及与另外免疫原的组合,诱导针对艰难梭菌的免疫应答的方法。本发明还涉及包含单独或与一种或多种另外的免疫原例如TcdA和TcdB组合的CDTa和/或CDTb蛋白的免疫原性组合物,特别是艰难梭菌疫苗,其能够诱导保护性免疫应答。
相关申请的交叉引用
本申请要求于2012年1月27日提交的美国临时申请号61/591,631、于2012年2月8日提交的美国临时申请号61/596,419、和于2012年9月20日提交的美国临时申请号61/703,754的权益,所述美国专利申请的内容在此整体通过引用并入本文。
对电子提交的序列表的提及
本申请的序列表经作为具有文件名“23192-PCT-SEQLIST-25JAN2013.TXT”的ASCII格式的序列表由EFS-Web电子提交,所述序列表具有2013年1月25日的创建日期和632 KB的大小。EFS-Web提交的这个序列表是说明书的部分并整体通过引用并入本文。
发明背景
梭菌属(Clostridium)是厌氧革兰氏阳性产孢子细菌属,其牵涉土壤和哺乳动物胃肠道的厌氧区室中的有机物质的分解和降解。为了影响其功能,许多梭菌属物种编码并表达靶向活物质的多种组分的降解酶。这些酶中的一些已进化出高度有效的毒素和细胞毒素。这些毒素的表达可以将一些梭菌属物种变成机会致病菌,包括破伤风梭菌(Clostridium tetani)、肉毒梭菌(Clostridium botulinum)、产气荚膜梭菌(Clostridium perfringens)和艰难梭菌,其分别引起破伤风、肉毒中毒、气性坏疽和抗生素相关性结肠炎。
艰难梭菌感染(CDI)是在工业化国家中最常见的医院内感染之一,由于由一种或多种抗菌剂引起的正常结肠菌群的破坏,通常在经历抗生素治疗的住院患者中流行。在美国每年估计有500,000个CDI病例(Campbell等人,Infect Control Hosp Epidemiol,30(6): 526-33(2009);Rupnik等人,Nature Reviews,7: 527-36(2009))。在美国报道的艰难梭菌病例数目已上升,如从2001年到2005年具有CDI出院诊断的住院治疗率增加92%所指出的。几乎所有抗生素包括克林霉素、头孢菌素、青霉素和氟喹诺酮均已与由产毒艰难梭菌引起的症状性病相关(Bartlett,J.G.,Clin Infect Dis. 46(10): 1489-92(2008);Gorbach,S.L.,N Engl J Med,341(22): 1690-1(1999);Kelly等人,N Engl J Med 330(4): 257-62(1994))。艰难梭菌相关疾病(CDAD)包括抗生素相关性腹泻、结肠炎和假膜性结肠炎,并且在一些情况下,可以进展至中毒性巨结肠、败血症和死亡。为此,CDI是重大健康护理事件。尽管患有CDAD的一些患者响应抗生素治疗的停止,但大多数需要用进一步试剂例如甲硝唑或万古霉素的治疗(在DuPont等人,Current Opinion in Infectious Diseases 21:500-507(2008)中综述)。现有治疗通常未能消除CDAD或导致复发性疾病,因此需要新的预防或治疗性处理的方法。
经过过去十年,严重或致命性感染的增加、护理标准治疗失败、毒力更高的流行株(NAP1/027/BI)的出现、增加的社区获得性感染和复发性感染的发病率(估计为~15-25%)已促成关于CDI的治疗和/或预防性处理的增加必要性(Aslam等人,Lancet, 365(9469): 1464(2005);Loo等人,N Engl J Med, 365(18): 1693-703(2006);McDonald等人,N Engl J Med, 353(23): 2433-41(2005);Muto,C.A., Infect Control Hosp Epidemiol, 26(1): 10-2(2005);Pépin等人,Cmaj,. 171(5): 466-72(2004);Zar等人,Clin Infect Dis,45(3): 302-7.(2007))。几项研究近期已提出社区获得性CDI的发病率已增加(MMWR Morb Mortal Wkly Rep, 57(13): 340-3(2008);Hirschhorn等人,J Infect Dis,. 169(1): 127-33(1994);Karlstrom等人,Clin Infect Dis, 26(1): 141-5(1998);Kyne等人,J Infect, 36(3): 287-8(1998);Lambert等人,Infect Control Hosp Epidemiol, 30(10): 945-51(2009);Noren等人,J Clin Microbiol, 42(8): 第3635-43页(2004);Paltansing等人,Clin Microbiol Infect,13(11): 1058-64(2007)。
产毒的艰难梭菌分离物通常编码称为毒素A和毒素B (“TcdA”和“TcdB”)的两种大的梭菌毒素,由于其罕见的大分子量(> 250 kDa)和作用机制,它们是称为大梭菌毒素(LCT)的较大毒素家族的部分(Schirmer和Aktories,Biochimica et Biophysica Acta1673: 66-74(2004))。另外的产毒株仅表达毒素B且不表达毒素A。TcdA和TcdB已证实是CDI的唯一致病因子。艰难梭菌株亚组还产生肌动蛋白-ADP-核糖基化毒素,称为艰难梭菌转移酶或CDT。通常称为二元毒素的这种毒素特别与毒力高的流行株例如NAP1/027/BI相关,并且由两种蛋白CDTa和CDTb组成。
CDTb亚单位形成七聚环状结构,其允许酶促活性的CDTa蛋白进入细胞质内。CDTa蛋白催化G-肌动蛋白在精氨酸177处的ADP-核糖基化,并且阻止肌动蛋白聚合(Barth,H.,等人,Binary bacterial toxins: biochemistry, biology, and applications of common Clostridium and Bacillus proteins.Microbiol Mol Biol Rev,68(3): 373-402(2004))。二元毒素在艰难梭菌疾病的发病机制中的精确作用定义不明;然而,这些毒素已显示作为肠毒素发挥作用(Geric,B.,等人,Journal of Infectious Diseases 193(8): 1143-1150(2006)),以诱导微管形成并从而增加艰难梭菌定殖(Schwan,C.等人,PLoS Pathog, 5(10): e1000626(2009)),并且最近已将其与和含非二元毒素的毒株相比较增加的死亡率相关联(Bacci等人,Emerg. Infect. Dis., 17(6): 976-82(2011);Goldenberg SD, Journal of Infection, 62: 355-62(2011))。
艰难梭菌疫苗开发的几种方法在文献中描述,包括用中和艰难梭菌毒素的多克隆血清的被动免疫接种(参见Thomas等人,WO 99/20304和WO 2005/058353)、用毒素片段的免疫接种(Ward等人,WO 98/59053;Lyerly等人,Current Microbiology 21: 29-32(1990);WO 00//061761)和用福尔马林失活的完全天然毒素的免疫接种(Torres等人,Infec tionand Immunity 63(12): 4619-4627(1995);Kim等人,Infection and Immunity 55(12): 2984-2992(1987));Fernie等人Devel.Biol.Standard 53: 325-32(1983);Libby等人,Infect Immun 36: 822-9(1982);Ghose等人,Infection and Immunity 75(6): 2826-2832)。还描述的是编码毒素片段的基于DNA的疫苗(Gardiner等人,Vaccine 27:3598-3604(2009)),包括二元毒素(J.E. Galen,WO 2011/060431)。Ellingsworth等人(WO 2012/028741)公开了包含TcdA和TcdB的受体结合结构域的重组融合蛋白。
还描述了基于重组毒素的疫苗的应用,在所述疫苗中促成毒性的特异性位点被突变。Ballard和Spyres描述了重组TcdB疫苗,其中在第395位处的半胱氨酸替换为另一种氨基酸(美国专利号7,226,597)。描述为对于酶活性重要的其他位点是在TcdB的第102位处的色氨酸残基(W102)和催化三联基序DXD,其是糖基转移酶A家族的标志基序,LCT是所述糖基转移酶A家族的部分(Busch等人,J.Biol.Chem. 275(18): 13228-234(2000))。Feng等人(WO 2011/068953)描述了具有单一、双重或三重突变的TcdA毒素和具有双重突变以消除毒性的TcdB毒素。
发明概述
本文描述的本发明涉及重组艰难梭菌毒素A (TcdA)蛋白,其包含选自下述的至少3个突变或由选自下述的至少3个突变组成:W101A、D287A、E514Q、D285A、S517A、W519A和C700A。在本发明的优选实施方案中,重组TcdA蛋白包含选自下述的一组突变或由选自下述的一组突变组成:(a) W101A、D287A和E514Q;(b) W101A、D287A、E514Q和W519A;(c) W101A、D287A、E514Q和D285A;(d) W101A、D287A、E514Q和S517A;以及(e) W101A、D287A、E514Q、W519A和C700A。在另外的实施方案中,TcdA蛋白是基本上纯化的。
本发明进一步涉及重组艰难梭菌毒素B (TcdB)蛋白,其包含选自下述的至少3个突变或由选自下述的至少3个突变组成:W102A、D288A、E515Q、D286A、S518A、W520A和C698A。在本发明的这个方面的优选实施方案中,纯化的TcdB蛋白包含选自下述的一组突变:(a) W102A、D288A和E515Q;(b) W102A、D288A、E515Q和W520A;(c) W102A、D288A、E515Q和D286A;(d) W102A、D288A、E515Q和S518A;以及(e) W102A、D288A、E515Q、W520A和C698A。在另外的实施方案中,TcdB蛋白是基本上纯化的。
本发明还涉及包含本文描述的TcdA和/或TcdB突变型毒素与药学可接受的载体的疫苗和免疫原性组合物。在优选实施方案中,所述组合物和疫苗包含TcdA和TcdB突变蛋白。组合物和疫苗可以进一步包含佐剂,例如铝佐剂和/或ISCOMATRIX®。
本发明进一步涉及通过在允许核苷酸序列表达以产生重组突变蛋白(其可以是分离的和/或纯化的)的条件下,在细胞培养中表达编码TcdA或TcdB蛋白序列的核苷酸序列,生产TcdA和TcdB突变蛋白的方法。在本发明的具体实施方案中,TcdA和TcdB突变型毒素蛋白使用在昆虫细胞培养中的杆状病毒表达系统来产生。
本发明还涉及包含选自下述的至少两种突变的重组二元毒素蛋白A (CDTa):C2A、Y67A、Y69A、Y253A、R255A、Y258A、R302A、Q307E、N342A、S345F、F356A、R359A、Y382A、E385Q、E385A、E387Q和E387D。在一些实施方案中,CDTa突变选自:C2A、R302A、S345F、E385Q、E385A、E387Q和E387D。在本发明的这个方面的实施方案中,CDTa蛋白包含选自下述的一组突变:(a) S345F、E385Q和E387Q;(b) R302A、E385A、E387D;(c) C2A、S345F、E385Q和E387Q;(d) R302A、S345F、E385Q和E387Q;(e) Y67A、Y69A和R255A;(f) R359A、Y67A和Q307E;(g) Y258A、F356A、S345F;以及(h) N342A、Y253A和Y382A。在本发明的优选实施方案中,CDTa蛋白缺少信号肽。在本发明的另外实施方案中,CDTa蛋白是基本上纯化的。
本发明进一步涉及包含核苷酸序列的核酸分子,所述核苷酸序列编码本文描述的TcdA、TcdB、CDTa和CDTb蛋白。在一些实施方案中,核苷酸序列对于在选择的宿主细胞中的最佳表达进行密码子最佳化,所述宿主细胞例如大肠杆菌(E.coli)或巨大芽孢杆菌(B.megaterium)。本文进一步提供的是包含本发明的核苷酸序列的载体和宿主细胞。
本文还提供了免疫原性组合物,其包含本文描述的TcdA、TcdB和/或CDTa蛋白和来自艰难梭菌的二元毒素蛋白B (CDTb)蛋白以及药学可接受的载体。在本发明的优选实施方案中,CDTb蛋白缺少信号肽。在另外的实施方案中。CDTb蛋白进一步缺少活化结构域。在优选实施方案中,所述组合物包含TcdA、TcdB、CDTa和CDTb蛋白,连同药学可接受的载体。在本发明的一些实施方案中,免疫原性组合物进一步包含佐剂。
本发明还涉及生产CDTa或CDTb蛋白的方法,其包括在允许核苷酸序列表达以产生CDTa或CDTb蛋白的条件下,用包含编码CDTa或CDTb蛋白的核酸序列的载体转化细胞。
本文还提供的是治疗和/或预防艰难梭菌相关疾病的方法,其包括给患者施用有效量的本发明的免疫原性组合物或疫苗。
如说明书和附加权利要求中自始至终使用的,除非上下文另有明确说明,否则单数形式“一个”、“一种”和“该/所述”包括复数的参考。
本申请自始至终,提及“包含”是开放式的,并且允许添加进一步的元件例如突变。在一个实施方案中,本申请自始至终提及“包含”可以是“由……组成”或“基本上由……组成”。在一些情况下,术语“由……组成”用于突出具有术语“由……组成”的实施方案的可能性。
在本文自始至终对毒素突变的一般讨论中,其中该讨论并未具体指TcdA或TcdB,氨基酸残基编号指TcdB的正确编号,除非另有说明(具体涉及TcdA的文本提供了TcdA的正确编号,并且具体涉及TcdB的文本提供了TcdB的正确编号)。由于TcdA中在残基编号13后的单个氨基酸缺失(相对于TcdB,关于TcdA和TcdB的比对参见图16),相对于毒素B,关于毒素A突变的残基编号对于毒素A移动一个单位(即,对于TcdA,本文描述的突变的残基编号相对于TcdB是n-1;例如TcdB中的W102是TcdA中的W101)。例外是TcdA中的半胱氨酸突变C700A,其是TcdB中的C698A,由于在TcdA的半胱氨酸蛋白酶结构域中的另外三个残基的存在(参见图16)。
如说明书和附加权利要求中自始至终使用的,应用下述定义和缩写:
“TcdA”和“TcdB”分别指毒素A和毒素B,其是来自产毒的艰难梭菌分离物的大羧菌毒素,及其在本文中描述的突变体和衍生物。提及“TcdA”和/或“TcdB”不要求蛋白使用艰难梭菌作为表达系统来产生,即可以采用如本文描述的不同表达系统。
“CDTa”和“CDTb”分别是来自肌动蛋白-ADP-核糖基化毒素的亚单位蛋白A和B,所述肌动蛋白-ADP-核糖基化毒素称为艰难梭菌转移酶或CDT,其也称为二元毒素。如本文定义的,“CDTa”和“CDTb”包括在本文中描述的天然或野生型CDTa和CDTb的突变体和衍生物。提及“CDTa”和/或“CDTb”不要求蛋白使用艰难梭菌作为表达系统来产生,即可以采用如本文描述的不同表达系统。
突变体在本文中一般如下提及:具有3个点突变的毒素A (3mTcdA)、具有3个点突变的毒素B (3mTcdB)、具有4个点突变的毒素A (4mTcdA)、具有4个点突变的毒素B (4mTcdB)、具有5个点突变的毒素A (5mTcdA)和具有5个点突变的毒素B (5mTcdB)。
二元毒素A的特异性突变体如下提及:3mCDTa1指具有下述突变的CDTa减去信号肽:R302A、E385A和E387D。3mCDTa2指具有下述突变的CDTa减去信号肽:S345F、E385Q和E387Q。4mCDTa3指具有下述突变的CDTa减去信号肽:R302A、S345F、E385Q和E387Q。3mCDTa4指具有下述突变的CDTa减去信号肽:Y67A、Y69A和R255A。3mCDTa5指具有下述突变的CDTa减去信号肽:R359A、Y67A和Q307E。3mCDTa6指具有下述突变的CDTa减去信号肽:Y258A、F356A和S345F。3mCDTa7指具有下述突变的CDTa减去信号肽:N342A、Y253A和Y382A。4mCDTa8指具有下述突变的CDTa减去信号肽:S345F、E385Q、E387Q和C2A。得自NAP1株R20291的关于CDTa减去信号肽(即无突变)、3mCDTa1和3mCDTa2的参考序列分别作为SEQ ID NO:38、SEQ ID NO:40和SEQ ID NO:42提供(参见图13)。源自Nap株的关于4mCDTa8的参考序列作为SEQ ID NO:67提供。相对于SEQ ID NO 38、40、42和67中提供的序列,CDTa和用于本文描述的研究的突变形式的CDTa的实际氨基酸序列含有另外N末端甲硫氨酸。关于可用于提供本发明的CDTa序列的参考序列而言,本领域技术人员将认识到来自表达二元毒素的其他毒株的CDTa序列也可以提供用于3mCDTa1、3mCDTa2或4mCDTa8的主链序列,并且此类序列可以具有不同于所提供的NAP1参考株序列的微小变化。在信号肽从天然CDTa氨基酸序列中缺失后,CDTa的残基编号在N末端残基(NAP1参考株中的缬氨酸)处开始(参见图15)。
本文采用的另外缩写包括下述:bm=巨大芽孢杆菌表达系统;bv=杆状病毒表达系统;cd=艰难梭菌表达系统;ec=大肠杆菌表达系统;NAP= NAP1/027/BI17株;VPI= VPI10463株。
“有效量”意指引入患者内以产生所需效应的足够的疫苗或免疫原性组合物,所述所需效应例如在患者中诱导针对艰难梭菌的免疫应答或预防艰难梭菌感染。本领域技术人员应认识到这个水平可以改变。
“MAA”意指无定形羟基磷酸铝硫酸盐佐剂。
如本文使用的,“ISCOM型佐剂”是包含免疫刺激复合物(ISCOM)的佐剂,其包含皂苷、胆固醇和磷脂,其一起形成特征性笼形颗粒,具有促成其功能的独特的球状、笼形结构(关于综述,参见Barr和Mitchell,Immunology and Cell Biology 74: 8-25(1996))。该术语包括用抗原产生且包含在ISCOM颗粒内的抗原的ISCOM佐剂,和不由抗原产生的空心ISCOM型佐剂的ISCOM基质佐剂。
如本文使用的,术语“分离的”指示与自然界中发现的不同的形式。蛋白(多肽)或核酸的不同形式可以是例如与自然界中发现的不同的纯度。在一个实施方案中,该术语指如上下文所述的蛋白或核酸,其基本上或实质上不含在其天然状态下通常与其伴随的组分。
如本文使用的,术语“衍生物”指相对于参考序列(例如本文描述的TcdA、TcdB、CDTa或CDTb序列),具有一种或多种改变的多肽,所述改变可以是氨基酸序列中的变化(包括氨基酸残基的添加和缺失)和/或化学修饰。在优选实施方案中,衍生物与改变前的原始参考序列至少85%、至少90%、至少95%、至少97%、至少99%相同。一般而言,衍生物保留参考序列的活性,例如诱导免疫应答。如本文使用的,术语“衍生物”并不限于野生型或天然参考序列的衍生物,还包括作为参考序列的突变序列的衍生物。在优选实施方案中,参考序列的任何指定突变被维持,但相对于突变的参考序列的改变/修饰,包括在衍生物序列中除参考序列的指定突变外的氨基酸残基处。如本文使用的,术语“衍生物”还包括具有相对于参考核苷酸序列的一种或多种改变的多核苷酸。
在一个实施方案中,衍生物是具有不同于基础序列的氨基酸序列的多肽,所述氨基酸序列通过一个或多个氨基酸取代由所述基础序列衍生。氨基酸取代可以是“保守的”(即氨基酸替换为来自相同氨基酸类别(非极性、极性/中性、酸性和碱性)的不同氨基酸,具有广泛相似的特性的氨基酸,或具有相似结构(脂肪族、含羟基或硫、环状、芳香族、碱性和酸性)的氨基酸),或“非保守的”(即氨基酸替换为不同类型的氨基酸)。广泛地说,更少的非保守取代将是可能的,而不改变多肽的生物活性。本发明的一些实施方案包括这样的衍生物,其包括相对于参考序列被取代的不超过25个氨基酸残基、20个氨基酸残基、15个氨基酸残基、12个氨基酸残基、11个氨基酸残基、10个氨基酸残基、9个氨基酸残基、8个氨基酸残基、7个氨基酸残基、6个氨基酸残基、5个氨基酸残基、4个氨基酸残基、3个氨基酸残基、2个氨基酸残基或1个氨基酸残基。
在另一个实施方案中,衍生物是具有不同于基础序列的氨基酸序列的多肽,所述氨基酸序列通过具有任意组合的一个或多个氨基酸缺失和/或添加由所述基础序列衍生。缺失或添加的氨基酸可以是连续的或单个的残基。在一些实施方案中,相对于参考序列,不超过25个氨基酸残基、不超过20个氨基酸残基、不超过15个氨基酸残基、不超过12个氨基酸残基、不超过10个氨基酸残基、不超过8个氨基酸残基、不超过7个氨基酸残基、不超过6个氨基酸残基、不超过5个氨基酸残基、不超过4个氨基酸残基、不超过3个氨基酸残基、不超过2个氨基酸残基或不超过1个氨基酸残基被缺失或添加。
在另一个实施方案中,衍生物是具有不同于基础序列的氨基酸序列的多肽,所述氨基酸序列通过具有蛋白的一种或多种化学修饰由所述基础序列衍生。化学修饰包括但不限于官能团的修饰(例如烷基化、羟基化、磷化、硫醇化、羧基化(carboxilation)等)、在蛋白合成过程中掺入非天然氨基酸和/或其衍生物以及使用交联剂及其他对多肽施加构象约束的方法。
本领域已知的任何方法均可用于测定参考序列(例如SEQ ID NO:44或67)和衍生物之间的差异程度。在一个实施方案中,序列同一性用于测定关联性。本发明的衍生物将与参考序列(例如SEQ ID NO:44)至少85%相同、至少90%相同、至少95%相同、至少97%相同、至少98%相同、至少99%相同。同一性百分比定义为相同残基数目除以总残基数目且乘以100。如果比对中的序列具有不同长度(由于缺口或延伸),则最长序列的长度将用于计算中,代表总长度的值。有利地,序列与参考序列的关联性可以例如通过比较序列信息进行测定,使用序列分析软件,例如可得自University of Wisconsin Genetics Computer Group (UWGCG)的GAP计算机程序,版本6.0。GAP程序利用如由Smith和Waterman (Adv.Appl.Math. 2:482,1981)修改的,Needleman和Wunsch (J.Mol.Biol. 48:443, 1970)的比对方法。
在另外的实施方案中,杂交用于测定关联性。编码本发明的衍生物的核酸将在高度严格条件下与编码参考序列的核酸杂交。杂交反应的严格性可容易地由本领域普通技术人员决定,并且一般是取决于探针长度、洗涤温度和盐浓度的经验计算。
“药学可接受的载体”是指促进施用的物质。例子包括液体填充剂、稀释剂或其他可以安全地用于全身施用中的物质。取决于具体施用途径,可以使用本领域众所周知的多种药学可接受的载体。这些载体可选自糖、淀粉、纤维素及其衍生物、麦芽、明胶、滑石、硫酸钙、植物油、合成油、多元醇、海藻酸、磷酸盐缓冲溶液包括磷酸盐缓冲盐水、乳化剂、等渗盐水和无热原水。特别地,药学可接受的载体可以含有不同组分例如缓冲液、无菌注射用水、生理盐水或磷酸盐缓冲盐水、蔗糖、组氨酸、盐和聚山梨醇酯。术语例如“生理学可接受的”、“稀释剂”或“赋形剂”可以互换使用。
“PD”意指“剂量后”,并且“PD1”、“PD2”、“PD3”等意指“剂量1后”、“剂量2后”、“剂量3后”等。
附图简述
图1显示了来自艰难梭菌毒素A和B (TcdA和TcdB)的大梭菌毒素的结构。
图2显示了来自多个梭菌属物种的LCT的多重序列比对,突出排列成活性位点的高度保守的残基。
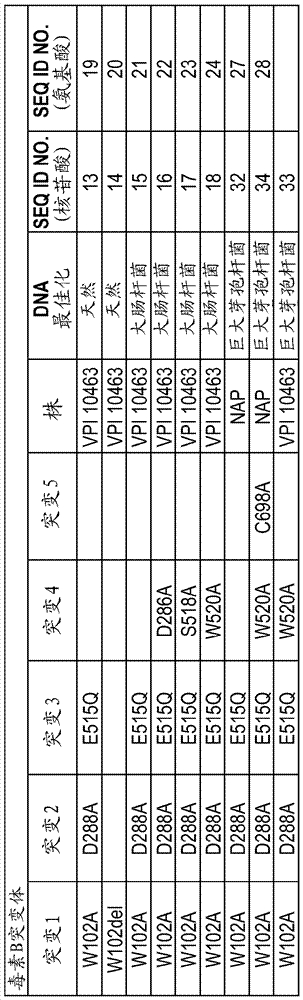
图3提供了本文实施例中所述的重组TcdA (图3A)和TcdB (图3B)突变体(全长和酶结构域)的突变的特性。
图4显示了Tcd突变酶结构域(催化结构域,TcdB突变体,上部;TcdA突变体,下部)在大肠杆菌中的表达。显示了纯化的带六-组氨酸标签的酶结构域通过SDS-PAGE随后为考马斯蓝染色的分析(左)。蛋白的同一性和完整性通过使用抗His MAb的蛋白印迹分析(右)证实。
图5显示了对TcdB和突变重组酶结构域的RhoA葡糖基化测定的结果,如实施例2中所述。数据针对具有100%活性的VPI10463 TcdB进行标准化。显示了关于天然全长TcdB毒素(TcdB)、和在巨大芽孢杆菌中表达的全长3mTcdB (mTcdB)的数据。还显示了下述催化结构域片段的数据:W102缺失(Delta),野生型VPI10463 TcdB (VPI),野生型TcdB (NAP1),三重突变体W102A、D288A、E515Q (Tri),四重突变体W102A、D288A、E515Q、S518A (QuadS),四重突变体W102A、D286A、D288A、E515Q (QuadD),和四重突变体W102A、D288A、E515Q、W520A (Quad W)。所有突变体片段均由VPI1-463株衍生。
图6显示了在基于细胞的测定中与多种TcdA和TcdB构建体相关的残留毒性。含有3、4或5个突变的不同突变体形式的TcdA和TcdB (毒素A突变体:nap_3mTcdA_bm、vpi_4mTcdA_bv、vpi_5mTcdA_bv,以及毒素B突变体:nap_3mTcdB_bm、nap_4mTcdB_ec和nap_5mTcdB_bv)与由艰难梭菌纯化的天然毒素(vpi_TcdA_cd、nap_TcdA_cd、vpi_TcdB_cd和nap_TcdB_cd)、或甲醛失活的天然毒素(vpi_ToxA_cd和vpi_ToxB_cd)、或甲醛失活的突变毒素(vpi_5mToxA_bv和nap_5mToxB_bm)相比较。TC50的增加指示毒素活性的减少。较低的虚线指示提议的安全阈值。
图7显示了TcdA_VPI_4M杆状病毒感染的Sf9昆虫细胞的上清液和全培养液样品通过SDS-PAGE的分析,如实施例6中所述。还显示了不含插入片段的杆状病毒(Bacmin),其充当阴性对照。
图8显示了在仓鼠攻击模型中的保护分析。显示了在多种表达系统中重组产生的,在顶端列出的多种突变型TcdA和TcdB毒素保护仓鼠免于用艰难梭菌孢子的致命攻击的能力的评估。如实施例7中所述,用VPI株攻击仓鼠。TcdA和TcdB突变体的来源如所示的(vpi或nap)。还指示了用于每种突变体的表达系统(cd (艰难梭菌)、bv (杆状病毒)、bm (巨大芽孢杆菌)和ec (大肠杆菌))。
图9提供了VPI_5mTcdA的氨基酸序列(SEQ ID NO: 26),其具有下述突变:W101A、D287A、E514Q、W519A和C700A。
图10提供了NAP_5mTcdB的氨基酸序列(SEQ ID NO: 28),其具有下述突变:W102A、D288A、E515Q、W520A和C698A。
图11显示了如实施例11中所述,在仓鼠攻击模型中进行的研究的存活曲线(随着时间过去的存活百分比)。仓鼠用nap_4mTcdA和nap_4mTcdB的组合(黑色三角形)、或甲醛失活的类毒素A和类毒素B (白色三角形)进行疫苗接种,两者均用ISCOMATRIX®佐剂在MAA上进行配制。还显示的是仅佐剂的对照(黑色圆圈)。动物用241 CFU原型株VPI10463 (图11A)或457 CFU流行株NAP1/027/BI17 (图11B)进行攻击。
图12显示了在仓鼠攻击模型中用NAP1/027/BI17株攻击后,CDTa和CDTb对艰难梭菌疫苗的保护功效的相关贡献。
图13显示了如上文描述的,来自天然CDTa蛋白的CDTa减去信号肽 (图13A,SEQ ID NO:38);3mCDTa1 (图13B,SEQ ID NO:40)和3mCDTa2 (图13C,SEQ ID NO:42)的氨基酸序列。显示了3mCDTa1和3mCDTa2中的突变残基。显示的CDTa序列来自NAP1株R20291。
图14显示了缺少来自天然CDTb蛋白的CDTb信号肽的proCDTb (图14A,SEQ ID NO:44)的氨基酸序列;和缺少来自天然CDTb蛋白的信号肽和活化结构域的CDTb (图14B,SEQ ID NO:46)的氨基酸序列。显示的CDTb序列来自NAP1株R20291。
图15显示了来自R20291参考株的CDTa的天然氨基酸序列(SEQ ID NO: 59)。天然CDTa的信号肽带有下划线。为了本文编号CDTa突变的目的,显示了残基编号1的位置。残基编号1在信号肽后的第一个氨基酸处开始。
图16提供了从残基1到残基719 (TcdA)和717 (TcdB),来自艰难梭菌VPI10463株的天然TcdA (SEQ ID NO:60)和天然TcdB (SEQ ID NO:61)的N末端部分的比对。本文公开的突变的位置带有下划线;然而,显示的残基来自天然序列。
图17提供了关于恒河猴“研究#11”(参见实施例12)的方案概述。每组在所示给药时间经肌内内接受vpi_5mTcdA_bv、nap_5mTcdB_bv、3m_CdtA2_bv和proCdtB_bv。除毒素疫苗组分之外,如所示的每组接受所示量的佐剂或佐剂组合(IMO= IMO2170,MAA= 羟基磷酸铝硫酸盐;和IMX= ISCOMATRIX®(CSL Ltd.))或不接受佐剂。如所示的一些疫苗组合物用甲醛进行处理。
图18显示了对于接受疫苗组合物的不同组的猴(研究#11),针对TcdA、TcdB、CDTa和CDTb的IgG滴度的ELISA测定结果,如三剂程序(图18A)或四剂程序(图18B)中所示,所述疫苗组合物包含与各种佐剂组合的四种组分中的每一种。
图19A-19C分别提供了用于TcdA、TcdB和二元毒素的中和测定。对于图19A和19B,从左到右对于每个剂量显示的条对应于组1-11。Nab测定用于测量抗血清抑制通过天然TcdA或TcdB杀死Vero细胞的能力。对于在研究11中的组,在第0天、第7/10天、第30天和第45天时提供针对TcdA和TcdB的中和抗体滴度(图19A和19B),并且对于二元毒素在第45天时提供(图19C)。
图20A提供了在还原和非还原条件下,CDTa (泳道1和2)和3mCDTa_1 (泳道3和4)的蛋白印迹分析结果。还显示的是大小标记(泳道1左侧)以及蛋白单体和二聚体条带的位置。图20B显示了随着时间过去的3mCDTa1的SEC-UV分析结果。显示了关于聚集体、二聚体和单体的峰。图20C提供了4mCDTa8的SEC MALS分析结果。提供的结果显示了随着时间过去的摩尔质量。对于4mCDTa8可见单个单体峰。
图21显示了用3mCDTa2或4mCDTa8免疫接种的叙利亚仓鼠的保护作用的比较。显示的是与仅佐剂的对照相比较,随着时间过去的存活百分比。
图22提供了与天然CDTa相比较,与胰凝乳蛋白酶活化的proCDTb混合的多种突变形式的CDTa对Vero细胞的细胞毒性测定结果,如实施例9中所述。
发明详述
本发明包括重组TcdA和TcdB突变毒素以及包含所述毒素的疫苗和免疫原性组合物,所述毒素包含显著减少或消除毒素酶促活性的特异性突变,并且在仓鼠攻击模型中有效提供针对用来自原型艰难梭菌株的艰难梭菌孢子的攻击的保护作用。因为TcdA和TcdB是CDI的唯一致病因子,所以这些毒素是用于预防和/或治疗这种疾病的希望靶。用这些毒素对于对象(subject)的疫苗接种可以潜在诱导保护性抗体的产生,所述保护性抗体将中和这些分子在所述对象中的毒性效应。然而,用这些毒素的疫苗接种对于人患者群体是不安全的,除非消除或急剧减少与该蛋白相关的毒性。本申请人已鉴定了显著降低且优选消除毒性的突变及其组合,并且同时含有这些突变的毒素保留诱导免疫原性的能力。
本发明还包括重组CDTa和CDTb毒素蛋白,以及包含与突变型TcdA和TcdB组合的所述毒素的疫苗和免疫原性组合物,其中CDTa毒素蛋白包含显著降低或消除毒素酶促活性的特异性突变。如本文描述的包含所述CDTa和CDTb毒素蛋白的疫苗和组合物在仓鼠攻击模型中有效提供针对用来自流行性艰难梭菌株的艰难梭菌孢子的攻击的保护作用(参见实施例11)。
在结构上,TcdA和TcdB包含在N末端处的酶促结构域(ED)、半胱氨酸蛋白酶结构域(CPD)、易位结构域(TD)和在C末端处的结合结构域(BD)(参见图1)。ED使小Rho GTP酶失活,导致细胞骨架中的急剧和不可逆改变,并且最终导致受累细胞通过细胞凋亡的死亡(Gerhard等人,Journal of Medical Microbiology, 57: 765-770(2008);Spyres等人,Infection and Immunity 71(6): 3294-3301(2003),Busch等人,J.Biol.Chem. 273(31): 19566-19572(1998))。CPD负责ED的自催化的切割。TD介导在BD与细胞表面结合后,ED易位到细胞质内。易位依赖于内吞途径,由此TD被认为在晚期核内体酸化后改变其构象并产生孔,允许ED进入细胞质内。BD是包含梭菌重复性寡肽(CROP)的多价、重复性受体结合结构域。CROP与表面碳水化合物(例如LewisX和LewisY)结合,并且介导毒素的附着和胞吞作用。这些重复是高度免疫原性的并且已成为针对艰难梭菌的疫苗的靶。
LCT家族目前包含5个成员:TcdA、TcdB(艰难梭菌)、TcnA (诺氏梭菌(Clostridium novyi))、TcsL (索氏梭菌(Clostridium sordellii))和TpeL (产气荚膜梭菌)。这些毒素共享相似组构、底物(RhoA GTP酶)、协同底物(UPD-葡萄糖、UPD-NAc葡糖胺)和辅因子(MnII)。它们的相互序列同一性范围为29-76%,然而,催化残基和排列成酶结构域的活性位点的残基几乎是100%保守的(参见图2),支持机械一致论(mechanistic unity)。TcsL、TcnA和TcdB的酶结构域已进行结晶(Reinert等人,J.Mol.Biol. 351: 973-81(200);Ziegler等人,J.Mol.Biol. 377: 1346-56(2007)),并且它们的结构是几乎相同的。另外,近期公开了TcdA的酶结构域的晶体结构(Pruitt等人,J.Biol.Chem. 287: 8013-8020(2012))。显示了TcdA事实上的确具有与其他LCT家族成员相似的结构。具体地,毒素的核心结构(UDP结合和催化活性)与TcdA相比较是保守的,但表面是趋异的。
一般而言,毒素诱变已得到描述。特别地,许多研究描述了与在第102位处的色氨酸残基(W102)和催化三联基序DXD相关的研究,所述DXD是糖基转移酶A家族的标志基序,LCT是所述糖基转移酶A家族的部分。Busch等人作出了关于W102A突变的首次描述(J.Biol.Chem. 275(18): 13228-234(2000)),其集中于TcsL的催化结构域。在该研究中,观察到当与野生型片段相比较时,W102A使糖基化活性降低约1,000倍并且W102Y使活性减少100倍,提示尿嘧啶环的芳香族叠加是这个残基的可能功能。Busch等人简短提及艰难梭菌rTcdB W102A片段,发现该片段在葡糖基化测定中的活性比野生型片段小得多。该作者得出结论,认为这种酶促活性的减少是由于对于协同底物的结合亲和力降低造成的。Teichert等人(Infect and Immun. 74: 6006-6010(2006))公开了在重组、全长TcdA中的W101A突变使在体外系统中(RhoA-葡糖基化,参见下文)的葡糖基转移酶活性减少380倍,以及对Swiss 3T3成纤维细胞的致细胞病变活性减少50倍。使突变型毒素的浓度增加50倍恢复完全细胞毒性。最后,Gerhard等人(Journal of Medical Microbiology 57,765-770(2008))检查全长W101A TcdA突变体的细胞凋亡效应。野生型TcdA能够在1nM浓度时诱导细胞凋亡,而用W101A突变体处理的细胞仅显示出最低限度的细胞凋亡征兆。然而,当TcdA W101A突变体的浓度增加50倍时,细胞的确显示出与野生型相似水平的细胞凋亡。Jank等人(J.Biol.Chem. 282(48): 35222-35231(2007))和Barroso等人(Microbial Pathogenesis16: 297-303(1994))还描述了毒素B的诱变。Sidhu等人(美国专利申请公开号2012/0269841)还描述了特异性突变毒素。
DXD基序对于酶促活性是必需的,因为它涉及MnII、UDP和葡萄糖结合,定向和极化。D288残基直接与MnII结合。D286与一般碱基(general base)(例如水解协同底物的水分子)、UDP和葡萄糖相互作用。Busch等人(Journal of Biological Chemistry 273: 18566-19572(1998))描述了对于TcsL的催化结构域片段中的这个位点的突变。发现用丙氨酸或天冬酰胺交换在第286或288位处的天冬氨酸使在体外测定中的葡糖基化活性降低约5,000倍(Busch等人,1998,同上)。对于D286A/D288A双重突变体未观察到酶促活性。当DXD基序中的天冬氨酸残基变成丙氨酸时,对于艰难梭菌TcdB片段观察到相似的抑制。另外,公开了在高浓度MnII的存在下,TcsL D288A酶的活性增加约20倍,仍导致活性降低数百倍(Busch等人,1998,同上)。高MnII浓度不调节双重突变体的活性。Teichert等人(2006)观察到与野生型TcdA相比较,使用全长D285/287N双重突变体在体外活性中的可比较的6,900倍降低。最后,Gerhard等人(Journal of Medical Microbiology 57: 765-770(2008))证实TcdA D285A/D287A双重突变体不能诱导细胞凋亡。
不希望受理论束缚,基于LCT的活性位点的精密检查,本发明人假设去除在第102位处的大芳环(W102A)将导致对于协同底物的UDP环的结合能的净丧失(通过芳香族叠加),导致具有更低催化周转率的酶。这通过在该位置处用酪氨酸替换丙氨酸仅部分得到恢复,因为酪氨酸的酚环比色氨酸的吲哚双环小得多,并且因此提供更少的结合能。DXD基序中的任一或两个天冬氨酸残基的突变对LCT的酶促活性具有显著影响,因为活性位点缺少参与MnII辅因子结合和极化,和最终UDP-葡萄糖中的O-糖苷键水解的关键残基。因此,推论组合W102/101袋和DXD基序中的突变将产生仅显示出部分的野生型活性的酶,因为它们在其结合协同底物并使协同底物极化的能力中严重受损。
全长遗传失活的TcdA和TcdB及其在巨大芽孢杆菌中的表达描述在WO 2011/068953中。具体地,WO 2011/068953描述了包含突变W101A、D287N和W519A的突变型TcdA,以及包含突变W102A和D288N的突变型TcdB。这些毒素在小鼠中进行评估,但不清楚是否评估动物在疫苗接种后的不利事件。
本发明人已发现由各自含有三个突变的TcdA和TcdB (3mTcdA;W101A、D287A、E514Q)和(3mTcdB;W102A、D288A、E515Q)组成的疫苗在仓鼠和猴中导致无法接受的毒性(局部炎症和肿胀),除非这种疫苗进一步被甲醛失活。在进一步降低毒性的尝试中,将三个另外的突变分开引入来自VPI10463株的TcdA和TcdB的催化结构域内,如本文描述的以及图3A和3B中显示的。具体地,测试了在第E514/515 (TcdA/TcdB)位处的MnII的电子等排结合位点的突变,以及在第S517/518和W519/520 (TcdA/TcdB)位处的突变(其影响O-糖苷键的极化)。另外,还构建了单一氨基酸缺失突变体并且在大肠杆菌中表达。
如在本文中图5和实施例2中所示,当使用RhoA葡糖基化测定分析时,没有明显对于4mTcdB的残留酶活性。尽管添加了这个第四个突变,但在动物免疫接种后仍观察到残留毒性。因此,引入第五个突变C700A/698A,其是半胱氨酸蛋白酶的关键残基,负责从易位结构域处切割催化结构域,从而允许其释放到细胞质内。这个突变的引入导致TcdB毒性另外降低十倍。
突变的TcdA和TcdB类毒素
如上所述,本文描述的实验显示包含3、4或5个突变的艰难梭菌突变毒素导致蛋白毒性至不同水平的降低。为此,本发明部分涉及包含设计为消除蛋白毒性的至少3个突变的分离的和/或纯化的重组TcdA蛋白。它进一步涉及包含设计为消除蛋白毒性的至少3个突变的分离的和/或纯化的重组TcdB蛋白。在本发明的优选实施方案中,本发明的重组TcdA和TcdB蛋白包含设计为消除毒性的至少4个突变或至少5个突变。包含至少3个突变的Tcd蛋白可以进一步通过化学方式例如甲醛失活进行解毒。
在本发明的一个实施方案中,TcdA突变体包含艰难梭菌毒素A蛋白(TcdA),其包含在选自下述的氨基酸残基处的至少3个、至少4个或至少5个突变,或由在选自下述的氨基酸残基处的至少3个、至少4个或至少5个突变组成:W101、D287、E514、D285、S517、W519和C700。在另外的实施方案中,TcdA突变体包含选自下述的至少3个、至少4个或至少5个突变,或由选自下述的至少3个、至少4个或至少5个突变组成:W101A、D287A、E514Q、D285A、S517A、W519A和C700A取代以及W101缺失。
充当用于引入突变的基础的天然艰难梭菌毒素A蛋白序列可以来自任何艰难梭菌株,例如NAP1/027/BI("NAP")或VPI 10463。在本发明的优选实施方案中,TcdA突变体包含得自VPI 10463株的氨基酸序列。可以形成本文TcdA突变体序列的主链的示例性TcdA氨基酸序列(即可以在指定位置处对参考“主链”序列作出本文指定的突变)包括下述中所示的氨基酸序列:SEQ ID NO:94 (VPI10463株,Genbank登记号CAA63564);SEQ ID NO:95 (630株(毒素型0,核糖核酸型012))、SEQ ID NO:96 (SE844 (毒素型IIIa,核糖核酸型080))、SEQ ID NO:97 (R12087 (NAP1))、SEQ ID NO:98 (K14 (毒素型0,核糖核酸型053))、SEQ ID NO:99 (BI6 (NAP1))、SEQ ID NO:100 (BI17 (NAP1))、SEQ ID NO:101 (CH6230 (毒素型IIIc))、和SEQ ID NO:102 (SE881 (毒素型V,核糖核酸型066))。在本发明的范围内还包括保留突变参考序列的突变的突变型TcdA蛋白的衍生物。例如,包含W101A取代或由W101A取代组成的,源自VPI参考株的突变型TcdA蛋白的衍生物将维持W101A取代,但可以包括在其他位置处对VPI氨基酸参考序列的改变/修饰。
本发明的具体实施方案提供了包含氨基酸取代W101、D287A和E514Q,或由氨基酸取代W101、D287A和E514Q组成的TcdA蛋白。进一步的实施方案提供了包含氨基酸取代W101A、D287A、E514Q和W519A,或由氨基酸取代W101A、D287A、E514Q和W519A组成的TcdA蛋白。本发明的另一个具体实施方案是包含氨基酸取代W101A、D287A、E514Q、W519A和C700A,或由氨基酸取代W101A、D287A、E514Q、W519A和C700A组成的TcdA蛋白。
在本文描述的本发明的可替代实施方案中,TcdA蛋白包含突变W101A、D287A、E514Q和D285A,或由突变W101A、D287A、E514Q和D285A组成。另一个可替代实施方案提供了包含突变W101A、D287A、E514Q和S517A,或由突变W101A、D287A、E514Q和S517A组成的TcdA蛋白。在这些可替代实施方案和本文描述的任何其他实施方案的任一个中,进一步突变可以加入TcdA蛋白中,例如C700A突变。在本发明的另外实施方案中,高达2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19或20个进一步突变可以加入本文描述的TcdA实施方案的任一个中。
在本发明的具体实施方案中,突变型TcdA蛋白包含如下述中所示的氨基酸序列、由如下述中所示的氨基酸序列组成,或基本上由如下述中所示的氨基酸序列组成:SEQ ID NO:7、SEQ ID NO:8、SEQ ID NO:9、SEQ ID NO:10、SEQ ID NO:11、SEQ ID NO:12、SEQ ID NO:25或SEQ ID NO:35。
在本发明的可替代实施方案中,TcdA蛋白包含在本文描述的任何氨基酸残基位置处(即W101、D287、E514、D285、S517、W519和C700)的突变,由在本文描述的任何氨基酸残基位置处的突变组成,或基本上由在本文描述的任何氨基酸残基位置处的突变组成,其中指定的氨基酸残基由天然序列中不存在的任何其他氨基酸残基取代,例如在第101位处的色氨酸由除色氨酸外的任何残基取代,在第287位处的天冬氨酸替换为除天冬氨酸外的任何氨基酸残基。例如,本发明的可替代实施方案包括由SEQ ID NO:26限定的TcdA序列的变体(5mTcdA: W101A、D287A、E514Q、W519A、C700A),其包含在W101、D287、E514、W519和C700位处的突变,或由在W101、D287、E514、W519和C700位处的突变组成;其中W101替换为除了色氨酸之外的任何氨基酸,D287替换为除天冬氨酸外的任何氨基酸,E514替换为除谷氨酸外的任何氨基酸,W519替换为除色氨酸外的任何氨基酸,并且C700替换为除半胱氨酸外的任何氨基酸,如SEQ ID NO:70中所示。
在优选实施方案中,所述氨基酸残基(W101、D287、E514、D285、S517、W519和C700)由氨基酸残基取代,所述氨基酸残基与由W101A、D287A、E514Q、D285A、S517A、W519A和C700A限定的突变在结构中相似或处于相同类别中。例如,在优选实施方案中,在第101位处的色氨酸替换为与丙氨酸在相同类别中或具有相似结构的氨基酸。
在本发明的另外实施方案中,突变型TcdA蛋白包含下文所示的氨基酸序列,由下文所示的氨基酸序列组成,或基本上由下文所示的氨基酸序列组成:
。
在另外的实施方案中,突变型TcdA蛋白是本文描述的TcdA蛋白的衍生物,例如包含下述中所示的氨基酸序列的TcdA蛋白的衍生物:SEQ ID NO:7、SEQ ID NO:8、SEQ ID NO:9、SEQ ID NO:10、SEQ ID NO:11、SEQ ID NO:12、SEQ ID NO:25、SEQ ID NO:26、SEQ ID NO:35或SEQ ID NO:70。此类衍生物包含与原始参考序列(例如SEQ ID NO:7、SEQ ID NO:8、SEQ ID NO:9、SEQ ID NO:10、SEQ ID NO:11、SEQ ID NO:12、SEQ ID NO:25、SEQ ID NO:26、SEQ ID NO:35或SEQ ID NO:70)至少85%、至少90%、至少95%、至少97%、至少98%或至少99%相同的氨基酸序列。在进一步的实施方案中,本发明包括这样的TcdA蛋白衍生物,其包含相对于SEQ ID NO:7、SEQ ID NO:8、SEQ ID NO:9、SEQ ID NO:10、SEQ ID NO:11、SEQ ID NO:12、SEQ ID NO:25、SEQ ID NO:26、SEQ ID NO:35和SEQ ID NO:70,对于不超过12个氨基酸残基、不超过11个氨基酸残基、不超过10个氨基酸残基、不超过9个氨基酸残基、不超过8个氨基酸残基、不超过7个氨基酸残基、不超过6个氨基酸残基、不超过5个氨基酸残基、不超过4个氨基酸残基、不超过3个氨基酸残基、不超过2个氨基酸残基或1个氨基酸残基的修饰/改变(例如取代)。
本发明还涉及包含艰难梭菌毒素B蛋白的TcdB突变体,其包含在选自下述的氨基酸残基处的至少3个、至少4个或至少5个突变,或由在选自下述的氨基酸残基处的至少3个、至少4个或至少5个突变组成:W102、D288、E515、D286、S518、W520和C698。在本发明的实施方案中,TcdB突变体蛋白包含选自下述的至少3个、至少4个或至少5个突变,或由选自下述的至少3个、至少4个或至少5个突变组成:W102A、D288A、E515Q、D286A、S518A、W520A和C698A取代以及W102缺失。
充当用于引入突变的基础的天然艰难梭菌毒素B蛋白序列可以来自任何艰难梭菌株,例如NAP1/027/BI ("NAP")株或VPI 10463株。可以形成本文TcdB突变体序列的主链的示例性TcdB氨基酸序列(即可以在指定位置处对参考“主链”序列作出本文指定的突变)包括下述中所示的序列:SEQ ID NO:86 (QCD-63q42 (核糖核酸型001,NAP2)、NCBI参考序列: ZP_05328744.10);SEQ ID NO:87 (NAP8,NCBI参考序列: ZP_06891228.1)、SEQ ID NO:88 (QCD-23m63 (核糖核酸型078,毒素型V)、NCBI参考序列: ZP_05400113.1)、SEQ ID NO:89 (CD196 (NAP1,027,毒素型III)、NCBI参考序列: YP_003213639.1);SEQ ID NO:90 (1470 (核糖核酸型017,毒素型VIII),GenBank登记号: CAA80815.1)、SEQ ID NO:91 (8864 (核糖核酸型036,毒素型X),GenBank登记号: CAC19891.1)、SEQ ID NO:92 (5340,GenBank登记号: AAG18011.1)和SEQ ID NO:93 (VPI10463(核糖核酸型087,毒素型0)、NCBI参考序列: ZP_05349824.1)。本领域技术人员能够在任何艰难梭菌株中的指定位置处引入所需突变。
在本发明的一个实施方案中,TcdB突变蛋白包含得自NAP1株的氨基酸序列(例如TcdB突变蛋白5mTcdB包含SEQ ID NO:28中所示的氨基酸序列,由SEQ ID NO:28中所示的氨基酸序列组成,或基本上由SEQ ID NO:28中所示的氨基酸序列组成)。在可替代实施方案中,TcdB突变蛋白包含得自VPI株的氨基酸序列(例如TcdB突变蛋白5mTcdB包含SEQ ID NO:58中所示的氨基酸序列,由SEQ ID NO:58中所示的氨基酸序列组成,或基本上由SEQ ID NO:58中所示的氨基酸序列组成)。在本发明的范围内还包括保留突变参考序列的突变的突变型TcdB蛋白的衍生物。例如,包含W102A取代或由W102A取代组成的,源自NAP1参考株的突变型TcdB蛋白的衍生物将维持W102A取代,但可以包括在其他位置处对NAP1氨基酸参考序列的改变/修饰。
本发明的具体实施方案提供了包含氨基酸取代W102A、D288A和E515Q,或由氨基酸取代W102A、D288A和E515Q组成的TcdB蛋白。进一步的实施方案提供了包含氨基酸取代W102A、D288A、E515Q和W520A,或由氨基酸取代W102A、D288A、E515Q和W520A组成的TcdB蛋白。本发明的另一个具体实施方案是包含氨基酸取代W102A、D288A、E515Q、W520A和C698A,或由氨基酸取代W102A、D288A、E515Q、W520A和C698A组成的TcdB蛋白。
在本文描述的本发明的可替代实施方案中,TcdB蛋白包含突变W102A、D288A、E515Q和D286A,或由突变W102A、D288A、E515Q和D286A组成。另一个可替代实施方案提供了包含突变W102A、D288A、E515Q和S518A,或由突变W102A、D288A、E515Q和S518A组成的TcdB蛋白。在这些可替代实施方案和本文描述的TcdB的任何其他实施方案的任一个中,进一步突变可以加入TcdB蛋白中,例如C698A突变。在本发明的另外实施方案中,高达2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19或20个进一步突变可以加入本文描述的TcdB实施方案的任一个中。
在本发明的具体实施方案中,突变型TcdB蛋白包含如下述中所示的氨基酸序列、由如下述中所示的氨基酸序列组成,或基本上由如下述中所示的氨基酸序列组成:SEQ ID NO:19、SEQ ID NO:20、SEQ ID NO:21、SEQ ID NO:22、SEQ ID NO:23、SEQ ID NO:24、SEQ ID NO:27、SEQ ID NO:28或SEQ ID NO:58。
在本发明的另外实施方案中,突变型TcdB蛋白包含如下所示的氨基酸序列,由如下所示的氨基酸序列组成,或基本上由如下所示的氨基酸序列组成:
。
在本发明的可替代实施方案中,TcdB蛋白包含在本文描述的任何氨基酸残基位置处(即W102、D288、E515、D286、S518、W520和C698)的突变,由在本文描述的任何氨基酸残基位置处的突变组成,或基本上由在本文描述的任何氨基酸残基位置处的突变组成,其中指定的氨基酸残基由天然序列中不存在的任何其他氨基酸残基取代,例如在第102位处的色氨酸由除色氨酸外的任何残基取代,在第288位处的天冬氨酸替换为除天冬氨酸外的任何氨基酸残基。例如,本发明的可替代实施方案包括由SEQ ID NO:28限定的TcdB序列的变体(5mTcdB: W102A、D288A、E515Q、W520A、C698A),其包含在W102、D288、E515、W520和C698位处的突变,或由在W102、D288、E515、W520和C698位处的突变组成;其中W102替换为除了色氨酸之外的任何氨基酸,D288替换为除天冬氨酸外的任何氨基酸,E515替换为除谷氨酸外的任何氨基酸,W520替换为除色氨酸外的任何氨基酸,并且C698替换为除半胱氨酸外的任何氨基酸,如SEQ ID NO:71中所示。
在优选实施方案中,所述氨基酸残基(W102、D288、E515、D286、S518、W520和C698)由氨基酸残基取代,所述氨基酸残基与由W102A、D288A、E515Q、D286A、S518A、W520A和C698A限定的突变在结构中相似或处于相同类别中。例如,在其包含W102突变的优选实施方案中,在第102位处的色氨酸由与丙氨酸在相同类别中或具有相似的结构的氨基酸取代。
在另外的实施方案中,突变型TcdB蛋白是本文描述的TcdB蛋白的衍生物,例如包含下述中所示的氨基酸序列的TcdB蛋白的衍生物:SEQ ID NO:19、SEQ ID NO:20、SEQ ID NO:21、SEQ ID NO:22、SEQ ID NO:23、SEQ ID NO:24、SEQ ID NO:27、SEQ ID NO:28、SEQ ID NO:58、SEQ ID NO:71。此类衍生物包含与原始参考序列(例如SEQ ID NO:19、SEQ ID NO:20、SEQ ID NO:21、SEQ ID NO:22、SEQ ID NO:23、SEQ ID NO:24、SEQ ID NO:27、SEQ ID NO:28、SEQ ID NO:58或SEQ ID NO:71)至少85%、至少90%、至少95%、至少97%或至少99%相同的氨基酸序列。在进一步的实施方案中,本发明包括这样的TcdA蛋白衍生物,其包含相对于SEQ ID NO:19、SEQ ID NO:20、SEQ ID NO:21、SEQ ID NO:22、SEQ ID NO:23、SEQ ID NO:24、SEQ ID NO:27、SEQ ID NO:28、SEQ ID NO:58和SEQ ID NO:71,对于不超过12个氨基酸残基、11个氨基酸残基、10个氨基酸残基、9个氨基酸残基、8个氨基酸残基、7个氨基酸残基、6个氨基酸残基、5个氨基酸残基、4个氨基酸残基、3个氨基酸残基、2个氨基酸残基或1个氨基酸残基的修饰/改变(例如取代)。
如下文更全面地描述的,本文描述的TcdA和TcdB突变毒素中任一种的组合可用于预防和/或治疗艰难梭菌感染或CDAD的免疫原性组合物和疫苗中,并且包含在本发明中。
本发明还涉及编码本文描述的TcdA和TcdB突变体的核苷酸序列。所述核苷酸序列可以是天然序列,其具有对天然核苷酸序列作出的变化以编码包含所需突变的蛋白,或密码子最佳化的核苷酸序列,其设计为利用更紧密匹配所需表达系统例如大肠杆菌和巨大芽孢杆菌的高表达基因的密码子使用的密码子。另外,可以对核苷酸序列作出其他变化,而不改变编码蛋白,以便增加表达或稳定性,例如可以修饰核苷酸序列的GC含量,可以添加另外的核苷酸用于克隆的目的,以添加纯化标签(例如His标签或GST标签),以添加限制位点等。还可以对核苷酸序列作出变化,以减少CROP区域的重复性以减少重组。
因此,本发明涉及编码TcdA和TcdB突变蛋白的分离的核苷酸序列。编码TcdA的本发明示例性核苷酸序列包含如下述中所示的核苷酸序列,由如下述中所示的核苷酸序列组成,或基本上由如下述中所示的核苷酸序列组成:SEQ ID NO:1、SEQ ID NO:2、SEQ ID NO:3、SEQ ID NO:4、SEQ ID NO:5、SEQ ID NO:6、SEQ ID NO:29、SEQ ID NO:30和SEQ ID NO:31。编码TcdB的示例性核苷酸序列包含如下述中所示的核苷酸序列,由如下述中所示的核苷酸序列组成,或基本上由如下述中所示的核苷酸序列组成:SEQ ID NO:13、SEQ ID NO:14、SEQ ID NO:15、SEQ ID NO:16、SEQ ID NO:17、SEQ ID NO:18、SEQ ID NO:32、SEQ ID NO:33、SEQ ID NO:34和SEQ ID NO:57。本发明的另外实施方案包括核苷酸序列,其为本文公开的核苷酸序列的衍生物的核苷酸序列,例如在高度严格条件下与核酸分子杂交的核苷酸序列,所述核酸分子包含如下述中所示的核苷酸序列,或由如下述中所示的核苷酸序列组成:SEQ ID NO:1、SEQ ID NO:2、SEQ ID NO:3、SEQ ID NO:4、SEQ ID NO:5、SEQ ID NO:6、SEQ ID NO:29、SEQ ID NO:30、SEQ ID NO:31、SEQ ID NO:13、SEQ ID NO:14、SEQ ID NO:15、SEQ ID NO:16、SEQ ID NO:17、SEQ ID NO:18、SEQ ID NO:32、SEQ ID NO:33、SEQ ID NO:34和SEQ ID NO:57。
本发明还涉及包含本发明的核酸和衍生物中的任一种的载体,以及包含本发明的载体中的任一种的宿主细胞。
来自二元毒素的CDTa和CDTb蛋白
本文描述的实验已证实,使用包含大梭菌毒素TcdA和TcdB以及二元毒素(CDTa和CDTb)的疫苗给仓鼠进行免疫诱导比包含单独的TcdA和TcdB的疫苗显著更高水平的针对用流行性NAP1/027/BI株的攻击的保护作用。为此,本发明涉及包含与CDTa和/或CDTb组合的,如上所述的突变型TcdA和/或TcdB蛋白的疫苗和免疫原性组合物。在优选实施方案中,本发明的疫苗和免疫原性组合物包含TcdA、TcdB、CDTa和CDTb蛋白。
因为二元毒素是活性毒素,所以希望制备遗传失活形式的毒素蛋白用于在疫苗或免疫原性组合物中使用。因此靶向毒素的酶促组分CDTa用于遗传失活。设计CDTa的六个三重突变型变体和CDTa的两个四重突变型变体,以使CDTa遗传失活:3mCDTa1 (R302A、E385A、E387D);3mCDTa2 (S345F、E385Q和E387Q);4mCDTa3(R302A、S345F、E385Q、E387Q);3mCDTa4 (Y67A、Y69A、R255A);3mCDTa5 (R359A、Y67A、Q307E);3mCDTa6 (Y258A、F356A、S345F);3mCDTa7 (N342A、Y253A、Y382A);和4mCDTa8 (S345F、E385Q、E387Q、C2A)。
对于3mCDTa1 (R302A、E385A、E387D),基于公开的对CDTa以及Iota毒素(Ia)进行的诱变/毒性研究选择突变,所述Iota毒素与CDTa具有~84.3%序列同一性。对于Iota毒素(Ia)中的催化活性必需的活性位点残基在CDTa中是高度保守的(Perelle等人,Production of a complete binary toxin actin-specific ADP-ribosyltransferase by Clostridium difficile CD196. Infection and Immunity 65(4): 1402-1407(1997);和Sundriyal等人,Structural basis for substrate recognition in enzymatic components of ADP-ribosyltransferase toxin CDTa from Clostridium difficile. Journal of Biological Chemistry 284(42)28713-28719(2009);Perelle等人,Evidence that Arg-295, Glu-378 and Glu-380 are active site residues of the ADP-ribosyltransferase activity of iota toxin. FEBS Letters 395: 191-194.(1996);Nagahama等人,Characterization of the enzymatic component of Clostridium perfringens Iota toxin. Journal of Bacteriology 182(8) 2096-2103(2000),和Tsuge等人,Crystal structure and site directed mutagenesis of the enzymatic components from Clostridium perfringens Iota toxin, J.Mol.Biol 325: 471-483.(2003))。因此,本发明人的假设是CDTa中的等价残基也应是催化活性必需的。
对于3mCDTa2,基于描述CDTa诱变的报道选择突变(S345F、E385Q和E387Q) (Nagahama等人;Tsuge等人;Perelle等人;同上;Sundriyal等人,Structural basis for substrate recognition in enzymatic components of ADP-ribosyltransferase toxin CDTa from Clostridium difficile. Journal of Biological Chemistry 284(42): 28713-28719(2009);和Gulke等人Characterization of the enzymatic component of the ADP-ribosyltransferase toxin CDTa from Clostridium difficile. Infection and Immunity 69(10): 6004-6011(2001))。另外,Sundriyal等人描述了CDTa的晶体结构,并且预测突变S345F将通过位阻取消底物结合。对于Ia S338A (S345的Ia等价物)观察到活性减少~20倍,但对于S338F突变体未观察到活性(Nagahama等人)。Gulke等人报道类似于对于Ia的观察,CDTa中的S345A仅导致活性的少量减少。
设计4mCDTa3 (R302A、S345F、E385Q、E387Q),以组合3mCDTa1和3mCDTa2中的突变。选择突变E385Q和E387Q超过E385A和E387D是因为这些取代的残基更类似于谷氨酸。
对于3mCDTa4 (Y67A、Y69A、R255A),基于此前的报道(Tsuge等人,Proc.Natl.Acad.Sci.USA 105: 7399-7404.(2008)),对参与肌动蛋白结合的残基作出突变,所述报道显示在产气荚膜梭菌的Iota毒素中的肌动蛋白识别和ADP-核糖基化残基的突变导致细胞毒性和酶促活性中的减少。
对于3mCDTa5 (R359A、Y67A、Q307E),使突变Y67A和对于与NAD相互作用的两个残基(R359和S307)的突变组合。在Iota毒素中的R352 (其是与CDTa中的R359等价的残基)显示对于NAD酶活性是必需的(Tsuge等人,(2003),同上)。另外,基于CDTa的结晶结构,预测NAD与R359和Q307相互作用(Sundriyal等人,(2009))。对于3mCDTa6 (Y258A、F356A、S345F),基于等价Iota毒素残基影响ART酶和NAD酶活性的能力(Tsuge等人,(2003))或基于CDTa晶体结构(Sundriyal等人,(2009)),选择另外的突变。
对于3mCDTa7 (N342A、Y253A、Y382A),对被认为涉及Iota毒素中的配体结合、NAD酶活性和/或ART酶活性(Sundriyal等人,(2009);Tsuge等人,(2003))的残基作出突变。另外,选择Y382是因为它是已知对于肌动蛋白特异性ADP-核糖基化毒素重要的保守的关键芳香族残基。
对于4mCDTa8,基于本发明人的野生型重组CDTa以及重组突变型CDTa具有二聚化的趋势的观察,将突变C2A加入3mCDTa2突变体的突变中。通过检查CDTa的序列,本发明人发现单个半胱氨酸残基,本发明人使其突变以尝试减少或消除CDTa的二聚化。解析分析显示这个半胱氨酸突变急剧减少突变型CDTa的寡聚化。参见实施例13和图20C。
相应地,本发明涉及重组二元毒素蛋白A (CDTa),其包含在选自下述的氨基酸位置处的至少两个突变,由在选自下述的氨基酸位置处的至少两个突变组成,或基本上由在选自下述的氨基酸位置处的至少两个突变组成:C2、Y67、Y69、Y253、R255、Y258、R302、Q307、N342、S345、F356、R359、Y382、E385、E385、E387和E387。在本发明的具体实施方案中,CDTa蛋白包含选自下述的高达2、高达3、高达4、高达5、高达6、高达7、高达8、高达9、高达10、高达11、高达12、高达13、高达14、高达15、高达16或高达17个突变或由其组成:C2A、Y67A、Y69A、Y253A、R255A、Y258A、R302A、Q307E、N342A、S345F、F356A、R359A、Y382A、E385Q、E385A、E387Q和E387D。在一些实施方案中,CDTa突变选自C2A、R302A、S345F、E385Q、E385A、E387Q和E387D。在本发明的一些实施方案中,重组CDTa蛋白包含选自下述的一组突变,或由选自下述的一组突变组成:(a) S345F、E385Q和E387Q;(b) R302A、E385A、E387D;(c) C2A、S345F、E385Q和E387Q;(d) R302A、S345F、E385Q和E387Q;(e) Y67A、Y69A和R255A;(f) R359A、Y67A和Q307E;(g) Y258A、F356A、S345F;以及(h) N342A、Y253A和Y382A。在本发明的优选实施方案中,CDTa蛋白缺少信号肽(即缺失天然CDTa的信号肽)。
在本发明的另外实施方案中,本文描述的CDTa突变体蛋白包含对于天然序列的一个进一步突变,例如由突变C2A、S345F、E385q和E387Q加上一个另外突变组成的CDTa蛋白。在本发明的另外实施方案中,高达2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19或20个进一步突变可以加入在本文的任何实施方案中所述的CDTa蛋白的任一种中。
在本发明的示例性实施方案中,上文描述的CDTa蛋白包含如下述中所示的氨基酸序列,由如下述中所示的氨基酸序列组成,或基本上由如下述中所示的氨基酸序列组成:SEQ ID NO:40、SEQ ID NO:42或SEQ ID NO:67。在本发明的另外实施方案中,CDTa蛋白包含具有另外的N末端甲硫氨酸的SEQ ID NO:40、SEQ ID NO:42或SEQ ID NO:67,由具有另外的N末端甲硫氨酸的SEQ ID NO:40、SEQ ID NO:42或SEQ ID NO:67组成,或基本上由具有另外的N末端甲硫氨酸的SEQ ID NO:40、SEQ ID NO:42或SEQ ID NO:67组成。
在本发明的可替代实施方案中,CDTa突变体蛋白包含在本文描述的任何氨基酸残基处的突变(即C2、Y67、Y69、Y253、R255、Y258、R302、Q307、N342、S345、F356、R359、Y382、E385、E385、E387和E387),由在本文描述的任何氨基酸残基处的突变组成,或基本上由在本文描述的任何氨基酸残基处的突变组成,其中指定的氨基酸残基由天然序列中不存在的任何其他氨基酸残基取代,例如在第2位处的半胱氨酸由除半胱氨酸外的任何残基取代,并且在第67位处的Y由除酪氨酸外的任何氨基酸残基取代。例如,本发明的可替代实施方案包括SEQ ID NO:67中所示的CDTa序列的变体(4mCdtA8: C2A、S345F、E385Q和E387Q),其包含在C2、S345、E385和E387位处的突变,或由在C2、S345、E385和E387位处的突变组成;其中C2替换为除了半胱氨酸之外的任何氨基酸,S345替换为除了丝氨酸之外的任何氨基酸,E385替换为除了谷氨酸之外的任何氨基酸残基,并且E387替换为除了谷氨酸之外的任何氨基酸,如SEQ ID NO:72中所示。
在优选实施方案中,所述氨基酸残基(即C2、Y67、Y69、Y253、R255、Y258、R302、Q307、N342、S345、F356、R359、Y382、E385、E385、E387和E387)由氨基酸残基取代,所述氨基酸残基与由C2A、Y67A、Y69A、Y253A、R255A、Y258A、R302A、Q307E、N342A、S345F、F356A、R359A、Y382A、E385Q、E385A、E387Q和E387D限定的突变在结构中相似或处于相同类别中。
在可替代实施方案中,本发明提供了包含上文描述的任何突变或其组合的突变型CDTa蛋白。充当用于引入突变的基础的天然CDTa蛋白序列可以来自任何艰难梭菌株,例如NAP1/027/BI("NAP")株或VPI 10463株(即上文描述的突变可以掺入任何CDTa参考序列内,例如SEQ ID NO:38中提供的CDTa序列,或来自不同株的CDTa序列)。可以形成本文CDTa突变体序列的主链的另外示例性CDTa氨基酸序列(即可以在指定位置处对参考“主链”序列作出本文指定的突变)包括下述中所示的氨基酸序列:SEQ ID NO:73 (AM478株(毒素型IV),GenBank登记号: AEC11575.1)、SEQ ID NO:74 (CD98株(核糖核酸型078,NAP7,毒素型V),GenBank登记号: AEC11569.1)、SEQ ID NO:75 (AE16株(核糖核酸型027,NAP1,毒素型III),GenBank登记号: AEC11560.1)、SEQ ID NO:76 (CD196株(NAP1,毒素型III,027),GenBank登记号: AAB67304.2)和SEQ ID NO:77 (A765株(毒素型IX,NAP1),GenBank登记号: AEC11578.1)。在本发明的另外实施方案中,本发明的重组CDTa蛋白中的任一种是基本上纯化的。
在另外的实施方案中,突变型CDTa蛋白是本文描述的CDTa蛋白的衍生物,例如包含下述中所示的氨基酸序列的CDTa蛋白的衍生物:SEQ ID NO:40、SEQ ID NO:42、SEQ ID NO:67或SEQ ID NO:72。此类衍生物包含与原始参考序列(例如SEQ ID NO:40、SEQ ID NO:42、SEQ ID NO:67或SEQ ID NO:72)至少85%、至少90%、至少95%、至少97%或至少99%相同的氨基酸序列。在进一步的实施方案中,本发明包括这样的CDTa蛋白衍生物,其包含相对于SEQ ID NO:40、SEQ ID NO:42、SEQ ID NO:67和SEQ ID NO:72,对于不超过12个氨基酸残基、11个氨基酸残基、10个氨基酸残基、9个氨基酸残基、8个氨基酸残基、7个氨基酸残基、6个氨基酸残基、5个氨基酸残基、4个氨基酸残基、3个氨基酸残基、2个氨基酸残基或1个氨基酸残基的修饰/改变(例如取代)。本发明的另外实施方案包括具有另外的N末端甲硫氨酸的本文所述CDTa蛋白(例如SEQ ID NO:40、SEQ ID NO:42、SEQ ID NO:67或SEQ ID NO:72)的任何衍生物。
本文还提供了分离的和/或纯化的重组二元毒素蛋白B (CDTb),其中CDTb蛋白缺少(即含有缺失)信号肽(本文称为“pro-CDTb”)。本发明的分离的proCDTb蛋白可以源自表达二元毒素的任何艰难梭菌株。可以用于本文描述的组合物和疫苗中的示例性CDTb序列包括下述中所示的氨基酸序列:SEQ ID NO:78 (CD98株(核糖核酸型078,NAP7,毒素型V),GenBank登记号: AEC11570.1)、SEQ ID NO:79 (K744株(核糖核酸型078,NAP8,毒素型V),GenBank登记号: AEC11585.1)、SEQ ID NO:80 (AM478株(毒素型IV),GenBank登记号: AEC11576.1)、SEQ ID NO:81 (AD667株(毒素型III,NAP1),GenBank登记号: AEC11558.1)、SEQ ID NO:82 (C421株(毒素型IX),GenBank登记号: AEC11564.1)、SEQ ID NO:83 (A765株(毒素型IX,NAP1),GenBank登记号: AEC11579.1)、SEQ ID NO:84 (CD196株(NAP1,毒素型III,027),GenBank登记号: AAB67305.1)、和SEQ ID NO:85 (AE16株(核糖核酸型027,NAP1,毒素型III),GenBank登记号: AEC11560.1)。
在大型多国临床试验和/或流行病学调查中(其中在美国和欧洲对证实的CDI临床病例进行毒株分型),占优势的表达二元毒素的核糖核酸型是027和078。在这些研究中,027和078的流行度范围为11%至40% (Bauer,等人,Lancet 377(9759): 63-73(2011);Gerding等人,Nat Rev Gastroenterol Hepatol. 8(2): 67-8(2011))。在本发明的这个方面的一些实施方案中,proCDTb氨基酸序列源自NAP1株。
在具体实施方案中,proCDTb蛋白,由如下所示的氨基酸序列组成,基本上由如下所示的氨基酸序列组成,或包含如下所示的氨基酸序列:
。
在本发明的另外实施方案中,分离的重组CDTb蛋白进一步缺少(即含有缺失)信号肽且含有CDTb的N末端活化结构域的进一步缺失(参见Barth等人,Microbiol.Mol.Biol.Rev. 68(3): 373-402(2004))。本发明的CDTb蛋白可以源自包含二元毒素的任何艰难梭菌株。在本发明的这个方面的一些实施方案中,CDTb蛋白序列源自NAP1株。在具体示例性实施方案中,CDTb蛋白由SEQ ID NO:46中所示的氨基酸序列组成,或基本上由SEQ ID NO:46中所示的氨基酸序列组成,包含SEQ ID NO:46中所示的氨基酸序列。在另外的实施方案中,CDTb蛋白是SEQ ID NO:46中所示的CDTb蛋白的衍生物,例如包含与SEQ ID NO:46至少85%、至少90%、至少95%、至少97%或至少99%相同的氨基酸序列的CDTb蛋白。在进一步的实施方案中,本发明包括CDTb蛋白衍生物,其包含相对于SEQ ID NO:46,对于不超过12个氨基酸残基、11个氨基酸残基、10个氨基酸残基、9个氨基酸残基、8个氨基酸残基、7个氨基酸残基、6个氨基酸残基、5个氨基酸残基、4个氨基酸残基、3个氨基酸残基、2个氨基酸残基或1个氨基酸残基的修饰/改变(例如取代)。
本发明还涉及编码本文描述的CDTa和CDTb突变体的核苷酸序列。所述核苷酸序列可以是天然序列,其具有对天然核苷酸序列作出的变化以编码包含所需突变的蛋白,或密码子最佳化的核苷酸序列,其设计为利用更紧密匹配所需表达系统例如大肠杆菌、巨大芽孢杆菌或昆虫细胞的高表达基因的密码子使用的密码子。另外,可以对核苷酸序列作出其他变化,而不改变编码蛋白,以便增加表达或稳定性,例如可以修饰核苷酸序列的GC含量,可以添加另外的核苷酸用于克隆的目的,以添加纯化标签(例如His标签或GST标签),以添加限制位点等。
因此,本发明涉及编码本发明的CDTa突变蛋白的分离的核酸。编码突变型CDTa蛋白的示例性核苷酸序列包含如下述中所示的核苷酸序列,由如下述中所示的核苷酸序列组成,或基本上由如下述中所示的核苷酸序列组成:SEQ ID NO:47、SEQ ID NO:48、SEQ ID NO:49、SEQ ID NO:50、SEQ ID NO:51、SEQ ID NO:52、SEQ ID NO:67和SEQ ID NO:68。在本发明的一些实施方案中,核酸是基本上纯化的。本发明的另外实施方案包括核苷酸序列,其为本文公开的CDTa核苷酸序列的衍生物,例如在高度严格条件下与核酸分子杂交的核苷酸序列,所述核酸分子包含如下述中所示的核苷酸序列,或由如下述中所示的核苷酸序列组成:SEQ ID NO:47、SEQ ID NO:48、SEQ ID NO:49、SEQ ID NO:50、SEQ ID NO:51、SEQ ID NO:52、SEQ ID NO:67和SEQ ID NO:68。
本发明还涉及编码如上所述的CDTb和proCDTb蛋白的分离的核苷酸序列。编码CDTb的示例性核苷酸序列包含如下述中所示的核苷酸序列,由如下述中所示的核苷酸序列组成,或基本上由如下述中所示的核苷酸序列组成:SEQ ID NO:53、SEQ ID NO:54、SEQ ID NO:55和SEQ ID NO:56。在本发明的一些实施方案中,核酸是基本上纯化的。本发明的另外实施方案包括核苷酸序列,其为本文公开的CDTb核苷酸序列的衍生物,例如在高度严格条件下与核酸分子杂交的核苷酸序列,所述核酸分子包含如下述中所示的核苷酸序列,或由如下述中所示的核苷酸序列组成:SEQ ID NO:53、SEQ ID NO:54、SEQ ID NO:55和SEQ ID NO:56。
本发明还涉及包含本发明的核酸中的任一种的载体,以及包含本发明的载体中的任一种的宿主细胞。
使用方法
本发明的实施方案还包括突变型TcdA、突变型TcdB、突变型CDTa和CDTb毒素中的一种或多种,或者包含本文描述的所述毒素的组合物,或者包含所述突变型毒素或组合物或由包含所述突变型毒素或组合物组成的疫苗:其(i) 用于在下述中使用,(ii) 用作用于下述的药物或组合物,或(iii) 用于在用于下述中的药物制备中使用:(a) 治疗(例如人体的治疗);(b) 医学;(c) 抑制艰难梭菌复制;(d) 治疗或预防艰难梭菌的感染;(e) 预防艰难梭菌感染的复发;(f) 减少与艰难梭菌感染相关的病理性症状的进展、发作或严重性和/或降低艰难梭菌感染的可能性,或(g) 治疗,预防或者延迟一种或多种艰难梭菌相关疾病的发作、严重性或进展,所述艰难梭菌相关疾病包括但不限于:腹泻、结肠炎和假膜性结肠炎。在这些用途中,突变型毒素、其组合物和/或包含所述突变型毒素或组合物或者由所述突变型毒素或组合物组成的疫苗可以任选与一种或多种抗菌试剂(例如下文描述的抗菌化合物;单克隆抗体或组合疫苗)组合使用。
相应地,本发明提供了用于预防和/或治疗性处理艰难梭菌感染或艰难梭菌相关疾病的方法,其包括给需要治疗的患者施用一种或多种所述毒素、或者包含本发明的所述一种或多种毒素的免疫原性组合物或疫苗。在本发明的这个方面的一些优选实施方案中,给患者施用TcdA蛋白、TcdB蛋白、CDTa蛋白和CDTb毒素蛋白,或者包含TcdA、TcdB、CDTa和CDTb的疫苗或免疫原性组合物。
“患者”(可替代地在本文中称为“对象”)指能够被艰难梭菌感染的哺乳动物。在优选实施方案中,患者是人。患者可以进行预防或治疗性处理。预防性处理提供足够保护性免疫以减少艰难梭菌感染或其效应即CDAD的可能性或严重性。可以进行治疗性处理,以减少艰难梭菌感染或CDAD的严重性或者预防艰难梭菌感染或CDAD的复发。
预防性处理可以使用含有本文描述的突变型或分离的毒素中的一种或多种的药物组合物进行。药物组合物可以施用于普通人群或处于增加的艰难梭菌感染危险中的那些人。
本发明的组合物可以作为包括另外试剂的治疗方案的一部分施用于患者。例如,本发明考虑了与抗生素组合的免疫原性组合物的使用,所述抗生素在治疗或减少CDAD的严重性或进展方面是有效的,例如甲硝唑、万古霉素和非达霉素(fidaxomycin)。在本文中,本发明还提供了治疗方案,其中将本文描述的免疫原性组合物施用于患者,所述患者经历用其他药物或试剂的治疗,所述其他药物或试剂在治疗或减少CDAD的严重性或进展方面是有效的,例如靶向艰难梭菌、毒素A或毒素B的单克隆抗体,例如由Ambrosino等人在美国专利7,625,559中描述的单克隆抗体。在此类情况下,本文描述的疫苗和组合物可以用于预防艰难梭菌感染的复发,或者可以减少艰难梭菌感染或其效应例如CDAD的严重性或发作或进展;用在已由艰难梭菌感染或处于由艰难梭菌感染的高度危险中的患者中。
“需要治疗”的那些人包括已具有艰难梭菌感染的那些人,以及易于具有感染的那些人或需要降低感染可能性的任何个人。具有增加的艰难梭菌感染危险的个人包括经历抗生素治疗例如用某些克林霉素、头孢菌素和氟喹诺酮治疗的那些人,以及老年(≥ 65岁)患者、免疫抑制的患者、其为健康护理工作者的患者、和经历长时间住院治疗的患者。
如上所述,本发明的特点是单独或者与一种或多种另外的抗原组合的,本文描述的突变型或分离的毒素在用于治疗具有艰难梭菌感染或对艰难梭菌感染易感的患者组合物(优选免疫原性组合物或疫苗)中的用途。适当地,所述组合物包含有效量的本文描述的突变毒素蛋白(TcdA、TcdB和CDTa)中的一种或多种,连同CDTb蛋白和药学可接受的载体。在上文描述的本发明的优选实施方案中,所述药物组合物用于人患者中。在可替代实施方案中,药物组合物用于非人患者中。
本发明的组合物可以进一步包含佐剂,例如ISCOM型佐剂或铝佐剂。包含佐剂可以增加通过给患者施用疫苗抗原(即TcdA、TcdB、CDTa和/或CDTb)引起的免疫应答,以便诱导长久保持的保护性免疫。除增加免疫应答之外,佐剂还可以用于减少引起所需免疫应答所需的抗原量或减少临床方案中所需的注射次数,所述临床方案用以诱导持久免疫应答,并且提供免于由艰难梭菌感染引起的疾病的保护和/或诱导由艰难梭菌感染引起的疾病的消退。
可以与本发明的突变和/或天然毒素蛋白结合使用的佐剂包括但不限于含有CpG寡核苷酸,或作用于toll样受体例如TLR 4和TLR9的其他分子的佐剂(关于综述,参见Daubenberger, C.A., Curr.Opin.Mol.Ther. 9(1):45-52(2007);Duthie等人,Immunological Reviews 239(1): 178-196(2011);Hedayat等人,Medicinal Research Reviews 32(2): 294-325(2012)),包括脂多糖、单磷酰脂质A和氨基烷基氨基葡糖苷4-磷酸盐。可用于本发明的组合物中的另外佐剂包括免疫刺激寡核苷酸(IMO;参见例如U.S. 7,713,535和U.S. 7,470,674);T-辅助表位、脂质A及其衍生物或变体、脂质体、磷酸钙、细胞因子(例如粒细胞巨噬细胞集落刺激因子(GM-CSF) IL-2、IFN-α、Flt-3L)、CD40、CD28、CD70、IL-12、热休克蛋白(HSP) 90、CD134 (OX40)、CD137、CoVaccine HT、非离子型嵌段共聚物、不完全弗氏佐剂、趋化因子、霍乱毒素;大肠杆菌不耐热肠毒素;百日咳毒素;胞壁酰二肽、胞壁酰二肽类似物、MF59,SAF、免疫刺激复合物、生物可降解的微球体、聚磷腈;合成多核苷酸。
用于与本文描述的组合物一起使用的另外佐剂是以ISCOM (“免疫刺激复合物”,关于综述参见Barr和Mitchell,Immunology and Cell Biology 74: 8-25(1996);以及Skene和Sutton,Methods 40: 53-59(2006))的特征形式,含有单独或与胆固醇和磷脂组合的皂苷(例如QS21)的佐剂。在本文提供的组合物和方法的具体实施方案中,突变毒素和/或毒素蛋白与ISCOM型佐剂或“ISCOM”组合,所述“ISCOM”是ISCOM基质颗粒佐剂,例如不用抗原制造的ISCOMATRIX® (在本文中也称为“IMX”) (ISCOM®和ISCOMATRIX®是澳大利亚帕克维尔CSL Limited的注册商标)。
另外,基于铝的化合物例如氢氧化铝(Al(OH)3)、羟基磷酸铝(AlPO4)、无定形羟基磷酸铝硫酸盐(AAHS)或所谓的“白矾”(KAl(SO4)·12H2O) (参见Klein等人,Analysis of aluminum hydroxyphosphate vaccine adjuvants by Al MAS NMR.,J.Pharm.Sci. 89(3): 311-21(2000)),可以与本文提供的组合物组合。在本文提供的本发明的示例性实施方案中,铝佐剂是羟基磷酸铝或AAHS,可替代地称为“MAA”。
上述组合物可以用作治疗性或预防性疫苗。相应地,本发明扩展至含有作为活性成分的一种或多种本发明的突变型或分离的毒素(“免疫原性试剂”)或其衍生物的疫苗的产生。任何合适的操作均考虑用于产生此类疫苗。示例性操作包括例如New Generation Vaccines (1997,Levine等人,Marcel Dekker,Inc. New York,Basel,Hong Kong)中所述的那些,所述文献通过引用并入本文。
本发明的突变毒素,包括衍生物毒素,或者包含所述突变毒素或衍生物或天然毒素例如CDTb (单独或与一种或多种免疫原组合)的组合物或疫苗,可以使用本领域众所周知的技术配制且施用于患者。通常的药物施用的指导提供在例如Vaccines, Plotkin和Orenstein编辑,W.B. Sanders Company, 1999;Remington's Pharmaceutical Sciences,第20版,Gennaro编辑,Mack Publishing, 2000;和Modern Pharmaceutics,第2版,Banker和Rhodes编辑,Marcel Dekker,Inc., 1990中。
相应地,本发明提供了用于在患者中诱导针对艰难梭菌感染的保护性免疫应答的方法,其包括给患者施用免疫有效量的本文描述的疫苗或药物组合物中的任一种的步骤。
本文还提供了用于治疗艰难梭菌感染,或用于治疗与艰难梭菌感染相关的任何病理状态的方法,该方法包括给患者施用免疫有效量的本文描述的疫苗或药物组合物中的任一种的步骤。
疫苗和/或组合物可以通过不同途径例如皮下、肌内、静脉内、粘膜、肠胃外或经皮进行施用。皮下和肌内施用可以例如使用针或喷射注射器进行。在本发明的优选实施方案中,疫苗和组合物通过肌内注射施用。
本文描述的组合物可以以与剂型相容的方式并以免疫有效的量进行施用,以治疗和/或减少艰难梭菌感染可能性。在本发明的上下文中,施用于患者的剂量应足以随着时间过去在患者中实现有利应答,例如艰难梭菌水平的减少,或减少由艰难梭菌感染的可能性。待施用的一种或多种免疫原性试剂的数量可以取决于待治疗的对象,包括其年龄、性别、重量和一般健康状况在内。在这点上,需要施用的一种或多种免疫原性试剂的精确量将取决于从业者的判断。在有待于在针对艰难梭菌感染的治疗或预防中施用的免疫原性试剂的有效量的测定中,医生可以评估循环血浆水平、疾病进展和抗艰难梭菌抗体的产生。在任何情况下,本发明的免疫原性试剂的合适剂量可以由本领域技术人员容易地决定。此类剂量可以是纳克至毫克级别的本发明的免疫原性试剂。
合适的给药方案优选考虑本领域众所周知的因素进行测定,包括患者的年龄、重量、性别和医学状况;施用途径;期望的效应;和采用的具体化合物。疫苗组合物可以以多剂的形式使用。
给药的时机取决于本领域众所周知的因素。在最初施用后,可以施用一个或多个另外剂量,以维持和/或加强抗体滴度。
对于组合疫苗,突变型毒素中的每一种可以在一种组合物中一起施用,或在不同组合物中分开施用。如本文描述的本发明的突变型或分离的Tcd或CDT毒素可以与一种或多种所需另外免疫原同时施用。术语“同时”并不限于在完全相同的时间施用治疗剂,相反它意指本文描述的突变型或分离的Tcd或CDT毒素和一种或多种其他所需免疫原依次且在一定的时间间隔内施用于对象,从而使得它们可以一起作用以提供比它们以其他方式施用增加的利益。例如,每种治疗试剂可以在相同的时间施用或以任何次序在不同时间点序贯施用;然而,如果并非在相同的时间施用,则它们应在时间上足够紧密地施用,以便提供所需疗效。每种治疗试剂可以分开地、以任何适当的形式且通过任何合适的途径进行施用。
依照上文描述的本发明的一个方面,提供了包含重组艰难梭菌毒素A (TcdA)蛋白和药学可接受的载体的免疫原性组合物,所述TcdA蛋白包含选自下述的至少3个突变:W101A、D287A、E514Q、D285A、S517A、W519A和C700A。在本发明的这个方面的实施方案中,TcdA蛋白包含选自下述的一组突变:(a) W101A、D287A、E515Q;(b) W101、D287A、E514Q和W519A;(c) W101A、D287A、E514Q和D285A以及(d) W101A、D287A、E514Q和S517A。在进一步的实施方案中,TcdA蛋白进一步包含C700A突变,例如下述突变组(e) W101A、D287A、E514Q、W519A和C700A。在另外的实施方案中,TcdA蛋白包含得自艰难梭菌NAP株或VPI 10463株的氨基酸序列。在进一步的实施方案中,TcdA蛋白包含选自下述的氨基酸序列:SEQ ID NO:7、SEQ ID NO:8、SEQ ID NO:9、SEQ ID NO:10、SEQ ID NO:11、SEQ ID NO:12、SEQ ID NO:25、SEQ ID NO:26和SEQ ID NO:35。
本发明还涉及包含重组艰难梭菌毒素B (TcdB)蛋白和药学可接受的载体的免疫原性组合物,所述TcdB蛋白包含选自下述的至少3个突变:W102A、D288A、E515Q、D286A、S518A、W520A和C698A。在本发明的这个方面的另外实施方案中,TcdB蛋白包含选自下述的一组突变:(a) W102、D288A和E515Q;(b) W102、D288A、E515Q和W520A;(c) W102A、D288A、E515Q和D286A;以及(d) W102A、D288A、E515Q和S518A。在进一步的实施方案中,TcdB蛋白进一步包含C698A突变,例如突变组(e)W102A、D288A、E515Q、W520A和C698A。在再进一步的实施方案中,TcdB蛋白包含得自艰难梭菌NAP株或VPI 10463株的氨基酸序列。在本发明的这个方面的具体实施方案中,TcdB蛋白包含选自下述的氨基酸序列:SEQ ID NO:19、SEQ ID NO:20、SEQ ID NO:21、SEQ ID NO:22、SEQ ID NO:23、SEQ ID NO:24、SEQ ID NO:27、SEQ ID NO:28和SEQ ID NO:58。在本发明的另外实施方案中,上文描述的突变型TcdB毒素蛋白中的任一种可以与说明书全文公开的任何其他突变型毒素蛋白,例如如上所述的突变型TcdA蛋白组合。
因此,本发明的一个实施方案是包含突变型TcdA蛋白、突变型TcdB蛋白和药学可接受的载体的免疫原性组合物。在本发明的这个方面的实施方案中,TcdA蛋白包含选自下述的至少3个突变:W101A、D287A、E514Q、D285A、S517A、W519A和C700A;并且TcdB蛋白包含选自下述的至少3个突变:W102A、D288A、E515Q、D286A、S518A、W520A和C698A。在进一步的实施方案中,本发明的免疫原性组合物包含如本文描述的突变型TcdA、如本文描述的突变型TcdB、突变型CDTa蛋白和CDTb蛋白,其中所述突变型TcdA蛋白包含选自下述的至少两个突变:C2A、Y67A、Y69A、Y253A、R255A、Y258A、R302A、Q307E、N342A、S345F、F356A、R359A、Y382A、E385Q、E385A、E387Q和E387D。在本发明的这个方面的实施方案中,CDTa和CDTb蛋白缺少信号肽。在另外的实施方案中,本文描述的免疫原性组合物中的任一种可以进一步包含佐剂。在再进一步的实施方案中,所述佐剂是ISCOM型佐剂,例如ISCOMATRIX (CSL,Ltd.)和/或铝佐剂,例如MAA。
本发明另外涉及包含来自艰难梭菌的重组二元毒素蛋白A (CDTa)和二元毒素蛋白B (CDTb)和药学可接受的载体的免疫原性组合物,所述CDTa包含选自下述的至少两个突变:C2A、Y67A、Y69A、Y253A、R255A、Y258A、R302A、Q307E、N342A、S345F、F356A、R359A、Y382A、E385Q、E385A、E387Q和E387D。在另外的实施方案中,所述免疫原性组合物包含CDTa蛋白,所述CDTa蛋白由选自下述的高达2、高达3、高达4、高达5、高达6、高达7、高达8、高达9、高达10、高达11、高达12、高达13、高达14、高达15、高达16或高达17个突变组成:C2A、Y67A、Y69A、Y253A、R255A、Y258A、R302A、Q307E、N342A、S345F、F356A、R359A、Y382A、E385Q、E385A、E387Q和E387D。在本发明的一些实施方案中,CDTa蛋白包含选自下述的一组突变,或由选自下述的一组突变组成:(a) S345F、E385Q和E387Q;(b) R302A、E385A、E387D;(c) C2A、S345F、E385Q和E387Q;(d) R302A、S345F、E385Q和E387Q;(e) Y67A、Y69A和R255A;(f) R359A、Y67A和Q307E;(g) Y258A、F356A、S345F;以及(h) N342A、Y253A和Y382A。在进一步实施方案中,CDTa蛋白包含如SEQ ID NO:40或SEQ ID NO:42中所示的氨基酸序列,或由如SEQ ID NO:40或SEQ ID NO:42中所示的氨基酸序列组成。
在再进一步的实施方案中,CDTa蛋白包含下文所示的序列或由下文所示的序列组成:
。
在可替代实施方案中,CDTa蛋白包含具有另外的N末端甲硫氨酸的SEQ ID NO:67,或由具有另外的N末端甲硫氨酸的SEQ ID NO:67组成。在另外的实施方案中,CDTa蛋白包含具有另外的N末端甲硫氨酸的SEQ ID NO:40或SEQ ID NO:42,或由具有另外的N末端甲硫氨酸的SEQ ID NO:40或SEQ ID NO:42组成。
在另外的实施方案中,免疫原性组合物包含本文描述的任何CDTa蛋白和缺少信号肽(即信号肽被缺失)的CDTb蛋白。在再进一步的实施方案中,所述组合物包含如本文描述的任何CDTa蛋白和缺少信号肽和活化结构域两者(即信号肽和活化结构域被缺失)的CDTb蛋白。
本发明还提供了包含下述的免疫原性组合物:任何如本文描述的重组突变型TcdA蛋白、任何本文描述的重组突变型TcdB蛋白、任何本文描述的重组突变型CDTa蛋白和缺少信号肽的CDTb蛋白,以及药学可接受的载体。在另外的实施方案中,所述组合物进一步包含佐剂。在进一步的实施方案中,所述佐剂是铝佐剂和/或ISCOM型佐剂。
在本发明组合物的可替代实施方案中,本文描述的突变型TcdA、TcdB和/或CDTa蛋白可以与天然TcdA、TcdB和/或CDTa蛋白(例如相对于相同株的天然TcdA或TcdB蛋白不包含任何突变,但已解毒,或其毒性已通过化学方法例如通过使用甲醛大幅减少的类毒素蛋白)组合。此类组合物还可以包括CDTb。在另外的可替代实施方案中,本文描述的突变型TcdA、TcdB和/或突变型CDTa蛋白也可以用甲醛处理,以进一步减少或消除残留毒性。
本发明还涉及包含下述的免疫原性组合物:(a) 包含突变W101、D287A、E514Q、W519A,或由突变W101、D287A、E514Q、W519A组成的TcdA蛋白;或包含突变W101、D287A、E514Q和W519A和C700A,或由突变W101、D287A、E514Q和W519A和C700A组成的TcdA蛋白;(b) 包含突变W102A、D288A、E515Q、W520A和C698A,或由突变W102A、D288A、E515Q、W520A和C698A组成的TcdB蛋白;或包含突变W102A、D288A、E515Q和W520A,或由突变W102A、D288A、E515Q和W520A组成的TcdB蛋白;(c) 缺少信号肽且包含突变S345F、E385Q、E387Q和C2A,或由突变S345F、E385Q、E387Q和C2A组成的CDTa蛋白;和(d) 缺少信号肽的CDTb蛋白。在可替代实施方案中,CDTb蛋白缺少信号肽和活化结构域。在进一步的实施方案中,所述组合物进一步包含铝佐剂或ISCOM型佐剂,例如ISCOMATRIX®。在再进一步的实施方案中,本文描述的组合物包含铝佐剂和ISCOM型佐剂。
重组毒素的表达
用于在本发明的方法中使用的本文描述的毒素蛋白或其衍生物重组产生,并且需要时例如用甲醛进行化学修饰。在本发明的优选实施方案中,在所选宿主细胞表达系统中重组表达后,重组毒素不通过化学方法例如甲醛失活进一步失活。多肽的重组表达要求构建含有多核苷酸的表达载体,所述多核苷酸编码目的多肽(即如本文描述的TcdA、TcdB、CDTa或CDTb多肽或其衍生物)。一旦已获得编码目的多肽的多核苷酸,就可以使用本领域众所周知的技术通过重组DNA技术生产用于生产目的多肽的载体。因此,在本文中描述了用于通过表达编码所述多肽的多核苷酸而制备目的多肽的方法。
本领域技术人员众所周知的方法可以用于构建含有目的多肽的编码序列以及适当的转录和翻译控制信号的表达载体。这些方法包括例如体外重组DNA技术、合成技术和体内遗传重组。因此,本发明提供了包含与启动子可操作地连接的编码目的多肽的核苷酸序列的可复制载体。
表达载体通过常规技术转移至宿主细胞,并且经转染的细胞随后通过常规技术在允许核苷酸序列表达的条件下进行培养以产生目的多肽。因此,本发明包括含有与异源启动子可操作地连接的编码目的多肽的多核苷酸的宿主细胞。如本文定义的,术语“宿主细胞”没有想要包括在转基因人类、转基因人胎儿或转基因人胚胎体内的宿主细胞。
在宿主细胞中表达编码TcdA、TcdB、CDTa、CDTb或其突变或衍生形式的重组核苷酸序列后,可以回收一种或多种毒素蛋白,以提供基本上纯化的蛋白用于在本文描述的免疫原性疫苗和组合物中使用。几种蛋白纯化操作是可获得的且适合于使用。重组毒素蛋白可以通过下述的各种组合或单独应用从细胞裂解产物和提取物中纯化:盐分级、离子交换层析、尺寸排阻层析、羟磷灰石吸附层析和疏水作用层析。另外,重组蛋白可以通过使用免疫亲和柱与其他细胞蛋白分离,所述免疫亲和柱由对于毒素蛋白或其衍生物或片段特异性的单克隆或多克隆抗体制备。
多种宿主表达载体系统可以用于表达目的多肽。此类宿主表达系统代表通过其可以产生且随后纯化目的编码序列的媒介物,还代表当用适当的核苷酸编码序列转化或转染时,可以原位表达目的多肽的细胞。这些包括但不限于用含有目的编码序列(即编码本文描述的突变型TcdA、突变型TcdB、CDTa和CDTb蛋白的序列)的DNA表达载体转化的微生物例如大肠杆菌或巨大芽孢杆菌;和用含有目的编码序列的重组病毒表达载体(例如杆状病毒)感染的昆虫细胞系统。
为了获得足够量的艰难梭菌毒素用于商业疫苗生产,可有利地开发在重组表达系统中表达重组TcdA和/或TcdB的方法,其克服与从在培养物中生长的艰难梭菌细菌中分离天然毒素相关的低生产产率和安全问题。因为艰难梭菌是厌氧产孢子生物,所以在培养物中生产Tcd毒素导致极低数量并且因此是昂贵的。另外,毒素的化学失活可以是困难的并且可以导致污染,这导致商业疫苗生产的安全问题。为此,本发明的优选实施方案涉及在如下所述的杆状病毒/昆虫细胞表达系统中重组表达艰难梭菌TcdA、TcdB、CDTa和CDTb蛋白,例如如上所述的TcdA、TcdB、CDTa和CDTb蛋白,或其衍生物的方法,其产生高数量的毒素A和毒素B。
如本文描述的,为了克服在重组TcdA和TcdB表达中遇到的困难,评估了几种表达平台。结果显示大肠杆菌不是最佳的表达平台,因为3mTcdA不能使用这种系统进行表达。另外,尽管3mTcdB在大肠杆菌中成功表达,但由在大肠杆菌系统中表达产生的表达产物含有比在其他表达系统中产生的重组蛋白更高量的降解材料。还评估4mTcdA/B在大肠杆菌中表达的能力。与用3mTcdA获得的结果一致,4mTcdA不能在这种宿主系统中表达,但4mTcdB以高水平(~1.6g/L)制备。还评估了巨大芽孢杆菌表达系统,并且成功制备低至中等量的5mTcdA (~75mg/L)和5mTcdB (~200-700mg/L)。
由于上文描述的亚最佳结果,其对于TcdA和TcdB两种突变体并不一致并且因此不是用于商业表达平台的良好候选物,评估了昆虫细胞/杆状病毒生产平台。在昆虫细胞/杆状病毒系统之前评估了可替代的表达系统,因为认为除非突变毒素完全缺少残留活性,否则由于毒素对Rho蛋白的作用,这些毒素的表达对于昆虫细胞宿主将是致死的。所有真核细胞包括昆虫细胞均产生Rho蛋白,其具有在细胞增殖和基因表达中的作用,并且控制肌动蛋白细胞骨架的组构。因为TcdA和TcdB二者的作用方式是使Rho GTP酶失活,导致细胞变圆和细胞死亡,所以任何残留毒素均可杀死细胞。因此,为了增加使用这种系统的成功毒素表达的机会和增加完整抗原的产率,本发明人测试了5mTcdA和5mTcdB。本文描述的结果显示5mTcdA/B毒素均在这种系统中成功表达。另外,用在这种系统中产生的解毒TcdA进行的稳定性测试显示当在培养基中在27℃下贮存6周时,没有可检测到的降解。
相应地,本发明涉及通过在细胞培养物中的杆状病毒表达系统中重组表达核苷酸序列而生产艰难梭菌毒素A和B的方法,所述核苷酸序列编码包含至少4或至少5个突变的Tcd蛋白。在本发明的优选实施方案中,所述宿主细胞是昆虫细胞。在上文描述的方法的优选实施方案中,在杆状病毒表达系统中表达包含突变W101A、D287A、E514Q、W519A和C700A的TcdA。在另外的优选实施方案中,在杆状病毒表达系统中表达包含突变W102A、D288A、E515Q、W520A和C698A的TcdB蛋白。
因此,本发明提供了生产艰难梭菌TcdA蛋白的方法,其包括在允许核苷酸序列表达以产生TcdA蛋白的条件下,用包含编码如本文描述的TcdA突变蛋白的核苷酸序列的载体转化宿主细胞,其中所述蛋白包含至少3、至少4或至少5个突变。在本发明的某些实施方案中,所述宿主细胞是昆虫细胞,并且所述载体是杆状病毒载体。
本发明进一步提供了生产艰难梭菌TcdB蛋白的方法,其包括在允许核苷酸序列表达以产生TcdB蛋白的条件下,用包含编码如本文描述的TcdB突变蛋白的核苷酸序列的载体转化宿主细胞,其中所述蛋白包含至少3、至少4或至少5个突变。在本发明的某些实施方案中,所述宿主细胞是昆虫细胞,并且所述载体是杆状病毒载体。
本发明进一步涉及通过重组表达编码如本说明书全文所述的CDTa蛋白或CDTb蛋白的核苷酸序列而生产来自艰难梭菌的二元毒素A和二元B的方法。
因此,本发明提供了生产CDTa蛋白的方法,其包括在允许核苷酸序列表达以产生CDTa蛋白的条件下,用包含编码如本文描述的CDTa突变蛋白的核苷酸序列的载体转化宿主细胞,其中所述蛋白包含至少2、至少3或至少4个突变。在本发明的某些实施方案中,所述宿主细胞是昆虫细胞,并且所述载体是杆状病毒载体。
本发明进一步提供了生产CDTb蛋白的方法,其包括在允许核苷酸序列表达以产生CDTb蛋白的条件下,用包含编码如本文描述的CDTb蛋白的核苷酸序列的载体转化宿主细胞。在本发明的某些实施方案中,所述宿主细胞是昆虫细胞,并且所述载体是杆状病毒载体。
本文提及的所有出版物均为了描述且公开可以与本发明结合使用的方法和材料的目的通过引用并入。本文的任何事物均不应解释为承认由于在先发明而使本发明不能先于这些公开。
已参考附图描述了本发明的优选实施方案,应当理解本发明并不限于这些精确实施方案,并且本领域技术人员可以在其中实现多种变化和修饰,而不背离如后附权利要求中定义的本发明的范围或精神。
实施例1
艰难梭菌TcdA和TcdB突变体在大肠杆菌中的表达。
使用Quick Change® XL定点诱变试剂盒(Stratagene Corp.,La Jolla,CA),对源自VPI10463株的艰难梭菌天然A和毒素B编码核苷酸序列进行诱变。如图3中所示,制备具有不同突变组合的几种突变毒素。突变体由大肠杆菌表达且纯化。另外,针对大肠杆菌表达系统进行密码子最佳化的核苷酸序列得自GenScript (GenScript USA,Inc., Piscataway, NJ)。在图3中所示的突变体中,仅大肠杆菌密码子最佳化的三重突变体TcdB (W102A、D288A、E515Q)以全长形式表达。测试了仅由在大肠杆菌中表达的催化结构域组成的另外构建体。
通过SDS-PAGE评估在大肠杆菌中的构建体表达。如图4中所示,Tcd突变酶结构域(催化结构域)在大肠杆菌中的表达分析一般展示~ 60kDa的一个主要重组体条带,和可变的降解产物谱。蛋白的同一性和完整性通过使用抗His MAb的蛋白印迹分析而证实(图4)。
实施例2
用RhoA葡糖基化测定的TcdA和TcdB酶活性测定。
使用RhoA葡糖基化活性测定在体外比较重组TcdA和TcdB酶结构域的毒性。简言之,将所述蛋白以两个不同的浓度水平(1和10 µg/mL)与反应缓冲液混合,所述反应缓冲液含有底物(重组RhoA-GTP酶)、辅因子(MnII)和放射性标记的协同底物(UDP-14C-葡萄糖)。反应在37℃下温育90分钟,并且用冰冷的10%三氟乙酸沉淀蛋白连同任何掺入的14C。沉淀的材料通过过滤回收并且将滤液在液体闪烁计数器中进行定量。
关于TcdB的测定结果显示于图5中。三重突变体W102A、D288A、E515Q催化结构域看起来保留天然催化结构域的约10%葡糖基化活性,这是与W102缺失突变体可比较的活性。QuadS (W102A、D288A、E515Q、S518A)、Quad W (W102A、D288A、E515Q、W520A)和Delta W102没有获得可检测到的酶促活性。对于全长毒素,3mTcdB保留全长毒素~0.5%的活性。在3mTcdB催化结构域片段和全长3mTcdB之间在毒性中的差异可能是由于折叠或纯度中的差异。对于TcdA观察到相似结果。
基于这个数据,将第四个突变引入全长突变型毒素内(W102A、D288A、E515Q、W520A)。4mTcdA和4mTcdB构建体在昆虫细胞(杆状病毒表达系统)和巨大芽孢杆菌中表达。尽管添加了这个第四个突变,但在动物免疫接种后仍观察到残留毒性。因此,引入第五个突变:C698A。C698A是半胱氨酸蛋白酶的关键残基,所述半胱氨酸蛋白酶负责从易位结构域切割催化结构域,允许其释放到细胞质内。这个突变的引入导致TcdB毒性降低另外十倍(图6)。
实施例3
TcdA和TcdB在杆状病毒表达系统中的表达。
为了克服在重组TcdA和TcdB表达中遇到的困难,评估了几种表达平台。最初测试了大肠杆菌;然而,本发明人不能使用这种系统表达3mTcdA,并且尽管3mTcdB被成功表达,但它是部分降解的。本发明人还评估了4mTcdA/B在大肠杆菌中的表达,并且再次,不能在这种宿主系统中成功制备4mTcdA,但4mTcdB以高水平(~1.6g/L)制备。还评估了巨大芽孢杆菌表达系统,并且成功制备低至中等量的5mTcdA (~75mg/L)和5mTcdB (~200-700mg/L)。
基于在这种生物中表达全长TcdA和TcdB的先前例子(Burger等人,Biochem Biophys Res.Commun. 307(3): 584-8(2003);Yang等人,BMC Microbiol. 8:192(2008)),评估了作为表达宿主的巨大芽孢杆菌。这种表达宿主获得增加的3mTcdA和3mTcdB生产(来源序列NAP1株),然而,在不存在甲醛失活的情况下,用这些分子的免疫接种仓鼠和猴揭示这些制剂含有无法接受的高水平的残留毒性。
为了增加完整抗原的产率,本发明人接下来评估了昆虫细胞/杆状病毒生产平台用于5mTcdA和5mTcdB的用途。对于在昆虫细胞中杆状病毒介导的表达,使用由W101A、D287A、E514Q、W519A突变(下文称作TcdA_VPI_4M)或W101A、D287A、E514Q、W519A和C700A突变(下文称作TcdA_VPI_5M)修饰的来自艰难梭菌株VPI10463的TcdA序列。另外,使用由下述突变修饰的来自艰难梭菌株NAP1/027/BI的TcdB序列:W102A、D288A、E515Q、W520A和C698A(下文称作TcdB_NAP_5M)。
如下文对于TcdA变体更详细地描述的,使用Bac-to-Bac表达系统(Invitrogen,Carlsbad,CA)制备重组杆状病毒。DNA序列通过基因合成制备并克隆到pFastBacTM转移载体中的NotI/SphI限制位点内。对于TcdA突变体的克隆,将NotI限制位点引入质粒VPIA_BM_pUC57中TcdA编码序列上游的BsrGI限制位点处。该构建体含有在pUC57中克隆的针对巨大芽孢杆菌最佳化的艰难梭菌毒素A序列(TcdA_BM)。用于插入NotI的寡核苷酸如下:BsrGI-NotI F- 5' G T A C A G C G G C C G C T 3'(SEQ ID NO:36)和BsrGI-NotI R- 5' T C G C C G G C G A C A T G 3'(SEQ ID NO:37)。所得到的VPIA_BM_pUC57_NotI用NotI和SphI消化以获得TcdA_BM基因,随后可以将TcdA_BM基因克隆到NotI/SphI消化的pFastbac1载体内。这产生转移质粒pFbTcdA_BM。
随后将pFastBacTM转化到携带亲本杆粒(bacmid)的感受态DH10BacTM大肠杆菌内,它在其中重组以获得重组杆粒。对于TcdA突变体,转化在两类细菌感受态细胞中完成。第一种是与Bac-to-Bac杆状病毒表达系统一起提供的标准的DH10BacTM大肠杆菌细胞(Invitrogen,Life Technologies),并且第二种是BacdCCDH10细胞(Dr. M van Oers (University of Wageningen,NL,S. Kaba等人Journal of Virological Methods 122(2004)113-118)。作为杆粒在BacdCC细胞中克隆的杆状病毒基因组含有v-组织蛋白酶和壳多糖酶基因的缺失,其被替换为氯霉素抗性基因。转化依照由Bac-to-Bac表达系统提供的手册进行。对于使用BacdCC DH10细胞的重组体选择,将25µg/ml氯霉素加入LB琼脂平板中。从在再划线培养后保持白色的菌落中,使培养物生长并且分离杆粒DNA并用于转染。
随后提取重组杆粒并且用于转染Sf9和Sf21昆虫细胞,在其中生成杆状病毒并且传代1-2次,以制备足够的重组杆状病毒用于实验。依照Invitrogen Bac-to-Bac手册进行转化。使用TCID50测定或商业杆状病毒滴定试剂盒(Expression Systems,LLC,Woodland,CA)进行杆状病毒滴定。除了在纯化过程条件中的轻微差异之外,两种TcdA变体和TcdB变体类似地进行处理。
实施例4
用于昆虫细胞生长和诱导的方法
SF9 (ATCC)、SF21细胞(Kemp Biotechnologies, Inc.),SF21CB (Intervet)和ExpresSF+ (Protein Sciences, Meriden, CT)使用无血清Sf900-II或Sf900-III培养基(Invitrogen, Carlsbad, CA)在摇瓶(培养体积= 1/5 – 1/2的瓶体积)中在27℃下进行培养,并且使用具有1-2"摆度(throw)的振荡器以50-150 RPM进行搅动。将细胞分开并且维持在0.5x106 – 5x106个活细胞/ml之间。对于表达研究,以≥ 0.1单位/活细胞的感染复数(MOI)感染活细胞浓度为1-2x106个细胞/ml的昆虫细胞。从感染后第2-6天监控细胞团块、上清液和全培养液样品中的产物水平。制备样品并且通过SDS-PAGE进行分析,其中TcdA (~300 kDa)和TcdB (~270 kDa)与大多数其他蛋白良好分离。用于纯化的收获通常在感染后第4-5天时进行。
实施例5
从昆虫细胞中收获和纯化TcdA和TcdB蛋白
在收获时全培养液样品中的TcdA水平从100-200 mg/l变化(由于实验变化和昆虫细胞类型差异),并且TcdB水平从200-500 mg/l变化。初始努力旨在从澄清上清液中回收产物。然而,重组蛋白存在于上清液和细胞团块二者中,并且可以从流或组合的全培养液中回收。细胞内的级分在不同昆虫细胞类型之间改变。从细胞团块或全肉汤中有效提取蛋白需要使用典型地用于诱导细胞裂解的浓度的非离子型去污剂(例如0.1-1.0% Triton X-100)。对于纯化,使用硫酸铵沉淀随后为使用强阴离子交换树脂的离子交换层析(IEX)的组合,从澄清上清液可回收至大于70%纯度的全长产物TcdA,伴随低产率。使用使用强阴离子交换树脂的IEX层析随后为羟磷灰石(HA)层析的组合,TcdA和TcdB均可从任何流中回收至>90%纯度,伴随>20%的产率。
实施例6
重组TcdA和TcdB蛋白的表征。
与在大肠杆菌(对于TcdB)和巨大芽孢杆菌中的细胞内表达相比较,TcdA和TcdB蛋白产物的完整性(产物的级分涉及全长的条带)在杆状病毒/昆虫细胞系统中得到最好保存。另外,所需产物与宿主杂质的比对于杆状病毒/昆虫细胞系统是最高的(由于更低的生物量和良好的每细胞生产力),从而使得下游加工更容易。为了举例说明这点,如图7中所示,使用4xLDS作为样品缓冲液(Invitrogen),通过SDS-PAGE (NuPAGE Novex 4-12% Bis-Tris凝胶)分析TcdA_VPI_4M杆状病毒感染的Sf9 (ATCC,Manassas,VA)昆虫细胞培养物的上清液和全培养液样品。作为阴性对照,使用不含插入片段的杆状病毒(Bacmin)。细胞以0.1 TCID50单位/细胞的MOI进行感染并且在感染后五天进行收获。
结果指示通过杆状病毒系统产生的突变毒素材料具有比在其他系统中产生的蛋白更高的质量。它显示出少得多的降解并且更容易纯化。
实施例7
在仓鼠攻击模型中评估TcdA和TcdB突变蛋白的免疫原性
仓鼠模型以下述方式进行。叙利亚仓鼠以三周间隔用疫苗肌内免疫接种四次。随后,在15天的休息期后,通过经口灌饲法给动物施用克林霉素(Cleocin)。发现这种处理破坏结肠微生物群,并且提供对艰难梭菌定殖的易感的窗口期。在Cleocin处理后五天,用致命剂量的艰难梭菌孢子攻击动物。对动物进行评估,每天两次,进行14天的时期。在用多种TcdA和TcdB蛋白免疫接种后,在这个模型中的保护结果在图8中提供。
在仓鼠致命攻击模型中,通过ELISA测量针对天然毒素的血清IgG,和在基于细胞的中和抗体测定中(数据未显示),重组突变毒素(大肠杆菌、巨大芽孢杆菌和杆状病毒/昆虫细胞)也显示在免疫学上是彼此相似的,并且与通过天然毒素的甲醛失活制备的类毒素疫苗相似。
实施例8
CDTa和CDTb的表达
所有二元毒素构建体均使用NAP1/027/BI序列作为模板生成。因为CDTa和CDTb序列对于迄今为止检查的所有毒株是>97%保守的,所以基于NAP1株选择的构建体应生成足够的抗体应答,以提供针对所有二元毒素表达株的保护作用。
编码CDTa的核苷酸序列(减去编码充当信号肽的前42个N末端氨基酸的密码子)通过Genscript (Genscript USA,Inc.,Piscataway,NJ)进行密码子最佳化,用于在大肠杆菌中的最佳表达(SEQ ID NO:47),并且含有侧翼NdeI和XhoI克隆位点并且不含终止密码子。由于排序错误,该序列含有在N末端处的额外甲硫氨酸。将该基因克隆到大肠杆菌表达载体pET30a (EMD Biochemicals USA,Gibbstown,NJ)的NdeI/XhoI位点内,并且与C末端6x组氨酸标签一起表达。表达研究使用大肠杆菌表达宿主BLR(DE3) (EMD Biochemicals)进行。CDTa是高度表达且可溶的。
首先进行尝试,以在大肠杆菌中表达具有6x组氨酸标签的成熟形式的CDTb (减去信号肽和活化结构域)。CDTb对于在大肠杆菌中的表达通过Genscript进行密码子最佳化,具有侧翼NdeI和XhoI克隆位点并且不含终止密码子(SEQ ID NO:53)。将该基因克隆到大肠杆菌表达载体pET30a的NdeI/XhoI位点内,并且与C末端6x组氨酸标签一起表达。表达研究使用大肠杆菌表达宿主BLR(DE3)进行。不幸的是,这种构建体证明是不稳定且不可溶的。发现>90%的CDTb蛋白在~40kD的降解产物中。
使用载体pGEX 6P-1 (GE Healthcare)稳定表达作为GST融合蛋白的CDTb先前由Sundriyal等人描述(Protein Expression and Purification 74: 42-48(2010))。基于这项研究,将上文描述的Genscript最佳化的CDTb进行PCR扩增,使用设计为使N末端克隆位点从NdeI变成BamHI且再引入终止密码子的引物。随后将CDTb与N末端GST标签一起在框内克隆到大肠杆菌表达载体pGEX 6P-1内,如由Sundriyal描述的(SEQ ID NO:54)。CDTb使用BLR(DE3)进行表达。融合蛋白是稳定的、高度表达且可溶的。然而,去除GST标签的蛋白酶解切割导致蛋白可溶性极大降低。
为了尝试增加CDTb的可溶性,构建第二种形式的CDTb。命名为proCDTb的这种构建体包括在原始CDTb构建体中排除的活化结构域,但仍缺少信号肽。proCDTb对于在大肠杆菌中的表达通过Genscript进行密码子最佳化,具有侧翼NdeI和XhoI克隆位点和终止密码子(SEQ ID NO:56)。如上所述将proCDTb克隆到表达载体pGEX 6P-1内。表达研究使用表达宿主BLR(DE3)进行。融合蛋白是稳定的、高度表达且可溶的。再次,去除GST标签的蛋白酶解切割导致蛋白可溶性极大降低。
进行仓鼠研究以评估二元毒素组分的安全性。仓鼠用与下述形式之一的CDTa组合的CDTb进行免疫接种:(1) 甲醛失活的重组天然(无突变) CDTa,(2) 甲醛失活的3mCDTa1 /3mCDTa2,或(3)重组3mCDTa1/3mCDTa2 (无甲醛失活)。动物在免疫接种后就全身和注射部位不利事件(AE)观察几天。在一个研究中,在含有甲醛失活的野生型序列CDTa的所有臂中均观察到显著AE,并且在含有甲醛失活的3mCDTa1和3mCDTa2的臂中观察到少和轻微的AE。在第二个研究中,在接受甲醛失活的野生型序列CDTa的一半臂(2/4)中观察到显著AE,并且再次在含有甲醛失活的3mCDTa1和3mCDTa2以及重组3mCDTa1/3mCDTa2 (无甲醛失活)的臂中观察到少和轻微的AE。研究结果指示单独的甲醛失活方案不足以使二元毒素完全失活。另外,AE可以主要归于CDTa组分,因为两种组分分开的免疫接种仅获得与CDTb相关的较小AE。
实施例9
遗传失活的CDTa蛋白的表达
因为二元毒素是活性毒素,所以还希望制备遗传失活形式的毒素蛋白。因此靶向毒素的酶促组分CDTa用于遗传失活。最初,设计CDTa的两个三重突变型变体,以尝试使CDTa遗传失活:3mCDTa1 (R302A、E385A、E387D)和3mCDTa2 (S345F、E385Q和E387Q)。再次,对基因以类似于CDTa (上文)的方式通过Genscript进行基因的密码子最佳化以用于在大肠杆菌中的最佳表达,但不含额外的甲硫氨酸。3mCDTa1和3mCDTa2也如上文对于CDTa所述进行克隆且表达。3mCDTa1和3mCDTa2是高度表达且可溶的。
设计CDTa的另外三个三重突变型变体和CDTa的两个四重突变型变体,以使CDTa遗传失活:4mCDTa3 (R302A、S345F、E385Q、E387Q);3mCDTa4 (Y67A、Y69A、R255A);3mCDTa5 (R359A、Y67A、Q307E);3mCDTa6 (Y258A、F356A、S345F);3mCDTa7 (N342A、Y253A、Y382A);和4mCDTa8 (S345F、E385Q、E387Q、C2A)。进行细胞毒性研究以比较突变体构建体与天然抗原的毒性。在毒性滴定测定中,CDTa (天然或突变型)和活化的CDTb以1:7 CDTa:CDTb摩尔比组合。毒素混合物在完全MEM培养基中系列稀释。随后将稀释的毒素应用于在384孔板中生长的Vero细胞,并且在加湿CO2温箱中在37℃下温育24小时。将细胞固定,渗透化处理,并且用Alexa 488鬼笔环肽染色,并且使用扫描细胞计量仪(ImageXpress Velos)获得单层的图像。使用GraphPad Prism软件绘制滴定曲线。结果指示相对于天然CDTa,对于所有突变型CDTa蛋白观察到较少的毒性(参见图22)。
实施例10
CDTa/CDTb的纯化
为了获得纯化的二元毒素蛋白用于在动物研究中使用,CDTa和CDTb作为加上亲和标签的抗原从大肠杆菌中纯化。使用亲和柱的纯化过程进行最佳化,以递送足够的产率和纯度用于动物研究。表1描述了用于纯化每种构建体的亲和标签,以及获得的产率和纯度。在一些情况下使用超过亲和层析的另外的纯化步骤,以实现用于进一步研究的足够纯度。
另外,未加上标签的二元毒素蛋白在巨大芽孢杆菌和使用杆状病毒载体在昆虫细胞中表达。通过SDS-PAGE测定来自每种系统的表达滴度。关于3mCDTa的表达滴度(由全裂解物/培养液测量)对于巨大芽孢杆菌为>1 g/L,并且对于杆状病毒系统为大约0.3 g/L。关于CDTb的表达滴度对于巨大芽孢杆菌为0.02 g/L,并且对于杆状病毒系统为0.2 g/L。对于proCDTb,表达滴度在杆状病毒中为>0.5 g/L。
实施例11
疫苗组合物的功效
如实施例7中所述,在仓鼠模型中评价包含二元毒素的疫苗组合物的功效。在一个研究中,仓鼠用疫苗进行免疫接种,所述疫苗包含(a)用羟基磷酸铝佐剂(“Merck Aluminum Adjuvant”或“MAA”)和ISCOMATRIX®佐剂配制的,重组TcdA (Nap_3mTcdA,使用巨大芽孢杆菌表达系统产生)和TcdB (Nap_3mTcdB,使用巨大芽孢杆菌表达系统产生),(b)用MAA和ISCOMATRIX®配制的甲醛失活的TcdA和TcdB (List Biological Laboratories, Inc., Campbell, CA),或(c)仅佐剂对照。结果指示接受重组疫苗的动物完全保护不受来自原型VPI10463株的孢子(95%纯的)的241 CFU/剂的致命攻击(图11A)。
然而,尽管这些动物幸存于用VPI株的攻击的事实,但在所有测试组中的动物中都发生大量重量减轻、腹泻和发病率。另外,艰难梭菌细菌和毒素可以在粪便和盲肠内容物中检测到。此外,在具有上文描述的相同三个测试组((a)–(c))的比较疫苗接种的分离的研究中,重组TcdA + TcdB疫苗不能大量增加受攻击的仓鼠的存活,所述攻击是使用来自高毒性的NAP1/027/BI17株的孢子(95%纯的)的457 CFU剂量的致命攻击(图11B)。
因此,评估另外的疫苗组分,以测定其他抗原是否可以加入疫苗中,以增加疫苗的保护功效并且减少与疾病相关的发病率。为此,进行文献和蛋白组学评估,以鉴定可能可以加入疫苗中的TcdA和TcdB毒素中的九种另外的抗原,并且在仓鼠模型中与TcdA和TcdB组合评价此类另外的抗原。根据这种评价,确定向含有TcdA和TcdB的疫苗添加二元毒素(CDTa和CDTb)一致地增加这种疫苗在NAP1/027/BI攻击后在仓鼠中的保护功效。
本发明人还在仓鼠攻击研究中研究了二元毒素的两个组分(CDTa和CDTb)都是由这种分子介导的保护所需的,还是一种蛋白将是足够的。在这个研究中,仓鼠用下述任一进行疫苗接种:(a)重组TcdA (5mTcdA,使用杆状病毒表达系统产生)、重组TcdB (5mTcdB,使用杆状病毒表达系统产生)和CDTa;(b)重组TcdA (5mTcdA,使用杆状病毒表达系统产生)、重组TcdB (5mTcdB,使用杆状病毒表达系统产生)和CDTb;或(c)重组TcdA (5mTcdA,使用杆状病毒表达系统产生)、重组TcdB (5mTcdB,使用杆状病毒表达系统产生)、CDTa和CDTb。所有组(a)–(c)均用MAA和ISCOMATRIX佐剂进行配制,并且与仅接受佐剂的第四组(d)相比较。结果表明,与TcdA和TcdB组合的CDTa和CDTb可以诱导针对NAP1/027/BI17株(470 CFU/动物)的致命攻击的保护,然而,这种保护是不完全的。相比之下,两种二元毒素蛋白(CDTa和CDTb)与TcdA和TcdB的组合完全恢复了在使用这种组合的先前研究中观察到的保护作用(图12)。
实施例12
疫苗组合物在非人灵长类动物中的免疫原性
11组4-10岁的恒河猴(n=3/组)用不同疫苗组合物进行肌内免疫接种,所述疫苗组合物包含20 µg vpi_5mTcdA_bv、20 µg nap_5mTcdB_bv、5 µg 3m_CdtA2_bv和5 µg proCdtB (“研究11”)。给予十个组的测试组合物还包含选自IMX、MAA、IMO2170的佐剂或这些佐剂的组合,如图17中所示。IMO2170是得自Idera Pharmaceuticals (Cambridge, MA)的toll样受体激动剂。
监测动物在整个研究中的不利事件。在整个研究中以及在给药1 (PD1)、PD2、PD3和PD4后4小时,对清醒动物每天通过应答机获得温度。在给药后4小时观察注射部位,并且每天观察,进行至少7天。在整个研究中,对于大多数猴,温度在正常范围内。对于任何动物在任何时间点均未观察到不利事件。
动物在指定时间点放血,并且使用ELISA测定测量针对所有四种抗原的免疫应答。对于每种疫苗组分,测量在恒河猴中通过疫苗接种诱导的血清抗毒素IgG的浓度。将抗原包被到96孔板上(TcdA以1 µg/ml,TcdB以0.5 µg/ml,CDTa以5 µg/ml和CDTb以5 µg/ml)。遵循一般的ELISA方案,包括洗涤和封闭步骤以及HRP山羊抗人IgG Fcg二抗和用于显色的底物。
对于接受三剂程序(three-dose schedule)的那些组,TcdA ELISA测定结果显示PD-1抗体滴度低于1:100。PD-2和PD-3滴度在组1-4和6 -7中是非常相似的。对于接受四剂程序的组,在第2剂后,抗体滴度仅在接受在IMX-MAA中的疫苗的组中观察到(组11,1:1863滴度)。在组9-10中的PD-3和PD-4滴度相似。另外,对于组11 PD-4观察到滴度下降。
对于接受三剂程序的组,TcdB ELISA测定结果显示PD-1抗体滴度低于1:100。PD-2和PD-3滴度在组2、4和7中是非常相似的。在组1、3、5和6中,在第三个剂后观察到滴度加强(>5X)。对于接受四剂程序的组,抗体滴度仅在接受在IMX-MAA中的疫苗PD-2的组中观察到(组11,1: 8144)。在组9和10中,PD-3和PD-4滴度是相似的。在第4剂后,对于组1观察到滴度加强(~8X),并且对于组11观察到滴度下降(2.5X)。
对于二元毒素组分,相对于CDTa,对于CDTb观察到总体更高的滴度。对于接受三剂程序的那些组,对于CDTa的PD-1抗体滴度低于1:100。在第2剂后,CDTa滴度仅在组5中观察到(1:10850)。在所有组中观察到PD-3滴度,其中在组5 (IMX-MAA-NoF)中观察到1:26,000的最大限度滴度。对于组6(IMO-MAA-F)和组7(无佐剂),观察到< 1:2000的滴度。对于接受四剂程序的所有组的CDTa滴度在直到第2剂后都是无法检测的。在第3剂后,在组8-10观察到滴度加强,并且在组11中观察到滴度下降。
对于接受三剂程序的所有组,对于CDTb的PD-1抗体滴度低于1:100。PD-2和PD-3抗体滴度在接受三剂程序的所有组中均很高。有趣的是,在接受不含佐剂的疫苗的组(组7)中存在滴度加强(5X)。对于接受四剂程序的组,与来自3剂程序组的PD-2滴度相比较,PD-2观察到低抗体滴度。然而,PD-3和PD-4滴度在所有组中是非常相似的。关于接受三剂程序的组的最终IgG ELISA滴度(第47天)在图18A中提供,并且关于接受四剂程序的组在图18B中提供。
还使用Nab测定测量中和抗体。在这个测定中,评价抗血清通过天然TcdA或TcdB杀死Vero细胞的能力。在毒素和抗血清稀释物与Vero细胞一起温育后,将细胞固定并用对F-肌动蛋白特异性的荧光探针进行染色。使用细胞扫描细胞计量仪测定染色的F-肌动蛋白量并随后测定细胞面积。结果指示在30天(PD-3)和45天(PD-3或PD-4,取决于组)后,在所有组中均观察到针对TcdA和TcdB的中和抗体。对于三剂程序,在第45天时,组4 (IMX/MAA-F)具有针对TcdA和二元毒素的最高中和抗体滴度;组1 (IMX-F)具有针对TcdB的最高中和滴度。来自四剂程序的第45天结果显示,在所有三个中和测定中,组8和11 (IMX-F和IMX/MAA-F)具有与MAA佐剂化的(组9和10)相比较的更高滴度。一般而言,用由IMX或IMX/MAA佐剂化的四组分疫苗免疫接种的猴具有最高中和滴度。Nab测定的结果在图19A-19C中提供。
实施例13
CDTa突变体4mCDTa8的解析和免疫原性分析
通过重组突变型3mCDTa2和重组野生型CDTa的蛋白印迹分析,本发明人观察到这些CDTa蛋白具有二聚化的趋势。将8-16% Tris-甘氨酸凝胶以5 µg或2 µg/孔装载,并且在还原和非还原条件下运行。参见图20A。结果指示在还原条件下,CDTa和3mCDTa2蛋白均主要作为单体存在(>97%)。然而,在非还原条件下,大约35-38%的CDTa和大约60-63%的3mCDTa2作为二聚体存在。
通过检查CDTa的序列,本发明人发现单个半胱氨酸残基(C2),本发明人使其突变以尝试减少或消除CDTa的二聚化,因为半胱氨酸可以导致蛋白通过二硫键寡聚化。本发明人因此制备4mCDTa8,在其中本发明人将突变C2A加入3mCDTa2中的突变中。进行SEC分析,以证实半胱氨酸突变是否事实上减少二聚化。对于3mCDTa1的SEC-UV分析,移动相为25 mM NaPi、0.3 M NaCl、0.05% w/v叠氮化钠,pH 6.8,并且使用TSKgel super SW3000 (Tsosoh Bioscience18675)与0.2 ml/分钟的流速进行分析。对于4mCDTa8的SEC-MALS分析,将样品注射到具有保护柱的Wyatt 010S5分析柱上。将柱平衡并且以在0.5 ml/分钟的PBS中运行。样品作为单个定义明确的峰洗脱,具有大约56.9 kDa的计算分子量,其比序列预测的单体分子量高约13%。
3mCDTa1的SEC-UV分析证实相对于单体,更高百分比的材料作为二聚体/更高级的聚集体存在(参见图20B)。相比之下,4mCDTa8的SEC-MALs分析显示蛋白材料几乎唯一地作为单体存在(图20C),证实半胱氨酸突变的确急剧降低CDTa的寡聚化。
为了测定C2A突变是否影响CDTa蛋白组分的免疫原性或在NAP1攻击后提供保护的能力,进行了仓鼠研究,其包括与其他疫苗组分(突变型TcdA/B、proCDTb)组合的3mCDTa 2和4mCDTa8的免疫原性比较。存活曲线显示在两个组中,80%仓鼠幸存于NAP1攻击,表明添加半胱氨酸突变没有不利地影响3mCDTa2的免疫原性(两种抗原在免疫学上是相似的)。
- 还没有人留言评论。精彩留言会获得点赞!