大丽轮枝菌产微菌核相关基因VdGAP的功能分析及其应用的制作方法
本发明涉及大丽轮枝菌(verticilliumdahliae)产微菌核相关基因vdgap的功能分析及其潜在应用的研究,属于植物病理学研究领域。
背景技术:
大丽轮枝菌(verticilliumdahliaekleb.)是引起植物黄萎病的重要土传病原真菌,寄主广泛,可危害多种重要的经济作物和园艺作物。传统的防治技术,如轮作、抗性品种选育、化学药剂等对黄萎病的防治有一定的效果,但是由于大丽轮枝菌具有丰富的遗传多样性,致病力易发生分化,且主要以抗逆性强的微菌核结构在土壤中存活,因而在世界范围内黄萎病的危害依然严重。
黄萎病是系统性侵染的单循环病害,微菌核是大丽轮枝菌在土壤中的主要休眠结构,能够在无寄主植物的土壤中长期存活达14年之久,成为植物黄萎病重要的初侵染来源,在病害循环中起重要作用,其形成的数量及存活情况直接影响着黄萎病的发生危害程度。因此明确微菌核形成和发育的机制对于深入研究该病害流行规律和制定防治措施具有重要意义。对于大丽轮枝菌微菌核的研究过去往往集中于微菌核的形态、大小、分布,微菌核的分离培养技术以及影响微菌核存活的土壤环境因素等。对于其发育的过程在形态学层次也已清晰,最初阶段是由单根或数根菌丝膨大,并产生大量的隔膜。然后,其隔膜的细胞继续增大,成为球状并产生侧芽。最后,在细胞壁内和细胞间沉积黑色素颗粒,黑色素对于形成完整的休眠结构是必须的。而对于其发育的分子机理则研究的很少。
目前报道的与大丽轮枝菌微菌核发育相关的基因不多,并且大多与致病性相关,其中研究最深入的是基因vdh1。通过构建大丽轮枝菌微菌核形成发育的est文库,在1000多个est文库中发现有29个与大丽轮枝菌微菌核形成有关。再进一步研究发现其中的classiihydrophobin基因(vdh1)参与了大丽轮枝菌微菌核的形成,该基因敲除后的突变体形成微菌核的能力大大降低,但其致病性几乎没有改变。通过与gfp融合表达,发现该基因定位于菌丝和分生孢子细胞出现融合的部位,也就是在大丽轮枝菌微菌核初始形成的阶段特异性表达,并且该基因表达受营养物质碳源的调控。另外,通过借鉴在其他病原真菌如核盘菌、立枯丝核菌菌核发育或致病性方面的研究成果,推测可能参与大丽轮枝菌微菌核发育的候选基因,通过基因敲除验证,发现了基因vmk1、vdpkac1、vdsnf1和vgb。其中vmk1编码促分裂原活化蛋白激酶(mitogen-activatedproteinkinase,mapk),该基因敲除后的突变体微菌核形成明显减少,致病力也显著下降。vdpkac1编码camp依赖性蛋白激酶a的一个催化亚基,该基因敲除后的突变体微菌核形成显著增加,而致病力则明显降低。vdsnf1编码sucrosenonfermenting1,该基因调控分解代谢阻遏,敲除后的突变体编码细胞壁降解酶的基因表达下降,致病力显著下降,微菌核形成的能力也显著下降。与其他植物病原真菌相比,大丽轮枝菌微菌核发育的分子机理研究相对滞后。目前已经发现的这些基因仅仅揭开了大丽轮枝菌微菌核发育机制的冰山一角,外界环境和内在因子引发大丽轮枝菌微菌核发育的途径、靶标位点以及作用方式都还需要进一步揭示。
另外大丽轮枝菌属于半知菌,病菌在不同生态条件下形态和致病力变异很大,而且受温度、营养、菌株特性等条件的影响大。在菌落形态上,根据在pda培养基上生长时形成微菌核的多少将大丽轮枝菌分为产生大量黑色微菌核的菌核型、产生少量黑色微菌核的中间型和不产微菌核的菌丝型这三种培养类型。从我们长期监测江苏省棉花黄萎病菌变异的结果来看,2002年调查时,菌丝型菌株在田间所占比例最高,而2008年再调查时,江苏棉花黄萎病菌已从菌丝型为主演变为菌核型为主。这种大丽轮枝菌微菌核产生的群体遗传变异规律也有待于揭示。
因此,我们希望通过分子生物学手段从大丽轮枝菌中直接挖掘更多参与微菌核发育相关的基因,探索其调控的信号网络途径,从分子层面来揭示大丽轮枝菌微菌核发育机理,为作物抗病育种、新型杀菌剂的研发提供新的思路和靶标。对于一部分细菌和真菌,在构建载体时,将其基因组上靶标基因两侧一定长度的同源序列进行克隆,并且将抗生素抗性基因盒连接到两者当中。利用如原生质体转化或农杆菌介导的转化方法,将这一带有同源序列的质粒导入该微生物细胞内后,细胞会在一定比例上启动核酸修复机制,而原靶标基因的核酸序列将被载体上带有抗性基因盒的序列替换,从而产生不含有靶基因的敲除突变体。而通过对敲除突变体和回补突变体表型的观察和检测,可以确定该基因的生物学功能及其在相关领域的应用前景。
技术实现要素:
从大丽轮枝菌v08df1菌株中克隆了vdgap基因,该基因全长为833个碱基,核苷酸序列为seqidno:1,该基因的cds序列具有seqidno:2所示的核苷酸序列。本发明同时提供了vdgap基因编码的蛋白质序列,该蛋白质序列具有seqidno:3所示的氨基酸序列。根据该基因及其侧翼的核酸序列,构建了针对该基因的敲除、回补载体。利用农杆菌介导的转化方法,获得了该基因的敲除突变体和回补突变体。通过对这些转基因材料的表型观察和分析,发现vdgap基因表达的蛋白影响大丽轮枝菌产微菌核的能力。
相关技术路线如下:
1.利用网上共享的生物信息资源,设计核酸引物,构建vdgap基因的敲除载体,并将载体导入大肠杆菌和农杆菌菌株中。
2.利用网上共享的生物信息资源,设计核酸引物,构建vdgap回补载体,并将载体导入大肠杆菌和农杆菌菌株中。
3.将敲除载体利用农杆菌转化的方法导入到大丽轮枝菌v08df1中,获得敲除突变体;将回补载体利用农杆菌转化的方法导入到δvdgap敲除突变体中,获得回补突变体。
4.观察野生型菌株以及敲除和回补突变体在不同培养基中产微菌核的差异。
附图说明
图1.vdgap敲除突变体荧光定量图(v08df1为野生菌株;1c2为t-dna筛选突变体;δvdgap敲除突变体3、δvdgap敲除突变体4、δvdgap敲除突变体5、δvdgap敲除突变体6、δvdgap敲除突变体7为5个敲除vdgap基因的突变体菌株)
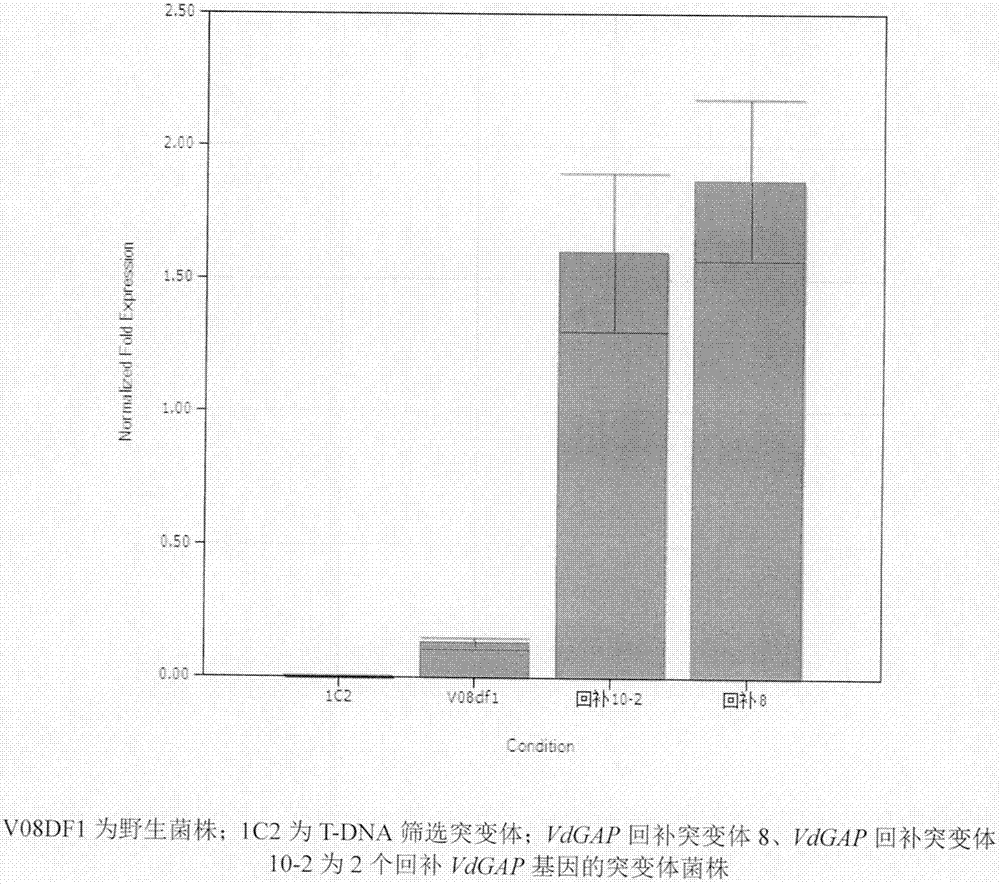
图2.vdgap回补突变体荧光定量图(v08df1为野生菌株;1c2为t-dna筛选突变体;vdgap回补突变体8、vdgap回补突变体10-2为2个回补vdgap基因的突变体菌株)
图3.野生型菌株v08df1和突变体1c2在不同培养基上产微菌核能力差异的分析结果(v08df1为野生菌株;1c2为t-dna筛选突变体)
图4.野生型菌株v08df1以及敲除和回补突变体在不同培养基上产微菌核能力差异的分析结果(v08df1为野生菌株;1c2为t-dna筛选突变体;δvdgap敲除突变体7为敲除vdgap基因的突变体菌株;vdgap回补突变体8、vdgap回补突变体10-2为2个回补vdgap基因的突变体菌株)
具体实施方式
1.vdgap基因的敲除载体和回补载体的构建,并将构建好的载体用化学转化法导入农杆菌agl-1菌株中,分别获得具有vdgap基因敲除功能以及回补功能的agl-1农杆菌各一个,具体流程描述如下:
a.利用网上共享的生物信息资源(参见:http://www.broadinstitute.org/annotation/genome/verticillium_dahliae/multihome.html),分别设计两对核酸引物,即5′侧翼扩增引物上游:ggggacagctttcttgtacaaagtggaaggtgcagataccaccattc;5′侧翼扩增引物下游:ggggactgcttttttgtacaaacttgtgttcacaaggctgtcaagag;3′侧翼扩增引物上游:ggggacaactttgtatagaaaagttgtttcgtaaagagcgagatagaa;3′侧翼扩增引物下游:ggggacaactttgtataataaagttgtgcacaggagtgggtaaaa。利用它们对大丽轮枝菌v08df1的基因组dna进行聚合酶链式反应(pcr)扩增,得到vdgap基因5′和3′两侧各约1kb的序列,然后将目的条带进行纯化。将纯化好的核酸片段各20ng与各60ng的pa-hyg-oscar和poscar质粒(购于fungalgeneticsstockcenter)混合,并加入1μlbp
b.利用网上共享的生物信息资源(参见:http://www.broadinstitute.org/annotation/genome/verticillium_dahliae/multihome.html),分别设计两条核酸引物,即上游:gacctgcaggcatgcaagctttcatgtacatgtacagggaccga;下游:acgacggccagtgccaagctttctttatcttgaatttgatgacgaact。利用该引物对大丽轮枝菌v08df1基因组dna进行pcr扩增,获得包括vdgap基因启动子,orf以及终止子在内的约2300bp的核酸片段。将该序列纯化,利用clonexpresstmii重组反应体系(购于vazyme生物技术公司)将该片段连接至用hindiii线性化的1300-ble载体(经本实验室改造)上,获得vdgap回补质粒。测序验证正确后,将该质粒导入农杆菌agl-1中,获得具有回补功能的农杆菌菌株。
2.vdgap基因的敲除突变体和回补突变体的获得
利用上一步骤中获得的具有vdgap基因敲除功能以及回补功能的agl-1农杆菌菌株分别对野生型v08df1菌株和获得的δvdgap敲除突变体进行转化,具体的转化步骤如下:大丽轮枝菌转接至液体pda中,28℃,150rpm培养3天;含有t-dna双元载体的农杆菌agl-1挑取单克隆接至液体mm培养基当中(现配现用,1升超纯水中溶解:2.05gk2hpo4,1.45gkh2po4,0.15gnacl,0.5gmgso4·7h2o,0.1gcacl2·6h2o,0.0025gfeso4·7h2o,0.5g(nh4)2so4和2.0gglucose),28℃,150rpm培养2天。之后将农杆菌用im培养基(在90mlmm培养基中加入0.5ml甘油和0.7808gmes,ph值调至5.3-5.5,并用mm定容至100ml)稀释到od600=0.15,预培养6小时,然后与等体积的、浓度为1.0×105孢子/ml的大丽轮枝菌孢子液混合。将混合物取适量涂布于灭菌的、置于cm固体培养基(与im培养基相似,但葡萄糖浓度为im培养基的一半,另外还包含200μm乙酰丁香酮)上的硝酸纤维素膜(0.45μm孔径)上,于28℃共培养48小时。之后,硝酸纤维素膜被转移至pda固体培养基上【200μmcefotaxime和50μg/mlhygromycinb(筛选vdgap敲除突变体)或25μg/mlbleomycin(筛选vdgap回补突变体)】。相同条件培养5-7天后,可见小的转化子菌落,用接种针分别挑取其孢子进行培养,经过单孢分离后以终浓度25%甘油,-70℃保存。
3.大丽轮枝菌v08df1野生菌株以及vdgap敲除和回补突变体在不同培养基中产微菌核的差异观察
获得敲除突变体和回补突变体后,将上述各菌株,挑取菌丝至1ml无菌水中,振荡均匀后,吸取10μl滴加至固体pda、查氏、添加棉花碎根的查氏(称取15g棉花幼苗的根须,研磨后,溶于70mlddw,25℃振荡1h,滤纸过滤,固体残渣分装2mleppendorf管(管盖已开孔),高温高压灭菌,滤出液用0.22μm滤器过滤至10ml无菌管中,使用时,每100ml培养基加入2管残渣和1管滤出液)三种培养基中,25℃培养,观察微菌核产生情况。
- 还没有人留言评论。精彩留言会获得点赞!