增强造血干细胞植入过程的材料和方法与流程
本申请是申请号为200980149522.5、发明名称为“增强造血干细胞植入过程的材料和方法”的中国专利申请的分案申请,该母案申请是2009年11月6日提交的pct国际专利申请pct/us2009/063654进入中国国家阶段的申请。
优先权要求
本申请要求2008年11月6日提交的美国临时专利申请no.61/112,018的权益,其以参考的方式整体并入本文。
政府权利声明
本发明开发的一部分是在政府资助下进行的,所述资助来自nih基金号hl069669,hl079654和dk07519。美国政府拥有本发明中的某些权利。
本文公开的一些方面和实施方案涉及增强造血干和祖细胞植入的材料和方法。
背景技术:
造血干和祖细胞(hematopoieticstemandprogenitorcell,hspc)移植是获准用于治疗某些恶性与非恶性血液病和代谢疾病的疗法。用于移植的hspc来源包括骨髓、动员的外周血和脐带血(umbilicalcordblood,ucb)(goldman和horowitz,2002;fruehauf和seggawiss,2003:broxmeyer,等,2006)。医生常规地进行骨髓、动员的外周血干细胞和脐带血的移植。这些过程要求从健康的正常供体或者从产生给定症状之前或处于恢复之中的患者收获足够数量的造血干和祖细胞。接着将所收获的材料施用给其造血系统以及推定地其患病或异常组织和细胞已被清除的患者。移植之后,移植的干细胞移动或“归巢”到适宜的骨髓微环境小生境(niche),在此小生境中存活、增殖并生成新的干细胞,这一过程被称作自我更新(porecha等,2006;broxmeyer,2006;hall等,2006)。所述细胞也分化成为谱系限制性祖细胞和成熟细胞,从而恢复了接受者的健康所必需的具有造血功能的造血系统。祖细胞经常存在于所移植的材料中,而且可能是这些移植物产生成熟细胞所必需的。然而,因为祖细胞并不是干细胞,不能自我更新,所以它们仅在有限的时间内参与移植治疗。
因为移植的过程对移植材料产生压力,所以成功的移植需要移植足够的细胞以应对该过程中杀死或破坏的细胞。这为脐带血移植物的移植带来了巨大的问题,因为这些移植物仅包含非常有限数目的干细胞。因此,脐带血移植物经常不能成功地用于移植成年人。类似地,10-25%的患者和正常供体无法动员足够的细胞以用于移植过程。在一些患者人群中,特别是用某些化学治疗药物治疗的人群,50%以上的患者无法动员。一般来说,可移植的细胞越多,移植成功的可能性就越大。例如,现今最好的做法建议外周血干细胞移植过程通常要求每公斤接受患者体重施用最少大约2百万个cd34+细胞,可获得的以及随后移植的cd34+细胞越多,患者的结局就越好(pulsipher,2009)。
干细胞数量不足、无法迁移/归巢到适宜的骨髓小生境或造血干和祖细胞的移植效率及自我更新差可不利地影响移植结局,其由多步骤的群体恢复(repopulation)过程来度量。尝试了许多方法在离体背景下扩增分离移植物中人造血干和祖细胞的数量,但是成果非常有限。提高hspc移植效率的策略需要克服医疗行业所面临的这一挑战。本发明中公开的一些方面和实施方案满足了这一需求。
技术实现要素:
本公开内容的一些方面涉及对造血干和祖细胞收获和/或植入的增强,其中一些方面包括但不仅限于离体存活、自我更新和归巢到适宜的骨髓小生境,以提高造血干和祖细胞治疗的成功率。
本公开内容的一些方面包括增加收获自供体的具有长期群体恢复能力的造血干和祖细胞数量的方法。这些方法中的一些包括如下步骤:鉴定抑制前列腺素(例如前列腺素e)生物合成的化合物,或拮抗参与前列腺素应答的至少一种前列腺素受体的化合物;以及在从供体外周血或骨髓收获造血干和祖细胞之前向供体提供药学有效量的所述化合物。在一个实施方案中,前列腺素e生物合成抑制剂和/或前列腺素e受体拮抗剂的施用与一种或多种临床上获准的造血干和祖细胞动员剂(例如,粒细胞集落刺激因子(granulocyte-colonystimulatingfactor,g-csf))联用,来增加可通过血液分离法(apheresis)收集的造血干和祖细胞的数量以用于造血移植物移植。在一个实施方案中,化合物选自环加氧酶抑制剂,包括例如吲哚美辛(2-{1-[(4-氯苯基)羰基]-5-甲氧基-2-甲基-1h-吲哚-3-基}乙酸)或其可药用盐。在另一些实施方案中,环加氧酶抑制剂选自:阿司匹林、布洛芬、塞来昔布、罗非昔布、美洛昔康、艾托考昔、伐地考昔、萘普生、双氯芬酸、利克飞龙、依托度酸、酮咯酸或其可药用盐。在一些实施方案中,环加氧酶抑制剂对cox-1和cox-2二者都有作用,通常情况下优先作用于cox-2。在一些实施方案中,可使用的另一种化合物是美洛昔康。
本公开内容的另一些方面包括增强收获的造血干和/或祖细胞(hematopoieticstemand/orprogenitor)移植物在接受者中长期群体恢复能力的方法。在接受者的造血系统有缺陷的情况下这是特别有用的。所述方法包括如下步骤:a)从供体中收获移植物,其中所述供体已经过有效量的抑制前列腺素e2生物合成的化合物和/或前列腺素e2受体拮抗剂的处理;b)将移植物与有效量的前列腺素e2或者一种或多种其衍生物离体接触;以及c)将处理过的移植物施用于接受者。在一个实施方案中,所述方法还包括下述步骤:向移植接受者提供有效量的前列腺素e2或一种其衍生物或任何具有pge2活性的分子,从而增强移植物向适宜治疗小生境的归巢。
本公开内容的另一些方面包括提高干细胞中病毒转导效力的方法。这些方法中的一些可包括如下步骤:提供含有至少一个待转导目的基因的病毒载体;提供至少一个经有效量的前列腺素e2或其衍生物离体处理过的干细胞,以及将所述病毒载体转染进经pge2或其衍生物处理的干细胞。
一些实施方案包括增强造血干和/或祖细胞动员的方法,包括如下步骤:鉴定造血干和/或祖细胞的来源;提供降低pge2生物合成和/或活性的化合物;和将造血干和/或祖细胞的来源与有效量的所述降低pge2生物合成和/或活性的化合物相接触。在一些实施方案中,所述降低pge2活性的化合物为非甾体抗炎化合物,其中所述非甾体抗炎化合物作用于环加氧酶-1和环加氧酶-2。在一些实施方案中,所述非甾体抗炎化合物主要作用于环加氧酶-2。在一些实施方案中,所述非甾体抗炎化合物选自:阿司匹林、塞来昔布、罗非昔布、艾托考昔、伐地考昔、布洛芬、萘普生、双氯芬酸、依托度酸,酮咯酸和利克飞龙。在另一些实施方案中,所述非甾体抗炎化合物为吲哚美辛,而在又一些实施方案中所述非甾体抗炎化合物为美洛昔康。
在一些实施方案中,向患者施用一段时间的所述非甾体抗炎化合物,所述时间段与使用至少一种增强造血干和祖细胞动员的额外化合物的共治疗相重叠。在一些实施方案中,所述增强造血干和祖细胞动员的化合物选自:g-csf和普乐沙福(plerixafor)。在一些实施方案中,向患者施用至少3天的所述非甾体抗炎化合物。
另一些实施方案包括增强来自供体的造血干和/或祖细胞动员的方法,其包括如下步骤:提供作为至少一种pge2受体之拮抗剂的化合物;以及在从供体收获造血干或祖细胞之前向所述造血干或祖细胞供体施用有效量的所述化合物。在一些实施方案中,至少一种pge2受体的拮抗剂选自:n-[[4′-[[3-丁基-1,5-二氢-5-氧代-1-[2-(三氟甲基)苯基]-4h-1,2,4-三唑-4-基]甲基][1,r-联苯基]-2-基]磺酰基]-3-甲基-2-噻吩甲酰胺(l-161,982)和4-(4,9-二乙氧基-1,3-二氢-1-氧代-2h-苯并[f]异吲哚-2-基)-n-(苯磺酰基)-苯乙酰胺(gw627368x)。
而另一些实施方案包括将造血干和或祖细胞(hematopoieticstemandorprogenitor)植入接受者,其包括如下步骤:从来源收获包含造血干和祖细胞的细胞群,所述来源已经过至少一种降低来源中pge2生物合成和/或活性的化合物处理;将所述造血干细胞群与增加pge2活性的化合物离体接触;以及将与所述增加pge2活性的化合物离体接触的所述造血干和祖细胞移植入接受者。在一些实施方案中,所述造血干细胞抽取自骨髓供体。而在另一些实施方案中,所述造血干细胞收获自从血液供体抽取的血液样品。而在又一些实施方案中,所述细胞抽取自脐带或胎盘。
一些实施方案包括增加造血干和/或祖细胞植入率的方法,其包括如下步骤:提供具有pge2活性的化合物;和将所述具有pge2活性的化合物与造血干和/或祖细胞群离体相接触。在一些实施方案中,具有pge2活性的化合物选自:任何e系列的前列腺素或e系列前列腺素的任何衍生物,例如pgei、pge2、pge3或pge1、pge2、pge3的二甲基衍生物,包括例如二甲基16,16-二甲基pge2。在一些实施方案中,所述具有pge2活性的化合物与所述造血干和/或祖细胞群接触至少1小时。一些实施方案包括用基本上没有pge2活性的缓冲液将与所述具有pge2活性的化合物相接触的所述造血干和/或祖细胞冲洗至少一次的步骤。而另一些实施方案还包括如下步骤:将与所述具有pge2活性的化合物相接触的所述造血干和/或祖细胞引入患者。
可参照下列非限制性附图、描述和权利要求更好地理解本发明的这些和其它特征、方面和优点。
附图说明
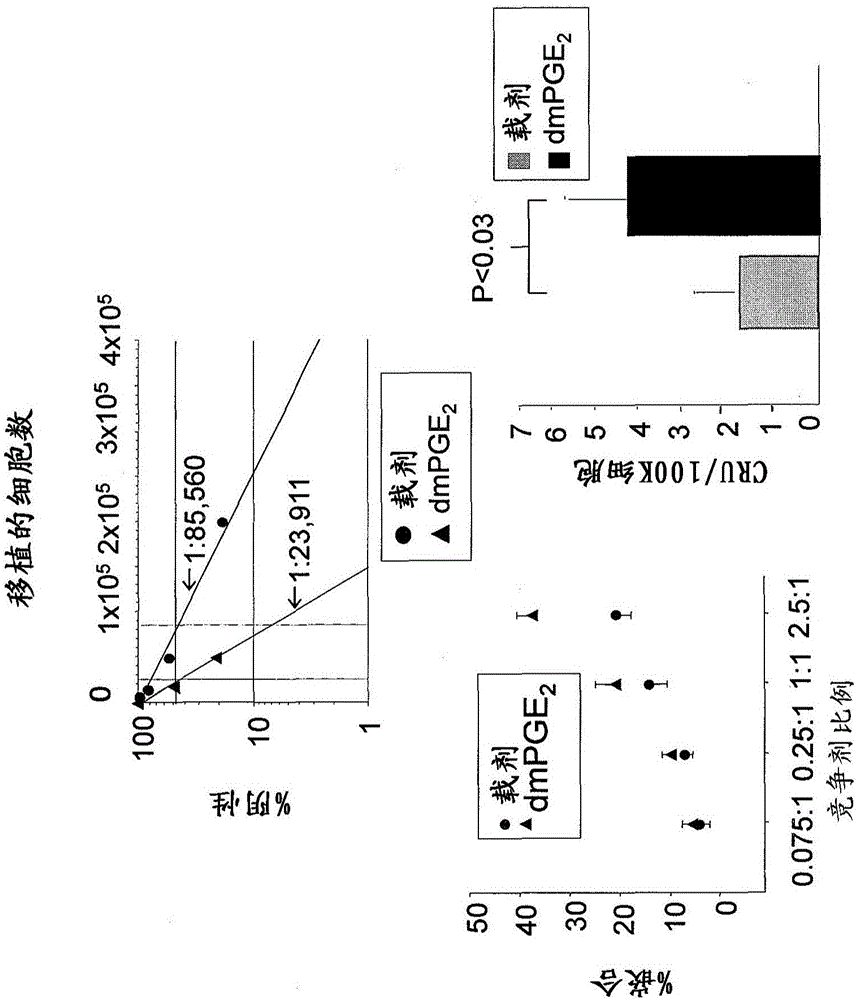
图1a.检测pge2增强造血干细胞植入之作用的实验的略图(上图);举例说明cd45.1和cd45.2细胞群的代表性流式细胞术结果图示(下图)。
图1b.阴性细胞百分比相对于植入细胞数的图(上图);嵌合百分比相对于竞争剂比例的散点图(左下图);不同条件下测定的cru/100k细胞的柱状图(右下图)。
图1c.概述在20周期间针对用和不用dmpge2处理的细胞绘制的恢复群体之细胞频率的表格。
图1d.多谱系重建的代表性facs图(髓细胞、b和t淋巴细胞,左上图)。计数/cd3的图(右上图)。中间行,在一级接受者中32周时(左图)和在二级接受者中12周时(右图)检测到的总wbc百分比的柱状图。在一级接受者中20周时和在二级接受者中12周时检测到的嵌合百分比图(下图)。
图2a.macs微珠除去lin-骨髓的代表性facs门控选择(facsgating),其显示lin-门控细胞(gatedcell)的c-kit+和sea-1+的门控选择(左图)。针对不同ep受体进行作图的计数(中图)和针对不同受体绘制的mrna相对于循环数的变化(右图)。
图2b.ssc相对于cd34的代表性facs图(左图);针对不同ep受体绘制的计数(中图)和针对不同受体绘制的mrna相对于循环数的变化(右图)。
图3a.实验略图(上图);针对不同处理进行作图的%cfse+(下图)。
图3b.举例说明实验方案的示意图;暴露于16,16-二甲基前列腺素e2(dmpge2)和多种对照后检测的%归巢效率(左下图)和改变倍数(右下图)。
图3c.实验略图(左图);针对不同处理的cd45.2相对于cd45.1的facs图(中图);针对不同处理绘制的%归巢效率图(右图)。
图4a.以灰色显示的cxcr4受体表达同工型对照的代表性流式细胞术图示(顶行);举例说明鼠和人hspc脉冲式暴露于pge2对cxcr4表达之结果的柱状图,针对不同条件进行作图的cxcr4变化(底行)。
图4b.针对不同处理的百分比归巢效率的柱状图。
图5a.作为dmpge2浓度的函数进行作图的膜联蛋白v+skl的百分比。
图5b.针对不同条件进行作图的存活素增加的倍数。
图5c.针对暴露于dmpge2的不同时间进行作图的,相对于对照活性归一化的变化百分比。
图6a.代表性流式细胞术图示,其显示门控skl细胞的dna含量(7aad染色),处于s+g2m期的skl百分比,以及经dmpge2处理的细胞周期中skl的增加倍数(左图);该图显示三个实验的合并数据(右图)。
图6b.实验略图以及显示在不同处理下测量的处于s+g2/m期的归巢skl细胞之增加倍数的柱状图。漫画图显示实验方案(左图);在有和没有dmpge2存在下测得的s+g2/m增加倍数的柱状图。
图7a.对不同处理后测定的cfu-gm/ml血作图的柱状图。
图7b.对不同处理后测定的cfu-gm/ml血作图的柱状图。
图8.总结举例说明pge2影响细胞周期中slamskl之数据的表格。
图9.经对照和dmpge2处理之细胞的迁移相对于sdf-i浓度的图。
图10.cd34+细胞迁移百分比相对于sdf-i浓度和/或amd3100的图。amd3100为(1,r-[1,4-亚苯基双(亚甲基)]双[1,4,8,11-四氮杂环十四烷]八氢溴化物二水合物),以商品名
图11.skl细胞归巢效率百分比相对于用dmpge2和/或amd3100以及多种对照处理的柱状图。
图12.facs图(左图);以及说明有和没有dmpge2时所测定的细胞周期中skl变化倍数的柱状图(右图)。
图13.实验方案的略图(左图);显示加入和不加入dmpge2所测定的s+g2/m期增加的柱状图。
图14.起始暴露于dmpge2(正方形)或对照(载剂(vehicle),菱形)后随时间测得的一系列移植物中嵌合百分比的图。
图15.使用载剂(浅灰色)吲哚美辛(深灰色)或黄岑苷元(灰色)所测定的cfu-gm/ml血的柱状图(左图);用g-csf(浅灰色);g-csf加吲哚美辛(灰色阴影)或g-csf加黄岑苷元(灰色)所收集数据的图。
图16.使用g-csf以及g-csf加吲哚美辛所测得的,相比于g-csf,cfu/ml血的增加倍数的柱状图。
图17.使用g-csf或g-csf加吲哚美辛处理后测得的,skl细胞(左侧)或slamskl细胞(右侧)之动员细胞的表型分析柱状图。
图18.使用载剂(vehicle)或吲哚美辛(左图)或者amd3100或amd3100加吲哚美辛(右图),对cfu-gm/ml血作图的柱状图。
图19.用载剂、吲哚美辛、amd3100;g-csf;amd3100加grobeta;amd3100加吲哚美辛或者g-csf加吲哚美辛处理后测得的,对cfu-gm/ml外周血作图的柱状图。
图20.载剂(透明)、g-csf(黑)、g-csf加美洛昔康(((8e)-8-[羟基-[(5-甲基-1,3-噻唑-2-基)氨基]甲叉]-9-甲基-10,10-二氧代-10λ6-硫杂-9-氮杂双环[4.4.0]十碳-1,3,5-三烯-7-酮)(浅灰)或g-csf加吲哚美辛(灰)处理后,cfu-gm/ml血的柱状图。
图21.用g-csf(浅灰)或g-csf加吲哚美辛(灰)(左图)测得的竞争性群体恢复单位(competitiiverepopulatingunit,cru)的柱状图;以及用载剂、g-csf对skl细胞测得的cxcr4mfi;不错开、错开一天或错开两天(右图)。
图22.用g-csf(菱形)或g-csf加吲哚美辛(正方形)测得的百分比嵌合相对于pbmc∶bm比的图(左图);用g-csf(浅灰)或g-csf加吲哚美辛(灰色)所测得的cru/2百万pbmc。
图23.pmn相对于g-csf(菱形)或g-csf加美达佳(美洛昔康)(正方形)动员的pbmc移植后天数的图。
图24.plt相对于g-csf(菱形)或g-csf加美达佳(美洛昔康)(正方形)动员的pbmc移植后天数的图。
图25.漫画图,其总结了设计用于测试单独g-csf或g-csf加美洛昔康处理狒狒之作用的实验。
图26.从用g-csf或g-csf加美洛昔康处理的4只不同的狒狒抽取的每ml血中cd34+细胞(左侧)或cfu-gm(右侧)的图。
图27.使用对cox-1或cox-2选择性不同的不同化合物所检测的cfu-gm/ml血的柱状图。
图28.g-csf或g-csf加不同量的阿司匹林或布洛芬处理细胞后,相比于g-csf,所检测的cfu-gm/ml血的变化倍数的柱状图。
图29.用g-csf或不同水平的美洛昔康处理后外周血中所测得的g-csf/ml血的柱状图。
图30.用g-csf或不同水平的美洛昔康处理后骨髓中所测得的g-csf/股骨的柱状图。
具体实施方式
为了促进对这项新技术原理的理解,接下来参照其优选实施方案,使用具体的语言对其进行描述。但是,需要理解的是,这并不是意在对这一新技术的范围进行限定,如本新技术相关技术领域的技术人员通常所进行地,考虑了对本新技术原理的改变、修改和进一步应用。
前列腺素e2(prostaglandine2,pge2)是大量存在的生理性类二十烷酸(eicosanoid),其是癌症、炎症和多种其它生理系统的已知中介物。已有多个研究小组研究了pge2在血细胞生成中的作用,但研究结果难以协调一致。例如,体内和体外研究证明,pge2可负调节髓细胞生成:pge2促进bfu-e和cfu-gemm集落形成,并增强cfu-s和cfu-gm的增殖。另一方面,pge2可刺激hspc并对血细胞生成具有两阶段作用(biphasiceffect):pge2短期离体处理骨髓细胞显示出刺激静息细胞群、可能从干细胞或更原始的祖细胞产生细胞周期中(cycling)人cfu-gm。另外,最近有显示离体暴露于16,16-二甲基pge2增加小鼠骨髓细胞的群体恢复(repopulating)能力以及斑马鱼中肾髓的恢复(north等,2007)。这些研究表明pge2参与调节血细胞生成,但没有将pge2与造血干细胞归巢(homing)相关联。换句话说,之前的研究倾向于认为pge2参与调节hspc分化,而对细胞归巢没有直接的作用。
如本文所证明的,pge2对长期的恢复群体的hspc具有直接的稳定的作用,并通过增强自我更新的hspc的存活、归巢和增殖来促进植入(engraftment)。
本文公开的一个方面是抑制环加氧酶活性,其增加造血干和祖细胞在外周血系统中循环的频率。在一个非限制性实施例中,施用环加氧酶抑制剂,例如在从接受动员剂给药前一天开始,通过经口或全身性途径向造血干细胞供体每天施用50μg吲哚美辛,增强了外周血中干细胞和祖细胞的动员。将环加氧酶抑制剂(例如吲哚美辛)与临床上批准的动员剂(例如g-csf)同时使用,产生协同作用从而动员祖细胞。
造血干细胞和祖细胞的动员也可通过向供体提供有效量的前列腺素e受体拮抗剂来实现。
本公开的一些方面证明离体暴露于pge2提高移植后的hspc频率,并为pge2处理的hspc提供了竞争性的优势。用pge2离体处理骨髓干细胞增强了小鼠中的总干细胞植入,导致干细胞存活增强、归巢效率提高并增加干细胞的自我更新。采用有限稀释竞争性移植模型证明dmpge2诱导的hspc频率提高,所述模型在同一动物中进行的直接头对头(head-to-head)分析中比较了对照和dmpge2处理的细胞的植入。例如,未处理的造血移植物或纯化的造血干细胞群(例如,小鼠中的skl细胞或人中的cd34+细胞)与每1ml培养基(例如imdm)中浓度为0.001-10μmpge2/1-10百万细胞的真正(authentic)pge2或其更稳定的类似物16,16-二甲基pge2(或任何其它的活性pge类似物)在冰上孵育1-6小时。孵育之后,用无菌盐水洗3遍,静脉内施用于接受者。基于用poisson统计学计算hspc频率和分析竞争性群体恢复单位(cru),该过程证明了pge2脉冲处理的hspc具有~4倍的竞争优势。采用1/4数量的pge2处理的细胞相对对照细胞的频率分析证明了等同的重建(reconstitution)。此外,使用对照或pge2处理的细胞在二级移植接受者中观察到了完全的造血重建,表明pge2对hspc的自我更新没有不良影响。事实上,观察到了ltrc活性增加的趋势,因为在系列移植之前没有对细胞或动物进行额外的处理,所以表明短期pge2暴露对一级移植中观察到的hspc的增强作用是长期的。经pge2处理细胞的植入增强在28周中保持稳定。移植后90天在二级植入动物中的分析显示出完全的多谱系重建和持续的高hspc频率,表明短期pge2处理对长期恢复群体的hspc的稳定作用。
增强的植入可源自hspc频率、归巢、存活和/或增殖的变化。north等提出,pge2不影响hspc归巢;但他们的研究并没有特别地评估hspc。出人意料地,在本文中pge2诱导的hspc频率的提高在超过20周的时间段内保持稳定,并在二级移植中维持。在竞争性移植模型中的直接比较显示,hspc短期暴露于pge2产生~4倍的竞争性优势。尽管在对照和pge2处理的细胞之间总移植细胞的归巢效率没有差异,但观察到经pge2处理的分选出的skl细胞的归巢效率提高,有力地表明pge2对hspc归巢具有直接的作用。
这些结果提示pge2对hspc或hspc长期群体恢复能力的更强的作用,而非如以前研究所提出的仅仅是短期作用。
为了说明而提供的并且不限于此的一种可能性是pge2对hspc功能的影响可能通过参与hspc归巢和自我更新的α-趋化因子受体cxcr4趋化因子受体的上调以及调节hspc存活和增殖的凋亡蛋白存活素(survivin)的抑制剂来介导。
流式细胞术和qrt-pcr显示,在sea-1+,c-kit+,lineage-(skl)小鼠骨髓细胞和cd34+人脐带血细胞(humancordbloodcell,ucb)上,所有4种pge2受体(ep1-ep4)都表达,没有显著的受体亚型表达的差异。在分析与hspc功能相关的几种功能性特征时,观察到在pge2暴露后skl细胞(26.8%)和cd34+ucb细胞(17.3%)上cxcr4表达显著增加,并且在暴露后6小时,cxcr4mrna显著上调。增加的cxcr4与经pge2处理的移植物的体内骨髓归巢活性的~2倍增加相一致,并且在未处理的骨髓(p<0.001,3次实验,n=6只小鼠/组/实验,独立检测)和在相同动物的头对头式竞争的纯化skl细胞中(p<0.001,3次实验,n=5只小鼠/组/实验,独立检测)都观察到,表明了pge2对hspc的直接作用。通过用选择性cxcr4拮抗剂amd3100进行处理,归巢效率的增加显著地降低。
pge2处理增加skl细胞的cxcr4mrna及表面表达。此外,cxcr4拮抗剂amd3100显著降低pge2对归巢的增强作用,表明,尽管不能排除对粘附分子表达和功能的额外作用,但增强的cxcr4表达和对骨髓sdf-i的趋化性在很大程度上导致了归巢增强。
本文公开的一个方面是用pge2处理接受者在体内增强了移植进接受者的干细胞的存活。在移植时和之后每天向接受者胃肠外施用pge2或其活性类似物来增强干细胞,可增加移植hspc的存活。例如,pge2或其活性类似物可在临接受造血移植物之前和之后每天以0.0001-10μm的量施用于患者。
pge2体外处理在处理后24小时内导致细胞周期中活跃的skl细胞比例的增加。此外,与仅用载剂(vehicle)脉冲处理的移植细胞相比,在brdu处理的接受者小鼠中经pge2处理细胞的移植显示多于~2倍的供体skl细胞处于细胞周期中的s+g2/m期。
存活素被认为是hspc进入细胞周期和前进所必需的,而存活素在条件敲除小鼠中的缺失表明其是hspc的维持所必须的。本文报道的研究发现在经pge2处理的skl细胞中,存活素mrna和蛋白水平升高,并伴有活性胱天蛋白酶-3(caspase-3)(一种介导凋亡的蛋白酶)的降低。存活分析表明,在体外pge2剂量依赖性地降低skl细胞的凋亡,与存活素蛋白表达的1.7倍增加和活性胱天蛋白酶-3的减少(减少23-59%;暴露后24-72小时)相一致。
由存活素介导的hspc存活的增强很可能对增强植入有贡献。脉冲暴露于pge2将细胞周期中hspc的比例增加了~2倍,同时hspc频率、cru和brdu+skl细胞归巢增加,并且在一级和二级移植中维持增加的hspc的频率。对这些结果的一个非限制性的解释是pge2的脉冲式暴露可引发单一轮次的hspc自我更新。例如,ep2和ep4受体活化与糖原合酶激酶-3(glycogensynthasekinase-3,gsk-3)的磷酸化以及β-联蛋白(β-catenin)信号转导增强有关(hull等,2004;regan,2003),其为wnt通路的下游,已显示涉及hspc存活和自我更新(fleming等,2008;khan和bendall,2006)。可能通过ep4但不排他的限于ep4的pge2信号转导可能直接增加β-联蛋白。已有人提出gsk-3和wnt通路之间存在协同的交互对话(cross-talk)(wang等,2004)。
存活素也促进hspc处于细胞周期中,其是通过p21waf1/cdkn1实现的(fukuda等,2004),其已知涉及hspc功能(cheng等,2000),并阻断胱天蛋白酶-3活性(li等,1998;tamm等,1998)。最近有报道称p21参与hspc的自我更新(janzen等,2008)。本文所报道研究中的一个发现是pge2上调存活素并降低胱天蛋白酶-3,表明存活素通路可能参与pge2对自我更新增加的作用。有趣的是,注意到存活素(peng等,2006)和cxcr4(staller等,2003;zagzag等,2005)的转录被转录因子低氧诱导因子-α(hypoxia-induciblefactor-1alpha,hif-iα)所上调,其可被pge2所稳定(liu等,2002;piccoli等,2007),有可能将某些一些pge2应答性通路与hspc的细胞存活、归巢和增殖/自我更新相关联。
这些研究表明,在pge2处理后观察到的hspc频率~4倍的增加来自于~2倍或更多的hspc归巢到接受者骨髓,以及增加~2倍的hspc经历自我更新。这些结果可以帮助定义pge2增强hspc功能的新作用机制,并且其表明出人意料的协助造血移植的治疗方法,尤其是针对细胞数有限导致植入潜力差的造血移植物。
本文公开的一个方面是增强干细胞基因治疗中病毒转导效率的方法。离体pge2处理干细胞增加这类细胞的自我更新分裂和存活,这是成功进行病毒载体介导的基因整合的重要因素。pge2促进的干细胞自我更新分裂/存活可合并到现有干细胞转导方案中,从而增加干细胞治疗中的总体基因转导效率。
本文报道的是使用pge2来增强hspc植入的一些方法,其为包括下列的多步骤过程:供体细胞的动员、hspc的维持和hspc在接受者体内的归巢。在一些条件下,这些方法导致hspc频率增加4倍,植入可能例如源自以下的累加作用:在pge2直接的影响下hspc归巢增加2倍和hspc细胞周期活性增加2倍。尽管确切的信号转导通路尚未确定,但是对该作用的一个非限制性解释是植入的增强是由于例如cxcr4和存活素的因子上调所导致的。
pge2提高hspc归巢和存活和/或增殖的能力可具有临床上的意义,尤其是在hspc数量有限(例如ucb和一些动员的pb产物)或者用于干细胞基因治疗中病毒转导的情况下。我们的有限稀释移植研究显示,与未经此处理的对照细胞相比,用1/4数目的经pge2处理的细胞能达到等同的植入结果。这些结果证明,在hspc数目有限的情况下使用pge2的功效。看上去全部四种ep受体亚型都在hspc上表达,并不清楚这些受体中那种(或它们的全部)参与植入功能。与这些结果相一致的是,增强的植入/恢复可通过体内施用pge2而实现,或者如果pge2体内施用,可进一步地有助于离体暴露于pge2的hspc的植入。
材料和方法
材料
小鼠:c57b1/6小鼠购自jacksonlaboratories(barharbor,me,usa)。b6.sjl-ptrcapep3b/boyj(boyj)和f1c57b1/6/boyj为内部培育的杂合体。所有的动物均采用微隔离笼饲养,并可连续摄食和饮用酸化水。移植研究用的小鼠在接受放射性辐照时并在移植后30天中接受饲喂多西环素(doxycycline)。印第安那大学医学院动物管理及使用委员会批准了全部的动物实验方案。
流式细胞术:除另有说明,所有的抗体均购自bdbiosciences。为检测和分选小鼠kl和skl细胞,使用与pe-cy7缀合的链霉亲和素(用于对生物素标记的macs谱系抗体(miltenyibiotech,auburn,ca)染色)、c-kit-apc、sea-1-pe或apc-cy7、cd45.1-pe和cd45.2-fitc。使用抗人cd34-apc检测ucbcd34+细胞。对于多谱系分析,使用apc-cy7-mac-1、pe-cy7-b-220和apc-cd3。用抗ep1、ep2、ep3和ep4的兔igg(caymanchemicals)和fitc-山羊-抗兔igg(southernbiotech,birmingham,al)第二染色来检测ep受体。使用链霉亲和素-pecy7、c-kit-apc、sca-1-apc-cy7和cxcr4-pe分析cxcr4的表达。用fitc-膜联蛋白-v检测凋亡。对于存活素和活性胱天蛋白酶-3的检测,用cytofix/cytoperm试剂盒(bd)透化并固定细胞,用抗活性胱天蛋白酶-3-fitc的流式细胞术试剂盒(bd)或存活素-pe(r&dsystems)染色。
对于细胞周期分析,用7aad或fitc-brdu流式细胞术试剂盒(bd)对细胞染色。所有的分析都在lsrii上进行,用facsaria或facsvantage分选仪(bd)进行分选。用cellquestpro和uiva软件(bd)进行数据获取和分析。
方法
有限稀释法竞争性和非竞争性移植
用无菌无热源pbs中1μmdmpge2(caymanchemical,annarbor,mi)/1×106细胞或0.01%乙醇在冰上处理wbm细胞(cd45.2)2小时。孵育后,将细胞洗两遍并与2×105同系cd45.1竞争剂骨髓细胞以0.075∶1、0.25∶1、1∶1和2.5∶1的比例混合并通过尾静脉注射(每个稀释度5只小鼠)移植进入经致死量辐照过的cd45.1小鼠(1100-cgy,分剂量)。每个月用流式细胞术测定pb中的cd45.1和cd45.2细胞。对于头对头(head-to-head)竞争性移植,用载剂或dmpge2处理来自cd45.1小鼠和cd45.2小鼠的wbm,将其与来自cd45.1/cd45.2小鼠的2×105个竞争剂骨髓细胞以0.075∶1、0.25∶1、1∶1和2.5∶1的比例相混合,并移植进入经致死量辐照的cd45.1/cd45.2小鼠。每月检测pb中cd45.1、cd45.2和cd45.1/cd45.2细胞。使用l-calc软件(stemcelltechnologies,vancouverbc,canada)通过poisson统计法对hspc频率进行定量,嵌合的分布<5%被认为是阴性接受者。按照(harrison,1980)的描述计算竞争性群体恢复单位(competitiverepopulatingunit,cru)。对于二级移植,在移植后第20周将来自之前1∶1比例移植的f1杂合小鼠的2×106个wbm细胞以非竞争性方式注射进入经致死量辐照的f1杂合小鼠,每月评价pb嵌合和三谱系重建。
hspc体内归巢至骨髓的分析:用cfse(molecularprobes,eugene,or)标记cd45.2wbm,清洗,用1μmdmpge2或载剂在冰上对其进行处理。处理后,清洗细胞,将2×107个细胞移植进经致死量辐照的cd45.2小鼠。16小时后,冲洗股骨和胫骨,用macs微珠(miltenyibiotech)去除小鼠骨髓的lin+细胞组分,用生物素(谱系)、c-kit(k)和sca-1(s)特异性的与荧光团缀合的抗体进行染色,测定cfse+wbm(非谱系去除)、kl和skl细胞的总数。对于同类系(congenic)归巢研究,在冰上用1μm的dmpge2、载剂或pbs处理lin-cd45.1细胞。孵育后,清洗细胞,将2×106个细胞移植进cd45.2小鼠。16小时后,收获接受者的骨髓,谱系去除,染色以及测定供体cd45.1skl细胞。对于使用分选的skl细胞的竞争性头对头归巢研究,对来自cd45.2和cd45.1小鼠的lin-细胞进行facs分选,用dmpge2或载剂将细胞处理2小时,清洗,将3×104个cd45.1(载剂或dmpge2处理的)加3×104个cd45.2(dmpge2或载剂处理的)skl细胞移植进经致死量辐照的f1杂合小鼠。为评价cxcr4在归巢研究中的作用,在冰上用载剂或1μm的dmpge2加10μm的amd3100(anormedinc.,vancouver,bc,canada)处理lin-cd45.2细胞,将2×106个处理过的细胞注射进经致死量辐照的cd45.1小鼠。移植后16小时分析归巢的skl细胞。
ep受体、cxcr4和存活素的表达:针对skl和每种ep受体对来自cd45.2小鼠的副本lin-细胞样品进行染色,通过facs测定kl和skl细胞上的表面受体表达。对于人ep受体,经过学术评审委员会的批准,ucb获自wishard医院,indianapolis,in。用ficoll-paquetmplus(amershambiosciences)分离单个核细胞(mononuclearcell),用macs微珠(miltenyibiotech)选择cd34+细胞(fukudaandpelus,2001)。对副本细胞样品进行cd34和每种ep受体的染色,通过facs检测表面表达。为了评价cxcr4、存活素和活性胱天蛋白酶-3,用1μmdmpge2或载剂将lin-细胞或cd34+ucb细胞在冰上处理2小时,清洗,然后在rpmi-1640+10%胎牛血清(fbs)中37℃下培养24小时。如上文所述对细胞进行skl(小鼠细胞)和cxcr4、存活素和/或活性胱天蛋白酶-3的染色,并用facs进行分析。
细胞周期分析:对于体外细胞周期分析,用1μmdmpge2或载剂将lin-细胞处理2小时,清洗,在含有rmscf(50ng/ml)(r&dsystems,minneapolis,mn)、rhflt-3和rhtpo(每种100ng/ml)(immunex,seattle,wa)的stemcellpro培养基(stemcelltechnologies)中培养。20小时后,对细胞进行skl染色,固定并透化,用7aad(bdbiosciences,sanjose,ca)染色。通过facs法检测dna的含量从而测定处于s+g2/m期的skl细胞比例。对于体内细胞周期分析,对cd45.2小鼠进行致死量的辐照,并移植5×106个来自cd45.1小鼠经1μmdmpge2或载剂处理2小时的lin-细胞。移植时,接受者小鼠在饮用水中接受1mg/mlbrdu(sigmaaldrich,st.louis,mo)以及1mg/小鼠的brdulp。16小时后,分离接受者的骨髓,谱系去除并对cd45.1、skl和brdu进行染色。采用facs法检测每只小鼠中brdu+的归巢(cd45.1+)skl细胞的比例。
凋亡测定:在冰上用0.1nm到1μm的dmpge2或载剂对照处理lin-细胞,清洗,在不含生长因子的rpmi-1640+2%fbs中37℃下孵育,诱导凋亡。24小时后,对细胞进行skl和膜联蛋白-v染色,用facs法检测膜联蛋白-v+的skl细胞的比例。
逆转录和qrt-pcr:使用绝对rna纯化试剂盒(stratagene,lajolla,ca)获取总rna。用随机引物(promega,madison,wi)和mmlv-逆转录酶(promega)在含有1mmdntp和rna酶抑制剂的50μl体积中逆转录恒定量的rna,如所述(fukuda和pelus,2001)。加入无dna酶与rna酶的水(ambion,austin,tx),获得等于10ngrna/μl的终浓度,将5μl用于qrt-pcr。为sybrgreenqrt-pcr设计引物,扩增子大小为75-150bp。在30μl总体积中进行qrt-pcr,其使用platinumsybrgreenqpcrsupermixudgwithrox(invitrogen,carlsbad,ca),在abi-7000(appliedbiosystems,carlsbad,ca)仪器中进行,活化步骤50℃2分钟,95℃变性2分钟,扩增45个循环,95℃-15秒、50℃-30秒、72℃-30秒,然后解离,以确保仅获得单一产物。
可用于实施本发明的一些方面的非甾体抗炎化合物包括但不仅限于例如下列的化合物:塞来昔布(celecoxib)(4-[5-(4-甲基苯基)-3-(三氟甲基)吡唑-1-基]苯磺酰胺),以商品名
作为至少一种pge2受体之拮抗剂的化合物包括但不仅限于可从caymanchemicalcompany(annarbor,mi,u.s.a.)或其它化学品供应公司获得的化合物。
统计学分析:所有的汇总的值都以平均值±sem的形式表示。适当地采用微软excel(microsoftcorp,seattle,wa)的成对或不成对、双尾t检验函数来确定统计学差异。本文中,尤其是在一些图中,术语‘dmpge2’和‘dmpge’可互换使用。
实施例
1.pge2增加长期群体恢复hspc的频率和植入
使用有限稀释竞争性移植模型,应用移植入cd45.1/cd45.2杂合小鼠的cd45.2和cd45.1同类系移植物,证明hspc短期暴露于pge2产生长期的对hspc和竞争性群体恢复单位(cru)频率的增强。接下来参照图1a,用载剂或dmpge2分别处理来自cd45.1或cd45.2小鼠的骨髓。cd45.1/cd45.2杂合体骨髓细胞用作竞争剂。将有限稀释物移植进经致死量辐照(1100-cgy,分剂量(splitdose))的cd45.1/cd45.2杂合小鼠,20周时分析pb中的嵌合情况。显示了检测每个细胞群的代表性流式细胞术图(底部的图)。
接下来参照图1b,通过poisson统计学确定的12周时经载剂(红)或dmpge2(蓝)脉冲处理的细胞的频率分析(上图);po=85,560(载剂)和po=23,911(经dmpge2处理)。显示12周时的pb中嵌合和cru分析(均值±sem)。数据代表2个汇总的实验,n=5只小鼠/组/实验,每个均独立检测。
接下来参照图1c,20周的时间段里在经载剂或dmpge2处理的骨髓的接受者中的hspc频率分析。变化倍数表明,与载剂相比,dmpge2脉冲处理的细胞的植入频率增加。
接下来参照图1d,代表性的多谱系重建(髓细胞、b和t淋巴细胞)的facs图。接下来参照图1d的中间图,一级移植(32周,左图)和接受来自20周时一级移植小鼠之移植物的一组4只小鼠的12周后分析(右图)。一级移植20周时(二级移植的时间)和二级移植后12周(下图),与载剂相比dmpge2处理的细胞显示出嵌合增加。20周一级移植的数据来自两个汇总的实验,n=5只小鼠/组/实验,每个均独立检测。12周二级移植的数据,n=4只小鼠/组,每个均独立检测。
仍然参照图1d,系列移植评估了移植的造血移植物中hspc的自我更新和扩增。为了研究离体暴露于dmpge2和载剂的长期群体恢复细胞(long-termrepopulatingcell,ltrc)的扩增,在移植后20周时从一级移植动物收获骨髓,并移植入二级接受者中。二级移植后12周的pb分析显示来自所有移植小鼠的细胞出现多谱系重建,标志着一级移植的ltrc的自我更新。在一级供体中观察到的由暴露于dmpge2引起的嵌合的增加也在未经任何额外处理的二级移植中见到。此外,在二级移植中也观察到以前经dmpge2处理过的hspc的竞争性增加的趋势,在二级移植中对髓细胞谱系的重建有轻微的偏好。
这一模型允许在同一动物中定量比较来自对照和dmpge2处理组的hspc植入和竞争性(图1a),以及宿主细胞的内源群体恢复。在移植后12周,对外周血(peripheralblood,pb)的分析显示,与载剂处理的细胞相比,经dmpge2处理的细胞的嵌合增加,hspc频率和竞争性群体恢复单位(cru)增加~4倍,其为公认的对长期群体恢复能力的量度(图1b)。在整个20周的移植后随访过程中,保持了~4倍的hspc频率增加,表明dmpge2脉冲暴露的作用是稳定的(图1c)。在移植后32周,观察到pb中b淋巴细胞和t淋巴细胞以及髓细胞谱系的重建,在未处理竞争剂细胞、经dmpge2或载剂处理的细胞之间没有可分辨的差异(图1d)。2.小鼠和人造血干和祖细胞(hematopoieticstemandprogenitor,hspc)表达pge2受体
据报道,pge2与4种特异性的高度保守的g蛋白偶联受体ep1-ep4相互作用(sugimoto和narumiya,2007;tsuboi等,2002)。ep受体的全部功能解释了pge2所引起的多种有时是相反的应答(breyer等,2001)。pge2受体亚型在hspc上的表达以前是未知的。现在参照图2a,富含造血祖细胞(hematopoieticprogenitorcell,hpc)的c-kit+lin-(kl)细胞和富含hspc的sea-1+c-kit+lin-(skl)细胞上ep受体的分析显示,所有4种ep受体(ep3+、ep2、ep1和ep4)都表达。此外,现在参照图2a(右图),qrt-pcr在facs法分选的kl和skl细胞中检测到全部4种ep受体的mrna。接下来参照图2a(中图),ep3解离曲线显示多个峰,与已知的ep3多剪接变体相一致(namba等,1993)。对于kl或skl细胞,观察到任何ep受体亚型之间的表面表达或mrna水平没有显著的定量差异。接下来参照图2b(右图),与小鼠细胞类似,全部四种受体亚型都在人cd34+ucb细胞表面表达,qrt-pcr分析检测到全部四种ep受体的mrna(图2b)。
3.短期pge2暴露增加hspc的归巢效率
脉冲式暴露于pge2后所观察到的hspc植入的增强可能是来自增加的hspc数和/或细胞周期状态有利于细胞的作用或者对宿主骨髓中hspc归巢或增殖的作用。不论其原因,骨髓小生境(niche)是hspc自我更新和分化所必需的,很可能只有那些归巢进入这些小生境的hspc才可提供长期群体恢复。接下来参照图3a,为评估hspc归巢,在冰上用dmpge2或载剂将cfse标记的全骨髓(wholebonemarrow,wbm)cd45.2细胞脉冲式处理2小时,清洗,静脉内(iv)注射进经致死量辐照的cd45.2宿主中。16小时后,对归巢到骨髓的总cfse+细胞以及kl和skl细胞群中归巢事件数进行定量。当评价总wbm细胞时,在dmpge2和载剂处理的细胞之间没有观察到归巢到骨髓的cfse+细胞百分比的差异;但是,与对照相比,显著更多的skl细胞归巢到骨髓。接下来参照图3b,在同类系模型中,与载剂处理的或未处置的细胞相比,对于dmpge2处理的细胞也观察到显著更高的skl细胞百分比。在未处理和载剂处理的细胞之间,未见到归巢效率的显著差异。
下面参照图3c,为了确定dmpge2对skl细胞归巢的增强作用是直接的还是间接的,将头对头式移植模型中富集的hspc归巢与其它细胞进行比较。通过facs分选从cd45.2和cd45.1小鼠中分离高度纯化的skl细胞,用dmpge2或载剂进行处理,将3×104个载剂处理的cd45.1细胞加3×104个dmpge2处理的cd45.2细胞移植进cd45.1/cd45.2小鼠。用同类系品系同时移植另外一组入同种品系,交换处理组以检测品系归巢的任何偏好。与使用wbm的研究相似,纯化skl细胞的dmpge2脉冲暴露将其归巢效率增加了2倍,有力地表明pge2对hspc的直接作用。尽管skl细胞不是均质的hspc细胞群,但是高度富含ltrc(okada等,1992;spangrude和scollay,1990)。
4.pge2增加hspc的cxcr4,而cxcr4拮抗剂amd3100阻断归巢的增强
下面参照图4a,基质细胞衍生因子-1α(stromal-cell-derivedfactor1alpha,sdf-1α)/cxcr4轴参与hspc的运输(trafficking)和归巢。本研究评价了经dmpge2处理的hspc归巢的提高是否是sdf-1α/cxcr4信号转导增加的结果。在kl和skl细胞上,lin-细胞脉冲暴露于dmpge2增加了cxcr4的表达(图4a);类似地,dmpge2脉冲暴露如预期地增加了cd34+ucb细胞上cxcr4的表达。qrt-pcr证明,与载剂相比,dmpge2处理的细胞中cxcr4mrna水平提高,在第6小时观察到最大提高(未显示数据)。
现在参照图4b,为了确定上调的cxcr4是否在pge2处理后所观察到的归巢增强中发挥作用,使用了选择性cxcr4拮抗剂amd3100,其体外抑制向sdf-i的迁移以及体内抑制hspc归巢。pge2脉冲暴露将skl细胞的归巢增加了~2倍,而经载剂或dmpge2脉冲处理的细胞与amd3100一起孵育降低了skl细胞的归巢,并消除了经dmpge2脉冲处理的细胞的归巢效率提高。
5.pge2减少hspc的凋亡,与存活素增加相一致
pge2的处理导致hspc频率和cru增加~4倍(图1),但归巢仅增强~2倍(图3),表明其它事件参与植入的增强。在正常和恶性血细胞生成中,凋亡是重要的调节过程,而pge2牵涉抗凋亡的信号转导。此外,ep受体的下游信号分子camp的活化抑制cd34+细胞的凋亡。一种与这些结果一致的假说是dmpge2处理影响hspc的存活和/或增殖,其导致了植入增强。为评价dmpge2对hspc存活的作用,用0.1nm-1μm的dmpge2或载剂脉冲处理lin-细胞,在减少血清的无生长因子的培养基中培养。在skl细胞中,脉冲暴露于dmpge2剂量依赖性地减小凋亡(图5a),1μm时达到~65%的抑制。
在正常和恶性造血细胞中,凋亡蛋白存活素的抑制剂是凋亡和增殖的重要调节子。现在参照图5b,这些结果证明,在hspc中pge2对存活素有影响。在dmpge2脉冲处理后第24小时,小鼠skl细胞和cd34+ucb细胞中,细胞内的存活素水平显著高于(分别为1.7和2.4倍)对照,qrt-pcr分析显示存活素mrna与对照相比升高。
现在参照图5c,相对于对照,降低的活性胱天蛋白酶-3与skl细胞暴露于dmpge2后24、48和72小时所见的存活素增加相一致。
6.pge2处理增加hspc增殖
存活素调节hspc进入细胞周期并在细胞周期中前进。另外,参与hspc增殖和自我更新的β-联蛋白位于ep受体通路的下游。pge2调节这些细胞周期调节蛋白的能力提示hspc自我更新和增殖的增加可能导致dmpge2脉冲处理的细胞的植入增强。为验证这一假说,分析了经dmpge2或载剂体外脉冲处理的skl细胞的细胞周期状态。现在参照图6a,脉冲暴露于dmpge2增加skl细胞中dna含量,标志着细胞周期中细胞增加(左图、右上象限)。在3个实验中,与对照相比,经dmpge2处理后,超过60%的skl细胞处于细胞周期的s+g2/m期(图6a右图)。对kl或lin-细胞的细胞周期比未见显著性作用(未显示数据);提示dmpge2选择性增加处于细胞周期中状态的早期hspc。
为验证体外观察到的dmpge2对hspc细胞周期增强的作用,用dmpge2脉冲处理骨髓细胞并注射进移植后用brdu处理的同类系小鼠,16小时后测定供体brdu+skl细胞的比例。现在参照图6b,对于移植前经dmpge2脉冲处理的细胞,观察到了s+g2/m期的归巢skl细胞比例增加~2倍,证实hspc短期暴露于dmpge2在体内刺激hspc进入细胞周期并在周期中前进。
7.双重cox1/cox2抑制剂吲哚美辛对内源性pge2生物合成的抑制动员hspc
因为pge2增加cxcr4受体表达并且sdf-1/cxcr4信号转导对于hspc的运输和在骨髓中的存留十分重要。因此,与这些结果相一致的一个假说是双重cox1/cox2抑制剂吲哚美辛对内源性pge2生物合成的抑制也会动员hspc。参照图7a和7b,显示了每天单独皮下(sc)150μg/kg吲哚美辛或150ug/kg黄岑苷元(图7a)或与g-csf联用(图7b)4天对cfu-gm动员的作用。现在参照图7a,每天一次共4天皮下施用150μg/kg吲哚美辛使得动员的祖细胞数增加4倍。现在参照图7b,吲哚美辛与g-csf共施用产生高度协同性的外周血干细胞动员增加。脂加氧酶抑制剂黄岑苷元(baicalein)对基线或g-csf诱导的cfu-gm动员没有作用,提示观察到的作用是对环加氧酶通路抑制特异性的。数据表示为动员的cfu-gm/ml血的均值±sem,n=3只小鼠,各自独立检测。
8.小鼠和人hspc脉冲暴露于pge2增加cxcr4表达。
为评价cxcr4,在冰上用1μmdmpge2或载剂对照处理谱系-的小鼠骨髓细胞或cd34+ucb细胞2小时,清洗,在rpmi-1640/10%热失活胎牛血清(hi-fbs)培养基中37℃下培养24小时,对skl(小鼠细胞)或cd34(人)和cxcr4进行染色并通过facs进行分析。现在参照图4a,dmpge2处理24小时后cxcr4在鼠kl和skl细胞和人类cd34+ucb细胞上的表达。数据以dmpge2或溶剂(n=3)处理所引起的cxcr4平均荧光强度(mfi)的变化的均值±sem的形式表示。qrt-pcr分析证明cxcr4的mrna增加2.65倍。
9.小鼠skl细胞脉冲暴露于pge2增加sdf-1α的迁移。
用dmpge2或载剂脉冲处理新分离的谱系-小鼠骨髓细胞2小时,清洗,重悬于10%hi-fcs中,并在37℃下培养16小时。孵育后,清洗细胞,重悬于rpmi/0.5%bsa,使其在侵袭小室(transwell)中向rmsdf-1α迁移4小时。通过流式细胞术检测总细胞迁移。现在参照图9,对于用dmpge2脉冲处理的细胞,总skl细胞迁移显著较高。数据是3次实验的迁移百分比的均值±sem。与载剂处理的细胞相比,dmpge2处理的细胞的
10.人cd34+细胞脉冲暴露于pge2增加向sdf-1α的迁移。
用dmpge2或载剂脉冲处理新分离的ucbcd34+细胞2小时,清洗,重悬于10%hi-fcs,在37℃下培养16小时。孵育后,清洗细胞,重悬于rpmi/0.5%bsa中,通过流式细胞术测定向rhsdf-1的迁移。为阻断cxcr4受体,在迁移测定之前,将副本细胞与5μg/ml的amd3100一起孵育30分钟。现在参照图10,数据为3次实验的迁移百分比的均值±sem。
11.阻断cxcr4受体阻断了pge2对skl细胞归巢的增强
为评价cxcr4在归巢中的作用,将谱系-cd45.2细胞与载剂或1μm的dmpge2加10μmamd3100共同孵育,将2×106个经处理的细胞注射进经致死量辐照过的cd45.1小鼠,移植后16小时,回收归巢的skl细胞并用facs分析。现在参照图11,在10μmamd3100不存在和存在时,经载剂和dmpge2处理的细胞归巢到骨髓的效率。在归巢测定之前,将细胞与amd3100一起孵育30分钟。
12.pge2在体外增加鼠skl细胞的细胞周期比率
用载剂或1μmdmpge2处理谱系-细胞2小时,清洗,在含rmscf、rhflt3和rhtpo的培养基中培养。20小时后,对细胞进行skl和hoechst-33342和派洛宁-y(pyronin-y)染色。通过facs检测处于细胞周期中的skl细胞的比例。现在参照图12,显示门控skl细胞之细胞周期分布的代表性流式细胞术图以及来自三次实验的经dmpge2处理的细胞与载剂对照相比细胞周期中细胞增加倍数的综合数据,平均值±sem,n=9只小鼠,各自独立检测。通过facs检测处于细胞周期中的skl细胞的比例。显示门控skl细胞之细胞周期分布的代表性流式细胞术图以及来自三次实验的经dmpge2处理的细胞与载剂对照相比细胞周期中细胞增加倍数的综合数据,平均值±sem,n=9只小鼠,各自独立检测。
13.在体外,pge2增加高度纯化的cd150+48-(slam)skl细胞的细胞周期比率。
现在参照图8,表中总结了使用lin-骨髓细胞收集的数据,所述细胞经1μmdmpge2或载剂处理2小时并在生长因子存在(50ng/mlrmscf,各100ng/ml的rhflt-3和rhtpo)的情况下培养20小时,针对slamskl、hoechst-33342和pyronin-y进行染色,通过facs对dna和rna的含量进行定量来测定处于go、gi、s和g2/m期的slamskl细胞的比例。数据是均值±sem,n=9只小鼠,各自独立检测。(b)gi+s+g2/m期的细胞百分比;n=9只小鼠的综合数据。(*)与载剂对照相比p<0.05。
14.脉冲式暴露于pge2在体内增加归巢skl细胞的增殖和细胞周期比率。
用dmpge2或载剂处理cd45.1谱系-骨髓细胞,并移植进经致死量辐照过的cd45.2小鼠。移植后立即在饮水中提供并通过腹膜内(ip)注射施用brdu。16小时后,分析骨髓,通过facs分析brdu+的cd45.1+skl细胞的比例。现在参照图13,用dmpge2或载剂处理cd45.1谱系-骨髓细胞,并移植进经致死量辐照的cd45.2小鼠。移植后立即在饮水中提供并通过ip注射施用brdu。16小时后,分析骨髓,以及通过facs分析brdu+的cd45.1+skl细胞的比例。较高比例的经dmpge2处理的skl归巢到骨髓。数据为均值±sem,n=5只小鼠,各自独立检测。
15.pge2脉冲式暴露后干细胞的长期群体恢复活性维持
对于头对头式的竞争性分析中,用载剂或dmpge2处理来自cd45.1和cd45.2小鼠的wbm,并以多种比例与2×105个来自cd45.1/cd45.2小鼠的竞争剂骨髓细胞混合并移植进经致死量辐照的cd45.1/cd45.2小鼠。每月检测外周血中cd45.1、cd45.2和cd45.1/cd45.2细胞的比例。对于二级、三级和四级移植,将来自之前以1∶1比例移植的cd45.1/cd45.2小鼠的2×106个wbm以非竞争性方式注射进经致死量辐照的cd45.1/cd45.2小鼠。每月检测外周血中cd45.1、cd45.2和cd45.1/cd45.2细胞的比例。现在参照图14,对于一级移植在第20周(二级移植的时间)以及在一个亚组中第32周(二级移植的12周分析的时间),对于二级移植在第12周和第24周,以及类似地对于三级和四级移植在各自在第12周,显示出与载剂相比经dmpge2处理的细胞的嵌合增加。20周一级移植的数据来自2个汇总的实验,n=5只小鼠/组/实验,各自独立检测。二级、三级和四级移植的数据,n=5只小鼠/组,各自独立检测。
16.针对吲哚美辛和g-csf的外周血干细胞(peripheralbloodstemcell,pbsc)的动员方案
每48小时共4天向小鼠皮下(sc)给予明胶中的150μg/kg吲哚美辛或150μg/kg黄岑苷元(脂加氧酶抑制剂),同时给以或不给以g-csf。按照以前的描述测定cfu-gm动员(pelus等,experimentalhematology33(2005)295-307)。现在参照图7a和7b,双重环加氧酶抑制剂吲哚美辛和g-csf的组合协同动员小鼠hspc。每天单独皮下(sc)施用150μg/kg吲哚美辛或150ug/kg黄岑苷元(脂加氧酶抑制剂)(图7a)或者与g-csf一起施用(图7b)4天对cfu-gm动员的作用。数据表示为均值±sem动员的cfu-gm/ml血,n=3只小鼠,各自独立检测。
现在参照图16,小鼠每天两次皮下注射g-csf(每只小鼠1μg)或g-csf+吲哚美辛(每只小鼠50μg)4天。按照(pelus等,experimentalhematology33(2005)295-307)的描述测定cfu-gm的动员。联用处理的小鼠显示cfu-gm/单位血的增加倍数大于仅用g-csf处理的动物。
通过facs分析用上述方案动员的来自小鼠外周血的低密度单个核细胞。对于skl和slam-skl细胞的检测,用sca-1-pe-cy7、c-kit-apc、cd150-pecy5、cd48-fitc、lineagecocktail-biotin染色,并用链霉亲和素-apc-cy7进行第二染色。现在参照图17,在bd-lsrii上进行分析。经g-csf或者g-csf和吲哚美辛组合处理的小鼠外周血中表型限定的hspc的流式细胞术分析。n=5只小鼠/组,各自独立检测。
17.吲哚美辛加amd3100联合动员hspc。
在4天里对小鼠通过每天两次皮下注射施用载剂或吲哚美辛(50μg/小鼠)。第五天对小鼠施用载剂或amd3100(5mg/kg)。1小时后,处死小鼠,按以前的描述检测cfu-gm动员(pelus等,experimentalhematology33(2005)295-307)。现在参照图18,载剂或吲哚美辛单独处理导致的cfu-gm动员(左图)。单次施用amd3100或吲哚美辛处理+amd3100导致的cfu-gm动员(右图)。数据表示为均值±sem,n=5只小鼠/组,各自独立检测。
18.吲哚美辛与多种动员方案联用的动员效率的比较
用载剂、吲哚美辛(50μg/只小鼠,每天两次皮下施用,4天)、amd3100(5mg/kg,第5天)、g-csf(1μg/只小鼠,每天两次皮下施用,4天)、amd3100+groβ(分别为5mg/kg和20mg/kg,第5天)、amd3100+吲哚美辛(吲哚美辛50μg/小鼠,每天两次皮下施用,4天;amd3100,5mg/kg,第5天)或g-csf+吲哚美辛(分别为1μg和50μg,每天两次皮下施用,4天)。按以前的描述检测cfu-gm的动员(pelus等,experimentalhcmatology33(2005)295-307)。现在参照图19,针对如上所述的多种处理方案绘制的cfu-gm/ml外周血。
用载剂、g-csf(1μg/小鼠,每天两次皮下施用,4天)、g-csf+吲哚美辛(50μg/小鼠,每天两次皮下施用,4天)或g-csf+美洛昔康(0.3mg/kg,每天两次皮下施用,4天)处理小鼠。按以前的描述检测cfu-gm的动员(pelus等,experimentalhematology33(2005)295-307)。现在参照图20,柱状图,其显示了吲哚美辛+g-csf以及类似作用的nsaid美洛昔康+g-csf诱导之动员的比较。数据表示为均值±sem,n=5只小鼠/组,各自独立检测。
19.nsaid错开给药使得hspc上cxcr4的表达恢复。
用g-csf(1μg/小鼠,每天两次皮下施用,4天)或g-csf+吲哚美辛(50μg/小鼠,每天两次皮下施用,4天)动员cd45.1小鼠,在第5天收集外周血单个核细胞(peripheralbloodmononuclearcell,pbmc)。pbmc以多种比例与cd45.2骨髓混合,移植入致死量辐照过(1100cgy,分剂量)的cd45.2小鼠。现在参照图21,显示移植后12周的竞争性群体恢复单位(左图)。数据表示为来自两次实验的平均值±sem,n=5只小鼠/组/实验,各自独立检测。因为与单独的g-csf相比,吲哚美辛与g-csf共同施用动员的pbmc的植入没有改善,所以猜想归巢的缺陷可能是cxcr4受体表达降低的结果,这可通过错开吲哚美辛和g-csf的处理而减轻。用g-csf(1μg/小鼠,每天两次皮下施用,4天)、g-csf+吲哚美辛不错开(50μg/小鼠,每天两次皮下施用,4天)、g-csf+吲哚美辛错开1天(吲哚美辛先开始并且施用4天,在吲哚美辛处理的第二天开始施用g-csf,施用4天,使得在收集pbmc之前有一天只施用g-csf而不施用吲哚美辛)或者g-csf+吲哚美辛错开2天(吲哚美辛先开始并且施用4天,在吲哚美辛处理的第三天开始施用g-csf,施用4天,使得在收集pbmc之前有两天只施用g-csf而不施用吲哚美辛)。现在参照图21(右图),显示skl细胞上cxcr4的表达。数据以均值±sem的形式表示,n=5只小鼠/组,各自独立检测。
20.与g-csf单独动员的pbmc相比,来自经g-csf加nsaid处理的小鼠的已动员pbmc显示显著增强的长期干细胞功能。
用g-csf或g-csf+吲哚美辛(错开1天)动员cd45.1小鼠,将pbmc与cd45.2竞争剂骨髓一起移植进经致死量辐照的cd45.2小鼠。现在参照图22,显示移植后12周多种供体:竞争剂比例下的嵌合(左图)和竞争性群体恢复单位(右图)。数据以均值±sem的形式表示,n=5只小鼠/组,各自独立检测。
21.与g-csf单独动员的pbmc相比,当移植入经致死量辐照的小鼠时,来自经g-csf加nsaid动员的小鼠的pbmc更快地恢复外周血中性粒细胞计数。
用g-csf或g-csf+美洛昔康(错开1天)动员小鼠,将2×106个pbmc移植进入经致死量辐照的接受者。用hemavet950fs(drewscientific)每两天对血中的中性粒细胞进行计数,直到完全恢复(与对照亚组相比)。用hemavet950fs(drewscientific)每两天对血中的血小板进行计数,直到完全恢复(与对照亚组相比)。现在参照图23,每两天对外周血(pb)中的中性粒细胞进行计数,直到完全恢复(与对照亚组相比)。这些数据以均值±sem的形式表示,n=10只小鼠/组,各自独立检测。现在参照图24,每两天对外周血(pb)中的血小板进行计数,直到完全恢复(与对照亚组相比)。数据以均值±sem的形式表示,n=10只小鼠每组,各自独立检测。
22.测定g-csf和美洛昔康在狒狒中对细胞动员的作用
现在参照图25,第一组狒狒按照如下的给药方案进行动员,10ug/kg体重的g-csf,2周清除(washout)期后,用10ug/kg的g-csf加0.1mg/kg的美洛昔康处理动物。第二组狒狒按照如下的给药方案进行动员,10ug/kg体重的g-csf加0.1mg/kg的美洛昔康,2周清除期后,用10ug/kg的g-csf。现在参照图26,通过facs分析检测pb中的cd34+细胞,按照以前的描述测定cfu-gm/ml血。g-csf和美洛昔康的共施用增加了cd34+细胞的动员(左图)以及所测定的cfu-gm/抽取的血单位。
23.小鼠中对pbsc动员的最佳增强需要抑制cox1和cox1酶。
用g-csf动员小鼠,将使用g-csf和多种nsaids组合(阿司匹林[cox-1和cox-2];利克飞龙[cox-2和5-lox];sc-560[cox-1];戊酰基水杨酸(valerylsalicylate)[cox-1];伐地考昔[cox-2];ns-398[cox-2])的动员方案的pb中cfu-gm进行比较。现在参照图27,总结了这些检测的结果,这些数据以均值±sem的形式表示,n=4只小鼠/组,各自独立检测。与被认为高度选择性地抑制这两种异构酶中仅一种的化合物相比,已知对cox1和2均有抑制作用的化合物更好地动员集落形成细胞。为检测两种常用的cox抑制剂的效果,用g-csf或者g-csf和阿司匹林或布洛芬的组合(口服(po),1天两次,4天)来动员小鼠,按照以前的描述测定cfu-gm。现在参照图28,对于仅用g-csf处理的对照组小鼠、用g-csf加10、20或40mg/kg阿司匹林处理的小鼠以及用g-csf加10、20或40mg/kg布洛芬治疗的小鼠,用柱状图形式表示g-csf+阿司匹林和g-csf+布洛芬动员cfu-gm至外周血的剂量应答性分析。数据以均值±sem的形式表示,n=4只小鼠/组,各自独立检测。
24.检测美洛昔康对小鼠cfu的剂量依赖性作用。
用g-csf和如下剂量的美洛昔康动员小鼠,0.0(对照)、0.02、0.2、0.5、1.5和3mg/kg体重的美洛昔康(每天两次皮下施用,四天),按照以前的描述检测cfu-gm。现在参照图29,在动物的外周血或动物骨髓样品中检测美洛昔康对hspc的剂量应答(图30),这些数据以均值±sem的形式表示,n=3只小鼠每组,各自独立检测。
尽管用图表和前述的描述说明并描述了这一新技术,但是其仅被认为是对特性的举例说明而非限制。需要理解的是,仅展示和描述了优选的实施方案,而要求对在此新技术精神范围内的所有改变和修改加以保护。同样,虽然使用特定的实施例、理论论证、说明和举例来例证这一新技术,这些例证以及所附的讨论不应以任何方式解释为对本技术的限制。本申请中参考的所有的专利、专利申请和文本的参考,科学论文、出版物等在此作为整体以参考的形式整合到本申请中。
本发明还涉及以下各项实施方案(它们对应于原申请的权利要求书):
1.增强造血干和祖细胞动员的方法,包括如下步骤:
鉴定造血干和/或祖细胞的来源;
提供降低pge2活性的化合物;和
将造血干和或祖细胞的来源与有效量的所述降低细胞和周围pge2活性的化合物相接触。
2.根据实施方案1的方法,其中所述降低pge2活性的化合物为非甾体抗炎化合物。
3.根据权利要求2的方法,其中所述非甾体抗炎化合物作用于环加氧酶-1和环加氧酶-2。
4.根据实施方案3的方法,其中所述非甾体抗炎化合物主要作用于环加氧酶-2。
5.根据实施方案2的方法,其中所述非甾体抗炎化合物选自:阿司匹林、塞来昔布、罗非昔布、艾托考昔、伐地考昔、布洛芬、萘普生、双氯芬酸、依托度酸、酮咯酸和利克飞龙。
6.根据实施方案2的方法,其中所述非甾体抗炎化合物为吲哚美辛。
7.根据实施方案2的方法,其中所述非甾体抗炎化合物为美洛昔康。
8.根据实施方案2的方法,其中所述非甾体抗炎化合物施用于患者一段时间,该时间段与至少一种额外的增强造血干和祖细胞动员的化合物的共处理重叠。
9.根据实施方案8的方法,其中所述增强造血干和/或祖细胞动员的额外的化合物选自:g-csf和普乐沙福。
10.根据实施方案2的方法,其中所述非甾体抗炎化合物施用于患者至少3天。
11.增强来自供体的造血干和/或祖细胞动员的方法,其包括如下步骤:
提供作为至少一种pge2受体之拮抗剂的化合物;和
从造血干和/或祖细胞的供体中收获造血干和祖细胞之前,向所述供体施用有效量的所述化合物。
12.根据实施方案11的方法,其中至少一种pge2受体的所述拮抗剂选自:n-[[4′-[[3-丁基-1,5-二氢-5-氧代-1-[2-(三氟甲基)苯基]-4h-1,2,4-三唑-4-基]甲基][1,1′-联苯基]-2-基]磺酰基]-3-甲基-2-噻吩甲酰胺和4-(4,9-二乙氧基-1,3-二氢-1-氧代-2h-苯并[f]异吲哚-2-基)-n-(苯磺酰基)-苯乙酰胺。
13.增强接受者中造血干和/或祖细胞植入的方法,包括如下步骤:
从经过至少一种降低来源中pge2活性的化合物处理的来源中收获包含造血干和/或祖细胞的细胞群;
将所述造血干和祖细胞的群与pge2活性离体相接触;和
将与具有pge2活性的所述化合物相接触的所述造血干和祖细胞体内移植入接受者。
14.根据实施方案13的方法,其中所述造血干和祖细胞的来源为从骨髓供体中抽取的骨髓样品。
15.根据实施方案13的方法,其中所述造血干和祖细胞的来源为从血液供体中抽取的血液样品。
16.根据实施方案13的方法,其中所述细胞来源为脐带或胎盘。
17.提高造血干和祖细胞植入率的方法,包括如下步骤:
提供具有pge2活性的化合物;和
将具有pge2活性的化合物与造血干和祖细胞的细胞群离体相接触。
18.根据实施方案17的方法,其中具有pge2活性的所述化合物选自pge2和dmpge2。
19.根据实施方案17的方法,其中具有pge2活性的所述化合物与所述细胞群接触至少1小时。
20.根据实施方案17的方法,其还包括如下步骤:
用基本上没有pge2活性的缓冲液将与具有pge2活性的所述化合物相接触的细胞群的至少一部分清洗至少一次。
21.根据实施方案16的方法,其还包括如下步骤:
将与具有pge2活性的所述化合物相接触的细胞群的至少一部分引入患者。
22.增强干细胞中病毒转导效率的方法,包括如下步骤:
提供含有至少一个待转导目的基因的病毒载体;
提供经有效量的具有pge2活性的分子离体处理的至少一个干细胞;和
用所述病毒载体转染经具有pge2活性的分子处理的干细胞。
23.根据实施方案22的方法,其中具有pge2活性的分子选自:e系列前列腺素和e系列前列腺素的二甲基衍生物。
24.根据实施方案22的方法,其中具有pge2活性的分子选自pge2和dmpge2。
25.增强造血干和/或祖细胞在移植接受者中存活的方法,包括如下步骤:
提供具有pge2活性的分子;
鉴定造血干或祖细胞移植的接受者;和
向所述移植接受者施用治疗有效量的具有pge2的分子。
26.根据实施方案25的方法,其中所述具有pge2活性的分子选自:e系列前列腺素和e系列前列腺素的二甲基衍生物。
27.根据实施方案25的方法,其中所述具有pge2活性的分子选自pge2和dmpge2。
参考文献
breyer,r.m,bagdassarian,c.k.,myers,s.a.,andbreyer,m.d.(2001).prostanoidreceptors:subtypesandsignaling.annu.rev.pharmacol.toxicol.41,661-690.
broxmeyer,h.e.(2006).cordbloodhematopoieticstemandprogenitorcells.inessentialsofstemcellbiology,elsevier,inc.),pp.133-137.
cheng,t.,rodrigues,n.,shen,h.,yang,y.,dombkowski,d.,sykes,m,andscadden,d.t.(2000).hematopoieticstemcellquiescencemaintainedbyp21cipl/wafl.science287,1804-1808.
fleming,h.e.,janzen,v.,lo,c.c,guo,j.,leahy,k.m.,kronenberg,h.m.,andscadden,d.t.(2008).wntsignalinginthenicheenforceshematopoieticstemcellquiescenceandisnecessarytopreserveself-renewalinvivo.cellstemcell2,274-283.
fruehauf,s.andseggewiss,r.(2003).it′smovíngday:factorsaffectingperipheralbloodstemcellmobilizationandstrategiesforimprovement[corrected].br.j.haematol.122,360-375.
fukuda,s.,mantel,c.r.,andpelus,l.m.(2004).survivinregulateshematopoieticprogenitorcellproliferationthroughp21waf1/cipl-dependentand-independentpathways.blood103,120-127.
fukuda,s.andpelus,l.m.(2001).regulationoftheinhibitor-of-apoptosisfamilymembersurvivininnormalcordbloodandbonemarrowcd34(+)cellsbyhematopoieticgrowthfactors:implicationofsurvivinexpressioninnormalhematopoiesis.blood98,2091-2100.
goldman,j.m.andhorowitz,m.m.(2002).theinternationalbonemarrowtransplantregistry.int.j.hematol.76suppl1,393-397.
hall,k.m.,horvath,t.l,abonour,r.,cornetta,k.,andsrour,e.f.(2006).decreasedhomingofretrovirallytransducedhumanbonemarrowcd34+cellsinthenod/scidmousemodel.exp.hematol.34,433-442.
janzen,v.,fleming,h.e.,riedt,t.,karlsson,g.,riese,mj.,lo,c.c,reynolds,g.,milne,c.d.,paige,c.j.,karlsson,s.,woo,m.,andscadden,d.t.(2008).
hematopoieticstemcellresponsivenesstoexogenoussignalsislimitedbycaspase-3.cellstemcell2,584-594.
khan,n.i.andbendall,l,j.(2006).roleofwntsignalinginnormalandmalignanthematopoiesis.histol.histopathol.21,761-774.
li3f.,ambrosini,g.,chu,e.y.,plescia,j.,tognin,s.,marchisio,p.c,andaltieri,d.c.(1998).controlofapoptosisandmitoticspindlecheckpointbysurvivin.nature396,580-584.
liu,x.h.,kirschenbaum,a.,lu,m.,yao,s.,dosoretz,a.,hollandj.f.,andlevine,a.c.(2002).prostaglandine2induceshypoxia-induciblefactor-1alphastabilizationandnuclearlocalizationinahumanprostatecancercellline.jbiolchem277,50081-50086.
muller-sieburg,c.e.andsieburg,h.b.(2006).clonaldiversityofthestemcellcompartment.curropinhematol13,243-248.
namba,t.,sugimoto,y.,negishi,m.,irie,a-,ushikubi,f.,kakizuka,a-,ito,s.,ichikawa,a.,andnarumiya,s.(1993).alternativesplicingofc-terminaltailofprostaglandinereceptorsubtypeep3determinesg-proteinspecificity.nature365,166-170.
north,t.e.,goessling,w.,walkley,c.r.,lengerke,c,kopani.k.r.,lord,a-m.,weber,g.j.,bowman,t.v.,jangj.h.,grosser,t.,fitzgerald,g.a.,daley,g.q.,orkin,s.h.,andzon,l,i.(2007).prostaglandine2regulatesvertebratehaematopoieticstemcellhomeostasis.nature447,1007-1011.
okada,s.,nakauchi,h.,nagayoshi,k.,nishikawa,s.,miura,y.,andsuda.t.(1992).invivoandinvitrostemcellfunctionofc-kit-andsea-1-positivemurinehematopoieticcells.blood80,3044-3050.
peng,x.h.,karna,p.,cao,z.,jiang,b.h.,zhou,m.,andyang,l.(2006).cross-talkbetweenepidermalgrowthfactorreceptorandhypoxia-induciblefactor-1alphasignalpathwaysincreasesresistancetoapoptosisbyup-regulatingsurvivingeneexpression.jbiolchem281,25903-25914.
piccoli,c,d′aprile,a-,ripoli,m.,scrima,r.,boffoli,d.,tabilio,a.,andcapitanio,n.(2007).thehypoxia-induciblefactorisstabilizedincirculatinghematopoieticstemcellsundernormoxicconditions.febslett581,3111-3119.
porecha,n.k.,english,k.,hangoc,g.,broxmeyer,h.e.,andchristopherson,k.w.(2006).enhancedfunctionalresponsetocxcl12/sdf-1throughretroviraloverexpressionofcxcr4onm07ecells:implicationsforhematopoieticstemcelltransplantation.stemcellsdev.75,325-333.
pulsipherma,chitphakdithaip,loganbr,leitmansf,anderlinip,kleinjphorowitzmm,millerjp,kingrj,conferdl,donor,recipient,andtransplantcharacteristicsasriskfactorsafterunrelateddonorpbsctransplantation:beneficialeffectsofhighercd34+celldose.blood.2009sep24;114(13):2606-16.epub2009jui16.
regan,j.w.(2003).ep2andep4prostanoidreceptorsignaling.lifesci.74,143-153.
spangrude,gj.andscollay,r.(1990).asimplifiedmethodforenrichmentofmousehematopoieticstemcells.exp.hematol.75,920-926.
staller,p.,sulitkova,j.,lisztwan,j.,moch,h.,oakeley,e.j.,andkrek,w.(2003).chemokinereceptorcxcr4downregulatedbyvonhippel-lindautumoursuppressorpvhl.nature425,307-311.
sugimoto,y.andnarumiya,s.(2007).prostaglandinereceptors.j.biol.chem.282,11613-11617.
tamm,l,wang,y.,sausville,e.,scudiero,d.a.,vigna,n.,oltersdorf,t.,andreed,j.c.(1998).iap-familyproteinsurvivininhibitscaspaseactivityandapoptosisinducedbyfas(cd95),bax,caspases,andanticancerdrugs.cancerres.58,5315-5320.
tsuboi,k.,sugimoto,y.,andichikawa,a.(2002).prostanoidreceptorsubtypes.prostaglandinsotherlipidmediat.68-69,535-556.
zagzag,d.,krishnamachary,b.,yee,h.,okuyama,h.,chiriboga,l.,ali,m.a.,melamedj.,andsemenza,g.l.(2005).stromalcell-derivedfactor-lalphaandcxcr4expressioninhemangioblastomaandclearcell-renalcellcarcinoma:vonhippel-lindauloss-of-functioninducesexpressionofaligandanditsreceptor.cancerres65,6178-6188.
- 还没有人留言评论。精彩留言会获得点赞!