一种抗除草剂抗虫融合基因、编码蛋白及应用的制作方法
本发明涉及一种抗除草剂抗虫融合基因、编码蛋白及应用。(二)
背景技术:
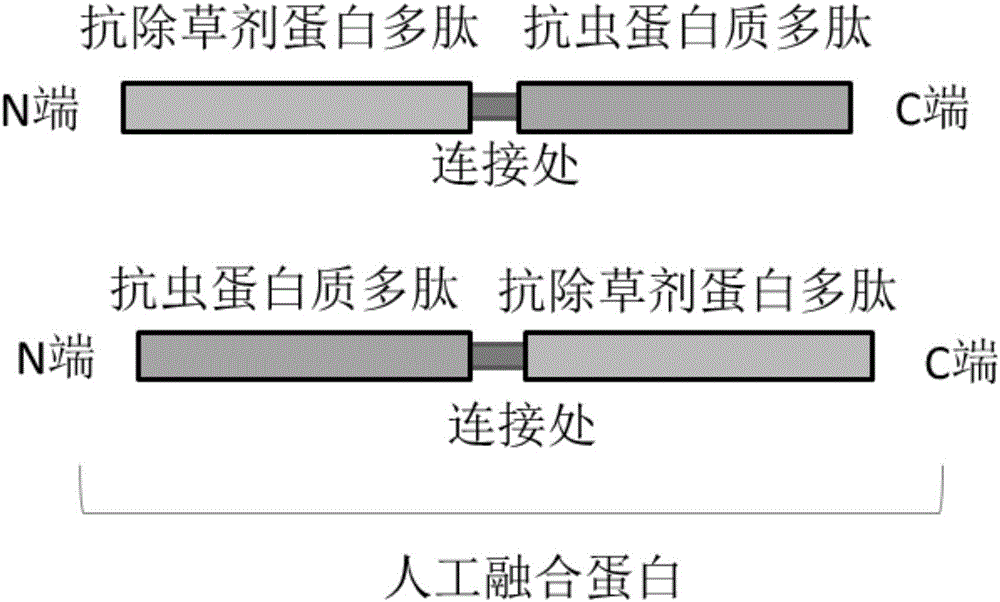
:杂草和病虫害是农业生产中影响产量的最重要的生物因素。如何降低杂草和病虫害的防治成本、提高防治效率、减少防治过程中给环境造成污染是农业科学研究最关键的问题。杂草时伴随着人类的生产活动而产生的,它们的存在是长期适应气候、土壤、作物、耕作制度及社会因素与栽培作物竞争的结果。人类从开始从事农业生产就在防治杂草,但是当时的杂草防治仅仅是一种极为粗放的初级劳动。随着科学的发展,特别是近代生命科学的发展进步,人类可以通过基因工程的方法赋予农作物耐受各种除草剂的特性,通过喷施除草剂就可以很好的防治杂草,把人类从初级劳动中解放出来,极大的提高效率。目前,国际上使用最广泛以及国内研究最多的是抗草甘膦转基因作物,其次是抗草铵膦转基因作物,最近陶氏益农公司研发了抗2,4-D作物,孟山都公司研发了抗麦草畏作物。能赋予农作物草甘膦抗性的基因很多,包括突变的对磷酸烯醇式丙酮酸具有高亲和性,而对草甘膦不敏感的EPSPS基因,例如CP4(Padgette,S.R,Kolacz,K.H,Delannay,X,etal.1995,CropScience,35(5):1451-1461)、aroA(ComaiL,SenLC,StalkerDM.1983,Science,221(4608):370-371)、G7(中国专利:200910098129.X)、G10(中国专利:201110009329.0);草甘膦解毒基因,包括可以把草甘膦转化为氨甲基磷酸的草甘膦氧化还原酶基因(glyphosateoxidoreductasegene,GOX)(Kishore,G.M.,&Barry,G.F.1995,Glyphosatetolerantplants)和可以把草甘膦转化为N-乙酰草甘膦的草甘膦乙酰转移酶编码基因(glyphosateN-Acetylation,GAT)(CastleLA;SiehlDL;GortonR;PattenPA;ChenYH;BertainS;ChoHJ;DuckN;WongJ;LiuD;LassnerMW.2004,Science,304(5674),1151-4;Castle,L.A.,Hong,C.Y.,Duck,N.B.,Giver,L.J.,Christina,I.,&Jeremy,M.,etal.2004,WO2002036782A3;2002,WO2002036782A2;GreenJM,HazelCB,RaymondFD,etal.2008,PestManagementScience,64(4):332-9.)。抗草铵膦的基因包括bar(ThompsonCJ;MovvaNR;TizardR;CrameriR;DaviesJE;LauwereysM;BottermanJ.1987,EmboJournal,6(9),2519-23;White,J.,Chang,S.Y.P.,Bibb,M.J.,&Bibb,M.J.1990,NucleicAcidsResearch,18(4),1062-1062.),pat(Wohlleben,W.,Arnold,W.,Broer,I.,Hillemann,D.,Strauch,E.,&Pühler,A.1988,gene70:25-37.Gene,70(1),25-37)。抗麦草畏的基因DMO(J.Biol.Chem.2005;美国专利:7022896)。抗2,4-D基因TfdA(LyonB.R.,LlewellynD.J.,HuppatzJ.L.,DennisE.S.,andPeacockW.J.,1989,PlantMol.Biol.,13(5):533-540)害虫给全球农业生产带来巨大的损失,目前害虫防治主要依靠化学农药,但是农药的使用加重了生产成本,并且农药残留对人体健康带来严重危害。因此,利用基因工程方法防治害虫具有重大经济、环境和社会价值。获得转基因抗虫作物的关键技术是克隆优良杀虫蛋白质。杀虫蛋白质有很多,使用得最为广泛的是苏云金芽孢杆菌(Bacillusthuringiensis,简称Bt)在产胞期间可以分泌一种伴胞晶体蛋白(ICP),如Cry1Ab,Cry1C等,这些蛋白对鳞翅目鞘翅目等昆虫(比如小菜蛾)有很强的杀伤作用(Schnepf,E.,Crickmore,N.,Van,R.J.,Lereclus,D.,Baum,J.,&Feitelson,J.,etal.1998,62(3),775-806.),目前已被深入研究和广泛使用,玉米、大豆、土豆、棉花等转Bt基因作物已得到大规模商业化种植。苏云金芽胞杆菌在营养生长期间,还会分泌一种与ICPs无氨基酸序列同源性、杀虫机理完全不同的蛋白,即营养期杀虫蛋白(vegetativeinsecticidalproteins,VIPs),如Vip3A、Vip3B、Vip3C、Vip3D、Vip3H,对一些对ICPs不敏感的害虫也有杀虫活性,并且不会发生交叉抗性(Estruch,J.J.,Warren,G.W.,Mullins,M.A.,Nye,G.J.,Craig,J.A.,&Koziel,M.G.1996,ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,93(11),5389-94.)。第一代转基因作物大多只具有抗虫或者抗除草剂的单一性状,而第二代转基因作物正在向同时抗虫抗除草剂等复合性状发展。获得具有复合性状的转基因作物有多种方法,比如通过对具有单一性状的转基因作物的杂交;通过将多个基因表达框构建在同一个表达框里面;通过共转化,将多个含有单基因的农杆菌混在一起,进行混合转化,筛选出多个基因同时整合了两个或多个质粒T-DNA的转基因植株。但是上述方法在实际操作和应用中都还存在一些问题。因此,获得更为简单高效的获得具有复合性状的转基因作物的方法是当下需要切实解决的问题。(三)技术实现要素:本发明目的是提供一种基因,这种基因具有抗除草剂和抗虫复合性状,可以用来生产抗除草剂抗虫植物。本发明采用的技术方案是:本发明提供一种抗除草剂抗虫融合基因,所述的基因包括编码抗除草剂蛋白的核苷酸序列和编码抗虫蛋白核苷酸序列;且上述2个核苷酸序列位于同一个开放阅读框内;抗除草剂蛋白和抗虫蛋白可以为全长或者截断后的活性多肽片段,具体所述融合基因由抗除草剂蛋白编码基因和抗虫蛋白编码基因构成;所述抗除草剂蛋白编码基因为下列之一:抗草甘膦基因CP4(Padgette,S.R,Kolacz,K.H,Delannay,X,etal.1995,CropScience,35(5):1451-1461)、抗草甘膦基因aroA(ComaiL,SenLC,StalkerDM.1983,Science,221(4608):370-371)、抗草甘膦基因G7(中国专利:200910098129.X)、抗草甘膦基因G10(中国专利:201110009329.0)、抗草甘膦基因GOX(Kishore,G.M.,&Barry,G.F.1995,Glyphosatetolerantplants)、抗草甘膦基因GAT(CastleLA;SiehlDL;GortonR;PattenPA;ChenYH;BertainS;ChoHJ;DuckN;WongJ;LiuD;LassnerMW.2004,Science,304(5674),1151-4;Castle,L.A.,Hong,C.Y.,Duck,N.B.,Giver,L.J.,Christina,I.,&Jeremy,M.,etal.2004,WO2002036782A3;2002,WO2002036782A2;GreenJM,HazelCB,RaymondFD,etal.2008,PestManagementScience,64(4):332-9.)、抗草铵膦基因bar(ThompsonCJ;MovvaNR;TizardR;CrameriR;DaviesJE;LauwereysM;BottermanJ.1987,EmboJournal,6(9),2519-23;White,J.,Chang,S.Y.P.,Bibb,M.J.,&Bibb,M.J.1990,NucleicAcidsResearch,18(4),1062-1062.)、抗草铵膦基因pat(Wohlleben,W.,Arnold,W.,Broer,I.,Hillemann,D.,Strauch,E.,&Pühler,A.1988,gene70:25-37.Gene,70(1),25-37)、抗麦草畏基因DMO(J.Biol.Chem.2005;美国专利:7022896)或抗2,4-D基因TfdA(LyonB.R.,LlewellynD.J.,HuppatzJ.L.,DennisE.S.,andPeacockW.J.,1989,PlantMol.Biol.,13(5):533-540);所述抗虫蛋白编码基因为下列之一:Cry1Ab、Cry1Ac、Cry2、Cry3、Vip3A、Vip3B、Vip3C、Vip3D或Vip3H。进一步,优选所述融合基因由抗除草剂蛋白编码基因Bar和抗虫蛋白编码基因Vip3A构成,所述融合基因的核苷酸序列为SEQIDNO.1所示。进一步,优选所述融合基因由抗除草剂蛋白编码基因GAT和抗虫蛋白编码基因Cry1Ab构成,所述融合基因的核苷酸序列为SEQIDNO.2所示。本发明还提供一种所述抗除草剂抗虫融合基因编码蛋白,当抗除草剂基因为bar,所述的抗虫基因为Vip3A时,所述编码蛋白的氨基酸序列为SEQIDNO.3所示,当抗除草剂基因为GAT,所述的抗虫基因为Cry1Ab时,所述编码蛋白的氨基酸序列为SEQIDNO.4所示。此外,本发明还提供一种所述抗除草剂抗虫融合基因编码蛋白在制备抗虫抗除草剂转基因作物中的应用,所述作物包括单子叶作物和双子叶作物,更优选所述作物为玉米、水稻、大豆、小麦和油菜中的一种。本发明设计的用抗除草剂蛋白和抗虫融合成的人工蛋白质分子,与现有的抗除草剂蛋白或者抗虫蛋白相比具有如下优点:同时具备抗除草剂和抗虫的特性;在生产转基因作物时,植株中抗除草剂蛋白质分子和抗虫蛋白质分子一样多。本发明提供的抗除草剂抗虫融合蛋白可以应用于单子叶植物和双子叶植物的抗虫抗除草剂方面,主要应用于抗除草剂抗虫玉米、水稻、大豆、小麦和油菜。(四)附图说明图1:抗除草剂抗虫融合蛋白的结构示意图。融合蛋白中的抗虫蛋白质多肽和抗除草剂蛋白质多肽可以分别在N端和C端或者分别在C端和N端。图2:融合蛋白表达载体T-DNA的结构示意图。pUBI为玉米泛素启动子,融合蛋白为抗虫抗除草剂融合蛋白基因,p35S为花椰菜花叶病毒35S启动子,G10EPSPS为抗草甘膦基因。(五)具体实施方式下面结合具体实施例对本发明进行进一步描述,但本发明的保护范围并不仅限于此:本发明以下实施例所使用的分子生物学和生物化学方法均为已知的技术。在Ausubel编写的JohnWileyandSons公司出版的CurrentProtocolsinMolecularBiology和J.Sambrook等编写ColdSpringHarborLaboratoryPress(2001)出版的MolecularCloning:ALabortoryManual,3rdED.等文献均有详细的说明。实施例1抗除草剂抗虫融合蛋白表达载体的构建抗草铵膦基因bar和抗虫基因Vip3A融合构成的融合基因被命名为bar-Vip3A基因,编码的蛋白质多肽序列如SEQIDNO:3所示。抗草甘膦基因Gat和抗虫基因Cry1Ab融合构成的融合基因被命名为Gat-Cry1Ab基因,编码的蛋白质多肽序列如SEQIDNO:4所示。bar-Vip3A(SEQIDNO:1)基因和Gat-Cry1Ab(SEQIDNO:2)基因都是通过人工合成获得,且都在5’端设置了BamHI酶切位点,都在3’端添加了人工合成的终止子,且设置了KpnI酶切位点。pUBI为玉米泛素蛋白启动子,通过PCR获得。设计PCR引物pUBI-F(5’GAAGCTTGCATGCCTACAGTGCAGCGTGACCC)和pUBI-R(5’GGGTGGATCCTCTAGAGTCGACCTGCAGAAGTAAC),以商业玉米品种郑单958的基因组DNA为模板,通过PCR扩增获得pUBI。PCR反应条件为:95℃3分钟;95℃15秒,68℃15秒,72℃2分钟,重复32个循环;然后72℃10分钟。将获得的大约2.0Kb的PCR产物克隆到T-载体pMD19中。然后,用HindIII和BamHI双酶切得到pUBI,并且DNA序列测定表明核苷酸序列正确(SEQIDNO:5)。G10EPSPS为抗草甘膦基因(中国专利:201110009329.0)。通过人工合成G10EPSPS基因(SEQIDNO:6)。合成的基因5’端连接有玉米乙酰乳酸合成酶AHAS叶绿体转运信号肽且设置有XhoI酶切位点,3’端连接有终止子且设置有XhoI酶切位点。农杆菌T-DNA载体的构建:双元载体pCambia1300-p35S-G10由载体pCambia1300修改而来,简单的说就是把pCambia1300载体中的抗潮霉素基因置换为抗草甘膦基因G10EPSPS。具体把pCambia1300载体经过XhoI酶切以后,去磷酸化处理,然后与经过XhoI酶切后的获得的人工合成的含有叶绿体转运信号肽和终止子的G10EPSPS基因片段进行两端连接,转化,获得的载体即为pCambia1300-p35S-G10。为了获得bar-Vip3A基因表达载体,用HindIII和KpnI对之前构建好的pCambia1300-p35S-G10进行双酶切,回收获得载体;用HindIII和BamHI酶切含有pUBI启动子的质粒,获得pUBI片段;用BamHI和KpnI对含有人工合成的bar-Vip3A基因及其终止子的质粒,回收得到bar-Vip3A片段。然后,把上述酶切后的载体和两个片段进行三段连接,获得终载体。获得的T-DNA结构为:“启动子-融合基因-终止子-启动子-G10EPSPS-终止子”。这个载体命名为:pCambia1300-pUBI-bar-Vip3A–p35S-1174(图2)。为了获得Gat-Cry1Ab基因表达载体,用HindIII和KpnI对之前构建好的pCambia1300-p35S-G10进行双酶切,回收获得载体;用HindIII和BamHI酶切含有pUBI启动子的质粒,获得pUBI片段;用BamHI和KpnI对含有人工合成的Gat-Cry1Ab基因及其终止子的质粒,回收得到Gat-Cry1Ab片段。然后,把上述酶切后的载体和两个片段进行三段连接,获得终载体。获得的T-DNA结构为:“启动子-融合基因-终止子-启动子-G10EPSPS-终止子”。这个载体命名为:pCambia1300-pUBI-Gat-Cry1Ab–p35S-1174(图2)。最后,通过电转的方法把上述2个T-DNA质粒转入农杆菌LBA4404中,通过含有15μg/ml四环素和50μg/mL的卡那霉素的YEP固体培养基筛选出阳性克隆,并保菌,用于接下来的植物转化。实施例2、水稻的转化转基因水稻的获得方法是采用现有技术(卢雄斌,龚祖埙(1998)生命科学10:125-131;刘凡等(2003)分子植物育种1:108-115)。选取成熟饱满的“秀水-134”种子去壳,诱导产生愈伤组织作为转化材料。分别取实施例1中构建的载体pCambia1300-pUBI-bar-Vip3A–p35S-1174和pCambia1300-pUBI-Gat-Cry1Ab–p35S-1174的农杆菌划板。挑单菌落接种,准备转化用农杆菌。将待转化的愈伤组织放入OD为0.6左右的农杆菌菌液中(农杆菌菌液的制备:将农杆菌接种至培养基,28℃培养至OD为0.6左右;培养基组成:3g/LK2HPO4、1g/LNaH2PO4、1g/LNH4Cl、0.3g/LMgSO4·7H2O、0.15g/LKCl、0.01g/LCaCl2、0.0025g/LFeSO4·7H2O、5g/L蔗糖、20mg/L乙酰丁香酮,溶剂为水,pH=5.8),让农杆菌结合到愈伤组织表面,然后把愈伤组织转移到共培养培养基(MS+2mg/L2,4-D+30g/L葡萄糖+30g/L蔗糖+3g/L琼脂(sigma7921)+20mg/L乙酰丁香酮)中,28℃共培养2-3天。用无菌水冲洗转化后的愈伤,转移到筛选培养基(MS+2mg/L2,4-D+30g/L蔗糖+3g/L琼脂(sigma7921)+20mg/L乙酰丁香酮+2mM草甘膦(Sigma))上,28℃筛选培养两个月(中间继代一次)。把筛选后,生长活力良好的愈伤转移到预分化培养基(MS+0.1g/L肌醇+5mg/LABA+1mg/LNAA+5mg/L6-BA+20g/L山梨醇+30g/L蔗糖+2.5g/Lgelrite)上,28℃培养20天左右,然后将预分化好的愈伤组织移到分化培养基上,每天14小时光照分化发芽。2-3周后,把抗性再生植株转移到生根培养基(1/2MS+0.2mg/LNAA+20g/L蔗糖+2.5g/Lgelrite)上壮苗生根,最后将再生植株洗去琼脂移植于温室,选择产量高、种子大或者生物量高等能够提高水稻产量的转基因株系,培育新品种。分别获得含上述转化载体和只含有筛选标记基因EPSPS的空载体的转基因水稻植株。实施例3、融合蛋白转基因水稻可以抗除草剂将实施例2方法制备的转基因水稻植株的T0代植株移栽到温室中,对转基因水稻植株和非转基因受体对照植株“秀水-134”的抗除草剂性能进行比较分析。我们获得的86个转pCambia1300-pUBI-bar-Vip3A–p35S-1174载体的转基因株系(命名为BV)进行不同浓度草铵膦抗性测定,抗性效果如表1所示:表1*:1x2x3x抗性转化事件个数655230*注:表一为对播种后15天的水稻植株喷施不同浓度草铵膦进行抗性检测。喷施草铵膦10天后株高、叶片数目和生长势与喷施空白对照的植株没有显著差异的株系为抗性转化事件。1x为保试达-18%草铵膦(德国拜耳)按体积比1:150稀释,2x为保试达18%草铵膦(德国拜耳)按体积1:75稀释,3x为保试达18%草铵膦(德国拜耳)按体积1:50稀释。我们获得的60个转pCambia1300-pUBI-Gat-Cry1Ab–p35S-1174载体的转基因株系(命名为GC)进行不同浓度草甘膦抗性测定,抗性效果如表2所示:表2*:1x2x3x抗性转化事件个数605858*注:表一为对播种后15天的水稻植株喷施不同浓度草铵膦进行抗性检测。喷施草甘膦10天后株高、叶片数目和生长势与喷施空白对照的植株没有显著差异的株系为抗性转化事件。1x为农达-41%草甘膦(美国孟山都)按体积比1:300稀释,2x为农达-41%草甘膦(美国孟山都)按体积比1:150稀释,3x为农达-41%草甘膦(美国孟山都)按体积比1:100稀释。实施例4、融合蛋白转基因水稻可以杀虫将实施例2方法制备的转基因水稻植株的T0代植株移栽到温室中,对转基因水稻植株和非转基因受体对照植株“秀水-134”的杀虫性能进行比较分析。我们获得的86个转pCambia1300-pUBI-bar-Vip3A–p35S-1174载体的转基因株系(命名为BV)进行抗虫性测定,抗性效果如表3所示:表3*:害虫有效转化事件个数杀虫率粘虫61100%甜菜夜蛾61100%*注:表一为对播种后30天的水稻叶片进行不同害虫的杀虫效果检测。每个实验都至少设置3个重复。我们获得的60个转pCambia1300-pUBI-Gat-Cry1Ab–p35S-1174载体的转基因株系(命名为GC)进行不同浓度草甘膦抗性测定,抗性效果如表4所示:表4*:害虫有效转化事件个数杀虫率玉米螟45100%棉铃虫45100%*注:表一为对播种后30天的水稻叶片进行不同害虫的杀虫效果检测。每个实验都至少设置3个重复。实施例5、玉米的转化玉米的转化技术已经比较成熟。参考文献如:VladimirSidorov&DavidDuncan(inM.PaulScott(ed.),MethodsinMolecularBiology:TransgenicMaize,vol:526;YujiIshida,YukohHiei&ToshihikoKomari(2007)Agrobacterium-mediatedtransformationofmaize.NatureProtocols2:1614-1622。基本方法如下:取授粉后8-10天的Hi-II玉米穗,收集所有的未成熟胚(大小为1.0-1.5mm)。将实施例1中制备的含有T-DNA载体pCambia1300-pUBI-bar-Vip3A–p35S-1174和pCambia1300-pUBI-Gat-Cry1Ab–p35S-1174的农杆菌与未成熟胚在共培养培养基上(MS+2mg/L2,4-D+30g/L蔗糖+3g/L琼脂(sigma7921)+40mg/L乙酰丁香酮)共培养2-3天(22℃)。转移未成熟胚到愈伤诱导培养基上(MS+2mg/L2,4-D+30g/L蔗糖+2.5g/Lgelrite+5mg/LAgNO3+200mg/L乙酰丁香酮),28℃暗培养10-14天。将所有的愈伤转到带有2mM草甘膦的筛选培养基(与愈伤诱导培养基相同)上,28℃暗培养2-3周。转移所有的组织到新鲜含草甘膦的筛选培养基上,28℃暗培养2-3周。然后,转移所有筛选后成活的胚性组织到再生培养基(MS+30g/L蔗糖+0.5mg/Lkinetin+2.5g/Lgelrite+200mg/L乙酰丁香酮)上,28℃暗培养10-14天,每皿一个株系。转移胚性组织到新鲜的再生培养基上,26℃光照培养10-14天。转移所有发育完全的植株到生根培养基(1/2MS+20g/L蔗糖+2.5g/Lgelrite+200mg/L乙酰丁香酮)上,26℃光照培养直到根发育完全。分别获得含转化载体pCambia1300-pUBI-bar-Vip3A–p35S-1174和pCambia1300-pUBI-Gat-Cry1Ab–p35S-1174的转基因玉米植株。实施例6、融合蛋白转基因玉米可以抗除草剂将实施例5制备的转基因玉米植株的T0代植株移栽到温室中,用商业化品种“郑丹958”的母本,“郑58”(Z58)的花粉进行授粉,收获T0代种子。然后将这些品系与商业化品种“郑丹958”的母本,“郑58”(Z58)进行回交转育,获得Z58近等位基因系。再对这些近等位基因系的除草剂抗性进行比较分析。我们获得的52个转pCambia1300-pUBI-bar-Vip3A–p35S-1174载体的转基因株系(命名为ZmBV)进行不同浓度草铵膦抗性测定,抗性效果如表5所示:表5*:1x2x3x抗性转化事件个数423219*注:表一为对3叶1心期的玉米植株喷施不同浓度草铵膦进行抗性检测。喷施草铵膦10天后株高、叶片数目和生长势与喷施空白对照(清水)的植株没有显著差异的株系为抗性转化事件。1x为保试达-18%草铵膦(德国拜耳)按体积比1:150稀释,2x为保试达18%草铵膦(德国拜耳)按体积比1:75稀释,3x为保试达18%草铵膦(德国拜耳)按体积比1:50稀释。我们获得的55个转pCambia1300-pUBI-Gat-Cry1Ab–p35S-1174载体的转基因株系(命名为ZmGC)进行不同浓度草甘膦抗性测定,抗性效果如表6所示:表6*:1x2x3x抗性转化事件个数555555*注:表一为对播种后15天的水稻植株喷施不同浓度草铵膦进行抗性检测。喷施草甘膦10天后株高、叶片数目和生长势与喷施空白对照的植株没有显著差异的株系为抗性转化事件。1x为农达-41%草甘膦(美国孟山都)按体积比1:300稀释,2x为农达-41%草甘膦(美国孟山都)按体积比1:150稀释,3x为农达-41%草甘膦(美国孟山都)按体积比1:100稀释。实施例7、融合蛋白转基因玉米可以杀虫将实施例5制备的转基因玉米植株的T0代植株移栽到温室中,对转基因玉米植株和非转基因受体对照植株“Z58”的杀虫性能进行比较分析。我们获得的52个转pCambia1300-pUBI-bar-Vip3A–p35S-1174载体的转基因株系(命名为ZmBV)进行抗虫性测定,抗性效果如表7所示:表7*:害虫有效转化事件个数杀虫率粘虫39100%甜菜夜蛾39100%*注:表一为对玉米叶片进行不同害虫的杀虫效果检测。每个实验都至少设置3个重复。我们获得的55个转pCambia1300-pUBI-Gat-Cry1Ab–p35S-1174载体的转基因株系(命名为ZmGC)进行抗虫性测定,抗性效果如表8所示:表8*:害虫有效转化事件个数杀虫率玉米螟42100%棉铃虫42100%*注:表一为对玉米叶片进行不同害虫的杀虫效果检测。每个实验都至少设置3个重复。实施例8.大豆转化这里使用的获得转基因大豆的步骤来自于已有的技术(Dengetal.,1998,PlantPhysiologyCommunications34:381-387;Maetal.,2008,ScientiaAgriculturaSinica41:661-668;Zhouetal.,2001,JournalofNortheastAgriculturalUniversity32:313-319)。选取健康、饱满、成熟的大豆,用80%乙醇消毒2分钟,再用无菌水清洗,然后放置在充满氯气(由50mlNaClO与2ml浓HCl反应生成)的干燥器中灭菌4-6个小时。灭菌后的“天隆1号”大豆在超净工作台里被播撒到B5培养基中,25℃条件下培养5天,同时光密度在90-150μmol光子/m2·s水平。当子叶变绿并顶破种皮,无菌的豆芽就会长出。去掉了下胚轴的豆芽在长度上被切成五五开,使得两片外植体都具有子叶和上胚轴。在子叶和上胚轴的节点处切外植体大约7-8处,即可用作被侵染的目标组织。分别含有载体pCambia1300-pUBI-bar-Vip3A-p35S-1174和pCambia1300-pUBI-Gat-Cry1Ab-p35S-1174的单克隆农杆菌被分开培养待用。准备好的外植体浸没在农杆菌悬浮液中共培养30分钟左右。然后,将侵染的组织上多余的细胞悬浮液用吸水纸吸收干净,再转移到1/10B5共培养培养基里,25℃暗培养3-5天。共培养的植物组织用B5液体培养基清洗,以除去多余的农杆菌,然后放置到B5固体培养基中25℃下培养5天,待其发芽。诱导发生的胚芽组织转移到含有0.1-0.5mM草甘膦的B5筛选培养基中,25℃光照培养4周,期间每两周更换一次培养基。筛选出来的胚芽组织再转移到固体培养基中,25℃培养,待其长成小苗。随后,将转基因植株苗转移到1/2B5培养基中进行生根诱导。最后,长成的小植株经清洗去除琼脂后栽种在温室中。实施例9、融合蛋白转基因大豆可以抗除草剂将实施例8方法制备的转基因大豆植株的T0代植株移栽到温室中,对转基因大豆植株和非转基因受体对照植株“天隆1号”的抗除草剂性能进行比较分析。我们获得的41个转pCambia1300-pUBI-bar-Vip3A-p35S-1174载体的转基因株系(命名为BV)进行不同浓度草铵膦抗性测定,抗性效果如表9所示:表9*:1x2x3x抗性转化事件个数312518*注:表一为对播种后20天的大豆植株喷施不同浓度草铵膦进行抗性检测。喷施草铵膦10天后株高、叶片数目和生长势与喷施空白对照的植株没有显著差异的株系为抗性转化事件。1x为保试达-18%草铵膦(德国拜耳)按体积比1:150稀释,2x为保试达18%草铵膦(德国拜耳)按体积比1:75稀释,3x为保试达18%草铵膦(德国拜耳)按体积比1:50稀释。我们获得的36个转pCambia1300-pUBI-Gat-Cry1Ab–p35S-1174载体的转基因株系(命名为GC)进行不同浓度草甘膦抗性测定,抗性效果如表10所示:表10*:1x2x3x抗性转化事件个数362821*注:表一为对播种后20天的大豆植株喷施不同浓度草铵膦进行抗性检测。喷施草甘膦10天后株高、叶片数目和生长势与喷施空白对照的植株没有显著差异的株系为抗性转化事件。1x为农达-41%草甘膦(美国孟山都)按体积比1:300稀释,2x为农达-41%草甘膦(美国孟山都)按体积比1:150稀释,3x为农达-41%草甘膦(美国孟山都)按体积比1:100稀释。实施例10、融合蛋白转基因大豆可以杀虫将实施例8方法制备的转基因大豆植株的T0代植株移栽到温室中,对转基因大豆植株和非转基因受体对照植株“天隆1号”的杀虫性能进行比较分析。我们获得的41个转pCambia1300-pUBI-bar-Vip3A–p35S-1174载体的转基因株系(命名为BV)进行抗虫性测定,抗性效果如表11所示:表11*:害虫有效转化事件个数杀虫率粘虫29100%甜菜夜蛾29100%*注:表一为对播种后30天的大豆叶片进行不同害虫的杀虫效果检测。每个实验都至少设置3个重复。我们获得的36个转pCambia1300-pUBI-Gat-Cry1Ab–p35S-1174载体的转基因株系(命名为GC)进行抗虫性测定,抗性效果如表12所示:表12*:害虫有效转化事件个数杀虫率玉米螟22100%棉铃虫22100%*注:表一为对播种后30天的大豆叶片进行不同害虫的杀虫效果检测。每个实验都至少设置3个重复。最后,还需要注意的是,以上列举的仅是本发明的具体实施例。显然,本发明不限于以上实施例,还可以有许多变形。本领域的普通技术人员能从本发明公开的内容直接导出或联想到的所有变形,均应认为是本发明的保护范围。当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 蝎神经毒素AaIT及其编码基因与应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种花狭口蛙的基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种泽陆蛙基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 降低种子重量的白桦AP2基因及其编码蛋白的制造方法与工艺
- 可编码ArtRD蛋白基因及其蛋白和应用的制造方法与工艺
- 蝎神经毒素AaIT及其编码基因与应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种花狭口蛙的基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种泽陆蛙基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 蛋白质及其编码基因在调控植物抗黄萎病性中的应用的制造方法与工艺
- 甘薯耐逆相关蛋白IbFBA2、编码基因、重组载体与应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 核盘菌凝集素SSL‑6蛋白及其编码基因与应用的制造方法与工艺
- 植物抗病毒相关蛋白GmSN1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 蛋白质及其编码基因在调控植物抗黄萎病性中的应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 核盘菌凝集素SSL‑6蛋白及其编码基因与应用的制造方法与工艺