微藻生物质的蛋白质富集方法与流程
大藻和微藻特别丰富,在很大程度上仍然未被探明。它们在饮食、化学制品或生物能源意图上的利用仍然是高度边缘状态的。然而,它们含有在丰富性和充裕性两方面都有很高价值的组分。
事实上,微藻是维生素、脂质、蛋白质、糖、色素和抗氧化剂的来源。
藻和微藻因此使工业部门感兴趣,在那里它们被用于制造食品补充剂、功能性食品、化妆品和药品,或用于水产养殖。
微藻是首要的光合微生物,定居于所有见光的群落生境。
在工业规模上,其单克隆培养在光生物反应器中进行(自养条件:光照加CO2),或者,有些也在发酵罐中进行(异养条件:存在碳源的情况下在黑暗中)。
有些微藻物种的确能够在没有光的情况下生长:小球藻属,菱形藻属(Nitzschia),小环藻属(Cyclotella),扁藻属(Tetraselmis),隐甲藻属(Crypthecodinium),裂壶藻属(Schizochytrium)。
此外,据估计,在异养条件下培养的花费比在光养条件下低10倍,因为对于本领域技术人员来说,这些异养条件允许:
-使用与用于细菌和酵母的发酵罐相同的发酵罐,能够使所有的培养参数受到控制,
-生物质的生产量远大于通过基于光的培养获得的量。
有利益的微藻利用一般需要控制发酵条件,使得可以积累它们的感兴趣组分,例如:
-色素(叶绿素a、b和c,β-胡萝卜素,虾青素,黄体素(lutein),藻青素,叶黄素(xanthophylls),藻红蛋白等),由于它们显著的抗氧化性质以及它们为食物提供天然着色剂这两方面,对所述色素的需求增加,
-脂质,以优化它们的脂肪酸含量(高达它们干固体重量的60%,或甚至80%),尤其是用于:
·生物燃料应用,以及
·当所选择的微藻产生“必需的”(即由饮食提供,因为它们不是由人类或动物天然产生的)多不饱和脂肪酸或PUFA时,在人类食用的食物或动物饲料中的应用,或
-蛋白质,以优化其营养质量或例如促进感兴趣的氨基酸的供应。
在供应感兴趣的氨基酸的情形下,具有富含精氨酸和谷氨酸的可用蛋白质源实际上可能是有利的。
精氨酸是一种在动物界具有许多功能的氨基酸。
精氨酸可以被降解并因此可充当吸收它的细胞的能量、碳和氮的来源。
在各种动物、包括哺乳动物中,精氨酸被分解为鸟氨酸和尿素。后者是一种含氮分子,其可以被清除(经由尿液排泄)以便调节动物生物体的细胞中存在的含氮化合物的量。
精氨酸允许经由NO合成酶合成一氧化氮(NO),从而参与动脉的血管舒张,它降低血管的僵硬性、增加血流量并由此改善血管的功能。
含有精氨酸的食物补充剂被推荐用于促进心脏健康和血管功能、防止“血小板聚集”(形成血块的风险)和降低动脉压。
精氨酸参与伤口愈合与它在脯氨酸形成中的作用有关,脯氨酸是胶原合成中的另一种重要的氨基酸。
最后,精氨酸是能量饮料中经常使用、特别是被运动者使用的组分。
至于谷氨酸,它不仅是用于蛋白质合成的基本结构单元之一,而且也是中枢神经系统(脑髓+脊髓)中最广泛分布的兴奋性神经递质,并且是GABA能神经元中的GABA前体。
在代码E620下,谷氨酸被用作食品中的增味剂。将它添加于食物制品来提高它们的味道。
除谷氨酸之外,食品法典委员会(Codex Alimentarius)还认可其钠盐(E621)、钾盐(E622)、钙盐(E623)、铵盐(E624)和镁盐(E625)作为增味剂。
谷氨酸(或其盐)经常存在于现成餐食(汤,调味汁,薯片和现成菜肴)中。它也常用于亚洲烹饪。
它目前经常与调味品相组合用于开胃酒(咸肉风味,奶酪风味)。这可以增强咸肉、奶酪等风味。很少发现不含任何谷氨酸的开胃酒。
它也存在于某些药物胶囊中,但不是由于它的味道功能。
最后,它是烹饪助剂(固体汤料,酱汁底料,调味汁等)的主要组分。
为了实现微藻代谢丰富性的探查,因此全面研究了用于获得高细胞密度(HCD)的第一发酵法以获得最高的蛋白质或脂质产量和生产力。
这些HCD培养的目的是在尽可能短的时间内获得最高可能浓度的目标产物。
这个原则例如通过由Chlorella zofingiensis生物合成虾青素证实,其中证明所述微藻的生长与该化合物的产生直接相关(Wang和Peng,2008,World J.Microbiol.Biotechnol.,24(9),1915-1922)。
然而,将生长维持在最高速率(μ,以h-1计)下并不总是与目标产物的高生产相关。
实际上,对于该领域的专业人员来说很快会明了,例如,当需要使微藻产生大量的脂质储备时,有必要使微藻经受限制它们生长的营养胁迫。
因此现在进行发酵方法中的生长/生产解耦以及细胞生长速率控制。
一般而言,本领域技术人员选择通过控制发酵条件(温度,pH等)或通过调节发酵培养基的营养组分供应(半连续条件称为“补料分批(fed batch)”)来控制微藻的生长。
如果他们选择通过碳源的供应来异养地控制微藻的生长,那么本领域技术人员一般选择根据所产生的代谢物(例如DHA型的多不饱和脂肪酸),使碳源(纯葡萄糖,乙酸盐,乙醇等)适应微藻(C.cohnii,细小裸藻(Euglena gracilis)等)。
温度也可以是一个关键参数:
-例如,据报道,在一些微藻物种中多不饱和脂肪酸的合成,例如微小小球藻(Chlorella minutissima)合成EPA,在低于所述微藻最佳生长所需的温度下被促进;
-另一方面,当生产温度从24℃增至35℃时,异养培养的原始小球藻中的黄体素产量更高。
事实上,原始小球藻被公认是最好的产油微藻之一。
在异养条件下,它迅速将碳水化合物转化成甘油三酯(超过干物质的50%)。
为了优化这种甘油三酯生产,本领域技术人员被引导至通过作用于发酵培养基的营养环境来优化碳流朝向产油。
因此,已知当有足够的碳供应但在氮缺乏的条件下时油积累。
因此,C/N比率在此是决定性因素,并且认为最佳结果通过直接作用于氮含量获得,葡萄糖含量不是限制性因素。
不出所料,这种氮缺乏影响细胞生长,这导致生长速率比所述微藻的正常生长速率低30%(Xiong等,Plant Physiology,2010,154,第1001-1011页)。
为了解释这个结果,上面提到的Xiong等的论文证明,如果小球藻生物质被分成其5个主要组分,即碳水化合物、脂质、蛋白质、DNA和RNA(占干物质的85%),虽然C/N比率对DNA、RNA或碳水化合物的含量没有影响,但它对蛋白质和脂质的含量变得显著。
因此,以低C/N比率培养的小球藻细胞含有25.8%的蛋白质和25.23%的脂质,而高C/N比率使得可能合成53.8%的脂质和10.5%的蛋白质。
为了优化它的产油,因此本领域技术人员必须控制碳流,通过将其转向产油来不利于蛋白质生产;当微藻被放置在缺氮培养基中时,碳流被重新分配并积累作为脂质储存物质。
然而,也可以有利地选择原始小球藻来产生蛋白质。
考虑到本领域技术人员关于管理用于产油的C/N比率(针对高C/N比)进行的分析,本领域技术人员因此被引导至偏爱低C/N比率,并且由此:
-在保持将转化为蛋白质的碳源原料不变的同时,向发酵培养基供应大量的氮源,以及
-刺激所述微藻的生长。
因此选择改变碳流朝向蛋白质(并因此朝向生物质)生产,而不利于储存脂质的生产。
本发明涉及微藻生物质、更特别是小球藻属、更加特别是原始小球藻物种的蛋白质富集方法。
本发明涉及用于某些微藻、更特别是原始小球藻的生物质的蛋白质富集的方法,所述生物质的蛋白质的精氨酸和谷氨酰胺含量显著高。
本发明更特别覆盖用于生产富含蛋白质的微藻的生物质的方法,其特征在于所述方法包括增加先前缺氮的生物质中的铵供应。
具体而言,在本发明的情形下,申请人公司在另一方面已经选择通过提出对本领域技术人员常规设想的方案的替代解决方案,或甚至与本领域技术人员会选择的恰好相反的方案,来探索新的路线。
本发明因此涉及异养培养的微藻的蛋白质富集方法,所述微藻是小球藻属的微藻,更加特别是原始小球藻,所述异养培养方法包括:
-第一步骤,所述第一步骤旨在限制铵供应以获得具有按N.6.25表示的小于50%、优选小于30%、优先在20%和25%之间的蛋白质含量的微藻生物质;
-第二步骤,在所述第二步骤中增加发酵培养基中的铵供应以获得大于50%、优选大于60%、更加优先大于65%的蛋白质含量。
如下文将说明的,根据本发明的方法的优先实施方式可以包括用NH3/KOH混合物调节第一步骤中的pH,由此限制铵供应并因此促进产生小蛋白质含量,然后在第二步骤中仅用NH3进行pH调节,以便将铵再供应给所述发酵培养基。
所述NH3/KOH混合物将使得它可以限制铵的供应。例如,对于第一和第二步骤之间相同浓度的NH3而言,所述混合物可以包括约1:1左右的NH3/KOH比率,例如约70-45%NH3和30-55%KOH,优选约65-55%v/v NH3和35-45%KOH,这些量以摩尔表示。
术语“约”旨在表示包含指示值的加或减10%的值范围,优选地为其加或减5%。例如,“约10”意指9和11之间,优选9.5和10.5之间。
在这个优先实施方式中,然后将NH3供应乘以约1.5至2,由此产生的氮消耗速度乘以5。
该第二步骤是这样的步骤,在此期间给之前缺氮的生物质增加铵供应,导致该生物质对该盐的散发性过度消耗,并显著导致蛋白质含量的提升,直至含量大于50%,优选大于60%,优先大于65%(百分比按N.6.25表示)。
由此发现,在氮缺乏阶段期间下降到小于0.005g/g/h的值的氮消耗比速度,在氮缺乏解除后增加到大于0.01g/g/h的值。
在一个优选实施方式中,生长速率保持基本不变。例如,在这两个阶段期间,生长速率保持在0.07h-1至0.09h-1,优选约0.08h-1。
为了说明这个概念,本发明更具体地覆盖了异养培养原始小球藻的方法,所述方法包括:
-分批阶段,其在接种所述发酵罐之后,供应20g/l的葡萄糖
-生长速率设定为0.08h-1的指数型补料分批阶段,其开始于所述分批中供应的葡萄糖被完全消耗时,
在此期间,为了获得含有少于25%蛋白质(按N.6.25表示)的生物质,通过利用NH3和KOH的混合物进行pH调节来限制铵供应,
以及然后
-具有设定为0.08h-1的相同生长速率的指数型补料分批阶段,在此期间通过利用100%氨水溶液调节pH来解除铵缺乏。
例如,当引入约2kg干葡萄糖时,启动其中解除铵缺乏的第二补料分批阶段。
对于本发明的目的而言,基本标准明确在于,在非常特定的条件下,通过发酵培养基的氮缺乏造成细胞胁迫、随后解除该胁迫,引发引入的氮的消耗以提升所产生的生物质的蛋白质含量。
因此,这种策略完全违背了技术偏见,所述技术偏见认为要提高生物质的蛋白质含量,从培养开始就增加氮的供应是绝对必要的。
此外,这些操作条件在此不仅通过蛋白质丰富度的增加而得到反映,而且还导致明显增加其精氨酸和谷氨酸含量。
更特别地,如下面将要说明的那样,原始小球藻物种的微藻异养培养包含在氮缺乏和然后在铵脉冲下培养的步骤,其导致产生相对于总氨基酸而言大于45%的谷氨酸和精氨酸。
因此,本发明还涉及用于富集异养培养的微藻、优选原始小球藻物种的微藻的谷氨酸和/或精氨酸含量的方法,所述方法包括所述微藻的异养培养,其包含旨在限制铵供应以获得贫蛋白质的微藻生物质的步骤,继之以维持生长速率并增加铵供应的步骤。
这些培养条件因此导致制备的微藻生物质包含相对于总氨基酸而言大于45%的谷氨酸和精氨酸。
从以下旨在说明性而非限制性的实施例中,将更清楚地理解本发明。
附图说明
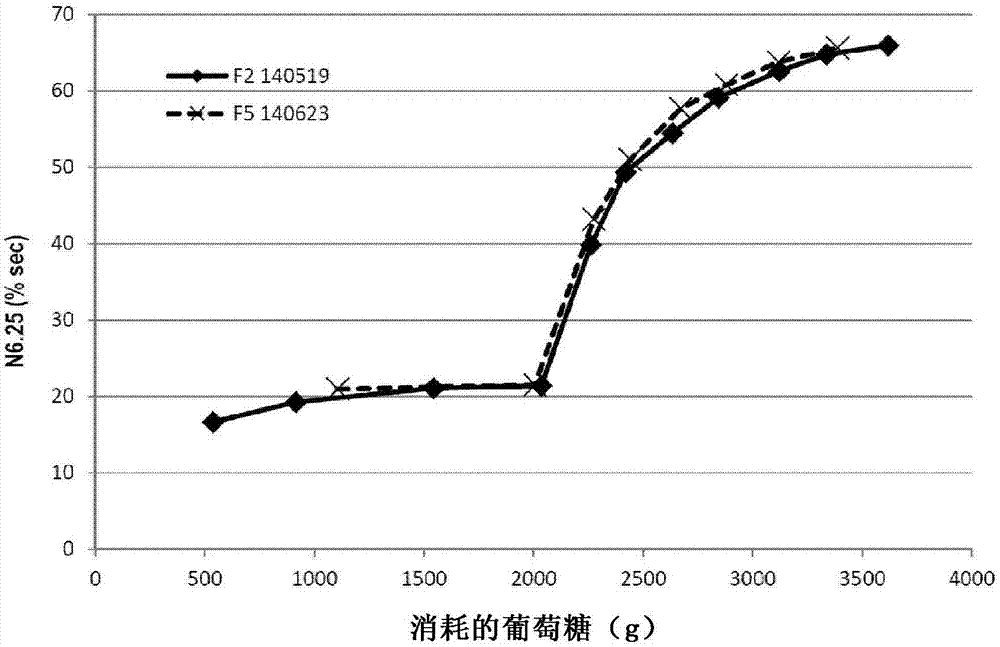
图1示出了N.6.25随着所消耗的葡萄糖的变化。
图2示出了氮消耗的比速率(qN)随着所消耗的葡萄糖的变化。
图3示出了N.6.25随时间的变化以及按干生物质重量百分比计的各氨基酸的量随时间的变化。
图4示出了N.6.25随时间的变化以及按干生物质重量百分比计的总脂肪酸或特定脂肪酸的量随时间的变化。
图5示出了N.6.25随时间的变化以及按干生物质重量百分比计的总糖或特定糖的量随时间的变化。
实施例
实施例1:富含具有高含量的谷氨酸和精氨酸的蛋白质的原始小球藻生物质的制备
使用的菌株是原始小球藻(菌株CCAP211/8D–藻类和原生动物培养物保藏中心(The Culture Collection of Algae and Protozoa),英国苏格兰)。
预培养:
-在500mL锥形瓶中的150mL培养基;
-培养基的组成:40g/L的葡萄糖+10g/L的酵母提取物。
在以下条件下进行温育:
-时间:72h;
-温度:28℃;
-振荡:110rpm(Infors Multitron培养箱)。
以分批和然后补料分批模式进行培养
制备和初始分批培养基
-制备和过滤400g/l的KOH(41%)/20%v/v的NH3(59%)的混合物;
-以121℃/20min灭菌20L发酵罐;
-用预培养物(OD600nm为15)接种2个500mL的锥形瓶;
-用KOH/NH3混合物调节pH为5.2;
-起始振荡速度为300rpm;
-通气:15L/min的空气;
-通过改变振荡将pO2调节在30%;
-温度:28℃
进料
-葡萄糖:500g/L
-硫酸铵:25g/L
-磷酸二氢钠:17g/L
-磷酸二氢钾:23g/L
-七水合硫酸镁:20g/L
-硫酸铁:120mg/L
-硝酸钙:610mg/L
-微量元素溶液:45mL/L
-维生素溶液:3.6mL/L
发酵程序
-接种前提供等量的20g/L葡萄糖
-当葡萄糖浓度=0g/L时,开始以补料分批模式供给葡萄糖;使用可以将生长速率设定在0.08h-1的流动速率
-用41%KOH/NH3混合物调节至pH 5.2
-当2kg葡萄糖已被微藻消耗时,将系统切换到仅用NH3进行pH调节,
-当生物质达到以干物质重量计为100g/L、并且已经供给了约3.5kg葡萄糖时,停止葡萄糖供给。
结果:
在这些相同的条件下进行两个试验,结果在表I和下面的图中给出:
表I.
图1示出了N.6.25随着所消耗的葡萄糖的变化。这两个试验反映了一些值得注意的结果:产生N.6.25含量大于65%的黄色生物质。
图2示出了氮消耗的比速率(qN)随着所消耗的葡萄糖的变化。
可以看出,在解除氮限制后(在消耗2kg葡萄糖时),氮消耗的比速率处于最大值,然后逐渐降低。两个试验之间的相似速率也反映了该方案的良好可重复性。
对临解除所述限制之前采集的样品和在所述脉冲之后的几个样品进行生物质中存在的氨基酸的全面分析。
结果在图3中显示。
注意到,在临解除氮限制之前,氨基酸的总和低(16.3%),并且各种氨基酸之间没有占优势的。
解除氮限制后1小时,注意到经历最大增加的氨基酸是谷氨酸,接着是精氨酸。其他氨基酸的含量也增加,但程度低得多。
因此N.6.25的增加首先与谷氨酸和精氨酸的增加相关。
除了这些分析之外,还对临解除氮限制之前采集的样品和在所述脉冲之后的几个样品进行生物质中存在的脂肪酸的全面分析。
结果在图4中显示。
生物质中脂肪酸的总含量在脉冲前为19.2%,降至10.2%。按照该曲线的优势脂肪酸是油酸。
因此当氮缺乏时,脂肪酸在所述生物质中积累。
对临解除氮限制之前采集的样品和对解除氮限制之后的几个样品也进行了生物质中存在的糖的全面分析。结果在图5中显示。
生物质中糖的总含量在解除氮限制之前为37.5%,降至20%,然后停滞。按照该曲线的优势糖是葡萄糖。
因此,当缺乏氮时,糖也存储在所述生物质中。
糖的含量然后显得稳定,不像脂肪酸含量那样持续下降。
所述生物质的盐含量通过测量煅烧残渣来测量:它是9%。
- 还没有人留言评论。精彩留言会获得点赞!