与植物抗病性相关的水稻OsGRDP1蛋白及其编码基因与应用的制作方法
本发明属于植物基因工程领域,涉及与植物抗病性相关的水稻osgrdp1蛋白及其编码基因与应用。
背景技术:
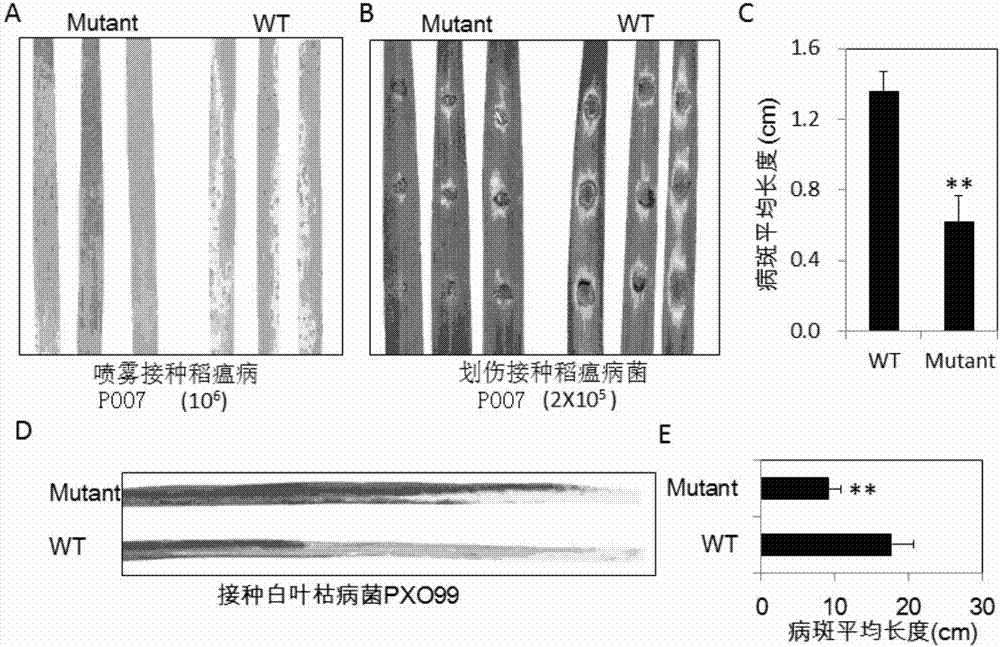
:过敏性反应(hypersensitiveresponse,hr)是植物中最为普遍且有效的防卫反应之一,其特征在于植物受到病原物侵染时,在侵染位点周围迅速产生局部细胞坏死,从而限制病原物向相邻细胞的扩展(morelanddangl,1997)。虽然过敏性反应在植物抗病性中的作用得到了广泛认可,但有关其机制的研究还不完善。为了研究过敏性反应的机制,不少研究者正在分离植物的类病斑突变体(lessionmimicmutants,lmms),期望首先通过遗传途径确定控制过敏性反应的基因,然后研究该基因的分子功能。类病斑突变体是植物某些基因表达异常的突变体。该类突变体在未受到病原物侵袭时,组成型表现类似病斑症状或过敏性细胞坏死的表型。已有的研究表明,多数lmms在未受到病原物侵染时,一些防卫基因组成型表达且抗病性增强,说明lmm基因是植物防卫反应中细胞坏死的负调控因子。因此,分离、鉴定lmm突变体、解析其类病斑形成及参与植物防卫反应的机制,是深入了解植物抗病机制的重要途径。迄今,已从水稻、玉米、拟南芥、大麦等多种植物中鉴定出了大量的lmm突变体。例如,通过自然变异和人工诱发突变筛选等方法,国内外已从水稻中至少鉴定出了43个不同类型的lmm突变体,其中多数突变体都表现了对稻瘟病和/或白叶枯病的抗性增强,如spl4、spl5、spl7、spl10、spl11、spl12、spl13、spl14、spl15、spl17、spl18、spl26、spl28和spl30。水稻中的lmm突变体,依据其突变表型的特点可以分为叶斑型(spottedleaf,spl)、褐斑型(brownleafspot,bl)、“斑马叶”坏死型(zebranecrosis,zn)、细胞坏死和抗性增强型(celldeathandresistance,cdr)、类稻瘟病斑(blastlesionmimic,blm)、黄化叶斑型(yellowleafspot,ysl),以及噻菌灵诱导型(probenazol,pbz)等。尽管如此,被克隆lmm基因依然为数很少。spl7是水稻第一个被克隆的lmm基因,该基因编码一热激转录因子,紫外线和高温可诱导其类病斑形成(yamanouchietal.2002)。spl11是另外一个控制类病斑形成的基因,它编码一个e3泛素连接酶。spl11突变体不仅对病原物具有广谱抗性,还调控水稻开花。ospti1a基因的突变体自发产生过敏性病斑,对稻瘟病抗性增强。ospti1a编码与番茄pti1同源的蛋白激酶,pti1通过与pto物理互作从而参与pto介导的抗病反应。spl18编码一个乙酰转移酶,该蛋白在烟草中的同源基因表达受hr反应诱导。spl1(也称为为sekiguchilesion)编码一个细胞色素p450单加氧酶家族成员,该酶具有色胺5’-羟基化酶活性,催化色胺形成5-羟色胺色胺。spl28编码网格蛋白相关的连接蛋白复合体中的中间亚基μ1(ap1m1)。spl28突变体的类病斑周围可累积高浓度的活性氧、植保素、胼胝质以及酚类化合物,对稻瘟病和白叶枯病的抗性均增强。spl5编码剪切因子3b的亚基3(sf3b3),说明spl5可能通过调控rna的剪切负向调控植物细胞死亡。oslms(lesionmimicandsenescence)编码一个含有两个双链rna结合域的蛋白质,该基因的突变导致产生类病斑和早衰表型,且对稻瘟病的抗性增强(undanetal.2012)。最近被克隆的lmr(lesionmimicresembling)基因编码aaa-typeatpase,该基因的突变使pr1和pbz1基因表达上调,对稻瘟病和白叶枯病的抗性增强。上述已克隆的lmm基因均为隐性基因,是防卫反应的负调控因子。但是,最近有研究报道oscard1(acceleratedcelldeathandresistance1)过表达导致自发产生过敏性病斑,该基因编码mapkkk激酶,其过表达增强了对稻瘟病的抗性,说明其是防卫反应的正调控因子。总的来说,国内外研究已鉴定了相当数量的lmm突变体,也已克隆了一定数量的lmm基因。目前已知lmm基因编码蛋白的功能呈现多样性,一定程度上反映了类病斑形成的复杂机制,也因此尚未建立一个完整的、有关过敏性反应形成的机制。但是,大多数lmm突变体表现广谱抗病性。因此,不断鉴定新型的lmm突变体,确定相关基因及其分子功能,将有助于全面揭示过敏性反应形成的机制,也将有助于深入揭示植物广谱抗病的分子机制,进而为抗病性的遗传改良提供依据。技术实现要素:本发明所要解决的技术问题是如何提高植物抗病性。为解决上述技术问题,本发明首先提供了一种与植物抗病性相关蛋白。本发明所提供的与植物抗病性相关蛋白的名称为osgrdp1,为如下a)或b)或c)或d)的蛋白质:a)氨基酸序列是序列2所示的蛋白质;b)在序列2所示的蛋白质的n端和/或c端连接标签得到的融合蛋白质;c)将序列2所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的具有相同功能的蛋白质;d)与序列2所示的氨基酸序列具有75%或75%以上的同源性且具有相同功能的蛋白质。其中,序列2由930个氨基酸残基组成。为了使a)中的蛋白质便于纯化,可在序列表中序列2所示的蛋白质的氨基末端或羧基末端连接上如表1所示的标签。表1、标签的序列标签残基序列poly-arg5-6(通常为5个)rrrrrpoly-his2-10(通常为6个)hhhhhhflag8dykddddkstrep-tagii8wshpqfekc-myc10eqkliseedl上述c)中的蛋白质osgrdp1,所述一个或几个氨基酸残基的取代和/或缺失和/或添加为不超过10个氨基酸残基的取代和/或缺失和/或添加。上述c)中的蛋白质osgrdp1可人工合成,也可先合成其编码基因,再进行生物表达得到。上述c)中的蛋白质osgrdp1的编码基因可通过将序列1所示的dna序列中缺失一个或几个氨基酸残基的密码子,和/或进行一个或几个碱基对的错义突变,和/或在其5′端和/或3′端连上表1所示的标签的编码序列得到。为解决上述技术问题,本发明又提供了与osgrdp1蛋白质相关的生物材料。本发明提供的与osgrdp1蛋白质相关的生物材料为下述a1)至a12)中的任一种:a1)编码osgrdp1蛋白质的核酸分子;a2)含有a1)所述核酸分子的表达盒;a3)含有a1)所述核酸分子的重组载体;a4)含有a2)所述表达盒的重组载体;a5)含有a1)所述核酸分子的重组微生物;a6)含有a2)所述表达盒的重组微生物;a7)含有a3)所述重组载体的重组微生物;a8)含有a4)所述重组载体的重组微生物;a9)含有a1)所述核酸分子的转基因植物细胞系;a10)含有a2)所述表达盒的转基因植物细胞系;a11)含有a3)所述重组载体的转基因植物细胞系;a12)含有a4)所述重组载体的转基因植物细胞系。上述生物材料中,a1)所述核酸分子为如下1)或2)或3)所示的基因:1)其编码序列是序列1所示的cdna分子或基因组dna分子;2)与1)限定的核苷酸序列具有75%或75%以上同一性,且编码osgrdp1蛋白质的cdna分子或基因组dna分子;3)在严格条件下与1)或2)限定的核苷酸序列杂交,且编码osgrdp1蛋白质的cdna分子或基因组dna分子。其中,所述核酸分子可以是dna,如cdna、基因组dna或重组dna;所述核酸分子也可以是rna,如mrna或hnrna等。其中,序列1由2970个核苷酸组成,整个序列1即为所述osgrdp1基因的编码序列(orf),编码序列表中序列2所示的蛋白质。本领域普通技术人员可以很容易地采用已知的方法,例如定向进化和点突变的方法,对本发明的编码osgrdp1的核苷酸序列进行突变。那些经过人工修饰的,具有与本发明分离得到的osgrdp1的核苷酸序列75%或者更高同一性的核苷酸,只要编码osgrdp1且具有相同功能,均是衍生于本发明的核苷酸序列并且等同于本发明的序列。这里使用的术语“同一性”指与天然核酸序列的序列相似性。“同一性”包括与本发明的编码序列2所示的氨基酸序列组成的蛋白质的核苷酸序列具有75%或更高,或85%或更高,或90%或更高,或95%或更高同一性的核苷酸序列。同一性可以用肉眼或计算机软件进行评价。使用计算机软件,两个或多个序列之间的同一性可以用百分比(%)表示,其可以用来评价相关序列之间的同一性。上述75%或75%以上同一性,可为80%、85%、90%或95%以上的同一性。上述生物材料中,a2)所述的含有编码osgrdp1的核酸分子的表达盒(osgrdp1基因表达盒),是指能够在宿主细胞中表达osgrdp1的dna,该dna不但可包括启动osgrdp1转录的启动子,还可包括终止osgrdp1转录的终止子。进一步,所述表达盒还可包括增强子序列。可用于本发明的启动子包括但不限于:组成型启动子;组织、器官和发育特异的启动子及诱导型启动子。合适的转录终止子包括但不限于:农杆菌胭脂碱合成酶终止子(nos终止子)、花椰菜花叶病毒camv35s终止子、tml终止子、豌豆rbcse9终止子和胭脂氨酸和章鱼氨酸合酶终止子。所述植物重组表达载体可用现有的植物表达载体构建。所述植物表达载体包括双元农杆菌载体和可用于微弹轰击的载体等,如pgreen0029、pcambia3301、pcambia1300、pcambia1301、pbi121、pbin19、pcambia2301、pcg1301或其他衍生植物表达载体。所述植物表达载体还可包含外源基因的3’端非翻译区,即包含多聚腺苷酸信号和任何其他参与mrna加工或基因表达的dna片段。所述多聚腺苷酸信号可引导多聚腺苷酸加入到mrna前体的3’端。使用所述基因构建重组表达载体时,在其转录起始核苷酸前可加上任何一种增强型、组成型、组织特异型或诱导型启动子,例如花椰菜花叶病毒(camv)35s启动子、泛素基因ubiquitin启动子(pubi)、胁迫诱导型启动子rd29a等,它们可单独使用或与其它的植物启动子结合使用;此外,使用本发明的基因构建重组表达载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是atg起始或邻近区域起始密码子等,但必须与编码序列的阅读框相同,以保证整个序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的,也可以是人工合成的。翻译起始区域可以来自转录起始区域或结构基因。为了便于对转基因植物细胞或植株进行鉴定和筛选,可对所用植物表达载体进行加工,如加入可在植物中表达的编码可产生颜色变化的酶或发光化合物的基因、具有抗性的抗生素标记物或是抗化学试剂标记基因等。也可以不加任何选择标记基因,直接以逆境筛选转化植株等。在本发明中,所述重组表达载体中启动所述编码基因osgrdp1转录的启动子可为ubiquitin启动子、35s启动子或actin1启动子。在本发明中,启动所述编码基因osgrdp1转录的启动子具体为ubiquitin启动子。上述生物材料中,所述载体可为质粒、黏粒、噬菌体或病毒载体。在本发明中,所述重组载体具体为将ubiquitin启动子和osgrdp1基因插入pcambia1301载体的saci和mlui酶切位点间后得到的载体。上述生物材料中,所述微生物可为酵母、细菌、藻或真菌,如农杆菌。在本发明中,采用的农杆菌具体为eha105。上述生物材料中,所述转基因植物细胞系均不包括繁殖材料。为解决上述技术问题,本发明还提供osgrdp1蛋白质或上述生物材料的新用途。本发明提供了osgrdp1蛋白质或上述生物材料在调控植物抗病性中的应用。本发明还提供了osgrdp1蛋白质或上述生物材料在调控植物类病斑的形成中的应用。本发明还提供了osgrdp1蛋白质或上述生物材料在培育抗病性提高的转基因植物中的应用。本发明还提供了osgrdp1蛋白质或上述生物材料在植物育种中的应用。上述应用中,所述调控植物抗病性为提高植物抗病性,具体体现在如下m1)或m2):m1)提高植物对水稻稻瘟病和/或水稻白叶枯病的抗性;m2)提高植物防卫相关基因的表达量。所述防卫相关基因具体为基因pr1a和/或pr1b和/或pr2和/或pr3和/或pr5和/或pr8和/或pr9和/或pr10。上述应用中,所述调控植物类病斑的形成为促进植物类病斑形成,具体体现在如下m3)或m4):m3)使植物自发形成类病斑;m4)在植物类病斑产生部位检测到活性氧突发。为解决上述技术问题,本发明最后提供了一种培育抗病性提高的转基因植物的方法。本发明提供的培育抗病性提高的转基因植物的方法包括提高受体植物中osgrdp1蛋白质的表达量和/或活性,得到转基因植物的步骤;所述转基因植物的抗病性高于所述受体植物。上述方法中,所述提高受体植物中osgrdp1蛋白质的表达量和/或活性的方法为在受体植物中过表达osgrdp1蛋白质。上述方法中,所述过表达的方法为将osgrdp1蛋白质的编码基因导入受体植物;所述osgrdp1蛋白质的编码基因的核苷酸序列是序列1所示的dna分子。上述方法中,所述将osgrdp1蛋白质的编码基因导入受体植物为将携带有所述osgrdp1蛋白质的编码基因的重组载体导入所述受体植物,具体可为使用ti质粒、ri质粒、植物病毒载体、直接dna转化、显微注射、电导、农杆菌介导等常规生物学方法转化受体植物组织或细胞,并将转化的植物组织培育成植株。所述重组载体可为将ubiquitin启动子和osgrdp1基因插入pcambia1301载体的saci和mlui酶切位点间后得到的载体。上述方法中,所述转基因植物的抗病性高于所述受体植物体现在转基因植物可自发形成类病斑。上述方法中,所述转基因植物理解为不仅包含将所述osgrdp1基因转化受体植物得到的第一代转基因植物,也包括其子代。对于转基因植物,可以在该物种中繁殖该基因,也可用常规育种技术将该基因转移进入相同物种的其它品种,特别包括商业品种中。所述转基因植物包括种子、愈伤组织、完整植株和细胞。在上述应用或方法中,所述抗病性为抗水稻稻瘟病和/或水稻白叶枯病。在上述应用或方法中,所述植物为单子叶植物或双子叶植物,所述单子叶植物为水稻、玉米、小麦等。在本发明中,所述植物为单子叶植物,所述单子叶植物为水稻,所述水稻品种具体为爱知旭。本发明利用一个水稻显性类病斑突变体spl-d,使用tail-pcr技术克隆出osgrdp1基因并进行了功能分析。突变体spl-d中t-dna插入在osgrdp1转录起始位点上游,导致osgrdp1表达增强,表现出自发形成类病斑,与野生型相比增强了对水稻稻瘟病和白叶枯病的抗性。进一步,利用ubiquitin启动子驱动osgrdp1基因在野生型水稻中超表达,超表达植株的表型与突变体spl-d一致,说明osgrdp1基因在增强水稻抗病性中具有较好的应用潜力。本发明为利用基因工程手段行进分子育种增强植物抗病性具有潜在的应用价值。附图说明图1为类病斑突变体spl-d与野生型(wt)的生长表型及其叶片活性氧的检测。其中,a为田间(北京)种植条件下叶片,叶鞘以及全株的类病斑的产生情况;b为dab染色检测类病斑周围活性氧的积累结果。图2为类病斑突变体spl-d与野生型(wt)接种稻瘟菌小种p007及水稻白叶枯菌pxo99的表型。其中,a为喷雾接种稻瘟菌小种p007的发病结果;b为划伤接种稻瘟菌小种p007的发病结果;c为划伤接种稻瘟菌小种p007后测量的病斑长度(数据为15个病斑长度的平均值,**:p<0.01,学生t测验);d为接种水稻白叶枯菌pxo99的发病结果;e为接种水稻白叶枯菌pxo99后测量的病斑长度(数据为10个病斑长度的平均值,**:p<0.01,学生t测验)。图3为利用实时定量pcr检测类病斑突变体spl-d与野生型(wt)的8个防卫基因的相对表达量结果(a-e)(**:p<0.01,学生t测验)。图4为利用tail-pcr克隆水稻osgrdp1基因的结果。a为t-dna在osgrdp1基因启动子区的插入位置;b为pcr鉴定t-dna的插入位点结果;c为利用实时定量pcr检测类病斑突变体spl-d与野生型(wt)中osgrdp1基因的相对表达量结果(**:p<0.01,学生t测验)。图5为osgrdp1的亚细胞定位结果。其中,a为osgrdp1-gfp在烟草表皮细胞中的定位结果;b为osgrdp1-gfp在水稻原生质体中的定位结果。图6为t0代转osgrdp1水稻株系中osgrdp1基因的表达分析及其叶片产生类病斑的表型。其中,a为利用实时定量pcr检测t0代转osgrdp1水稻株系中osgrdp1基因的相对表达量结果(**:p<0.01,学生t测验);b为t0代转osgrdp1水稻株系的叶片产生类病斑的结果。具体实施方式下述实施例中所使用的实验方法如无特殊说明,均为常规方法。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。下述实施例中的定量试验,均设置三次重复实验,结果取平均值。下述实施例中的稻瘟菌小种p007记载于文献“稻瘟病菌生理小种及其毒性”中,公众可从中国农业大学获得。下述实施例中的水稻白叶枯病菌株pxo99记载于文献“小粒野生稻抗白叶枯病新基因的鉴定与初步定位”中,公众可从中国农业大学获得。实施例1、水稻osgrdp1蛋白及其编码基因的获得一、突变体spl-d的获得及其抗病性状检测1、突变体spl-d的获得及其表型从t-dna转化水稻品种爱知旭的后代中筛选获得突变体spl-d。和野生型水稻品种爱知旭相比,突变体spl-d自发产生类病斑,该突变体的表型如图1中a所示。其中,wt为野生型水稻品种爱知旭,spl-d为突变体水稻。在长日照条件下(北京,5月-10月,日照时间约12-14小时),此突变体在水稻幼苗期和野生型几乎没有差异。随着发育的进行,进入分蘖期以后从下部叶片至上部叶片陆续出现类病斑,类病斑可扩展到叶鞘;在短日照条件下(北京,11月至翌年3月,日照时间约8-10小时),此突变体在苗期(四叶一心期)即可出现类病斑。2、叶片活性氧的检测利用3’3’-diaminobenzidine(dab)对突变体spl-d和野生型水稻爱知旭叶片进行染色实验。具体步骤参照文献“qunenliu,yuesening,yingxinzhang,ningyu,chundezhao,xiaodengzhan,weixunwu,daibochen,xiangjinwei,guo-liangwang,shihuacheng,liyongcao,2017,oscul3anegativelyregulatescelldeathandimmunitybydegradingosnpr1inrice,plantcell,29(2):345-359,doi:10.1105”中的方法进行。结果如图1中b所示。突变体spl-d的类病斑产生部位可明显地检测到活性氧突发,而野生型则检测不到。3、接菌试验(1)利用稻瘟菌小种p007对突变体spl-d和野生型水稻爱知旭叶片进行喷雾和离体划伤接种。结果如图2所示。划伤接种后120小时,突变体spl-d的病斑长度仅为野生型的1/2,说明突变体spl-d对稻瘟病菌的抗性显著增强。(2)利用水稻白叶枯病菌株pxo99接种突变体spl-d和野生型水稻爱知旭叶片。结果如图2所示。接种2周后,突变体spl-d的病斑长度显著小于野生型,说明突变体spl-d对白叶枯病菌的抗性显著增强。4、防卫反应基因的表达情况利用荧光定量pcr方法分析突变体spl-d和野生型水稻爱知旭中防卫反应基因的表达情况。具体步骤如下:取突变体spl-d和野生型水稻相同部位完全展开叶片,采用trizol试剂法(invitrogen)提取其总rna,采用mmlv反转录酶(takara)并参照相应的使用方法进行反转录,然后利用实时荧光定量pcr技术,根据厂家(takara)提供的使用方法,在pcr体系中加入sybrgreeni荧光染料,在荧光定量pcr仪(abi7500,usa)检测防卫相关基因pr1a、pr1b、pr2、pr3、pr5、pr8、pr9、pr10的表达情况,以水稻actin1基因为内参。引物序列如表1。实验设三次重复。数据处理采用comparativect方法,即ct值为pcr管中荧光信号达到设定的阈值时所经历的循环数,δct=ct(待测基因)-ct(actin1),以2-δct值衡量基因转录水平,对突变体及野生型水稻中的防卫基因进行比较分析。表1、actin1及防卫基因的引物序列结果如图3所示。所检测的防卫基因pr1a、pr1b、pr2、pr3、pr5、pr8、pr9、pr10均呈现显著的上调表达。二、采用tail-pcr技术克隆osgrdp1基因1、osgrdp1基因的获得对上述步骤1所获得的突变体spl-d进行遗传分析,结果表明,此突变体的类病斑表型与t-dna插入共分离,为单位点插入的显性突变体。利用tail-pcr技术分离t-dna侧翼序列。分析发现t-dna插入位点在第11号染色体上。在基因组上设计引物p1(5’-ttagcacctctaatcttgcaaa-3’)、p2(5’-ccaaacaagattaacccctga-3’)和t-dna边界引物p3(5’-gatgccgaccggatctgtcgatc-3’)组合进行pcr扩增(图4b),测序分析发现t-dna插入到一个注释为富含甘氨酸结构蛋白编码基因的启动子区域,如图4中a所示,距其翻译起始密码子atg上游495bp,将此基因命名为osgrdp1基因。osgrdp1基因含有8个外显子和7个内含子,其开放阅读框(orf)如序列表中的序列1所示,其编码的蛋白(osgrdp1蛋白)氨基酸序列如序列表中的序列2所示。与野生型水稻爱知旭相比,本发明的突变体spl-d为仅在野生型水稻爱知旭基因组中osgrdp1基因起始密码子atg上游发生t-dna插入突变,t-dna插入位点距osgrdp1基因起始密码子atg上游495bp。2、osgrdp1基因表达量检测利用荧光定量pcr法对突变体spl-d和野生型水稻爱知旭中osgrdp1基因表达进行分析,检测方法以及数据处理同步骤一的4。引物序列如下:qrt-grdp1-f:5’-agcagttgcattgcttgaca-3’;qrt-grdp1-r:5’-gccgtcttcgacattgatgt-3’。结果如图4中c所示,t-dna插入导致osgrdp1基因上调表达3倍以上。实施例2、osgrdp1蛋白的亚细胞定位1、利用pcr方法,以gfp-grdp1-f(5’-ataggtaccatggacggggagcaagag-3’,下划线部分为kpni位点)为正向引物,以gfp-grdp1-r(5’-tatggatccggtgctcgcgttg-3’,下划线部分为bamhi位点)为反向引物,从水稻品种爱知旭的cdna中扩增出osgrdp1基因的开放阅读框序列。pcr产物回收后连入pmd-18t载体(takara)中,经测序正确后,利用kpni和bamhi进行双酶切,酶切产物连入植物亚细胞定位表达载体pcg1301(购自普如汀生物技术(北京)有限公司),即得到含有osgrdp1基因的重组表达载体pcg1301-osgrdp1-gfp。重组表达载体pcg1301-osgrdp1-gfp的结构描述为:在pcg1301载体的多克隆位点kpni和bamhi之间插入序列表中序列1的第1-2970位得到的重组质粒。在重组表达载体pcg1301-osgrdp1-gfp中,驱动osgrdp1基因表达的为35s启动子。2、按照文献“li,x.,2011,infiltrationofnicotianabenthamianaprotocolfortransientexpressionviaagrobacterium.bio-protocolbio101:e95.doi:10.21769/bioprotoc.95)和文献(wang,k.,liu,y.andli,s.,2013,bimolecularfluorescencecomplementation(bifc)protocolforriceprotoplasttransformation.bio-protocol3(22):e979.doi:10.21769/bioprotoc.979.”中的方法,将重组表达载体pcg1301-osgrdp1-gfp分别转化烟草叶肉细胞和水稻原生质体,在激光共聚焦显微镜下观察osgrdp1蛋白的定位情况。结果如图5所示,从图中可以看出osgrdp1蛋白在烟草叶肉细胞和水稻原生质体中均定位于细胞膜上。实施例3、osgrdp1基因超表达植株的获得及表型鉴定本实施例中所涉及的基因为实施例1获得的水稻品种爱知旭中的osgrdp1基因,其核苷酸序列为序列表中的序列1,编码序列表中序列2所示的蛋白(osgrdp1)。序列1由2973个核苷酸组成,序列2由930氨基酸组成。一、osgrdp1基因超表达植株的获得及鉴定1、重组表达载体pcambia1301-ubi-osgrdp1的构建(1)利用pcr方法,采用引物5’-atagagctcgtgcagcgtgacccggt-3’(下划线部分为saci位点)和5’-ataggatccaagtaacaccaaacaacagggt-3’(下划线部分为bamhi位点),以双元载体pubigusplus(普如汀生物技术(北京)有限公司)为模板,扩增ubiquitin启动子区域。将扩增产物回收后连接pmd18-tsimple(takara)载体并测序验证。将测序验证正确的质粒用saci和bamhi双酶切,酶切产物回收后连入pcambia1301载体(普如汀生物技术(北京)有限公司)的saci和bamhi双酶切位点,得到pcambia1301-ubi。(2)利用pcr方法,采用引物oe-grdp1-f(5’-ataggatccatggacggggagcaagag-3’,下划线部分为bamhi位点)和oe-grdp1-r(5’-tatacgcgttcaggtgctcgcgttg-3’,下划线部分为mlui位点),从水稻品种爱知旭的cdna中扩增出osgrdp1基因的开放阅读框序列。pcr产物回收后连入pmd-18t载体(takara)中,经测序正确后,利用bamhi和mlui进行双酶切,酶切产物连入步骤(1)中的植物表达载体pcambia1301-ubi,即得到含有osgrdp1基因的重组表达载体pcambia1301-ubi-osgrdp1。所述重组表达载体pcambia1301-ubi-osgrdp1为将ubiquitin启动子和osgrdp1基因插入pcambia1301载体的saci和mlui酶切位点间后得到的载体。在重组表达载体pcambia1301-ubi-osgrdp1中,驱动所述osgrdp1基因表达的为ubiquitin启动子。在重组表达载体pcambia1301-ubi-osgrdp1的构建过程中,也可以人工合成序列表中序列1所示的osgrdp1基因作为模板。2、osgrdp1基因超表达水稻的获得将上述步骤1所构建的重组表达载体pcambia1301-ubi-osgrdp1,通过农杆菌eha105(普如汀生物技术(北京)有限公司)导入到水稻品种爱知旭(oryzasatival.cv.aichiasahi)的胚性愈伤组织中。具体转化方法参见文献“易自力、曹守云、王力、储成才、李祥、何锶洁、唐祚舜、周朴华、田文忠,提高农杆菌转化水稻频率的研究,遗传学报,2001,28(4):352-358”一文。3、osgrdp1基因超表达水稻的鉴定(1)pcr初步鉴定从步骤2中所获得的t0代转入pcambia1301-ubi-osgrdp1的转基因水稻中提取基因组dna,利用引物5’-gctgcgccgatggtttctacaa-3’和5’-cacggcctccagaagaagatgttg-3’检测转基因水稻中的新霉素磷酸转移酶基因(hptii)片段,pcr扩增产物为514bp的片段的即为转基因阳性植株。经上述pcr鉴定,将其中4个转入pcambia1301-ubi-osgrdp1的转基因水稻株系分别记做t0代转osgrdp1水稻株系oe-1、oe-2、oe-3和oe-4。(2)转录水平分析(rna表达量)以上述获得的t0代转osgrdp1水稻株系oe-1、oe-2、oe-3、oe-4和野生型水稻爱知旭为材料进行荧光定量pcr分析,扩增osgrdp1基因的引物为实施例1中qrt-grdp1-f和qrt-grdp1-r;以actin1基因为内参,扩增引物为表1中的qrt-osrac1-f和qrt-osrac1-r。检测方法以及数据处理同实施例1步骤一中的4。各试验材料中osgrdp1基因的实时荧光定量pcr结果如图6中a所示。从图中可以看出:与未转基因的野生型水稻爱知旭相比,本发明获得的t0代转osgrdp1水稻株系oe-1、oe-2、oe-3和oe-4中osgrdp1基因的转录水平均显著提高。二、osgrdp1基因超表达水稻类病斑表型鉴定以上述获得的t0代转osgrdp1水稻株系oe-1、oe-2、oe-3、oe-4和野生型水稻爱知旭为材料,按常规方法在温室种植,在短日照条件下(日照时间8小时)观察各材料四叶一心期的表型。结果如图6中b所示,t0代转osgrdp1水稻株系oe-1、oe-2、oe-3、oe-4均不同程度产生类病斑,而非转基因的野生型水稻爱知旭则不产生类病斑。且t0代转osgrdp1水稻株系oe-1、oe-2、oe-3和oe-4产生类病斑的严重度与其中osgrdp1基因的表达量呈正相关。这一结果进一步说明,超表达osgrdp1基因可导致产生类病斑及提高转基因水稻的抗病性。序列表<110>中国农业大学<120>与植物抗病性相关的水稻osgrdp1蛋白及其编码基因与应用<160>2<170>patentinversion3.5<210>1<211>2793<212>dna<213>人工序列(artificialsequence)<400>1atggacggggagcaagaggcgcggtggctggcggcgcagggggtcgccgtaggtgccgac60atggtcgcggcggcgctccggcagctggagttcctcgcggcggtggaccgccgccggtgg120ctctacgaggggccgctgctcgagagggccatccacaggtacaaatcatgctggcttcct180ctcctttccaagcacactcaggctgctgttgtagatgggcctttagttgttcctcttgat240tgcgaatggatatggcactgccatcggcttaacccggtacaatacttaaaggactgcaag300agattgtatggcagaatactcgacaatagcaatgttcagtcttcgattagagcagaatcc360aagcaccagtctgagaaggtttgggctgagcaatatcctaaggagccctttgagctggag420tacacaagctcttctgacaactcaatttatgcaaatgctggagctgcagaggacatttct480tatgatttggtcgcagctgttaagagacagtcttctttcttctaccaggttgacacacca540actatgcatgatcagcgatttctggaagaagctttagctcggtacaaaggattcttgtac600ctgatcaagacgaatcaggagaacaaaatgaagctctttcgcgtgccaacctatgatgtg660gatgtcatctggcacactcaccaactgcatcctgctacctactgccatgatatgctgaag720ctcattggacgagttctggagcatgatgacaccgatgacgatcgatcagaaggaaagaag780ctcgatactggattttcagggactaccaaacagtttgagaatgcctttggtgcaagatac840tggaaggctggtgctatgtaccgtggcaacttgccgtctcctgtgacatccaatcctcag900atgtttattagtgaggtggatggtgaatttagtgttggcaaagctgagtcgcaaattacc960attcttgaaacaacagttatagagttattcttgcaaattgtggacatcaagaacttgcca1020cctgcaattccaaaggaaaatgtgtacatatggttcacaaagaatcaaccagacatgttc1080atcagtgatggtggcaggctggatatctctacaaaaacagggaagtcgattggagccagt1140atccaatgtgagcctactggtgaactcattctaacagtactggttgatcgggaatcatca1200tcgaagaaacccaaaaagattgggaagatttctatccctcttcaagagtttacatggtct1260gattccaagctctcctttgagaggtggtttgaattgaaacctcatgatgggcatgccagt1320tcacctatcgtcagccttcgagttgctgcctcttccactgtcccagtgaaggctcagcag1380gttcttagcatgatcaggacagaacctttctctcttaagagttttctgtcaccgaattcc1440atcaaggatcaaaagatgagctgctggactcactttgtgtatgattgcaatactgaactt1500atccgtcttcaaattagggatcagaaggcaaagaacggcatggttgtcgctcgggagttg1560gtcggagtaacaaagtcatcaaagaaaccatttaagctagcagagttcgtggacaacaag1620tggtcactgagcaactctaacttatgcatcactaatgacatgaaaccaagcaaggatggc1680tctatacttgagctcaaatgtgacaacaaaacgatcaaactctaccagggaaagaggcta1740gaatttcaacggaagtgctgcaacaatcacgcagaggagaatgcttcagctattactgct1800gtgaagttctctgctgaacatccatatggcaaagcagttgcattgcttgacaccaaatct1860gagctgataatggtaaatgaggattggtttctgcttccttggatcgtgatgtcgttcctg1920ttccaagacatcaatgtcgaagacggctcgaagctcatcggtggcgccattgctcagaaa1980ggcgcgatctctgagcccgacactgctgccatggcgatgtcagccgagacggtggctgct2040cctgctaactgcggcacttgcggcacggcgtgcgacagtaacatggcaggcgacaaggtg2100gtggccgccgcgagatgcaaggccgcgcgatgcaaggctgtcgcggcaagcggcggcggc2160ggcggccaggcagagtcggctgggtgcggctcgggatgcggcggcggctgtggcggcggt2220gttgccaaggtcgttgaggccaccaaggctggcgtcggcggccatggcaagtccggtgga2280tgtggctcgggctgcggtggtggctgtggcggcggcgggtgcggcgccatggccgtcgag2340agctccaaggatgatgtccatgcgaagtgtgcaggttgtggctcaggctgcggcggcggc2400tgtggcggcggctgtggcggtggcatggtcatggaagcctccaaggctggccatgtcaag2460tccggcgggtgcggctcaggctgcggcggcggctgtggtggaggctgcggtggcggcgtg2520gccatggaaagctccaaggctggccatgtcaagtccggcggatgcggctcaggctgcggc2580ggcggctgtggtggcggttgtggcggcggcatggtcatggaaagctccaaggctgtccat2640gtcaagtccggcggctgcggcggaggctgtggtggcggcgtggccatggaaagctccacc2700gttggacatgccaagtccggtggctgcggttcaggctgtggcggcggctgcggtggcggc2760ggttgcggcgcgatgctcaacgcgagcacctga2793<210>2<211>930<212>prt<213>人工序列(artificialsequence)<400>2metaspglygluglnglualaargtrpleualaalaglnglyvalala151015valglyalaaspmetvalalaalaalaleuargglnleuglupheleu202530alaalavalaspargargargtrpleutyrgluglyproleuleuglu354045argalailehisargtyrlyssercystrpleuproleuleuserlys505560histhrglnalaalavalvalaspglyproleuvalvalproleuasp65707580cysglutrpiletrphiscyshisargleuasnprovalglntyrleu859095lysaspcyslysargleutyrglyargileleuaspasnserasnval100105110glnserserileargalagluserlyshisglnserglulysvaltrp115120125alagluglntyrprolysgluprophegluleuglutyrthrserser130135140seraspasnseriletyralaasnalaglyalaalagluaspileser145150155160tyraspleuvalalaalavallysargglnserserphephetyrgln165170175valaspthrprothrmethisaspglnargpheleugluglualaleu180185190alaargtyrlysglypheleutyrleuilelysthrasnglngluasn195200205lysmetlysleupheargvalprothrtyraspvalaspvaliletrp210215220histhrhisglnleuhisproalathrtyrcyshisaspmetleulys225230235240leuileglyargvalleugluhisaspaspthraspaspaspargser245250255gluglylyslysleuaspthrglypheserglythrthrlysglnphe260265270gluasnalapheglyalaargtyrtrplysalaglyalamettyrarg275280285glyasnleuproserprovalthrserasnproglnmetpheileser290295300gluvalaspglyglupheservalglylysalagluserglnilethr305310315320ileleugluthrthrvalilegluleupheleuglnilevalaspile325330335lysasnleuproproalaileprolysgluasnvaltyriletrpphe340345350thrlysasnglnproaspmetpheileseraspglyglyargleuasp355360365ileserthrlysthrglylysserileglyalaserileglncysglu370375380prothrglygluleuileleuthrvalleuvalasparggluserser385390395400serlyslysprolyslysileglylysileserileproleuglnglu405410415phethrtrpseraspserlysleuserphegluargtrpphegluleu420425430lysprohisaspglyhisalaserserproilevalserleuargval435440445alaalaserserthrvalprovallysalaglnglnvalleusermet450455460ileargthrglupropheserleulysserpheleuserproasnser465470475480ilelysaspglnlysmetsercystrpthrhisphevaltyraspcys485490495asnthrgluleuileargleuglnileargaspglnlysalalysasn500505510glymetvalvalalaarggluleuvalglyvalthrlysserserlys515520525lysprophelysleualagluphevalaspasnlystrpserleuser530535540asnserasnleucysilethrasnaspmetlysproserlysaspgly545550555560serileleugluleulyscysaspasnlysthrilelysleutyrgln565570575glylysargleuglupheglnarglyscyscysasnasnhisalaglu580585590gluasnalaseralailethralavallyspheseralagluhispro595600605tyrglylysalavalalaleuleuaspthrlyssergluleuilemet610615620valasngluasptrppheleuleuprotrpilevalmetserpheleu625630635640pheglnaspileasnvalgluaspglyserlysleuileglyglyala645650655ilealaglnlysglyalailesergluproaspthralaalametala660665670metseralagluthrvalalaalaproalaasncysglythrcysgly675680685thralacysaspserasnmetalaglyasplysvalvalalaalaala690695700argcyslysalaalaargcyslysalavalalaalaserglyglygly705710715720glyglyglnalagluseralaglycysglyserglycysglyglygly725730735cysglyglyglyvalalalysvalvalglualathrlysalaglyval740745750glyglyhisglylysserglyglycysglyserglycysglyglygly755760765cysglyglyglyglycysglyalametalavalgluserserlysasp770775780aspvalhisalalyscysalaglycysglyserglycysglyglygly785790795800cysglyglyglycysglyglyglymetvalmetglualaserlysala805810815glyhisvallysserglyglycysglyserglycysglyglyglycys820825830glyglyglycysglyglyglyvalalametgluserserlysalagly835840845hisvallysserglyglycysglyserglycysglyglyglycysgly850855860glyglycysglyglyglymetvalmetgluserserlysalavalhis865870875880vallysserglyglycysglyglyglycysglyglyglyvalalamet885890895gluserserthrvalglyhisalalysserglyglycysglysergly900905910cysglyglyglycysglyglyglyglycysglyalametleuasnala915920925serthr930当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1