包括SNP3的分子标记及其在湖羊辅助育种中的应用的制作方法
本发明属于分子标记领域,具体地涉及一种湖羊snp分子标记、相关引物对、试剂盒,以及它们在湖羊分子标记辅助育种中的应用,进一步涉及一种筛选湖羊体重性状的方法及其在湖羊分子标记辅助育种中的应用。
背景技术:
自20世纪中叶以来,我国绵羊遗传改良经历了地方品种的选优培育和提纯,杂交改良地方品种,再到新品种培育,生产方向也从“毛主肉辅”向“肉主毛辅”转变。目前国内畜牧养殖业中养羊业的发展速度仅次于家禽养殖[1]。
我国虽然开展了多项肉用绵羊新品种的培育研究工作,并培育了巴美肉羊[2]、昭乌达肉羊[3]和察哈尔羊[4]等具有自主知识产权的肉羊品种。但在育种工作中还是存在“重引进,轻选育”的现象[1]。以湖羊、小尾寒羊为例,这两个品种是我国著名的多胎绵羊品种,但是这些品种毛用价值不高,产肉性能偏低,不能满足羊肉消费市场的需求。因此,进行遗传改良,提高养羊的生产效率是绵羊选育工作中亟待解决的问题,也是摆在广大动物遗传育种研究者面前的一项刻不容缓的任务。
在人类单倍体型图计划、多物种基因组信息的陆续公布,商业化中高密度snp芯片的出现和基因数据分析方法的成熟的大背景下,基于全基因组范围内搜寻与动物重要经济性状相关联的遗传变异研究,为分子育种研究带来了新思路和新途径,而全基因组关联分析(genome-wideassociationstudy,gwas)已成为复杂性状功能基因鉴定的分析方法和手段之一。人们已经在与绵、山羊经济性状上开展了很多gwas的研究[5-29]。ovinesnp50beadchip芯片由illumina公司与国际羊基因组协会专家联合开发,包含54241个snp位点,平均每46kb有一个标记,覆盖整个绵羊基因组。该芯片已经成为绵、山羊经济性状gwas分析的重要工具,目前已有的研究结果大都使用该芯片获得。
gwas在羊的相关性状基因的研究中主要分为两类:(1)无关个体基础上的联系。(2)家系基础上的联系剖析。病例-对照分析的方法主要用来分析病例组和对照组的全基因组中基因型的分布特点和差异性。随机群体基础上的分析法在动物的应用中主要用来分析数量性状。目前各国研究者已经对复杂疾病和经济性状开展了许多的gwas研究分析,发现了许多与疾病或经济性状相关联的snps,已经证实的相关基因或位点就超过一千余个,而且近年来公开报道gwas研究成果的数量呈逐年递增态势[30]。
虽然gwas分析在相关的研究中取得了优异的成绩和效果,也有学者对其持质疑的态度[31],有关研究和分析也在不断的修缮过程中[32-34]。目前对于gwas研究和分析我们还需要从多方面入手,将科研转化到实际中去。
随着生活水平的提高,人们对羊肉的消费需求也越来越多,国内一些地方肉用绵羊品种如湖羊、小尾寒羊等,因繁殖力强,肉质鲜美等优点深受消费者的喜爱,但与肉用品种相比,这些品种具有产肉量较低,生产周期长等,不能满足消费者的需求,湖羊为我国特有的宝贵绵羊品种,集繁殖力强,早期生长快,肉品质好等优点于一体,但也存在产肉性能不高,肉用体型不理想的缺陷。因此,进行湖羊肉用系选育,提高湖羊的生产性能,是湖羊选育工作中亟待解决的重要问题。
技术实现要素:
为了解决现有技术中的技术欠缺和生活中的现实需求,本发明以湖羊为研究对象,运用ovinesnp50genotypingbeadchip芯片在湖羊肉用新类群核心群的亲代(g1)和子代(g2)群体进行基因分型,对肉用核心群体长、体高、胸围、尾长、尾宽等体尺性状进行全基因组关联分析,得到与体高显著相关的位点11个,与胸围显著相关的位点1个,并对这些位点进行湖羊肉用系核心群g3代个体的snp群体验证,一般线性模型分析结果显示,所有snps均不影响湖羊肉用系核心群g3代个体的体尺性状,而与g3代个体的体重性状显著相关。
本发明是首次对湖羊体尺性状的gwas研究,利用gwas技术筛选湖羊体尺性状候选功能基因及snps,可定位绵羊体尺性状候选基因,为探索绵羊体尺性状功能基因提供重要的理论依据和参考。通过snps的群体验证,获得了可用于分子标记辅助育种的snps,它们可用于湖羊肉用性状分子标记辅助育种和湖羊肉用系核心群个体的早期选育。
本发明的目的是提供与湖羊体重性状相关的snp分子标记及其在筛选湖羊体重性状或湖羊分子标记辅助育种中的应用。在一个实施方案中,所述分子标记包括位点位于seqidno:1的第209bp位点的snp3,其为g或a。在一个实施方案中,所述分子标记包括位点位于seqidno:2的第129bp位点的snp5,其为t或c。在一个实施方案中,所述分子标记包括位点位于seqidno:3的第303bp位点的snp7-1,其为t或c。在一个实施方案中,所述分子标记包括位点位于seqidno:3的第373bp位点的snp7-2,其为t或c。在一个实施方案中,所述分子标记包括位点位于seqidno:4的第87bp位点的snp10-1,其为g或a。在一个实施方案中,所述分子标记包括位点位于seqidno:4的第207bp位点的snp10-2,其为t或c。在一个实施方案中,所述分子标记包括位点位于seqidno:4的第211bp位点的snp10-3,其为c或a。
本发明的另一目的是提供与湖羊体重性状相关的snp分子标记及其在筛选湖羊体重性状或湖羊分子标记辅助育种中的应用。在一个实施方案中,所述分子标记的序列如seqidno.1所示,其中第209bp(基因定位为oar6_95218086.1上游44bp)位点的碱基为g或a。在另一个实施方案中,所述分子标记的序列如seqidno.2所示,其中第129bp(基因定位为s10476.1下游16bp)位点的碱基为t或c。在另一个实施方案中,所述分子标记的序列如seqidno.3所示,其中第303bp(基因定位为oar1_164254640.1下游192bp)位点的碱基为t或c。在另一个实施方案中,所述分子标记的序列如seqidno.3所示,其中第373bp(基因定位为oar1_16425464下游235bp)位点的碱基为t或c。在另一个实施方案中,所述分子标记的序列如seqidno.3所示,其中第303bp(基因定位为oar1_164254640.1下游192bp)位点的碱基为t或c;和第373bp(基因定位为oar1_16425464下游235bp)位点的碱基为t或c。在另一个实施方案中,所述分子标记的序列如seqidno.4所示,其中第87bp(基因定位为oar6_90337552.1上游41bp)位点的碱基为g或a。在另一个实施方案中,所述分子标记的序列如seqidno.4所示,其中第207bp(基因定位为oar6_90337552.1下游79bp)位点的碱基为t或c。在另一个实施方案中,所述分子标记的序列如seqidno.4所示,其中第211bp(基因定位为oar6_90337552.1下游83bp)位点的碱基为c或a。
本发明另一目的是提供一种检测与湖羊体重性状相关的上述snp3、snp5、snp7-1、snp7-2、snp10-1、snp10-2、snp10-3的引物对、包含该引物对的试剂盒以及它们在筛选湖羊体重性状或湖羊分子标记辅助育种中的应用。在一个实施方案中,所述引物对的核苷酸序列见表3-1。在一个实施方案中,所述引物对为表3-1中的3f+3r,即seqidno.5和seqidno.6。在另一个实施方案中,所述引物对为表3-1中的5f+5r,即seqidno.7和seqidno.8。在另一个实施方案中,所述引物对为表3-1中的7f+7r,即seqidno.9和seqidno.10。在另一个实施方案中,所述引物对为表3-1中的10f+10r,即seqidno.11和seqidno.12。
本发明的另一目的是提供上述snp分子标记、引物对或试剂盒在筛选湖羊体重性状或湖羊分子标记辅助育种中的应用。
本发明的另一目的是提供一种筛选湖羊体重性状的方法,包括如下步骤:提取湖羊基因组dna,利用上述引物对进行pcr扩增,对扩增产物中的上述snp3、snp5、snp7-1、snp7-2、snp10-1、snp10-2、snp10-3进行检测,从而筛选湖羊体重性状。在一个实施方案中,所述引物对的核苷酸序列见表3-1。在一个实施方案中,所述引物对为表3-1中的3f+3r,即seqidno.5和seqidno.6。在另一个实施方案中,所述引物对为表3-1中的5f+5r,即seqidno.7和seqidno.8。在另一个实施方案中,所述引物对为表3-1中的7f+7r,即seqidno.9和seqidno.10。在另一个实施方案中,所述引物对为表3-1中的10f+10r,即seqidno.11和seqidno.12。
在上述任一筛选湖羊体重性状的方法的具体实施方案中,其中pcr扩增的反应程序为:95℃预变性10min;95℃变性30s,53℃退火30s,72℃延伸30s,35个循环;72℃延伸10min;pcr扩增的反应体系如下:
本发明的另一目的是提供上述筛选湖羊体重性状方法在湖羊分子标记辅助育种中的应用。
本发明的有益效果在于:所述分子标记、引物对及相关试剂盒可以用于筛选湖羊体重性状或湖羊分子标记辅助育种,用于湖羊肉用系核心群个体的早期选育。
附图说明
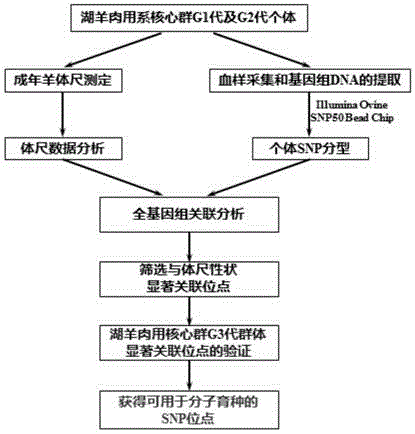
图1为获得可用于分子育种的snp位点的技术路线。
图2-1为湖羊基因组dna琼脂糖凝胶电泳检测结果(marker:dl2000plus)。
图2-2为glm体高全基因组关联分析的曼哈顿图(manhattanplots)。
图2-3为glm胸围全基因组关联分析的曼哈顿图(manhattanplots)。
图2-4为mlm体高全基因组关联分析的曼哈顿图(manhattanplots)。
图2-5为mlm胸围全基因组关联分析的曼哈顿图(manhattanplots)。
图2-6为湖羊体高q-qplot图(a:glm;b:mlm)。
图2-7为湖羊胸围q-qplot图(a:glm;b:mlm)。
图3为湖羊体高性状相关snps检测引物的pcr扩增结果。m:dl2000plus;泳道1~8分别对应针对oar6_95218086.1,oar15_18440393.1,s10476.1,oarx_120998827.1,oar1_164254640.1,s10347.1,s11279.1,oar6_90337552.1设计的扩增产物。
具体实施方式
下面通过实施例对本发明做进一步说明。
实施例1湖羊体尺性状的全基因组关联分析
ovinesnp50genotypingbeadchip芯片是通过illumina的iselect项目与国际羊基因组协会合作开发,整合了多个羊品种基因差异,平均每46kb有1个标记,提供足够的snp密度覆盖整个基因组,可应用于全基因组关联研究中。本发明人从2006年开始,在湖州和萧山分别组建了湖羊肉用新类群核心群,经过选育,核心群的亲代和子代群体在体长、体高、胸围、尾长、尾宽等体尺指标上表现出了显著差异。我们利用ovinesnp50genotypingbeadchip芯片在湖羊肉用新类群核心群的亲代和子代群体进行基因分型,分别使用一般线性模型(generallinearmodel,glm)和混合线性模型(mixedlinearmodel,mlm)对湖州太湖湖羊养殖专业合作社构建的肉用核心群体长、体高、胸围、尾长、尾宽等体尺性状进行全基因组关联分析(genomewideassociationstudies,gwas),运用最新绵羊基因组ovis_aries_v3.1序列信息和gwas方法进行的绵羊体尺性状、肉用性状主效基因的鉴定研究。该研究结果可定位绵羊体尺性状候选基因,也可为探索绵羊体尺性状功能基因提供重要的理论依据和参考。
2.1试验材料
2.1.1试验动物
本试验选用240只湖羊均来自湖州太湖湖羊养殖专业合作社湖羊肉用系核心群,饲养管理方式依据肉羊标准的饲养管理方法进行饲养。
2.1.2样品采集和处理
每只试验羊采集颈静脉血10ml,于含edta抗凝剂的采血管中暂存,并置于-20℃长期保存。
2.1.3主要试剂
蛋白酶k:美国amresco公司
nacl、无水乙醇、琼脂糖、冰乙酸、硼酸、naoh:北京化学试剂公司
dnamarker15000:杭州擎科梓熙生物有限公司
dna芯片杂交试剂盒:illumina公司
2.1.4主要溶液
以萨姆布鲁克所著的《分子克隆实验指南》第三版为参考,配制所需溶液,溶剂为超纯水,蒸汽高温高压灭菌40min。
(1)1mol/l的tris-hcl(ph=8.0,1l):121.1gtris-base溶于800ml超纯水后,以浓盐酸将溶液ph值调至8.0,定容至1l,高压灭菌。
(2)5×tbe(tris硼酸缓冲液):称取54gtris-base,27.5g硼酸,3.72gna2edta·2h2o,加超纯水定容至1l,混匀。
(3)0.5medta(ph=8.0):将186.1gna2edta·2h2o溶解于800ml双蒸水中,将ph调至8.0,定容至1000ml,高温高压灭菌。
(4)0.5mnacl:5.844gnaci溶于超纯水,并定容至200ml,高温高压灭菌。
(5)10%sds:将10gsds溶于65℃的超纯水,定容至100ml,用0.2nm的滤膜滤菌后保存。
(6)te缓冲液(ph=8.0)含20mmtris·hcl(ph=8.0),1mmedta(ph=8.0):2ml1mtris·hcl(ph=8.0),0.2ml0.5medta(ph=8.0),定容至100ml,高压灭菌。
(7)蛋白酶k(20mg/ml):将100mg蛋白酶k溶解于5ml的超纯水中,并按照每管400μl分装后-20℃冻存。
(8)3mnaac(ph=5.2):取12.305g无水naac加双蒸水溶解,定容至50ml,加冰乙酸调节ph至5.2,高压灭菌。
(9)pbs:8.0gnacl,0.2gkcl,3.48gna2hpo4·12h2o,0.2gkh2po4,定容至200ml,高压灭菌。
(10)ste:5ml1mtris·hcl(ph=8.0),20ml的0.5medta(ph=8.0),20ml的0.5mnacl,10ml的10%sds,加入ddh2o双蒸水至100ml,混匀。
2.1.5主要仪器设备
恒温水浴锅:江苏太仓实验设备公司
thermofisher-80℃超低温冰箱:美国
海尔4℃/-20℃冰箱:山东
centrifuge58108高速低温冷冻离心机:德国eppendorf公司
f100icematic制冰机:意大利
bio-radchemidoc凝胶成像分析系统:美国
tomyes-315型高压蒸汽灭菌锅:日本
sartorius电子天平:德国
dyy-7c型电泳仪:北京六一
dyy-iii32型电泳槽:北京六一
ql-901涡旋振荡器:江苏
dshz-300多用途水浴恒温振荡器:江苏太仓实验设备厂
astrageneastranet紫外分光光度计:英国
10μl、100μl、200μl、1000μl移液器:德国eppendorf公司
infinium全基因组snp分析系统:illumina公司
2.1.6数据分析软件及在线软件
2.1.6.1数据分析处理软件
1.tassel3.0
tassel的主要功能是研究表型与基因型之间的关系,是使用人口和数量遗传学工具评估基因型和性状关联的java软件,具有最新和统计上最强大的复杂种群关联作图方法,包括一般线性模型(generallinearmodel,glm)和混合线性模型(mixedlinearmodel,mlm)。本研究中使用tassel软件包中的glm和mlm对湖羊体高、体(斜)长、胸围、尾长和尾宽等5个体尺性状进行全基因组关联分析。
2.plink1.09
gwas分析软件,可对基因型或表型数据、样本进行质量管理控制,对snps进行分析处理,估算基因型和表型值。
3.r语言
用于全基因组关联分析结果中曼哈顿图及q-qplot图的绘制。
4.mutationsurveyorversion5.02
测序峰图分析和序列组装软件。
2.1.6.2在线网站及数据库
(1)ucsc在线网站:http://genome.ucsc.edu/。
(2)ncbi在线网站:http://www.ncbi.nlm.nih.gov。
(3)david在线网站:http://david.abcc.ncifcrf.gov。
2.2试验方法
2.2.1体尺性状的测定
湖羊肉用系核心群个体的体尺测量包括体(斜)长、体高、尾长、尾宽和胸围。测量使用卷尺等工具,保证羊处于水平地面,安静,放松站立。测量人员尽量保证为同一人,以减少人为原因引起的测量误差,每只羊至少测量2次,取平均值作为最终的测量结果。具体测定方法如下:
体长:由肩脚骨前端到坐骨结节后端的直线距离;
体高:由髻甲最高点到地面的垂直距离;
胸围:沿肩胛骨后缘绕胸一周的长度;
尾长:由尾根到尾端的距离;
尾宽:尾幅最宽处的距离。
2.2.2技术路线
技术路线参见图1。
2.2.3外周血基因组dna的提取
-20℃冻存的外周血解冻后,吸取0.5ml到一个新的1.5mlep管中,以tris饱和酚法进行血液基因组dna的提取。
利用琼脂糖凝胶电泳检测基因组dna的完整性。
取基因组dna溶液2μl,测定dna的浓度和纯化效果,保证所提取和纯化的dna浓度在100ng/μl以上,od值260/280处于1.8~2.0。
2.2.4湖羊肉用系核心群g1代和g2代个体snps的基因分型
使用illumina公司与国际羊基因组协会专家联合开发的ovinesnp50beadchip芯片进行个体snps的基因分型。
2.2.4.1基因分型步骤
(1)样本标准化:将dna浓度稀释成50ng/μl。
(2)dna的扩增:向样品中加入浓度为1n的naoh溶液,再加入基因组扩增所需的试剂,然后室温放置12h。
(3)dna的片段化:使用酶制剂将dna变成片段。
(4)沉淀dna:使用无水异丙醇将dna沉积在管壁。
(5)重悬dna:室温干燥后加入相应的缓冲溶剂使其完全溶解。
(6)dna与芯片的杂交:步骤(5)得到的dna样品与芯片杂交后,置于杂交炉中充分反应。
(7)芯片的延伸和染色:用清洗液洗去未结合和非特异性结合的dna。
(8)包被芯片:将上述的芯片置于xc4试剂中,涂上包被液后,在真空条件下放置1h。
(9)芯片扫描:将步骤(8)处理好的芯片放到扫描仪中进行芯片扫描。
2.2.4.2基因型数据质量控制管理
芯片处理后,把数据输入到beadstudio软件中进行相关的分析。软件输出的基因型经整理和校对后,进行统计分析,使用软件剔除或矫正不精确的snp位点,获得全基因组所有数据的特征。
2.2.4.3snp位点分型和质量管理
利用全基因组基因型分型系统对所有样本的全基因组核酸进行分型研究,然后使用genomestudio软件将数据变成可视化的基因分型结果,保存为txt格式,并输出。
2.2.5数据处理
2.2.5.1表型数据统计分析
利用spss20统计软件,对测定的肉用系核心群体尺指标进行统计分析,计算每个体尺指标的平均值和标准差等。
2.2.5.2gwas处理
基因型数据经质控处理后,使用tassel3.0软件的一般线性模型(glm)和混合线性模型(mlm)进行snp的gwas分析,挖掘与湖羊肉用系核心群体尺性状表型相关的snps。
glm模型校正了性别、群体结构2个混杂因素。由于分析的所有个体均为来自同一饲养场相同饲养环境和管理条件的羊只,因此数据建模时不包括场效应。
具体模型为:y=xβ+e
其中,
y:湖羊肉用系核心群体尺性状、体重性状表型值向量;
β:表型均值、snp、群体结构、性别等固定效应向量;
e:残差效应向量;
x为β的关联矩阵。
mlm模型校正了性别、群体结构和亲缘关系3个混杂因素。
具体模型为:y=xβ+sα+qv+zu+e
其中:
y:湖羊肉用系核心群体尺性状表型值向量;
β:除snp和群体结构之外的固定效应向量;
α:snp效应向量;
v:群体结构效应向量;
u:多基因背景效应向量;
e:残差效应向量;
x、s、q、z分别为β、α、v、u的关联矩阵。
2.2.5.3多重假设检验
进行湖羊肉用系核心群体尺性状关联分析时,多重假设检验如出现错误,需要对p值进行处理分析和扶正。分别采用glm和mlm分析计算出f值和p值,然后进行检验,具体公式如下:
若位点的p值小于α,我们就认为该snp位点与体尺性状具有显著关联。
2.2.5.4群体分层
进行湖羊肉用系核心群体尺性状关联分析时,群体分层和假阳性影响较大。通过对湖羊肉用系核心群体高、体长、胸围、尾长和尾宽性状绘制q-qplot图,判别是否出现偏差和样本群体的分层现象。
2.2.5.5候选基因的注释及挖掘
全基因组关联分析获得显著性snps位点后,下载显著关联snp位点上、下游各500bp的碱基序列,并与ncbi和ovisaries_v4.0(ucsc)等数据库进行序列的blast,以确定snp的定位信息和相邻基因信息。
2.3数据分析结果
2.3.1湖羊肉用系核心群g1代和g2代个体体尺性状的t检验
进行数据gwas分析前,首先对湖羊肉用系核心群g1代(n=161)和g2代(n=79)个体的体尺性状进行了t检验。
2-1湖羊核心群g1代和g2代个体体尺性状的t检验
注:同一行不同小写字母表示差异显著(p<0.05),不同大写字母表示差异极显著(p<0.01),相同字母表示差异不显著。
结果显示,g1代和g2代个体在体高、体长、胸围、尾长和尾宽这5个体尺性状上都存在极显著差异,经过选育后湖羊肉用系核心群g2代个体的体高、体长、胸围、尾长和尾宽均极显著高于g1代个体(p<0.01)(表2-1)。
2.3.2湖羊肉用系核心群6月龄重与体尺性状的相关分析
湖羊肉用系核心群g1代和g2代个体生产记录包括6月龄重,对湖羊肉用系核心群g1代(n=161)和g2代(n=79)合计240个个体的体高、体长、胸围、尾长和尾宽这5个体尺性状和6月龄重进行相关分析,结果如表2-2所示,湖羊核心群个体的5个体尺性状和6月龄重呈现极显著的正相关,与6月龄重相关系数最低的体尺性状为尾宽(r=0.640,p<0.01),与6月龄重相关系数最高的体尺性状为胸围(r=0.893,p<0.01)。另外,各体尺性状间也存在极显著的正相关,其中体高和胸围的相关系数最高(r=0.896,p<0.01),体长和尾宽的相关系数最低(r=0.589,p<0.01)。
2-2湖羊肉用系核心群g1代和g2代个体(湖州)体尺性状和6月龄重的相关分析
注:*表示显著相关(p<0.05);**表示极显著相关(p<0.01)。
2.3.3基因组dna检测
所有个体提取和纯化的血液基因组dna均进行了片段长度、纯度和浓度的检测。基因组dna1%琼脂糖凝胶电泳检测结果如图2-1所示,基因组均达到“条带单一,明亮,无拖尾,同时基因组dnaod值260/280处于1.8~2.0”的标准,可用于snp分型。
检测结果参见图2-1。
2.3.4snp分型及质量管理
检测合格的样本基因组dna通过全基因组分型检测平台进行检测,采用plink1.09软件对采集的240个个体样本和54241个snps位点作了以下的质量控制管理。对于snp位点,我们剔除了:(1)3577个分型成功率小于90%的染色体位点;(2)4435个等位基因频率等于0.05的染色体位点;(3)20个不符合hwe检验的染色体位点。对于个体。我们剔除了12个分型成功率小于90%的个体。
根据以上的质量管理原则,最终筛选出228个个体样本和46209个有效的位点进行gwas分析。
2.3.5染色体间显著水平的确定
为减少多重检验带来的假阳性率,以连锁不平衡修正的bonferroni校正对全基因组关联分析结果p值进行校正。最后估算的ld块和单个独立snp数目为35161,因此bonferroni校正的达5%基因组水平显著的p值阈值为1.42203×10-6(0.05/35161),即p值低于此阈值的snps则认为与表型显著关联;达到基因组水平极显著p值阈值为2.844×10-7(0.01/35161)。2.3.6体尺性状gwas结果
根据“材料与方法”2.2.5中提供的软件和模型,对湖羊肉用系g1代和g2代核心群个体进行gwas分析。
2.3.6.1体尺性状glm的gwas结果
glm分析结果显示,4个snps在基因组水平上与体高达到显著相关,分别为位于6号染色体的oar6_90337552.1、8号染色体的s11279.1、10号染色体的s44173.1和17号染色体的s55179.1;7个snps在基因组水平上与体高达到极显著相关,分别为位于1号染色体的oar1_164254640.1、2号染色体的s10476.11、6号染色体的oar6_95218086.1、9号染色体的s10347.1、15号染色体的oar15_18440393.1、27号染色体的oarx_76354330.1和oarx_120998827.1(表2-3);同时,位于27号染色体的oarx_76354330.1在基因组水平上还与胸围极显著相关(表2-4)。没有snp与体长、尾宽和尾长这3个体尺性状在基因组水平达到显著相关。
表2-3湖羊与体高在基因组水平显著相关的snps(glm分析结果)
注:带灰色底纹的snp为基因组水平极显著相关的snp。下同。
表2-4湖羊与胸围在基因组水平显著相关的snps(glm分析结果)
glm体高和胸围全基因组关联分析的曼哈顿图(manhattanplots)如图2-2、2-3所示,曼哈顿图的x轴是snp位点所在的染色体位置,y轴是snp的p值(-log10),y值越大说明p值越显著。
2.3.6.2体尺性状mlm的gwas结果
mlm分析结果显示,4个snps在基因组水平上与体高达到显著相关,分别为位于1号染色体的oar1_164254640.1、位于9号染色体的s10347.1、位于15号染色体的oar15_18440393.1和27号染色体的oarx_120998827.1;2个snps在基因组水平上与体高达到极显著相关,分别为位于6号染色体的oar6_90337552.1和位于27号染色体的oarx_76354330.1(表2-5);同时,位于27号染色体的oarx_76354330.1在基因组水平上还与胸围显著相关(表2-6)。没有snp在基因组水平与体长、尾宽和尾长这3个体尺性状达到显著相关。
表2-5湖羊与体高在基因组水平显著相关的snps(mlm分析结果)
表2-6湖羊与胸围在基因组水平显著相关的snps(mlm分析结果)
mlm分析结果与glm分析结果不同的是,位于2号染色体的s10476.11、6号染色体的oar6_90337552.1、8号染色体的s11279.1、10号染色体的s44173.1和17号染色体的s55179.1在全基因组水平上与体高均未达到显著性水平。
与glm分析结果相同的是,mlm分析同样得到“27号染色体的oarx_76354330.1在全基因组水平与体高和胸围均呈现显著或极显著相关”;位于“6号染色体的oar6_95218086.1和27号染色体的oarx_120998827.1在基因组水平上与体高达到极显著相关”的结论。
mlm体高和胸围全基因组关联分析的曼哈顿图(manhattanplots)如图2-4、2-5所示。2.3.7群体分层评估
群体分层是指群体内存在别的亚群体从而导致在进行关联分析时有假阳性结果的出现,进而影响结果。
本研究分别采用glm和mlm对检测出有显著关联的体高(图2-6)和胸围(图2-7)性状绘制q-qplot图。
q-qplot图主要用来衡量观测值和预测值之间的差异。图2-6和图2-7的横坐标表示实际观测值对应的分位数结果,纵坐标表示所建模型理论上的预测值对应的分位数结果,两个取值应该近似相等,图中的斜线代表预测线,如果出现偏离的情况说明实际值和预测值有偏差,如果出现较大的偏离则说明是这个snp位点突变所产生的遗传作用造成的。
体高和胸围中glm(图2-6a,图2-7a)和mlm(图2-6b,图2-7b)q-qplot图的实际值基本上都落在预测线上,且图形基本一致,表明glm和mlm的关联分析结果可靠,试验群体经过校正后不存在群体分层的现象。
2.3.8全基因组水平显著关联snps的基因注释
根据“2.2.5.5候选基因的注释及挖掘”中描述的方式,对湖羊体高和胸围gwas全基因组水平显著关联的snp位点进行blast,确认snp位点与绵羊全基因组内的散步处所和主导的功效等信息联系,然后注释。
在全基因组水平,与湖羊体高或胸围显著关联的11个snps位点注释信息见表2-7。这些snps中有5个位于基因内,如oarx_76354330.1位于capn6基因内含子2;oar1_164254640.1位于cadm2基因内含子7;s11279.1位于rnf217基因内含子3;s10347.1位于samd12基因内含子3;oar15_18440393.1位于ddx10基因内含子8。另外,还有6个全基因组水平显著关联的snp位点位于基因间隔区,如,s55179.1位于acads和sppl3之间,acads下游16kb;s10476.1位于slc38a11和cobll1基因之间,在slc38a11下游93kb;oar6_95218086.1位于gc和npffr2之间,在gc下游209kb;oar6_90337552.1位于epha5和loc101120496之间,epha5下游1446kb;s44173.1位于loc101106088和fam124a之间,loc101106088下游25kb;oarx_120998827.1位于loc101103048和prr32基因之间,loc101103048下游270kb;
在分布的染色体上,与湖羊体高或胸围在全基因组水平显著关联的11个snps位点分布在9条染色体上,相应为1号染色体1个位点,2号染色体1个位点,6号染色体2个位点,8号染色体1个位点,9号染色体1个位点,10号染色体1个位点,15号染色体1个位点,17号染色体1个位点,27号染色体2个位点。
表2-7与湖羊体高或胸围显著关联的11个snps上、下游各500bp序列的比对结果(glm)
2.4讨论
2.4.1表型性状的选择
我们对湖羊肉用系核心群个体体高、体长、胸围、尾长和尾宽和6月龄体重的相关分析结果显示,体尺和体重存在显著的正相关,该结果与研究结果和生产实践的情况是吻合的。如,高志英发现多浪羊的体高、胸围等与体重显著相关[35],赵子贵发现体重与体高、胸围、体长等体尺性状存在极显著的线性回归关系[36]。而在养羊生产中,通过测量动物的体尺和体重可以间接预测其以后的生产性能,达到早期选育的目的。
目前已开展了羊的gwas研究多集中于繁殖性状[37,38]、肉质性状[39]、产奶性状[40]、疾病抗性[41]、毛品质[42]、毛色[19,28,43]、角型[44,45]及尾型[46,47]等,与体尺性状相关的gwas分析研究不多,但al-mamun等也在绵羊6号染色体上发现了ncapg和lcorl基因可影响澳大利亚美利奴羊体尺指标[6]。本研究中,我们对湖羊体高、体长、胸围、尾长和尾宽这5个体尺指标进行了gwas分析,这些指标与湖羊的生长性状和品种特点密切相关,该研究结果不仅有助于获得与湖羊体尺性状相关的候选基因,相关的snp还可以用于湖羊肉用系核心群个体的早期选育,对于湖羊肉用性能的提升具有重要意义。
2.4.2群体分层
群体分层会引起gwas分析产生假阳性现象,这种现象虽然会表现为显著相关,但是它与显著相关的性状并没有关系,而是出现了一种假关联[48]。因此群体分层也被认为是影响gwas结果最重要的原因之一[49]。群体分层与假阳性现象交织在一起,会使许多相关性难以分辨,并且还影响群体间的验证[50]。
在之前的研究中,牛和猪的gwas研究中都考究了其对分析结果的影响[51]。从取样群体来看,由于试验羊来自同一个试验场构建的半开放式核心群,理论上应不存群体分层。体高和胸围glm和mlm的q-qplot图的实际值与预测线基本吻合,且glm和mlm的q-qplot图形基本一致。结合glm和mlm分析结果获得的显著关联位点情况看,胸围glm和mlm分析结果完全一致,而体高glm和mlm差异不大,glm仅比mlm多5个显著位点,表明glm和mlm的关联分析结果可靠,试验群体经过校正后不存在群体分层的现象。
2.4.3基因注释
由于我们分析得到的显著的snps位点有11个,散布在9条染色体上。对这11个snps进行基因注释后(基因注释结果主要依据2018年5月11日的ncbi数据库中对oar_v4.0全基因组鸟枪法测序拼接结果),我们获得了一些可能与湖羊的体高和胸围有关的候选基因。
2.4.3.1cadm2(celladhesionmolecule2)
oar1_164254640.1位于cadm2(geneid:101120371)基因内含子7。细胞粘附分子(cadm)由蛋白质家族组成,其功能包括维持细胞极性和肿瘤抑制,包括肝细胞癌(hcc)的几种癌症中可以观察到cadm2基因表达的低表达[52]。cadm2基因在人类的肾细胞癌中被dna启动子甲基化和/或杂合性丢失所抑制。作为一种新型肿瘤抑制剂起作用,并且可能成为人类肾细胞癌的潜在治疗靶点[53]。还在日本人群中使用微卫星的全基因组关联研究(561个病例和561个对照)鉴定了cadm2,其为牛皮癣的候选基因[54]。有学者将cadm2识别为全身能量稳态的有力调节剂,降低cadm2表达可以逆转包括肥胖,胰岛素抵抗和葡萄糖稳态受损在内的多种与代谢综合征相关的症状[55]。
2.4.3.2rnf217(ringfingerprotein217)
s11279.1位于rnf217(geneid:101119141)基因内含子3。人类rnf217编码高度保守的ring指蛋白,主要在具有不同剪接变体的睾丸和骨骼肌中表达。rnf217含有tm结构域的rbr泛素连接酶亚家族成员之一,包含rnf144a和rnf144b,rnf19a/dorfin,rnf19b和rnf217(也称为ibrdc1)的所有5种含跨膜(transmembrane,tm)的rbre3连接酶具有rbr-tm(gxxxg)上层结构。某些人类白血病rnf217的高表达,显示该基因的失调可能与白血病发生有关[56]。另外,人类胃癌、胃腺癌和肝癌中也发现存在rnf217蛋白gxxxg基序的突变[57]。2.4.3.3samd12(sterilealphamotifdomaincontaining12)
s10347.1位于samd12(geneid:101114621)内含子3。samd12是sam结构域(sterilealphamotif,sam)家族成员之一,samd12可能主要通过sam结构域发挥影响雄性不育的功能和作用[58]。研究显示在samd12的内含子4中的tttca和tttta重复的异常扩增与成人的肌阵挛性癫痫有关,在samd12中tttca和tttta的重复估计在2.2~18.4kb的范围内,对应于440~3680个重复单位[59]。
2.4.3.4ddx10(dead-boxhelicase10)
oar15_18440393.1位于ddx10(geneid:101106358)内含子8。ddx10编码rna解旋酶,涉及卵巢癌肝癌、急性髓系白血病[60]等病理过程。
2.4.3.5capn6(calpain-6)
oarx_76354330.1位于capn6(geneid:101110122)内含子2。钙蛋白酶6(capn6)是钙依赖性细胞内非溶酶体蛋白酶之一。capn6是一种具有微管结合和稳定活性的非蛋白分解蛋白酶,可促进破骨细胞中的细胞骨架结构和微管稳定性[61]。capn6作为rac1活性的潜在调节器,通过与rho鸟嘌呤核苷酸交换因子gef-h1相互作用,控制层脂膜形成和细胞运动[62]。胚胎组织中表达的capn6可作为微管稳定蛋白,参与微管动力学和细胞骨架组织的调节[63]。胚胎发生过程中,骨骼和心肌发育过程中可观察到capn6mrna的表达,肺、肾和胎盘的特定细胞,及各种上皮细胞类型中该基因也有表达[64]。
通过基因注释发现,与湖羊体高或胸围在全基因组水平显著关联的11个snps位点分布在9条染色体上,其中1、2、8、9、10、15、17号染色体均有1个位点与湖羊肉用核心群体尺性状显著相关,而6、27号染色体各有2个位点与湖羊肉用核心群体尺性状显著相关。研究人员在该染色体上已经发现了多个与绵羊体重[6]、多胎性[65]、产毛量[66]等生产性状相关的候选基因。绵羊的27号染色体为性染色体,该染色体上的snp大多与绵羊的脂肪沉积、尾脂丰满度和尾型有关[67-69]。已有的研究结果显示,6号和27号染色体对绵羊的生产性状和种质特点具有重要影响,因此,我们在这些染色体上发现的4个snps可能对湖羊肉用系品种选育具有重要意义。
另外,基因注释的结果显示,11个与湖羊体高或胸围在全基因组水平显著关联的snps位点中6个snp定位在基因间隔区,5个已经定位在基因内的snps也分布在基因的内含子上,所以,这些snps是通过何种方式来影响湖羊的体尺性状仍需进一步的研究和分析。
实施例2湖羊体高性状相关snps的群体验证
应用基因组中数以百万计的snps,gwas可以进行全基因组水平上的分子遗传标记对照分析或相关性分析,通过比较发现影响复杂性状的基因变异。gwas研究目前主要采用两阶段或多阶段方法。第一阶段基于覆盖全基因组范围snp芯片进行不同群体的对照分析,经过不同数学模型的统计分析后,获取少量在基因组水平的阳性snp。第二阶段或随后的多阶段中采用更大的样本验证基因分型和数据模型分析的结果。这样的设计需要保证第一阶段中筛选与目标性状相关snp的敏感性和特异性,尽量减少分析的假阳性或假阴性,第二阶段中使用足够大的样本群进行基因分型验证。
随着基因组学研究以及基因芯片技术的发展,通过gwas方法发现并鉴定了大量与复杂性状相关联的遗传变异,并在农业动物重要经济性状主效基因的筛查和鉴定中得到了广泛应用。我们利用ovinesnp50genotypingbeadchip芯片,通过对比分析本发明人组建的湖羊肉用新类群核心群的g1代和g2代群体,发现了11个在基因组水平显著影响湖羊体高和胸围的snps。这些snps能否用于湖羊肉用系核心群个体的早期选育仍需要大群体的反复验证。
目前尚未有湖羊经济性状gwas的报道,我们通过下载11个在基因组水平显著影响湖羊体高和胸围的snps位点上、下游500bp的碱基序列,设计pcr扩增引物,以杭州庞大农业开发有限公司构建的另1个湖羊肉用系核心群g3代个体为研究对象,进行湖羊肉用系核心群g3代203个个体11个snps的基因分型和群体遗传学分析,以及snps与湖羊肉用系核心群体尺性状(体高、胸围)和体重性状的关联分析,11个位点的gwas验证结果可为后续功能基因验证,解释性状变异的分子遗传机制,并为湖羊肉用性状分子标记辅助育种奠定重要基础。
3.1材料及样品的采集
3.1.1动物群体的选择
试验动物均来自杭州庞大农业开发有限公司构建的湖羊肉用系核心群g3代个体。203头湖羊个体进行不同位点的snp基因型分析,并进行对应位点的群体遗传学分析。
3.1.2血样采集及保存
详见实施例1。
3.2主要试剂、溶液的配置和试验仪器设备
详见实施例1。
3.3试验方法
3.3.1表型的描述性统计分析
利用spss20软件对湖羊肉用系核心群g3代个体体高、胸围、初生重、断奶重、6月龄重和成年重表型值进行初步的统计学分析,包括最小值、最大值、平均值和标准差。
3.3.2基因组dna提取及质量控制
详见实施例1。
3.3.3pcr扩增及检测
(1)引物设计与合成
以实施例1中筛选出的11个与湖羊体高和胸围性状显著相关的snps位点为中心,在ucsc数据库中,下载11个snp上、下游各500bp的序列,并在snp上、下游设计扩增引物。其中针对oarx_76354330.1位点设计的2f和2r、针对s44173.1位点设计的11f和11r、针对s55179.1位点设计的12f和12r在预试验中出现较为明显的非特异扩增,且由于序列的特殊性无法进行引物设计的修订,因此仅能验证oar6_95218086.1、oar15_18440393.1、s10476.1、oarx_120998827.1、oar1_164254640.1、s10347.1、s11279.1、oar6_90337552.1、s44173.1和s55179.1共8个位点的基因型情况,各位点引物序列详见表3-1
表3-1引物设计结果
(2)湖羊体高性状相关snp检测的pcr扩增体系
使用30μl的pcr反应体系进行扩增,pcr反应程序为:95℃预变性10min;95℃变性30s,53℃退火30s,72℃延伸30s,35个循环;72℃延伸10min,反应结束后4℃保存。具体反应试剂的使用量见表3-2。
表3-2湖羊体高性状相关snp检测的pcr扩增体系
(3)检测目的片段并测序
取5μl的pcr产物与1μl的6×orangeloadingbuffer均匀混合后点样于1.5%的琼脂糖凝胶,使用dl2000plus为marker标记,200v电压电泳15min左右,eb染色10min后,观察pcr产物有无扩增条带,记录,拍照保存。每个样本的每个snp位点均进行pcr扩增产物的上、下游引物的直接测序。
3.3.4序列分析
使用mutationsurveyor5.02软件对每个个体的正、反相测序峰图进行分析,确定各样本不同位点扩增产物测序结果的突变位点和突变方式。测序结果异常或无峰图或者峰图异常的进行二次测序,直至获得稳定和一致的测序结果。
3.3.5统计分析
3.3.5.1基因频率和基因型频率
使用popgen32软件计算各snps的基因频率和基因型频率。
①群体中某基因型频率=基因型个体数/该群体个体数×100%;
②体中某基因频率=该性状纯合基因型频率+1/2×杂合基因型频率。
3.3.5.2hardy-weinberg平衡检测
运用χ2鉴定各品种基因型频率和基因频率是否符合hardy-weinberg平衡。
其中:m代表基因型数目;fi表示观察到的第i个基因型的个体数;n代表总的样本数;pi表示第i个基因型的理论基因型频率。
3.3.5.3多态信息含量(polymorphisminformationcontent,pic)
其中:pi和pj分别为第i个和第j个等位基因频率;n为等位基因数。
3.3.5.4位点杂合度(h)
其中:pi为基因频率;m为等位基因数;r为位点数;h为平均杂合度。
3.3.5.5shannon信息含量(shannoninformationcontent,sic)
sic=-clogpi
其中:pi为第i个等位基因在群体中的频率,c为常数。
3.3.5.6关联分析
使用spss20软件,基于一般线性模型(glm)挖掘与湖羊肉用系核心群体尺性状表型相关的snps。
由于分析的所有个体均为来自同一饲养场相同饲养环境和管理条件的羊只,因此数据建模时不包括场效应。
具体模型为:y=xβ+e
其中,
y:湖羊肉用系核心群体尺性状、体重性状表型值向量;
β:表型均值、snp等固定效应向量;
e:残差效应向量;
x为β的关联矩阵。
当y为湖羊肉用系核心群初生重表型值向量时,按照以下模型分析:
y=xβ+sα+e;
其中:
y:湖羊肉用系核心群体尺性状表型值向量;
α:同胞数的固定效应向量;
β:snp效应向量;
e:残差效应向量;
x、s分别为β、α的关联矩阵。
3.4结果与分析
3.4.1表型的描述性统计分析
湖羊肉用系核心群g3代个体(萧山)体尺、体重性状相关指标的描述统计见表3-3,相关分析结果见表3-4。
相关分析结果显示,湖羊肉用系核心群g3代个体的同胞数与初生重呈极显著负相关(r=0.640,p<0.01),与断奶重呈显著负相关(r=-0.164,p<0.05);体高和周岁体重呈极显著正相关(r=0.281,p<0.01),胸围与周岁体重呈极显著正相关(r=0.721,p<0.01)。另外,体高与胸围也呈极显著正相关(r=0.186,p<0.01)。
表3-3湖羊肉用系核心群g3代个体(萧山)体尺和体重性状的
描述性统计分析
表3-4湖羊肉用系核心群g3代个体(萧山)体尺性状和体重的相关分析
3.4.2湖羊体高性状相关snps检测引物的pcr扩增
由图3可知,各引物pcr产物亮度较高,条带单一,无非特异性扩增,实际pcr产物与预计pcr扩增产物大小一致,可进行后续pcr-产物直接测序试验。
3.4.3湖羊体高性状相关snps位点扩增产物突变分析
8个湖羊体尺性状相关snps位点扩增产物突变分析结果如表3-5所示。
表3-5湖羊体高性状相关snps位点及突变类型、方式
除引物对9f和9r的扩增产物检测到的突变仅对应目的snp位点外,其余7对引物的扩增产物在目的snp位点附近还检测到其它突变。
其中,3f和3r(扩增产物长度385bp)、4f和4r(扩增产物长度418bp)、6f和6r(扩增产物长度500bp)这3对引物共检测到2个突变位点;5f和5r的扩增产物为3个突变位点(扩增产物长度269bp);7f和7r(扩增产物长度439bp),8f和8r(扩增产物长度307bp)的扩增产物检测到4个突变位点;10f和10r的扩增产物检测到突变最多,在长度为262bp的扩增产物中发现了突变位点竟高达9个,这8对引物湖羊肉用系核心群g3个体发现的27个突变及其突变方式详见表3-4。
该结果表明,湖羊群体中,ovinesnp50genotypingbeadchip芯片使用的snp附近可能还存在很多其它类型的突变有待深入发掘。
另外,一个比较有意思的结果是,这些8对引物pcr产物测序直接发现的突变位点虽然为27个,但5f和5r的扩增产物中s10476.1下游100bp的突变方式却有2种,因而该位点的等位基因不是常见的2个,变为3个。相应的,27个突变位点,28种突变方式中转换占82.1%(23/28),颠换占17.9%(5/28)。
从27个snps的基因注释结果看,其中22个位于基因间隔区,另外5个均位于基因内含子,分别为oar15_18440393.1位于ddx10基因内含子8,s10347.1位于samd12基因内含子3,s11279.1位于rnf217基因内含子3,oar1_164254640.1位于cadm2基因内含子7,oarx_76354330.1位于capn6基因内含子2。
3.4.4湖羊体高性状相关snps的遗传参数
27个snp位点的有效等位基因数、shannon信息含量和平均杂合度计算结果见表3-6。
其中,5f和5r引物对pcr扩增产物中s10476.1下游100bp对应的a→g和a→c突变遗传参数均为最高,有效等位基因数、shannon信息含量和平均杂合度分别为2.1584、0.8353和0.5367,具有丰富的遗传多态。而3f和3r引物对pcr扩增产物中oar6_95218086.1(c→t),7f和7r引物对pcr扩增产物中oar1_164254640.1下游149bp处c→a突变的遗传参数最低,有效等位基因数、shannon信息含量和平均杂合度分别为1.0713、0.1500和0.0666,遗传变异范围相对较窄。
3.4.5湖羊体高性状相关snps的群体遗传学分析
多态信息含量(pic)是用来判定和分析一个遗传标识所表达的信息含量,当pic>0.5是高度多态位点,当0.25<pic<0.5是中度多态位点,pic<0.25是低度多态位点。
3-6湖羊体高性状相关snps位点的遗传参数
表3-7湖羊体高性状相关snps位点的群体遗传学分析
表3-8湖羊体高性状相关snps位点的hardy-weinberg平衡检验
表3-9湖羊体高性状相关snps位点与体重性状的关联分析
注:相同一列数据后面如果没有字母,或者有相同的字母则表示差异不显著(p>0.05)。有不同小的字母则表示差异显著(p<0.o5)
不同大写字母表示差异极显著(p<0.01)。
本试验中有14个位点的pic值小于0.25,分别为oar6_95218086.1上游44bpg→a突变、oar6_95218086.1c→t突变、s10476.1a→c突变、s10476.1下游16bpt→c突变、oarx_120998827.1上游126bpa→c突变、oarx_120998827.1c→t突变、oar1_164254640.1g→a突变、oar1_164254640.1下游149bpc→a突变、oar1_164254640.1下游192bp和235bp处的t→c突变、s10347.1下游72bpt→c突变、s10347.1下游83bpt→c突变、oar6_90337552.1上游41bpg→a突变、oar6_90337552.1下游83bpc→a突变,显示这些位点为低度多态。剩余的13个snps的pic值都在0.25~0.5之间,属于中度多态(表3-7)。
3.4.6湖羊体高性状相关snps的hard-weinberg平衡检验
χ2检验结果表明,除oar6_90337552.1下游50bpt→c突变,和oar6_90337552.1下游79bpc→t未达到hard-weinberg平衡状态(p<0.05),其余25个snp均已处于hard-weinberg平衡状态(表3-8)。
3.4.7湖羊体高性状相关snps与体尺性状、体重性状的关联分析
利用“3.3.5.6关联分析”列出的glm模型对湖羊体高性状相关snps与体尺性状、体重性状进行关联分析(表3-9)。
3.4.7.1湖羊体高性状相关snps与湖羊肉用系核心群g3代个体体尺性状的关联分析
分析结果显示,检测的8对引物的27个突变位点对湖羊肉用系核心群g3代个体体尺性状的影响均未达到显著性水平(p>0.05)。
3.4.7.2湖羊体高性状相关snps与湖羊肉用系核心群g3代个体体重性状的关联分析
分析结果显示,检测的8对引物中有4对引物的7个突变位点可显著影响湖羊肉用系核心群g3代个体的体重性状(p<0.05或p<0.01)。
引物对3f和3r扩增产物中oar6_95218086.1上游44bpg→a的突变显著影响湖羊肉用系核心群g3代个体断奶重(p<0.05)和6月龄重(p<0.05)。其中突变纯合型aa个体的断奶重(17.83±2.35kg)极显著高于突变杂合型ga个体(16.21±1.76kg)(p<0.01),突变纯合型aa个体的6月龄重(37.82±2.64kg)显著高于突变杂合型ga个体(36.19±2.09kg)(p<0.05),该位点与断奶重和6月龄重显著相关。
引物对5f和5r扩增产物中s10476.1下游16bpt→c突变虽然可以显著影响湖羊肉用系核心群g3代个体的断奶重(p<0.05)和6月龄体重(p<0.01),该位点与断奶重和6月龄重显著相关。
引物对7f和7r扩增产物中oar1_164254640.1下游192bpt→c突变、oar1_164254640.1下游235bpt→c突变完全连锁,这两个位点单独或组合都可以显著影响湖羊肉用系核心群g3代个体的断奶重和6月龄重(p<0.05),极显著影响湖羊肉用系核心群g3代个体的初生重(p<0.01)。oar1_164254640.1下游192bp或235bp的突变型cc个体的初生重(3.59±0.04kg)极显著高于tt型个体(2.91±0.33kg)和突变型tc个体(2.88±0.32kg)(p<0.01);突变型cc单倍个体的断奶重(22.55±1.63kg)极显著高于tt型个体(17.66±2.36kg),和突变型tc个体(17.57±2.10kg)(p<0.01)。突变型cc单倍个体的6月龄重(45.90±0kg)极显著大于tt型个体(37.62±2.65kg)和tc型个体(37.66±.33kg)(p<0.01)。oar1_164254640.1下游192bp和235bp的突变纯合单倍型cccc个体的初生重(3.59±0.04kg)极显著高于野生纯合单倍型tttt个体(2.91±0.33kg)和突变杂合单倍型tctc个体(2.88±0.32kg)(p<0.01);突变纯合型cccc单倍个体的断奶重(22.55±1.63kg)极显著高于野生纯合型tttt单倍型个体(17.66±2.36kg),和突变杂合单倍型tctc个体(17.57±2.10kg)(p<0.01)。突变纯合型cccc单倍个体的6月龄重(45.90±0kg)极显著大于tttt单倍型个体(37.62±2.65kg)和tctc单倍型个体(37.66±.33kg)(p<0.01)。
引物对10f和10r扩增产物中oar6_90337552.1上游41bpg→a突变可以显著影响湖羊肉用系核心群g3代个体的初生重、断奶前日增重、断奶重和6月龄体重。
引物对10f和10r扩增产物中oar6_90337552.1下游79bpc→t极显著影响湖羊肉用系核心群g3代个体的断奶重和6月龄重(p<0.01),ct基因型个体的断奶重(18.10±2.28kg)和6月龄重(38.13±2.44kg)极显著大于tt个体断奶重(16.11±2.31kg)和6月龄重(35.92±2.79kg)(p<0.01)。虽然不同基因型个体间的初生重、断奶前日增重、断奶到6月龄日增重没有显著差异,但呈现了ct>cc>tt的趋势。这对引物扩增产物中oar6_90337552.1下游83bpc→a突变还可显著影响湖羊肉用系核心群g3代个体的6月龄重和6月龄到周岁的日增重(p<0.05),突变杂合型ca个体的6月龄重(38.09±2.81kg)、6月龄到周岁日增重(0.23±0.06kg)均显著大于突变纯合型aa个体的6月龄体重(33.80±3.91kg)和6月龄到周岁日增重(0.31±0.10kg)(p<0.05),野生型个体cc的6月龄重(37.64±2.51kg,p<0.01)、6月龄到周岁日增重(0.24±0.06kg,p<0.05)均显著大于突变纯合型aa个体。
3.5讨论
3.5.1snp位点及突变类型的研究
3.5.1.1snp的突变形式
snp是由单个碱基的颠换、转换、缺失或插入的突变导致的dna序列多态性[72-74],通常变异频率大于1%[73]。由于snp位点涉及的是单个碱基的变异,这种变异可能是转换(嘧啶与嘧啶,嘌呤与嘌呤之间的突变一般称为转换,即
在研究的湖羊肉用系核心群中,我们共在27个位点发现了28种类型的突变,其中转换突变的比例为82.1%(23/28),颠换占17.9%(5/28),转换高于基因组中一般发生的比例,颠换则低于基因组中一般发生的比例。另外,从理论上说一个snp位点应该有4种等位基因,即a、t、c、g,但一般却只出现2种等位基因,因此又叫双等位基因。在27个突变位点中,我们发现了个有意思的结果是,5f和5r的扩增产物中s10476.1下游100bp的突变方式有2种,即a→g、a→c,其中a→g占91.6%(186/203),a→c占8.4%(17/203),该位点的等位基因对应为a、g、c这3个。snp三等位基因和四等位基因的报道并不罕见[75],但其产生的具体机制尚不明确。s10476.1下游100bp的突变位于基因间隔区,与其他位点相比,因为该位点呈现三等位基因,表现出比其他位点更高的多态信息含量。
日本科学家m.kimura(1968)提出的中性学说认为在分子水平上,大多数进化演化和物种内的大多数变异,不是由自然选择引起的,而是通过那些对选择呈中性或近中性的突变等位基因的遗传漂变引起的。即这些位点的突变形式对生物个体的生存和生长来说既无坏处,也没有益处。虽然s10476.1下游100bp的a→g、a→c突变是gwas筛选出的与湖羊肉用系核心群g1和g2代个体体尺性状相关的snp位点,但在湖羊肉用系核心群g3代个体的群体验证中,该位点的glm分析结果显示,该snp并不影响湖羊肉用系核心群g3代个体的体尺和体重性状,因此,我们认为该三等位基因的snp位点更倾向于中性突变。
3.5.1.2snp突变位置分布
snp常常被人为的划分为两种形式:一种是功能性突变,主要发生在基因编码区。另一种是单碱基的突变,主要发生在非编码区。由于存在选择压力,基因组中snp的分布常常是不均匀的,在非编码区的突变频率要远远大于编码区的频率。以人类基因组snp为例,100万个snp中,处于编码区的大概有24~40万个,处于非编码区的大概有50万个,但大概只有20%~30%的编码区突变会引起非同义编码snps,从而引起蛋白质的功能变化。
本试验对gwas分析结果获得的snps位点验证结果显示,设计的引物中仅有4对引物7个位点可以显著影响湖羊肉用系核心群g3代个体的体重指标,而其中5个snp位于基因间隔区,2个snp位于基因的内含子。
3.5.2snps的遗传多样性分析
现代遗传学家普遍认为,遗传变异是有机体适应环境变化的先决条件[76]。遗传多样性的存在使物种的生存能力极大地提升。衡量群体内遗传变异大小的指标不仅包括遗传杂合度,还包括多态信息含量。杂合度能够反应群体在多个基因座位上遗传变异的程度,平均杂合度的值越大,说明群体内发生的遗传变异程度也就越大,反之越小。多态信息含量是用来描述一个遗传标记所表达的信息含量。
通过对试验群体进行遗传变异和群体杂合性分析,研究发现湖羊的snp位点的pic除了上述出现的14个表现为低度多态以外,其余均表现为中度多态性。这表明了该群体的基因多样性较高。
湖羊肉用系核心群g3代个体引物对10f和10r中,oar6_90337552.1下游50bpt→c(p<0.05)和oar6_90337552.1下游79bpc→t(p<0.01)均显著或极显著地偏离hardy-weinberg平衡,这两个位点突变杂合型的观察值数均超过其理论数,而野生型个体和突变纯合型个体的观察数则小于相应的理论数。群体未达到hardy-weinberg平衡可能是因为存在人工选择或近亲等其他因素引起的的基因频率的改变[77]。
我们对8对引物snp位点的gwas分析结果显示,oar6_90337552.1下游79bpc→t这个位点可极显著影响与湖羊肉用系核心群g3代个体的断奶重和6月龄体重。突变杂合型ct个体的断奶重(18.10±2.28kg)和6月龄重(38.13±2.44kg)极显著大于突变纯合型tt个体断奶重(16.11±2.31kg)和6月龄重(35.92±2.79kg)(p<0.01),另外,ct个体的断奶重和6月龄重也高于野生型cc个体断奶重(17.41±2.29kg)和6月龄重(37.40±2.71kg),但二者差异不显著(p>0.05)。由于该位点与湖羊核心群g3代个体的断奶重和6月龄体重极显著相关,在湖羊肉用系人工选育的过程中会因突变杂合型个体表现出高断奶重和6月龄重而被选择,最终引起突变杂合型基因型频率的显著上升。而该位点的
hardy-weinberg平衡检测结果也可以对这一推论进行佐证。该结果既说明了我们在该羊场湖羊肉用系的选育工作非常有成效,也能看到湖羊肉用系选育的分子标记辅助育种工作用于后续个体的早期选育的广阔前景。
3.5.3群体遗传变异与分化
杂合度(h)即群体中被检测的位点上杂合子的频率,如果pic越大,则可以说明ne和h就越大,当这些参数比较大时表明群体在改为点的变异性高。本研究结果表明,s10476.1下游100bpa→g、a→c各遗传参数均为最高,具有较高的遗传多样性。
3.5.4与湖羊肉用系核心群g3代个体体重性状相关的snps
3.5.4.1与体重相关
3f+3r扩增产物中oar6_95218086.1上游44bpg→a突变glm分析结果显示,该snp与湖羊肉用系核心群g3代个体的断奶重和6月龄体重呈显著相关(p<0.05)。该位点位于gc和npffr2基因间隔区,gc基因下游209kb处。npffr2属于rf氨基肽家族成员,npff在体液平衡、疼痛、摄食以及心血管功能中发挥重要的作用[76]。因此该位点的突变可能与npff基因在摄食调控中的功能有关,而其作用方式还需进一步的研究证实。
5f+5r扩增产物s10476.1下游16bpt→c突变与湖羊肉用系核心群g3代个体的与断奶重(p<0.05)和6月龄重显著相关(p<0.01)。该位点位于slc38a11和cobll1基因间隔区,slc38a11下游93kb。该突变的突变杂合型个体和突变纯合型个体的断奶重和6月龄重不存在显著差异(p>0.05),但在数值上均高于野生型个体。研究表明,cobll1涉及慢性淋巴细胞白血病和b细胞发育[77]。因此,突变杂合型和突变纯合型个体是否表现出较高的b细胞发育水平和更完善的机体免疫力,因而疾病抗性更强,表现出不易腹泻等表型,最终达到更高的断奶重和6月龄重?当然这种推断还需要后续大群体的验证。
7f+7r的扩增产物oar1_164254640.1下游192bpt→c和下游235bpt→c这两个位点单独或组合都与湖羊肉用系核心群g3代个体的初生重(p<0.01)、断奶重(p<0.05)和6月龄体重(p<0.05)显著相关。该位点位于cadm2基因内含子7。cadm2为细胞粘附分子2,为细胞粘附分子家族成员之一,可维持细胞极性和抑制肿瘤[77],及全身能量稳态的有力调节剂。研究发现,降低cadm2表达可以逆转包括肥胖,胰岛素抵抗和葡萄糖稳态受损在内的多种与代谢综合征相关的症状[55]。我们的研究结果显示,突变型cc个体的初生重(3.59±0.04kg)极显著高于tt型个体(2.91±0.33kg)和突变型tc个体(2.88±0.32kg)(p<0.01);突变型cc单倍个体的断奶重(22.55±1.63kg)极显著高于tt型个体(17.66±2.36kg),和突变型tc个体(17.57±2.10kg)(p<0.01)。突变型cc单倍个体的6月龄重(45.90±0kg)极显著大于tt型个体(37.62±2.65kg)和tc型个体(37.66±.33kg)(p<0.01)。突变纯合单倍型cccc个体的初生重(3.59±0.04kg)极显著高于野生纯合单倍型tttt个体(2.91±0.33kg)和突变杂合单倍型tctc个体(2.88±0.32kg)(p<0.01);突变纯合型cccc单倍个体的断奶重(22.55±1.63kg)极显著高于野生纯合型tttt单倍型个体(17.66±2.36kg),和突变杂合单倍型tctc个体(17.57±2.10kg)(p<0.01)。突变纯合型cccc单倍个体的6月龄重(45.90±0kg)极显著大于tttt单倍型个体(37.62±2.65kg)和tctc单倍型个体(37.66±2.33kg)(p<0.01)。但是该位点突变纯合单倍型个体的基因型频率仍然偏低,显示在该位点进行突变纯合单倍型个体的定向选育可能在肉用性状选育方面达到更好的选育效果。
10f+10r的扩增产物oar6_90337552.1上游41bpg→a突变可以显著影响湖羊肉用系核心群g3代个体的初生重、断奶前日增重、断奶重和6月龄体重。oar6_90337552.1下游79bpc→t与湖羊肉用系核心群g3代个体的断奶重和6月龄体重极显著相关,oar6_90337552.1下游83bpc→a与湖羊肉用系核心群g3代个体的6月龄体重和6月龄到周岁日增重显著相关。这3个snp都位于epha5和loc101120496基因间,距离epha5下游1446kb。epha5为受体酪氨酸激酶,是一种新的dna损伤修复调节剂。研究显示,该基因不仅可以调节电离辐射诱导的dna损伤修复过程,也是肺癌的功能性靶点[78],同时在胚胎发育过程中也发挥着关键作用[79]。因此,10f+10r的扩增产物检测到的3个位点是否可以通过epha5基因影响动物的生长发育,继而影响湖羊的肉用性能?这一推论仍需进一步的试验验证。
3.5.4.2湖羊肉用系核心群体尺性状相关snps的gwas分析结果及其群体验证的意义
在生产实践中,测量畜禽的体尺和体重,并间接的预测畜禽以后的产肉性能、产奶性能等指标,可以缩短育种年限,实现早期选育。已经有很多学者进行了畜禽体尺和体重指标的相关研究。例如,吴占福(2014)发现胫长、胸围可以较好的估测坝上长尾鸡公鸡体重[80]。石碧儒(2010)发现塞北乌骨鸡复冠鸡体重和胸角呈极显著相关,胸角是肉用型鸡测定的体尺指数,该指数可以用于鸡肉用品系的选育[81]。2012年俄木曲者等建立了羊的体尺与体重数据的线性回归模型,通过该数据模型获得的预测值与实际的测定值之间差异不显著,在实际育种工作中可以通过体尺性状测量指标预估群体中母羊的体重变化情况,他认为在简阳大耳羊黑色类群成年母羊的品系选育和生产研究中,应该把胸围作为第一选育指标,统筹体长这一指标,从而获得比较理想的育种繁殖效果[82]。鉴于体尺性状与体重性状的相关关系,我们对实施例1gwas分析获得的8个snps进行了群体验证,期望获得在大群体范围内可以用于湖羊体尺性状或体重性状早期选育的snp。
验证结果显示,群体中芯片检测的位点普遍存在,除了oar1_164254640.1(g→a)、s10476.1(a→c)和oar6_95218086.1(c→t)表现为低度多态外,其余5个snps均表现为中度多态。但是这些位点与湖羊肉用系核心群g3代的体尺、体重数据的关联分析结果显示,这些位点并不影响湖羊肉用系核心群g3代的体尺性状,不仅如此,这8个snp位点同样不对湖羊肉用系核心群g3代的体重性状造成显著影响。单纯从试验结果看,似乎gwas分析获得的结果并没有实际意义。
但我们比较两次数学模型的数据及其来源发现了以下问题:
(1)不同批次试验羊体尺指标测量的稳定性与准确性不能完全保证。
湖羊进行体尺测量时要求将被测个体牵引到平整地面使之稳定,并固定测量者,以减小试验误差。实施例1用于gwas分析的羊只均来自湖州太湖湖羊养殖专业合作社湖羊肉用系核心群,测量人员为同一人,因此测量的试验误差较小。但第三章群体验证使用的试验羊来自杭州庞大农业开发有限公司构建的湖羊肉用系核心群,测量人员为多人,因此该批次湖羊体尺指标测定的人为误差较大。与体尺测量对试验羊和测量者有较高要求不同的是,体重测量对环境和测量的要求较低,试验误差极小,因此湖羊体重数据在实际生产实践中不仅更容易采集,更能够保证数据的准确性。
(2)湖羊肉用系核心群体尺数据与体重数据显著相关。
有研究表明,生长速度性状与体尺性状的相关主要由6月龄重、胸围和体长等密切相关[83]。湖州太湖湖羊养殖专业合作社用于gwas分析的羊只周岁体高和6月龄体重的相关系数可达0.839(p<0.01),周岁胸围和6月龄体重的相关系数更高达0.893(p<0.01)。而杭州庞大农业开发有限公司群体验证使用的湖羊,周岁体高和6月龄体重(r=-0.147,p<0.05)、成年重(r=-0.164,p<0.05)的相关系数却呈现显著的负相关),周岁胸围和6月龄体重(r=0.281,p<0.01)、成年重(r=0.712,p<0.01)的均达到极显著的正相关。
由于gwas分析时所用群体的体尺指标和体重指标存在极显著的相关关系,因此,gwas分析获得的在全基因组水平与体尺性状显著相关的snp位点也可能与体重性状存在较高的相关关系,因此造成我们在后续的群体验证工作中,群体验证工作显示了这些位点与湖羊的初生重、断奶重、6月龄重和成年重在不同程度上的相关。
由于不同场间,试验羊试验管理水平和条件也不完全相同,且选育方向也不完全一致,因此,这一相关分析结果是否由测量误差所导致,还是由于选育方向造成仍需对两个场肉用系核心群不同代次之间的体尺和体重数据进行比较和分析。另外,两个场的相关分析虽然明确湖羊胸围与湖羊成年重的极显著相关关系,但很遗憾的是,在进行群体验证时,gwas分析中获得的唯一一个与胸围在全基因组水平显著相关的snp位点,oarx_76354330.1,未能设计出相应的单一扩增产物的引物,导致该位点的验证工作无法进行,因此,后续工作中能否再进行相应位点的扩增引物和扩增体系的优化,补充该位点的验证信息,对gwas结果的可靠性将是一个重要补充。
(3)gwas分析的作用。
根据我们的验证结果,我们使用的8对引物中仅有1对在目的扩增区域只检测到了相应待检测的位点,另外的7对都检测到了2个及其以上的snps。并且,检测出的27个snp位点(含新snps位点19个),后期验证中发现有多个新发现的snps与湖羊体重性状存在显著或极显著相关,并且这些位点的基因频率和基因型频率都或多或少地显示出与我们进行的湖羊肉用系核心群育种工作的相关性。而高达22.2%以上功能性snps位点的筛出比例与前期大量的gwas分析工作密不可分。这些有意义的位点正是由于与gwas分析获得的阳性位点距离较近,才能够纳入我们的分析视野当中。
因此,我们认为gwas是候选功能基因和候选功能性snp筛选的重要工具,它能够提供一种类似路标的作用,为后期的验证工作指明研究方向,而不会让研究人员被浩如烟海的序列信息所淹没。当然,如果能研制出密度更高的snp检测芯片将会大大提高结果的可靠性。gwas结合群体验证的结果表明,gwas是候选功能基因和候选功能性snp筛选的重要工具,所筛选出的体尺性状相关snp位点附近能够较大几率地筛选到与湖羊肉用性能显著相关的snps,这些snps可为后期的验证工作提供一种类似路标的作用,从而发现更有价值的候选功能性snps,并用于后续湖羊肉用系核心群个体的早期选育。
参考文献
[1]魏彩虹,杜立新.我国肉用绵羊育种现状与未来发展方向[j].中国草食动物科学,2012,s1):454-457.
[2]文伟,陶格素,图亚.巴彦淖尔成功培育具有知识产权肉羊品种[j].北京农业,2008,15):70-70.
[3]魏景钰,胡大君,隔日勒图雅.昭乌达肉羊新品种简介[j].中国畜牧兽医文摘,2013,8):39-40.
[4]内蒙古晨报.肉羊新品种被农业部命名为“察哈尔羊”[j].湖北畜牧兽医,2014,3):15-15.
[5]zhaofuping,weicaihong,zhangli,etal.agenomescanofrecentpositiveselectionsignaturesinthreesheeppopulations[j].journalofintegrativeagriculture,2016,15(1):162-174.
[6]almamunhawladera.,kwanpaul,clarksamuela.,etal.genome-wideassociationstudyofbodyweightinaustralianmerinosheeprevealsanorthologousregiononoar6tohumanandbovinegenomicregionsaffectingheightandweight[j].geneticsselectionevolution,2015,47(1):66.
[7]max.,guanl.,xuanj.,etal.effectofpolymorphismsinthecamkmtgeneongrowthtraitsinujumqinsheep[j].animalgenetics,2016,47(5):618-622.
[8]lizhang,liujiasen,zhaofuping,etal.genome-wideassociationstudiesforgrowthandmeatproductiontraitsinsheep[j].plosone,2013,8(6):e66569.
[9]matikaoswald,riggiovalentina,anselme-moizanmarie,etal.genome-wideassociationrevealsqtlforgrowth,boneandinvivocarcasstraitsasassessedbycomputedtomographyinscottishblackfacelambs[j].geneticsselectionevolutiongse,2016,48(1):11.
[10]demarsjulie,fabrestéphane,sarryjulien,etal.genome-wideassociationstudiesidentifytwonovelbmp15mutationsresponsibleforanatypicalhyperprolificacyphenotypeinsheep[j].plosgenetics,2013,9(4):e1003482.
[11]储明星,桑林华,王金玉,etal.小尾寒羊高繁殖力候选基因bmp15和gdf9的研究[j].journalofgenetics\s&\sgenomics,2005,32(1):38-45.
[12]gholizadehm,rahimimianjig,nejatijavaremia,etal.genomewideassociationstudytodetectqtlfortwinningrateinbaluchisheep[j].journalofgenetics,2014,93(2):489-493.
[13]garcíagámezelsa,gutiérrezgilbeatriz,sahanagoutam,etal.gwaanalysisformilkproductiontraitsindairysheepandgeneticsupportforaqtninfluencingmilkproteinpercentageinthelalbagene[j].2012,7(10):e47782.
[14]mouselm.r.,reynoldsj.o.,whites.n.genome-wideassociationidentifiesslc2a9andnlngeneregionsasassociatedwithentropionindomesticsheep[j].plosone,2015,10(6):e0128909.
[15]jawasrehk,boettcherp.j.,stellaa.genome-wideassociationscansuggestsbasisformicrotiainawassisheep[j].animalgenetics,2016,47(4):504-506.
[16]niggelera.,tetensj.,
[17]ebrahimif,gholizadehm,rahimi-mianjig,etal.detectionofqtlforgreasyfleeceweightinsheepusinga
[18]bolormaas,swana.a.,brownd.j.,etal.multiple-traitqtlmappingandgenomicpredictionforwooltraitsinsheep[j].geneticsselectionevolution,2017,49(1):62.
[19]lim.h.,tiirikkat.,kantanenj.agenome-widescanstudyidentifiesasinglenucleotidesubstitutioninasipassociatedwithwhiteversusnon-whitecoat-colourvariationinsheep(ovisaries)[j].heredity,2014,112(2):122-131.
[20]高中元,林婄婄,袁亚男,etal.plin基因多态性及其与绵羊尾形和屠宰性状的关联研究[j].山西农业大学学报(自然科学版),2012,32(2):158-164.
[21]刘真.肥尾和瘦尾羊全基因组选择信号检测.中国农业科学院,2015.
[22]zhucaiye,fanhongying,yuanzehu,etal.genome-widedetectionofcnvsinchineseindigenoussheepwithdifferenttypesoftailsusingovinehigh-density600ksnparrays[j].scirep,2016,6(27822.
[23]xus.s.,renx.,yangg.l.,etal.genome-wideassociationanalysisidentifiesthegeneticbasisoffatdepositioninthetailsofsheep(ovisaries)[j].animalgenetics,2017,48(5).
[24]martinp,palhièrei,tosser-kloppg,etal.heritabilityandgenome-wideassociationmappingforsupernumeraryteatsinfrenchalpineandsaanendairygoats[j].journalofdairyscience,2016,99(11):8891-8900.
[25]兰蓉,朱兰,姚新荣,etal.山羊产羔数全基因组关联分析[j].畜牧兽医学报,2015,46(4):549-554.
[26]martinp,palhièrei,maroteauc,etal.agenomescanformilkproductiontraitsindairygoatsrevealstwonewmutationsindgat1reducingmilkfatcontent[j].scientificreports,2017,7(1):1872.
[27]davenportc.b.theoriginofblacksheepintheflock[j].science,1905,22(569):674-675.
[28]mariemartinpauline,isabellepalhière,annericard,etal.genomewideassociationstudyidentifiesnewlociassociatedwithundesiredcoatcolorphenotypesinsaanengoats[j].plosone,2016,11(3):e0152426.
[29]beckerd,ottom,ammannp,etal.thebrowncoatcolourofcopperneckedgoatsisassociatedwithanon-synonymousvariantatthetyrp1locusonchromosome8[j].animalgenetics,2015,46(1):50-54.
[30]潘东东,李正帮,张维,etal.全基因组关联研究综述[j].应用概率统计,2014,30(1):84-103.
[31]凃欣,石立松,汪樊,etal.全基因组关联分析的进展与反思[j].生理科学进展,2010,41(2):87-94.
[32]yangj.,zaitlenn.a.,goddardm.e.,etal.advantagesandpitfallsintheapplicationofmixed-modelassociationmethods[j].naturegenetics,2014,46(2):100-106.
[33]xus.mappingquantitativetraitlocibycontrollingpolygenicbackgroundeffects[j].genetics,2013,195(4):1209-1222.
[34]张统雨,朱才业,杜立新,etal.羊重要性状全基因组关联分析研究进展[j].遗传,2017,39(6):491-500.
[35]高志英,於建国,阿曼姑阿巴斯,etal.多浪羊羔羊断奶体重体尺指标及其相关性研究[j].草食家畜,2009,4):29-31.
[36]赵子贵.隆林山羊微卫星标记多态性及其与生长性状的相关性研究.广西大学,2013.
[37]martinez-royoa,alabartj.l.,sartop,etal.genome-wideassociationstudiesforreproductiveseasonalitytraitsinrasaaragonesasheepbreed[j].theriogenology,2017,99(21-29.
[38]王珊,李晓林,牛志刚,etal.多浪羊多胎基因全基因组关联分析研究[j].江西农业学报,2017,29(5):77-81.
[39]张莉.绵羊肉用性状全基因组关联分析.中国农业科学院,2013.
[40]atlijam,arranzj.j.,martinezvalladaresm,etal.detectionandreplicationofqtlunderlyingresistancetogastrointestinalnematodesinadultsheepusingtheovine50ksnparray[j].geneticsselectionevolutiongse,2016,48(1):4.
[41]cecchif,russoc,iamartinod,etal.identificationofcandidategenesforparatuberculosisresistanceinthenativeitaliangarfagninagoatbreed[j].tropicalanimalhealth&production,2017,49(6):1135-1142.
[42]刘书东.中国美利奴羊(新疆型)毛品质性状全基因组关联分析.石河子大学,2017.
[43]seroussie,rosova,shiraka,etal.unveilinggenomicregionsthatunderliedifferencesbetweenafec-assafsheepanditsparentalawassibreed[j].geneticsselectionevolutiongse,2017,49(1):19.
[44]kardosm,luikartg,bunchr,etal.wholegenomeresequencinguncoversmolecularsignaturesofnaturalandsexualselectioninwildbighornsheep[j].molecularecology,2015,24(22):5616.
[45]johnstons.e.,mcewanj.c.,pickeringn.k.,etal.genome-wideassociationmappingidentifiesthegeneticbasisofdiscreteandquantitativevariationinsexualweaponryinawildsheeppopulation[j].molecularecology,2011,20(12):2555-2566.
[46]朱才业.不同尾型绵羊全基因组关联分析、拷贝数变异及选择信号检测.中国农业大学,2017.
[47]任雪.利用全基因组关联分析(gwas)鉴定绵羊多角性状和脂尾性状的候选功能基因.中国科学院大学,2016.
[48]landereric,kruglyakleonid.geneticdissectionofcomplextraits:guidelinesforinterpretingandreportinglinkageresults[j].naturegenetics,1995,11(3):241-247.
[49]lidauerm,
[50]谭贤杰,吴子恺,程伟东,etal.关联分析及其在植物遗传学研究中的应用[j].植物学报,2011,46(1):108-118.
[51]张立敏,张猛,周正奎,etal.基于高密度snp芯片技术对中国西门塔尔牛scd1基因与肉质性状的相关性分析[j].畜牧兽医学报,2012,43(6):878-886.
[52]lidongliang,zhangyongjian,zhanghe,etal.cadm2,asanewtargetofmir-10b,promotestumormetastasisthroughfak/aktpathwayinhepatocellularcarcinoma[j].journalofexperimental&clinicalcancerresearch,2018,37(1):46.
[53]hewei,lixuesong,xushuping,etal.aberrantmethylationandlossofcadm2tumorsuppressorexpressionisassociatedwithhumanrenalcellcarcinomatumorprogression[j].biochemical&biophysicalresearchcommunications,2013,435(4):526-532.
[54]hirumaa,ikedas,teruit,etal.anovelsplicingvariantofcadm2asaprotectivetranscriptofpsoriasis[j].biochemical&biophysicalresearchcommunications,2011,412(4):626.
[55]yanx.,wangz.,schmidtv,etal.cadm2regulatesbodyweightandenergyhomeostasisinmice[j].molmetab,2017,8(c).
[56]krauselucianam.fontanari,jappannasophia,krausealexandre,etal.identificationandcharacterizationofostl(rnf217)encodingaring-ibr-ringproteinadjacenttoatranslocationbreakpointinvolvingetv6inchildhoodall[j].scientificreports,2014,4(4):6565.
[57]hoshiuhrong,leeyuju,linweeichin.regulationofrnf144ae3ubiquitinligaseactivitybyself-associationthroughitstransmembranedomain[j].journalofbiologicalchemistry,2015,290(38):23026-23038.
[58]杨乐,黄林,金素钰,etal.牦牛和犏牛睾丸samd12基因的克隆测序及其mrna水平比较[j].西南民族大学学报(自然科学版),2016,42(5):525-530.
[59]ishiurah,doik,mitsuij,etal.expansionsofintronictttcaandttttarepeatsinbenignadultfamilialmyoclonicepilepsy[j].naturegenetics,2018,88(13):1296.
[60]王颖.ddx10在肝癌中的表达及促进增殖的相关机制研究.山东大学,2017.
[61]minhongjung,teitelbaumstevenl,tae-hokim,etal.calpain-6,atargetmoleculeofglucocorticoids,regulatesosteoclasticboneresorptionviacytoskeletalorganizationandmicrotubuleacetylation[j].journalofbone&mineralresearchtheofficialjournaloftheamericansocietyforbone&mineralresearch,2011,26(3):657-665.
[62]tonamik,kuriharay,arimas,etal.calpain-6,amicrotubule-stabilizingprotein,regulatesrac1activityandcellmotilitythroughinteractionwithgef-h1[j].journalofcellscience,2011,124(8):1214-1223.
[63]tonamik,kuriharay,aburatanih,etal.calpain6isinvolvedinmicrotubulestabilizationandcytoskeletalorganization[j].molecular&cellularbiology,2007,27(7):2548-2561.
[64]deart.n.,boehmt.diversemrnaexpressionpatternsofthemousecalpaingenescapn5,capn6andcapn11duringdevelopment[j].mechdev,1999,89(1–2):201-209.
[65]管峰,艾君涛,庞训胜,etal.黄淮山羊微卫星多态性及其与产羔数相关性的研究[j].中国畜牧杂志,2006,42(23):4-7.
[66]赵宗胜,王根林,马玉萍,etal.绵羊微卫星标记与部分毛用性状的关系研究[j].畜牧兽医学报,2006,37(9):864-869.
[67]甘尚权,张伟,沈敏,etal.绵羊x染色体59383635位点多态性与脂尾性状的相关性分析[j].遗传,2013,35(10):1209-1216.
[68]王世银,张伟,沈敏,etal.绵羊x染色体59194976位点多态分析及其与尾(臀)脂性状相关性研究[j].基因组学与应用生物学,2013,32(5):575-580.
[69]张伟,沈敏,李欢,etal.绵羊x染色体59571364与59912586位点在脂尾、瘦尾绵羊群体中的多态性检测及分析[j].遗传,2013,35(12):1384-1390.
[70]周勇,朱万斌.湖羊与引进肉羊杂交后代产肉性能及肉品质研究[j].畜牧兽医杂志,2016,35(5):1-4.
[71]martine.r.,laie.h.,gilbertj.r.,etal.snpingawayatcomplexdiseases:analysisofsingle-nucleotidepolymorphismsaroundapoeinalzheimerdisease[j].americanjournalofhumangenetics,2000,67(2):383-394.
[72]张建勤.宁夏固原地区两个鸡群体遗传多态性研究.西北农林科技大学,2004.
[73]张建勤.固原鸡耐寒性状微卫星标记及其相关基因遗传变异研究.西北农林科技大学,2008.
[74]张宏礼,张鸿雁.关于最大信息熵原理与群体遗传平衡一致性的探讨[j].遗传,2006,28(3):324-328.
[75]crousec.a.,rogerss,amiotte,etal.analysisandinterpretationofshorttandemrepeatmicrovariantsandthree-bandedallelepatternsusingmultiplealleledetectionsystems[j].journalofforensicsciences,1999,44(1):87-94.
[76]张晓媛.带有荧光标签的npff2受体重组细胞模型建立.兰州大学,2013.
[77]
[78]staquicinifernanda,dobroffandrey,ferrarafortunato,etal.abstractlb-006:receptortyrosinekinaseepha5isafunctionalmoleculartargetinhumanlungcancer[j].cancerresearch,2015,75(15supplement):lb-006-lb-006.
[79]staquicinif.i.,qianm.d.,salameha,etal.receptortyrosinekinaseepha5isafunctionalmoleculartargetinhumanlungcancer[j].journalofbiologicalchemistry,2015,290(12):7345.
[80]吴占福,刘海斌,于文文,etal.坝上长尾鸡体重与体尺指数相关与回归分析.京津冀一体化畜牧兽医科技创新研讨会暨“瑞环杯”新思想、新方法、新观点论坛,2014.
[81]石碧儒.塞北乌骨鸡体重与体尺指数相关性分析[j].中国畜禽种业,2010,06(7):122-123.
[82]俄木曲者,范景胜,陈天宝,etal.简阳大耳羊黑色类群成年母羊体重与体尺指数回归分析[j].中国草食动物科学,2012,32(5):13-16.
[83]陈海燕,朱海平,傅衍,etal.大约克新品系生长速度和体尺性状间的典型相关分析[j].畜牧与兽医,2003,35(5):21-22.
[84]郭家中.奶牛重要经济性状的全基因组关联分析.西北农林科技大学,2013.
序列表
<110>浙江省农业科学院
<120>包括snp3的分子标记及其在湖羊辅助育种中的应用
<160>12
<170>siposequencelisting1.0
<210>1
<211>385
<212>dna
<213>cp011891.1
<220>
<221>mutation
<222>((209))..((209))
<223>risgora
<400>1
gtcctacttgtggtctctgtttcccctcaatgggccttacaaatgctaatacatgtttaa60
gggtaattaacattacttaggttgattaacatggaaattaaaaagaaaaaaagctgaaga120
gaaagttgagacttcttccctacatggtagaaatttaaagggactcattcccaaaagaca180
aatctatagatgtttattaaagaaatggraagagtgagacagacattaggattaaagcaa240
caggttttgatacggtgctacctaagattagatgggccaacatgaaaggggcccaaacaa300
atccactctacatttatagtacagaactagggtaaatatttaaaaggttataaaaaattg360
cactgagatattaggccagcaacac385
<210>2
<211>269
<212>dna
<213>cp011887.1
<220>
<221>mutation
<222>((129))..((129))
<223>yistorc
<400>2
gtctgatggatgtgctgtagtttctaggccaactgagttcacagtggtcaagataattat60
aaagccattgaattgcttttcttctttgtgctcacgtggttgcacgttacacacaaagtg120
gaagtggcygagagaggagcagtctcctcctcctttttgtcaggtatcccagctccagga180
caatcaggaaatagacagggtccctatcttacaggacagcctctgtgttttgcggtctca240
gcagctctcgttattctcaggtgtgcatg269
<210>3
<211>439
<212>dna
<213>cp011886.1
<220>
<221>mutation
<222>((303))..((303))
<223>yistorc
<220>
<221>mutation
<222>((373))..((373))
<223>yistorc
<400>3
gctgggatgaaagagattaaccattagctagtttggcagaaagctaaagaagagatctgg60
aaggttttaagaaaccaagactaagtgattaacagggttcatgagtggttgataatccag120
ggtttgcattgtataaagcctcgttctagcatgtttcccagttcccttatgctagtctta180
ttttaaccacaaaattaccctgttgtaaaaggtttacaaagacccttagactgttttcat240
tttatcgataaggacagtgggccttgtggaggttatttgaccttttctagtcacacagta300
aayaaatgatagaatggggattacaaaaatactttaagaaattttaacatcactgatggt360
tacattttcttaytttcacagtattttcacttctgtcatgttggcttttcaggataaccc420
agggaagttgacaaggatg439
<210>4
<211>262
<212>dna
<213>cp011891.1
<220>
<221>mutation
<222>((87))..((87))
<223>risgora
<220>
<221>mutation
<222>((207))..((207))
<223>yistorc
<220>
<221>mutation
<222>((211))..((211))
<223>miscora
<400>4
cagtctgaatcccaattatcactaacatgttataaatgtaggaaaacttcagttttgctc60
acccacaaattgttggccatcactgtrtctgggtgttaagtgagaagatgaatcttgaat120
gcttagcacagtgctgagaacaatatagtgaagtgagtgaagtcgcttagtcatgtctga180
ccctttgcaactccatggactgtagcytacmagcctcatctgtctgtgagattttccagg240
caatagtactggagtggatttc262
<210>12
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>12
gtcctacttgtggtctctgttt22
<210>6
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>6
gtgttgctggcctaatatctca22
<210>7
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>7
gtctgatggatgtgctgtagttt23
<210>8
<211>24
<212>dna
<213>人工序列(artificialsequence)
<400>8
catgcacacctgagaataacgaga24
<210>9
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>9
gctgggatgaaagagattaacca23
<210>10
<211>24
<212>dna
<213>人工序列(artificialsequence)
<400>10
catccttgtcaacttccctgggtt24
<210>11
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>11
cagtctgaatcccaattatcact23
<210>12
<211>25
<212>dna
<213>人工序列(artificialsequence)
<400>12
gaaatccactccagtactattgcct25
- 还没有人留言评论。精彩留言会获得点赞!