植物株型相关蛋白PROG2及其编码基因与应用的制作方法
本发明属于生物
技术领域:
,具体涉及一种植物株系相关蛋白PROG2及其编码基因与应用。
背景技术:
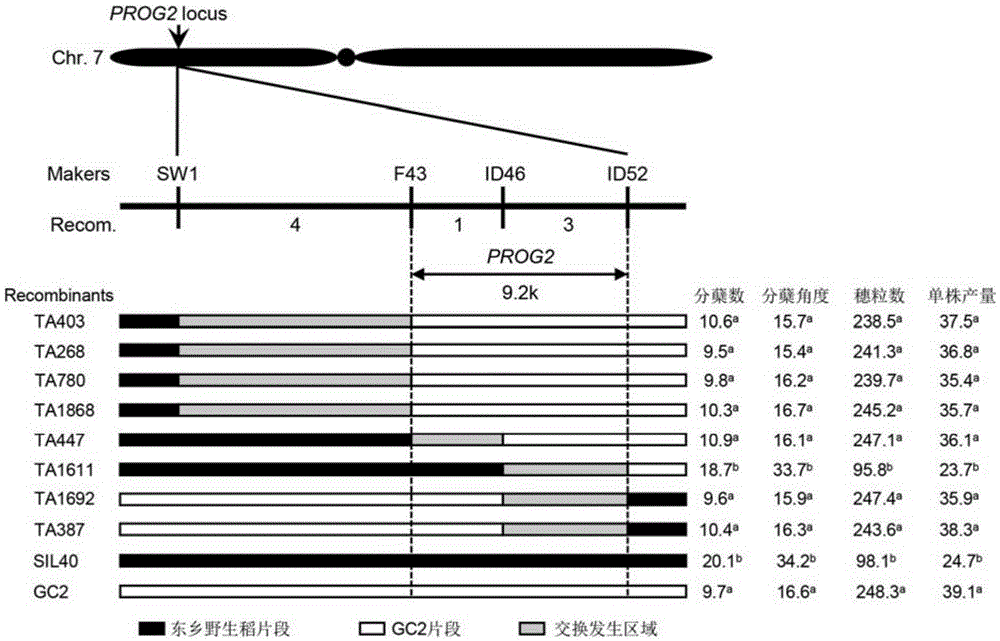
:水稻,作为世界上最重要的粮食作物之一,养育了世界上超过50%的人口。然而,最近的研究表明,在水稻三大生产国,中国、印度和印度尼西亚分别有78%、37%和81%的水稻生产区的水稻产量从1961年到2008年一直停滞不前,水稻栽培品种遗传背景狭窄和育种技术落后,是造成这一现象的主要原因。普通野生稻是亚洲栽培稻的祖先种,野生稻在演化成栽培稻的过程中,经过自然选择和人工选择,基因多样性降低、等位基因数减少。据统计,栽培稻的等位基因数约为野生稻的60%,从而导致当前水稻品种选育所面临的遗传瓶颈问题。因此从水稻近缘野生种的普通野生稻OryzarufipogonGriff.基因组中发掘和利用在栽培稻中已丢失或削弱的优异驯化基因,并把它们应用于水稻育种生产中具有十分重要的理论意义和实践价值,也是解决当前水稻育种难题的一条行之有效的途径。水稻株型是影响产量的关键因素之一,塑造理想株型也一直以来都是水稻育种者提高水稻产量的重要途径。我国野生稻资源丰富,从野生稻中发掘、定位和克隆水稻株型相关基因不仅有助于拓宽水稻栽培品种的遗传多样性,同时也为水稻现代分子育种提供重要参考和理论基础,对加强我国野生稻基因资源的保护、将资源优势变成经济优势具有重要的意义。技术实现要素:本发明所要解决的技术问题是如何提高水稻产量。为解决上述问题,本发明首先提供了一种植物株系相关蛋白。本发明所提供的植物株系相关蛋白,名称为PROG2,来源于江西东乡野生稻,为如下a1)或a2)或a3):a1)氨基酸序列是序列表中序列2所示的蛋白质;a2)在序列表中序列2所示的蛋白质的N端和/或C端连接标签得到的融合蛋白质;a3)将a1)或a2)所示的蛋白质经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的与植物株型相关的蛋白质。其中,序列表中序列2由188个氨基酸残基组成。为了使a1)中的蛋白质便于纯化,可在序列表中序列2所示的蛋白质的氨基末端或羧基末端连接上如表1所示的标签。表1、标签的序列标签残基序列Poly-Arg5-6(通常为5个)RRRRRPoly-His2-10(通常为6个)HHHHHHFLAG8DYKDDDDKStrep-tagII8WSHPQFEKc-myc10EQKLISEEDL上述a3)中的蛋白质PROG2,所述一个或几个氨基酸残基的取代和/或缺失和/或添加为不超过10个氨基酸残基的取代和/或缺失和/或添加。上述a3)中的蛋白质PROG2可人工合成,也可先合成其编码基因,再进行生物表达得到。上述a3)中的蛋白质PROG2的编码基因可通过将序列表中序列1所示的DNA序列中缺失一个或几个氨基酸残基的密码子,和/或进行一个或几个碱基对的错义突变,和/或在其5′端和/或3′端连上表1所示的标签的编码序列得到。编码所述PROG2的核酸分子也属于本发明的保护范围。所述编码所述PROG2的核酸分子可为如下(b1)或(b2)或(b3)或(b4)所示的DNA分子:(b1)核苷酸序列是序列表中序列1所示的DNA分子;(b2)其编码序列是序列表中序列1自5’末端起第125-691位核苷酸的DNA分子;(b3)与(b1)或(b2)限定的核苷酸序列具有75%或75%以上同一性,且编码权利要求1所述蛋白质的DNA分子;(b4)在严格条件下与(b1)或(b2)限定的核苷酸序列杂交,且编码权利要求1所述蛋白质的DNA分子。其中,所述核酸分子可以是DNA,如cDNA、基因组DNA或重组DNA;所述核酸分子也可以是RNA,如mRNA或hnRNA等。其中,序列表中序列1由892个核苷酸组成,序列表中序列1的核苷酸编码序列表中序列2所示的氨基酸序列。本领域普通技术人员可以很容易地采用已知的方法,例如定向进化和点突变的方法,对本发明的编码PROG2的核苷酸序列进行突变。那些经过人工修饰的,具有与本发明分离得到的PROG2的核苷酸序列75%或者更高同一性的核苷酸,只要编码PROG2且与植物株型相关,均是衍生于本发明的核苷酸序列并且等同于本发明的序列。这里使用的术语“同一性”指与天然核酸序列的序列相似性。“同一性”包括与本发明的编码序列表的序列2所示的氨基酸序列组成的蛋白质的核苷酸序列具有75%或更高,或80%或更高,或85%或更高,或90%或更高,或95%或更高同一性的核苷酸序列。同一性可以用肉眼或计算机软件进行评价。使用计算机软件,两个或多个序列之间的同一性可以用百分比(%)表示,其可以用来评价相关序列之间的同一性。含有所述编码所述PROG2的核酸分子的表达盒、重组载体、重组微生物或转基因细胞系也属于本发明的保护范围。所述表达盒包括启动子、编码所述PROG2的核酸分子和终止子。所述启动子具体可为PROG2启动子(序列3自5’末端起第1至1258位所示);所述编码所述PROG2的核酸分子可如序列3自5’末端起第1383至1949位所示;所述终止子具体可为PROG2终止子(序列3自5’末端起第2151至4214位所示)。所述重组载体可为将所述PROG2的编码基因(即序列表中序列1自5’末端起第125-691位所示的DNA分子)通过含有所述PROG2的编码基因的表达盒插入出发质粒得到的重组质粒。所述重组载体具体可为用序列表中序列3所示的DNA分子替换pCAMBIA3301的EcoRI和SmaI识别序列间的片段(pCAMBIA3301被限制性核酸内切酶EcoRI和SmaI切成一个大片段和一个小片段,该DNA为该小片段),得到的重组质粒D81,重组质粒D81表达序列表中序列2所示的PROG2。所述pCAMBIA3301与D81的差别仅在于将pCAMBIA3301的EcoRI和SmaI识别序列间的DNA片段(pCAMBIA3301被限制性核酸内切酶EcoRI和SmaI切成一个大片段和一个小片段,该DNA为该小片段)替换为序列表中序列3所示的DNA分子。所述重组微生物可通过将所述重组载体导入出发微生物得到。所述出发微生物可为酵母、细菌、藻类或真菌。所述细菌可为革兰氏阳性细菌或革兰氏阴性细菌。所述革兰氏阴性细菌可为根癌农杆菌(Agrobacteriumtumefaciens)。所述根癌农杆菌(Agrobacteriumtumefaciens)可为根癌农杆菌EHA105。所述转基因植物细胞系均不包括繁殖材料。所述转基因植物理解为不仅包含将所述PROG2基因转化受体植物得到的第一代转基因植物,也包括其子代。对于转基因植物,可以在该物种中繁殖该基因,也可用常规育种技术将该基因转移进入相同物种的其它品种,特别包括商业品种中。所述转基因植物包括种子、愈伤组织、完整植株和细胞。所述PROG2,或,所述编码所述PROG2的核酸分子,或,含有所述编码所述PROG2的核酸分子的表达盒、重组载体、重组微生物或转基因细胞系,在调控植物株型和/ 或产量中的应用也属于本发明的保护范围。所述PROG2,或,所述编码所述PROG2的核酸分子,或,含有所述编码所述PROG2的核酸分子的表达盒、重组载体、重组微生物或转基因细胞系,在培育株型改变和/或产量改变的转基因植物中的应用也属于本发明的保护范围。上述应用中,所述产量改变可为单株产量降低。上述应用中,所述株型改变可为株高降低和/或分蘖数增加和/或分蘖角度增大和/或一次枝梗数减少和/或二次枝梗数减少和/或主茎穗粒数减少。上述应用中,所述植物可为d1)-d4)中的任一种:d1)单子叶植物;d2)双子叶植物;d3)水稻;d4)水稻品种ZH17。为解决上述技术问题,本发明还提供了培育转基因植物的方法。本发明所提供的培育转基因植物的方法,包括将编码所述PROG2的核酸分子导入受体植物中,得到转基因植物的步骤;所述转基因植物与所述受体植物相比单株产量降低和/或株高降低和/或分蘖数增加和/或分蘖角度增大和/或一次枝梗数减少和/或二次枝梗数减少和/或主茎穗粒数减少。上述方法中,所述受体植物可为d1)-d4)中的任一种:d1)单子叶植物;d2)双子叶植物;d3)水稻;d4)水稻品种ZH17。实验证明,利用本发明提供的植物株型相关蛋白PROG2及其编码基因能调控植物株型:T0代转PROG2基因阳性植株表现出类似SIL40的表型;与受体植物ZH17相比,T0代转PROG2基因阳性植株株高降低,分蘖数增加,分蘖角度增大,一次枝梗数减少,二次枝梗数减少,主茎穗粒数减少和单株产量降低。可见,植物株型相关蛋白PROG2对调控水稻株型和产量具有非常重要的作用,在培育高产水稻新品种中具有广阔前景。附图说明图1为桂朝2号与东乡普通野生稻渗入系SIL40的表型比较。图2为PROG2基因的精细定位。图3为T0代转PROG2基因水稻植株与中花17的表型比较。具体实施方式下面结合具体实施方式对本发明进行进一步的详细描述,给出的实施例仅为了阐明本发明,而不是为了限制本发明的范围。下述实施例中的实验方法,如无特殊说明,均为常规方法。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。以下实施例中的定量试验,均设置三次重复实验,结果取平均值。桂朝2号记载于如下文献中:ZhangX,ZhouSX,FuYC,etal.IdentificationofadroughttolerantintrogressionlinederivedfromDongxiangcommonwild rice(O.rufipogonGriff.).PlantMolBiol,2006,62:247~259,公众可以从中国农业大学获得。桂朝2号在下文中简称GC2。江西东乡野生稻记载于如下文献中:TianF,LiDJ,FuQ,ZhuZF,FuYC,WangXK,SunCQ.2006.Constructionofintrogressionlinescarryingwildrice(OryzarufipogonGriff.)segmentsincultivatedrice(O.sativaL.)backgroundandcharacterizationofintrogressedsegmentsassociatedwithyield-relatedtraits.TheoreticalandAppliedGenetics,112,570-80.公众可以从中国农业大学获得。东乡普通野生稻渗入系SIL40为GC2与江西东乡野生稻多次杂交和回交的后代,其记载于如下文献中:TianF,LiDJ,FuQ,ZhuZF,FuYC,WangXK,SunCQ.2006.Constructionofintrogressionlinescarryingwildrice(OryzarufipogonGriff.)segmentsincultivatedrice(O.sativaL.)backgroundandcharacterizationofintrogressedsegmentsassociatedwithyield-relatedtraits.TheoreticalandAppliedGenetics,112,570-80.公众可以从中国农业大学获得。东乡普通野生稻渗入系SIL40在下文中简称SIL40。ZH17记载于如下文献中:TanLB,LiXR,LiuFX,SunXY,LiCG,ZhuZF,FuYC,CaiHW,WangXK,XieDXandSunCQ.Controlofakeytransitionfromprostratetoerectgrowthinricedomestication.NatureGenetics,2008,40(11):1360-1364,公众可以从中国农业大学获得。下述实施例中的根癌农杆菌EHA105(文献中的名称为AgrobacteriumtumefaciensstrainEHA105)记载于如下文献中:GLUTELINPRECURSORACCUMULATION3encodesaregulatorofpost-Golgivesiculartrafficessentialforvacuolarproteinsortinginriceendosperm.PlantCell.2014Jan;26(1):410-25.公众可从中国农业大学获得,以重复本申请实验。实施例1、株型改变和/或产量改变的转基因水稻的获得及鉴定一、PROG2基因的发现将GC2与SIL40的表型进行比较及统计,实验结果见图1(a为分蘖期整体株型对比,b为灌浆期整体株型对比,c为主茎穗对比,d为分蘖数统计结果,e为分蘖角度统计结果,f为一次枝梗数比较,g为二次枝梗数比较,h为主茎穗粒数比较,i为单株产量比较,其中*表示P<0.05差异显著,**表示P<0.01差异极显著)。结果表明,与GC2相比,SIL40倾斜生长,分蘖数增加,分蘖角度增大,一次枝梗数减少,二次枝梗数减少、主茎穗粒数减少,单株产量减少。SIL40与轮回亲本GC2回交,构建次级分离群体。利用次级分离群体将PROG2基 因(序列表序列1所示)初步定位于水稻第7染色体短臂SW1与ID52标记之间。进一步,利用4000隐性纯合单株(直立生长),将PROG2基因精细定位于F43和ID52两标记之间(见图2),实际物理距离为9.2kb。PROG2基因的开放阅读框如序列表中序列1自5’末端起第125-691位所示,编码的蛋白命名为PROG2,其氨基酸序列如序列表中序列2所示,由188个氨基酸残基组成。二、PROG2基因功能验证1、重组载体和重组农杆菌的构建(1)以江西东乡野生稻2周苗为实验材料,提取基因组DNA为模板,采用引物D81-F:5′-CGGAATTCAAGCTTTGGAGAATAAGTCT-3′(下划线部分为限制性内切酶EcoRI识别位点)和引物D81-R:5′-TCCCCCGGGTCTAGAAAGGGATTTATCCC-3′(下划线部分为限制性内切酶SmaI识别位点)进行扩增,得到约4200bp的PCR产物。(2)用限制性内切酶EcoRI和SmaI双酶切步骤(1)获得的PCR产物,回收酶切产物。(3)用限制性内切酶EcoRI和SmaI双酶切植物表达载体pCAMBIA1300,回收约8.9kb载体骨架。(4)将步骤(2)的酶切产物和步骤(3)的载体骨架连接,得到重组质粒D81。根据测序结果,对步骤(4)获得的重组质粒D81进行结构描述如下:向植物表达载体pCAMBIA1300的EcoRI和SmaI酶切位点之间插入核苷酸序列是序列表中的序列3所示的DNA分子。重组质粒D81表达序列表中序列2所示的PROG2蛋白。重组质粒D81含有一个表达盒,所述表达盒包括PROG2启动子、PROG2蛋白的编码基因和PROG2终止子;所述PROG2启动子如序列3自5’末端起第1至1258位所示,所述PROG2蛋白的编码基因如序列3自5’末端起第1383至1949位所示,所述PROG2终止子如序列3自5’末端起第2151至4214位所示。将重组质粒D81导入根癌农杆菌EHA105中,得到含有重组质粒D81的重组农杆菌EHA105/D81。将空载体质粒pCAMBIA1300转入根癌农杆菌EHA105,得到含有质粒pCAMBIA1300的重组农杆菌EHA105/pCAMBIA1300。2、转PROG2基因水稻的获得(1)将水稻品种ZH17的种子脱壳灭菌,然后采用Hiei等的方法(HieiY,OhtaS,KomariT&KumashiroT.Efficienttransformationofrice(OryzasativaL.)mediatedbyAgrobacteriumandsequenceanalysisoftheboundariesoftheT-DNA.PlantJ.1994,6:271–282)用重组农杆菌EHA105/D81转化ZH17,获得T0代拟转PROG2基因水稻植株。(2)分别提取T0代拟转PROG2基因水稻植株的基因组DNA并作为模板,采用引物HPT-F:5'-TACTTCTACACAGCCATC-3'和引物HPT-R:5'-CGTCTGTCGAGAAGTTTC-3'扩增,如果得到947bp的扩增产物,则该T0代拟转PROG2基因水稻植株为T0代转PROG2基因阳性植株。按照上述(1)和(2)的方法,最终获得56株T0代转PROG2基因阳性植株。随机选择3个T0代转PROG2基因阳性植株的株系,分别命名为D81-1、D81-2和D81-3,进行下述实验。按照上述方法,将重组农杆菌EHA105/D81替换为重组农杆菌EHA105/pCAMBIA1300,其他步骤均相同,得到T0代转空载体阳性植株。3、转PROG2基因阳性植株的表型鉴定将水稻品种ZH17、T0代转空载体阳性植株和D81-1的种子分别种植在装有营养土和蛭石的盆中(营养土和蛭石体积比为1:1),25℃、光照交替培养,在生长发育过程中对水稻植株表型进行比较及统计。实验重复三次,每次重复30株。按照上述步骤,将D81-1分别替换为D81-2和D81-3,其它步骤均相同,得到各水稻植株表型的统计结果。实验结果见图3(a为分蘖期整体株型对比,b为灌浆期整体株型对比,c为主茎穗对比,d为株高统计结果,e为分蘖数统计结果,f为一次枝梗数比较,g为二次枝梗数比较,h为主茎穗粒数比较,i为单株产量比较,其中*表示P<0.05差异显著,**表示P<0.01差异极显著)。结果表明,T0代转PROG2基因阳性植株表现出类似SIL40的表型;与ZH17相比,T0代转PROG2基因阳性植株株高降低约四分之一,分蘖数增加约一倍,分蘖角度增大,一次枝梗数减少约四分之一,二次枝梗数减少约三分之二,主茎穗粒数减少约二分之一和单株产量降低约三分之一。T0代转空载体阳性植株与ZH17的株高、分蘖数、一次枝梗数、二次枝梗数、主茎穗粒数和单株产量均无显著差异。结果表明,PROG2基因对调控水稻株型和产量具有非常重要的作用。当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 蝎神经毒素AaIT及其编码基因与应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种花狭口蛙的基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种泽陆蛙基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 降低种子重量的白桦AP2基因及其编码蛋白的制造方法与工艺
- 可编码ArtRD蛋白基因及其蛋白和应用的制造方法与工艺
- 蝎神经毒素AaIT及其编码基因与应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种花狭口蛙的基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种泽陆蛙基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 蛋白质及其编码基因在调控植物抗黄萎病性中的应用的制造方法与工艺
- 甘薯耐逆相关蛋白IbFBA2、编码基因、重组载体与应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 核盘菌凝集素SSL‑6蛋白及其编码基因与应用的制造方法与工艺
- 植物抗病毒相关蛋白GmSN1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 蛋白质及其编码基因在调控植物抗黄萎病性中的应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 核盘菌凝集素SSL‑6蛋白及其编码基因与应用的制造方法与工艺