一种菌及其构建方法和应用与流程
本发明属于生物工程领域,涉及一种菌及其构建方法和应用。
背景技术:
石油基聚合物为人们的生产生活带来了极大的便利。但是,随着石化资源的日益减少,以及使用石化资源带来的环境问题,寻找新的环境友好的非石油基聚合物受到越来越多的重视。生物基材料具有易生物降解、原料来源广和易化学改良等优点,使其在很多领域中都有重要的应用。目前,聚对苯二甲酸丙二醇酯(PTT)和聚乳酸(PLA)是受关注度较高的两种生物基材料。PTT是一种性能优异的聚酯类新型纤维,由对苯二甲酸和1,3-丙二醇(1,3-PD)缩聚而成,它综合了聚对苯二甲酸乙二醇酯(PET)和聚对苯二甲酸丁二醇酯(PBT)的优势,具有很好的柔软性、蓬松性、抗污性、弹性和可以常温染色等特点,可以广泛用于服装、装饰和工程塑料等各个领域。PLA也称为聚丙交酯,是以乳酸(LAC)为主要原料聚合得到的聚合物,它具有较好的热稳定性、抗溶剂性和生物相容性,可进行多种方式的加工,可用于制造包装材料、纤维和非织造物等在服装、建筑、农业和医疗卫生等领域用途广泛。此外,PLA制品具有良好的生物可降解性,使用后能被自然界中微生物完全降解,是公认的环境友好材料。
生物基材料PTT和PLA巨大的市场需求,推动了生物法生产生物基材料单体1,3-PD和光学纯LAC的发展。目前生物法生产1,3-PD的方法主要分为两类:第一类是利用基因工程大肠杆菌,以葡萄糖等糖类为底物,生产1,3-PD(例如CN201110093628;ZL200710104008);第二类是利用肠道细菌,以甘油为底物,生产1,3-PD(ZL200410100479;CN201180064621)。最近,由于生物柴油产业的发展,甘油作为其副产物,价格逐渐下降,是1,3-PD生产的理想底物。可利用甘油为底物生产1,3-PD的菌株主要包括肺炎克雷伯氏菌(Klebsiella peneumoniae)、产酸克雷伯氏菌(Klebsiella oxytoca)、弗氏柠檬酸杆菌(Citrobacterfreundii)和丁酸梭菌(Clostridium butyricum)等。用于生产PLA的LAC必须具有高光学纯度,因此,研究者筛选得到了很多可生产光学纯LAC的细菌,例如鼠李糖乳杆菌(Lactobacillus rhamnosus)、凝结芽孢杆菌(Bacillus coagulans)和土芽孢乳杆菌(Sporolactobacillus terrae)等野生细菌,都可以以糖类为原料生产高浓度的光学纯L-乳酸(L-LAC)或D-乳酸(D-LAC)(ZL200710176057;ZL201210115365;CN201410022868)。
利用上述的菌株和生产方法,可实现1,3-PD和LAC中一种化合物的生物法生产,但是,目前尚无可同时生产1,3-PD和光学纯LAC(L-LAC或D-LAC)的菌株。肺炎克雷伯氏菌和产酸克雷伯氏菌在以甘油为底物,生产1,3-PD的过程中,还会生产少量的2,3-丁二醇、乙醇、LAC、琥珀酸、乙酸和甲酸等副产物,虽然这两种菌株发酵甘油的发酵液中,同时具有1,3-PD和LAC,但由于两种化合物的浓度差距大(一般不在同一数量级)导致其中一种化合物只能被当做副产物处理,且醇类产物和酸类产物种类多,成分复杂,难以分离得到高纯度的LAC,无法实现同时生产1,3-PD和LAC。在工业化大规模生产中,必须考虑产物浓度、转化率,提取成本等因素,如果能够利用一个菌株,以甘油为底物,以1,3-PD和光学纯LAC(L-LAC或D-LAC)为主要产物,高转化率的实现同时生产1,3-PD和光学纯LAC(L-LAC或D-LAC),将大大降低生产成本,提高生产效率。
技术实现要素:
针对现有技术的不足,本发明提供一种菌,所述菌具有联产1,3-丙二醇和L-乳酸的特性。联产是指同时生产。
进一步地,所述菌是由野生菌改造获得的人工菌。
进一步地,所述野生菌具有如下代谢途径:
1)甘油→1,3-丙二醇;
所述野生菌还具有如下代谢途径中一个或多个:
2)丙酮酸→α-乙酰乳酸,α-乙酰乳酸合成酶(budB)是催化该代谢途径的酶;
3)α-乙酰乳酸→乙偶姻,α-乙酰乳酸脱羧酶(budA)是催化该代谢途径的酶;
4)丙酮酸→乙酸,丙酮酸氧化酶(poxB)是催化该代谢途径的酶;
5)乙酰辅酶A→乙酰磷酸,乙酰磷酸转移酶(pta)是催化该代谢途径的酶;
6)乙酰磷酸→乙酸,乙酰激酶(ackA)是催化该代谢途径的酶;
7)乙酰辅酶A→乙醛,醛脱氢酶(adhE)是催化该代谢途径的酶;
8)富马酸→琥珀酸,富马酸还原酶(frdA)是催化该代谢途径的酶;
9)丙酮酸→D-乳酸,D-乳酸脱氢酶(dldh)是催化该代谢途径的酶;
所述改造包括:阻断所述代谢途径中的2)-9)中的一个或多个。
进一步地,所述野生菌为产酸克雷伯氏菌(Klebsiella oxytoca)。
进一步地,所述野生菌为产酸克雷伯氏菌(Klebsiella oxytoca)PDL-0,所述产酸克雷伯氏菌(Klebsiella oxytoca)PDL-0于2016年4月8日保藏于中国典型培养物保藏中心,保藏登记号为CCTCC M 2016184。
进一步地,所述改造包括:通过抑制或去除酶的活性来阻断所述代谢途径中的2)-9)中的一个或多个。
进一步地,所述改造包括:通过改变酶的基因来抑制或去除酶的活性。
进一步地,所述改造包括:通过基因重组的办法来改变酶的基因。
进一步地,编码α-乙酰乳酸脱羧酶基因的序列如SEQ ID NO:1所示;
编码α-乙酰乳酸合成酶基因的序列如SEQ ID NO:2所示;
编码醛脱氢酶基因的序列如SEQ ID NO:3所示;
编码乙酸激酶和乙酰磷酸转移酶基因的序列如SEQ ID NO:4所示;
编码丙酮酸氧化酶基因的序列如SEQ ID NO:5所示;
编码富马酸还原酶基因的序列如SEQ ID NO:6所示;
编码D-乳酸脱氢酶基因序列如SEQ ID NO:7所示。
进一步地,所述人工菌的budA、budB、adhE、ackA-pta、poxB、frdA和dldh中的一种基因或者多种基因缺陷。
进一步地,所述基因缺陷通过同源重组的方法产生。
进一步地,所述改造还包括外源引入L-乳酸合成途径。
进一步地,所述外源引入L-乳酸合成途径指:将凝结芽孢杆菌(Bacillus coagulans)2-6的L-乳酸脱氢酶基因lldh通过质粒转化到所述野生菌中,得到产L-乳酸的菌。
进一步地、如权利要求12所述的菌,其特征在于,所述外源引入L-乳酸合成途径指:将凝结芽孢杆菌(Bacillus coagulans)2-6的L-乳酸脱氢酶基因lldh替换所述野生菌中的budB基因,并通过质粒转化到所述野生菌中,得到产L-乳酸的菌。
进一步地,所述人工菌的基因型包括ΔbudA::lldh ΔbudB ΔadhE ΔackA-ptaΔpoxB ΔfrdA Δdldh。
进一步地,所述人工菌的基因型是ΔbudA::lldh ΔbudB ΔadhE ΔackA-ptaΔpoxB ΔfrdA Δdldh。
从另一角度描述,所述人工菌是由所述野生菌的budA、budB、adhE、ackA-pta、poxB、frdA和dldh基因缺陷,并外源引入了lldh基因获得;所述人工菌产1,3-PD和L-LAC,且1,3-PD和L-LAC的总转化率超过90%。总转化率计算公式为:总转化率=1,3-丙二醇的摩尔转化率+L-乳酸的摩尔转化率;其中,1,3-丙二醇的摩尔转化率=(1,3-PD的最终浓度×最终发酵液体积×甘油的摩尔质量92)/(消耗的甘油质量×1,3-PD的摩尔质量76);L-乳酸的摩尔转化率=(L-LAC的最终浓度×最终发酵液体积×甘油的摩尔质量92)/(消耗的甘油质量×L-LAC的摩尔质量90)。
进一步地,budA、budB、adhE、ackA-pta、poxB、frdA和dldh基因缺陷的获得采用同源重组的方法,且各基因缺陷的获得不存在固定的先后顺序;所述同源重组的方法是指通过PCR扩增以上所述基因的上游同源片段和下游同源片段,并将所述上游同源片段和所述下游同源片段构建到自杀质粒中转化至大肠杆菌(Escherichia coli)中,得到供体菌;将所述供体菌与相应的受体菌进行双亲本杂交,使所述上游同源片段和所述下游同源片段与所述受体菌的基因组发生同源重组,从而获得以上所述基因缺陷的菌株,即所述人工菌。
进一步地,所述野生菌是产酸克雷伯氏菌(Klebsiella oxytoca)PDL-0,所述产酸克雷伯氏菌(Klebsiella oxytoca)PDL-0于2016年4月8日保藏于中国典型培养物保藏中心,保藏登记号为CCTCC M 2016184。
进一步地,所述budA的DNA序列如SEQ ID NO:1所示;和/或所述budB的DNA序列如SEQ ID NO:2所示;和/或所述adhE的DNA序列如SEQ ID NO:3所示;和/或所述ackA-pta的DNA序列如SEQ ID NO:4所示;和/或所述poxB的DNA序列如SEQ ID NO:5所示;和/或所述frdA的DNA序列如SEQ ID NO:6所示;和/或所述dldh的DNA序列如SEQ ID NO:7所示;和/或所述lldh的DNA序列如SEQ ID NO:8所示。
在一较佳实施方式中,所述改造包括向所述野生菌中引入外源1,3-PD合成途径和/或L-LAC合成途径。
进一步地,所述引入外源1,3-PD合成途径是将甘油脱水酶编码基因dhaB及1,3-PD氧化还原酶编码基因dhaT转化到所述野生菌中。
进一步地,所述引入外源L-LAC合成途径是将L-乳酸脱氢酶基因lldh通过质粒转化到所述野生菌中,得到产L-乳酸的菌。
进一步地,所述外源引入的L-乳酸脱氢酶基因lldh来自于凝集芽孢杆菌(Bacillus coagulans)2-6。凝集芽孢杆菌(Bacillus coagulans)2-6为CN101173242A中的凝结芽孢杆菌(Bacillus coagulans)CASH,其保藏号为CGMCC No2184。
进一步地,所述改造还包括:通过基因重组的办法来改变budA、budB、adhE、ackA-pta、poxB、frdA和/或dldh基因。
在一个较佳实施方式中,所述菌是产酸克雷伯氏菌(Klebsiella oxytoca)PLL,所述产酸克雷伯氏菌(Klebsiella oxytoca)PLL于2016年4月8日保藏于中国典型培养物保藏中心,保藏登记号为CCTCC M 2016186。
本发明提供的菌具有如下代谢途径:
1)甘油→1,3-丙二醇;
2)甘油→丙酮酸→L-乳酸。
本发明提供的一种菌具有联产1,3-丙二醇和L-乳酸的特性,其中联产得到的:1,3-丙二醇的摩尔转化率≥35%;L-乳酸的摩尔转化率≥40%。
进一步地,联产得到的:1,3-丙二醇的摩尔转化率≥40%;和/或L-乳酸的摩尔转化率≥50%。
本发明提供的一种菌具有联产1,3-丙二醇和L-乳酸的特性,其中联产得到的:1,3-丙二醇和L-乳酸的总转化率超过90%。
本发明提供的一种菌具有联产1,3-丙二醇和L-乳酸的特性,其中联产得到的1,3-丙二醇和L-乳酸的质量比为1:0.1-10。进一步地,联产得到的1,3-丙二醇和L-乳酸的质量比为1:0.2-5。进一步地,联产得到的1,3-丙二醇和L-乳酸的质量比为1:0.5-2。
进一步地,本发明提供的所述菌来自于真菌中的曲霉属、酵母属、接合酵母属、毕赤酵母属、克鲁维酵母属、假丝酵母属、汉逊酵母属、德巴利酵母属、毛霉属、球拟酵母属或链霉菌属;或者来自细菌中的甲基菌属、沙门氏菌属、芽孢杆菌属、假单胞菌属、克雷伯氏菌、乳杆菌属、肠杆菌属、柠檬酸杆菌属、暗杆菌属、泥杆菌属或者梭菌属。
本发明还公开了一种如上所述的菌的构建方法,包括以下步骤:
步骤一、从土壤样品中筛选能生产1,3-丙二醇和乳酸的产酸克雷伯氏菌;
步骤二、使得步骤一中得到的产酸克雷伯氏菌的α-乙酰乳酸脱羧酶基因、α-乙酰乳酸合成酶基因、醛脱氢酶基因、乙酸激酶和乙酰磷酸转移酶基因、丙酮酸氧化酶基因、富马酸还原酶基因和D-乳酸脱氢酶基因缺陷,得到过程菌株一;
步骤三、将凝结芽孢杆菌(Bacillus coagulans)2-6的L-乳酸脱氢酶基因插入到所述步骤二中得到的过程菌株一的α-乙酰乳酸脱羧酶基因在基因组中上游的启动子之后,得到具有L-乳酸脱氢酶活性的菌株,即所述菌。
进一步地,所述步骤一中得到的产酸克雷伯氏菌是产酸克雷伯氏菌(Klebsiella oxytoca)PDL-0,所述产酸克雷伯氏菌(Klebsiella oxytoca)PDL-0于2016年4月8日保藏于中国典型培养物保藏中心,保藏登记号为CCTCC M2016184。
进一步地,所述budA的DNA序列如SEQ ID NO:1所示;所述budB的DNA序列如SEQ ID NO:2所示;所述adhE的DNA序列如SEQ ID NO:3所示;所述ackA-pta的DNA序列如SEQ ID NO:4所示;所述poxB的DNA序列如SEQ ID NO:5所示;所述frdA的DNA序列如SEQ ID NO:6所示;所述dldh的DNA序列如SEQ ID NO:7所示;所述lldh的DNA序列如SEQ ID NO:8所示。
进一步地,所述步骤二包括以下步骤:
步骤2-1、使所述产酸克雷伯氏菌PDL-0 CCTCC M 2016184的α-乙酰乳酸脱羧酶基因和α-乙酰乳酸合成酶基因缺陷,得到α-乙酰乳酸脱羧酶和α-乙酰乳酸合成酶活性丢失的菌株,命名为产酸克雷伯氏菌PDL-1;
步骤2-2、使所述产酸克雷伯氏菌PDL-1的醛脱氢酶基因缺陷,得到醛脱氢酶活性丢失的菌株,命名为产酸克雷伯氏菌PDL-2;
步骤2-3、使所述产酸克雷伯氏菌PDL-2的乙酸激酶和乙酰磷酸转移酶基因缺陷,得到乙酸激酶和乙酰磷酸转移酶活性丢失的菌株,命名为产酸克雷伯氏菌PDL-3;
步骤2-4、使所述产酸克雷伯氏菌PDL-3的丙酮酸氧化酶基因缺陷,得到丙酮酸氧化酶活性丢失的菌株,命名为产酸克雷伯氏菌PDL-4;
步骤2-5、使所述产酸克雷伯氏菌PDL-4的富马酸还原酶基因缺陷,得到富马酸还原酶活性丢失的菌株,命名为产酸克雷伯氏菌PDL-5;
步骤2-6、使所述产酸克雷伯氏菌PDL-5的D-乳酸脱氢酶基因缺陷,得到D-乳酸脱氢酶基因活性丢失的菌株,命名为产酸克雷伯氏菌PDL-6;
经过以上步骤后得到了α-乙酰乳酸脱羧酶基因、α-乙酰乳酸合成酶基因、醛脱氢酶基因、乙酸激酶基因、乙酰磷酸转移酶基因、丙酮酸氧化酶基因、富马酸还原酶基因和D-乳酸脱氢酶基因缺陷的所述过程菌株一;
所述α-乙酰乳酸脱羧酶基因序列如SEQ ID NO:1所示;所述α-乙酰乳酸合成酶基因序列如SEQ ID NO:2所示;所述醛脱氢酶基因序列如SEQ ID NO:3所示;所述乙酸激酶和乙酰磷酸转移酶基因序列如SEQ ID NO:4所示;所述丙酮酸氧化酶基因序列如SEQ ID NO:5所示;所述富马酸还原酶基因序列如SEQ ID NO:6所示;所述D-乳酸脱氢酶基因序列如SEQ ID NO:7所示。
进一步地,所述步骤三中的凝结芽孢杆菌2-6的L-乳酸脱氢酶基因序列如SEQ ID NO:8所示。
进一步地,所述步骤二中,使得产酸克雷伯氏菌PDL-0 CCTCC M 2016184的α-乙酰乳酸脱羧酶基因、α-乙酰乳酸合成酶基因、醛脱氢酶基因、乙酸激酶和乙酰磷酸转移酶基因、丙酮酸氧化酶基因、富马酸还原酶基因和D-乳酸脱氢酶基因缺陷是通过PCR扩增以上所述基因的上游同源片段和下游同源片段,并将所述上游同源片段和所述下游同源片段构建到自杀质粒pKR6K上,并转化至大肠杆菌(Escherichia coli)S17-1(λpir)中,得到供体菌;将所述供体菌与相应的受体菌进行双亲本杂交,使所述上游同源片段和所述下游同源片段与所述受体菌的基因组发生同源重组,从而获得以上所述基因缺陷的菌株;所述受体菌包括所述产酸克雷伯氏菌PDL-0 CCTCC M 2016184、所述产酸克雷伯氏菌PDL-1、所述产酸克雷伯氏菌PDL-2、所述产酸克雷伯氏菌PDL-3、所述产酸克雷伯氏菌PDL-4和所述产酸克雷伯氏菌PDL-5;
所述步骤三中,将凝结芽孢杆菌(Bacillus coagulans)2-6的L-乳酸脱氢酶基因lldh插入到所述产酸克雷伯氏菌PDL-6的α-乙酰乳酸脱羧酶基因在基因组中上游的启动子之后,是通过PCR扩增所述α-乙酰乳酸脱羧酶基因在基因组中上游同源片段和下游同源片段,以及凝结芽孢杆菌2-6的L-乳酸脱氢酶基因片段后,构建到自杀质粒pKR6K上,并转化至大肠杆菌S17-1(λpir)中,得到带有pKR6K-ΔbudA::lldh的供体菌;将所述带有pKR6K-ΔbudA::lldh的供体菌与所述产酸克雷伯氏菌PDL-6进行双亲本杂交,得到所述菌。
本发明还提供另一种如上所述的菌的构建方法,其特征在于,通过基因工程改造向菌株中引入外源1,3-PD合成途径和/或外源L-LAC合成途径。
进一步地,所述引入外源1,3-PD合成途径指:将甘油脱水酶编码基因dhaB及1,3-PD氧化还原酶编码基因dhaT通过质粒转化到所述菌株中,得到菌株A,所述菌株A产1,3-PD。
进一步地,所述引入外源1,3-PD合成途径指:选用产酸克雷伯氏菌中的1,3-PD合成途径中的甘油脱水酶编码基因dhaB及1,3-PD氧化还原酶编码基因dhaT,进行PCR克隆后连接到质粒DNA pet-Duet上,并转化到所述菌株中,得到菌株A,所述菌株A产1,3-PD。
进一步地,所述引入外源L-LAC合成途径指:将L-乳酸脱氢酶基因lldh通过质粒转化到所述菌株中,得到菌株B,所述菌株B产L-LAC。
进一步地,所述引入外源L-LAC合成途径指:选用来自于凝结芽孢杆菌(Bacillus coagulans)2-6的L-乳酸脱氢酶基因lldh通过质粒转化到所述菌株中,得到菌株B,所述菌株B产L-LAC。
进一步地,还包括如下操作:向引入外源1,3-PD合成途径和/或引入外源L-LAC合成途径的菌株,采用同源重组的方法产生budA、budB、adhE、ackA-pta、poxB、frdA和/或dldh基因缺陷的菌株。
本发明还涉及如上所述的菌在生产1,3-丙二醇或L-乳酸或联产1,3-丙二醇和L-乳酸中的应用。
尤其是产酸克雷伯氏菌(Klebsiella oxytoca)PLL CCTCC M 2016186在生产1,3-丙二醇或L-乳酸或联产1,3-丙二醇和L-乳酸中的应用。
本发明提供一种通过发酵如上所述的菌联产1,3-丙二醇和L-乳酸的方法。通过发酵如上所述的菌联产1,3-丙二醇和L-乳酸的方法,实际上是指如上所述的菌的应用。
进一步地,如上所述的联产1,3-丙二醇和L-乳酸的方法,其特征在于,所述菌是产酸克雷伯氏菌(Klebsiella oxytoca)PLL CCTCC M 2016186。
进一步地,发酵时通气量为0-2.0vvm。进一步地,发酵时通气量为1.0vvm。
进一步地,发酵时采用氢氧化钙和水的混合乳液作为中和剂调节发酵液的pH。
进一步地,包括以下步骤:
步骤(1)、菌株选择:选用产酸克雷伯氏菌(Klebsiella oxytoca)PLL CCTCC M 2016186;
步骤(2)、种子培养;
步骤(3)、发酵:发酵过程中,使用中和剂调节发酵液pH为5.5-7.5。
进一步地,包括以下步骤:
步骤(1)、菌株选择:选用产酸克雷伯氏菌(Klebsiella oxytoca)PLL CCTCC M 2016186;
步骤(2)、种子培养:选用所述步骤(1)的菌株,在无菌条件下接种至甘油培养基中,培养温度为25-40℃,摇床震荡转速为100-300rpm,培养时间为6-24小时,制得种子培养液;
步骤(3)、发酵:将所述步骤(2)中制得的种子培养液接种至装有甘油培养基的发酵罐中,接种量v/v为0.5-10%,发酵温度为25-40℃,通气量为0.3-2.0vvm,搅拌转速为50-400rpm,发酵过程中,使用中和剂调节发酵液pH为5.5-7.5;发酵方式为分批发酵或分批补料发酵,进行分批发酵时,当甘油培养基中的甘油耗尽时,停止发酵;进行分批补料发酵时,当甘油培养基中的甘油耗尽时,通过向发酵罐中补加400-800g/L的甘油溶液,控制发酵液中甘油浓度为5-40g/L,当发酵液中1,3-丙二醇或L-乳酸浓度不再升高时,停止发酵。
进一步地,所述步骤(2)中培养温度为30-37℃,摇床震荡转速为150-250rpm,培养时间为10-16小时;所述步骤(3)中接种量v/v为2-6%,发酵温度为30-37℃,通气量为0.7-1.5vvm,搅拌转速为150-300rpm,调节发酵液pH为6.0-7.0,发酵方式为分批补料发酵,补加的甘油溶液中甘油浓度为500-700g/L,控制发酵液中甘油浓度为10-30g/L;所述步骤(3)中的中和剂包括氢氧化钠水溶液、氢氧化钾水溶液、氨水水溶液,以及氢氧化钙和水的混合乳液中的任一一种或者多种。
本发明的有益效果包括:
1、使用产酸克雷伯氏菌PLL CCTCC M 2016186,得到的发酵液中目标产物浓度高,副产物种类少且浓度低。可生产浓度超过70.7g/L的1,3-PD和浓度超过95.4g/L的L-LAC,1,3-PD和L-LAC的总转化率超过90%,生产的L-LAC具有高光学纯度(纯度大于99.9%)。发酵液中仅能检测到少量的副产物。
联产1,3-PD和光学纯LAC(L-LAC或D-LAC)的好处是:由于克雷伯氏菌代谢甘油属于甘油歧化途径,分为甘油氧化途径和甘油还原途径。甘油到1,3-丙二醇的转化属于甘油的还原途径,转化过程需要氧化途径中产生的辅因子NADH和ATP来提供还原力及能量。乳酸在2004年被美国能源部评选为前三十种重点发展的平台化合物之一,鉴于其广阔的用途,在2010年又被选为最有前途的平台化合物之一。乳酸是一种重要的单体,可以用来合成可生物降解、可生物相容且环境友好的生物多聚体-聚乳酸(PLA)。除了乳酸的经济价值外,合成乳酸的过程中净产1mol NADH,可以为合成1,3-PD提供还原力,从而达到还原力平衡的状态;从甘油到乳酸的转化过程中,是三碳到三碳的转化,没有碳损失,可以保证碳的回复率;另外,乳酸具有非常好的生物相容性,这对工业上高产乳酸至关重要;由于乳酸的生物合成已经实现了工业级别的生产,因此,其下游纯化技术也相对成熟。这一切的特征都使得乳酸成为与1,3-PD联产的最佳选择。而事实也证明联产1,3-PD和L-LAC获得了非常高的总转化率(超过90%)。根据现有报道(Biotechnol Bioeng.Metabolism in 1,3-propanediol fed-batch fermentation by a D-lactate deficient mutant of Klebsiella pneumoniae.2009;104(5):965-72.;Bioresour Technol.Production of optically pure d-lactate from glycerol by engineered Klebsiella pneumoniae strain.2014;172:269-75.;Hong A A,Cheng K K et al.Strain isolation and optimization of process parameters for bioconversion of glycerol to lactic acid[J].Journal of chemical technology and biotechnology,2009,84(10):1576-1581.),在克雷伯氏菌中,由甘油直接转化合成1,3-PD的最高产量为95.4g/L,从甘油到1,3-PD的摩尔转化率为0.48mol/mol(即48%);从甘油直接转化L-乳酸的最高产量为85.4g/L,转化率为0.9mol/mol(即90%),生产强度为0.97g/Lh-1。但是根据现在已有报道,尚无可同时生产1,3-PD和光学纯LAC(L-LAC或D-LAC)的菌株。
由于在工业化大规模生产中,必须考虑产物浓度、转化率,提取成本等因素,如果能够利用一个菌株,以甘油为底物,以1,3-PD和光学纯LAC(L-LAC或D-LAC)为主要产物,高摩尔转化率的实现同时生产1,3-PD和光学纯LAC(L-LAC或D-LAC),将大大降低生产成本,提高生产效率。
2、产酸克雷伯氏菌PLL CCTCC M 2016186易于离心和过滤,有利于生物材料单体1,3-PD和L-LAC的高效生物法生产和提取。
3、产酸克雷伯氏菌PLL CCTCC M 2016186,具有重要的实际应用价值。
4、产酸克雷伯氏菌PLL联产得到的1,3-PD和L-LAC的浓度差距不大,其质量比都在1:0.1-10范围内,尤其是在1:0.2-5范围内,进一步地在1:0.5-2范围内。
附图说明
图1是对产酸克雷伯氏菌(Klebsiella oxytoca)进行基因改造的示意图。
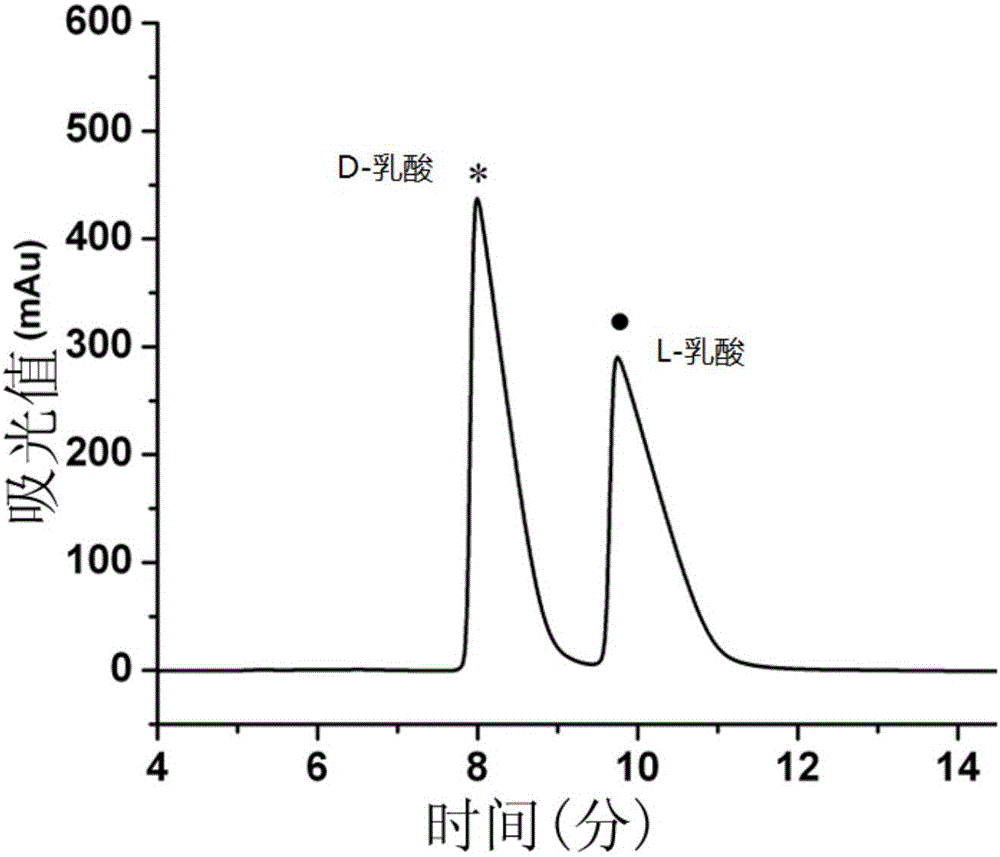
图2是D-乳酸和L-乳酸的标准品混合物检测图。
图3是产酸克雷伯氏菌PLL CCTCC M 2016186的发酵产物乳酸手性分析检测图。
具体实施方式
下面结合实施例对本发明的技术内容做进一步的说明:下述实施例是说明性的,不是限定性的,不能以下述实施例来限定本发明的保护范围。下述实施例中所使用的实验方法如无特殊说明,均为常规方法。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
本发明所述的基因工程产酸克雷伯氏菌PLL CCTCC M 2016186,由于budA、budB、adhE、ackA-pta、poxB、frdA和dldh基因缺陷,导致α-乙酰乳酸脱羧酶、α-乙酰乳酸合成酶、醛脱氢酶、乙酸激酶、乙酰磷酸转移酶、丙酮酸氧化酶、富马酸还原酶和D-乳酸脱氢酶活性丢失,因此2,3-丁二醇、乙醇、乙酸、琥珀酸和D-LAC代谢途径失活;另外,凝结芽孢杆菌2-6的lldh基因插入到budA基因在基因组中上游的启动子之后,使菌株具有了L-乳酸脱氢酶的活性,如图1所示。从图1中可以看出,甘油可转化得到1,3-丙二醇和L-乳酸,L-乳酸脱氢酶催化丙酮酸生成L-乳酸,并且以下代谢途径失活:α-乙酰乳酸合成酶催化丙酮酸生成α-乙酰乳酸;α-乙酰乳酸脱羧酶催化α-乙酰乳酸生成乙偶姻;丙酮酸氧化酶催化丙酮酸生成乙酸;乙酰磷酸转移酶催化乙酰辅酶A生成乙酰磷酸;乙酰激酶催化乙酰磷酸生成乙酸;醛脱氢酶催化乙酰辅酶A生成乙醛;富马酸还原酶催化富马酸生成琥珀酸;D-乳酸脱氢酶催化丙酮酸生成D-乳酸。在图1中,差号表示该酶活性丢失,相应的代谢途径失活。
由于α-乙酰乳酸脱羧酶、α-乙酰乳酸合成酶、醛脱氢酶、乙酸激酶、乙酰磷酸转移酶、丙酮酸氧化酶、富马酸还原酶和D-乳酸脱氢酶在产酸克雷伯氏菌中不利于甘油向1,3-PD和L-LAC转化,因此当这些酶功能缺陷后,则有利于产酸克雷伯氏菌以甘油为底物生产1,3-PD和L-LAC。同时,插入L-乳酸脱氢酶基因到该基因工程产酸克雷伯氏菌中,也有利于丙酮酸向L-LAC转化,见图1。本发明正是在这样的基因改造思路下进行相应的基因工程菌株构建,得到了高产1,3-PD和L-LAC、易纯化的上述基因型的基因工程产酸克雷伯氏菌。以下将以具体的菌株对其有益效果进行说明和验证,但并不表示只有下述具体菌株才具有高产1,3-PD和L-LAC、易纯化的有益效果,理论上应该是凡是具备该基因型的产酸克雷伯氏菌均有这样的有益效果。
如图2和3所示,使用产酸克雷伯氏菌PLL CCTCC M 2016186,以甘油为底物,可生产高浓度的1,3-PD和高浓度的L-LAC。发酵液中检测不到D-LAC,因此,生产的L-LAC具有高光学纯度(纯度大于99.9%)。发酵液中检测不到2,3-丁二醇、乙醇和甲酸,仅能检测到少量的乙酸和琥珀酸,发酵液中产物检测结果如表1所示。
表1:产酸克雷伯氏菌PLL CCTCC M 2016186以甘油为底物发酵液中产物组成
如表1所示,产酸克雷伯氏菌PLL CCTCC M 2016186,以甘油为底物,可生产70.0g/L的1,3-PD和104.0g/L的L-LAC,其中,1,3-PD的摩尔转化率达到42.5%,L-LAC的摩尔转化率达到53.4%,两种主要产物1,3-PD和L-LAC的总转化率超过90%。发酵液中仅能检测到少量的副产物。
实施例1:以1,3-PD和LAC为主要产物的菌株的筛选和鉴定
称取2g土壤样品加入50mL甘油液体培养基中,置于摇床中37℃培养24小时,摇床转速为200rpm。然后用无菌的生理盐水稀释培养液,分别稀释10倍、100倍、1000倍和10000倍后涂布到含甘油固体培养基的培养皿中,37℃静置培养24小时。待长出单菌落后,挑选菌落面积和产酸透明圈面积大的菌落,接种到甘油液体培养基中,置于摇床中37℃培养24小时,摇床转速为200rpm。对培养液进行离心,测定培养液中1,3-PD和LAC的产量,挑取一株1,3-PD和LAC产量高,同时易于离心的菌株。
将上述菌株多次在甘油固体培养基上划线分离纯化,然后再进行10个循环的培养测试,10次循环培养产生的1,3-PD和LAC产量和转化率基本保持原有水平,证明上述菌株即是目标菌株,将该菌株命名为PDL-0。测定菌株PDL-0培养液中D-LAC和L-LAC的比例,结果表明,菌株PDL-0生产的LAC中,D-LAC的比例大于99.9%,L-LAC的比例小于0.01%。
抽提菌株PDL-0的全基因组,然后PCR扩增菌株PDL-0的16S rRNA的基因序列,将PCR产物测序,测序得到的16S rRNA基因序列如SEQ ID NO:9所示。菌株PDL-0的16S rRNA基因序列与NCBI数据库(http://www.ncbi.nlm.nih.gov/)中其他产酸克雷伯氏菌的16S rRNA基因序列具有99%的同源性,分析结果表明,菌株PDL-0为产酸克雷伯氏菌(Klebsiella oxytoca)。
其中,所述甘油液体培养基的配方为:酵母粉3g/L、K2HPO4·3H2O 10g/L、KH2PO42g/L、NH4Cl 1g/L、MgSO4·7H2O 0.5g/L、FeCl3·6H2O 20mg/L、CoCl2·6H2O 50mg/L和甘油20g/L;121℃灭菌20分钟。
其中,所述甘油固体培养基的配方为:酵母粉3g/L、K2HPO4·3H2O 10g/L、KH2PO42g/L、NH4Cl 1g/L、MgSO4·7H2O 0.5g/L、FeCl3·6H2O 20mg/L、CoCl2·6H2O 50mg/L,甘油20g/L和琼脂粉15g/L;121℃灭菌20分钟。
实施例2:产酸克雷伯氏菌PDL-0中α-乙酰乳酸脱羧酶基因(budA)和α-乙酰乳酸合成酶基因(budB)的缺陷
(1)用于在产酸克雷伯氏菌PDL-0中部分缺失budA基因的载体的构建
根据budA基因序列(如SEQ ID NO:1所示)设计引物,PCR扩增budA基因的上游和下游同源片段。以产酸克雷伯氏菌PDL-0的基因组DNA为模板,使用引物budA-1:5’-ACATGATTACGAATTCATGAACCATTCTGCTGAATG-3’(如SEQ ID NO:10所示)和引物budA-2:5’-AACGGGCTGGCATCACCGCGAAGGGCGTGC-3’(如SEQ ID NO:11所示)进行PCR扩增,得到上游同源片段;使用引物budA-3:5’-CGCGGTGATGCCAGCCCGTTTTCCGCTTCA-3’(如SEQ ID NO:12所示)和引物budA-4:5’-TACCGAGCTCGAATTCTTAGTTTTCGACTGAGCGAA-3’(如SEQ ID NO:13所示)进行PCR扩增,得到下游同源片段。PCR扩增条件为:95℃5分钟;95℃30秒,60℃30秒,72℃1分钟,共30个循环;72℃5分钟。PCR反应结束后,将PCR扩增产物进行1.0%琼脂糖凝胶电泳,回收并纯化,得到上游和下游同源片段。
使用限制性内切酶EcoRI酶切自杀质粒pKR6K(Wang et al.,J.Biol.Chem.2014,289:6080-6090),将酶切产物进行1.0%琼脂糖凝胶电泳,回收并纯化,得到线性化的质粒pKR6K。自杀质粒pKR6K可通过将质粒pK18mobsacB(优宝生物公司)的复制子替换成质粒pCAM140的复制子获得。质粒pCAM140的复制子的序列可通过基因合成的方式获得,质粒pCAM140的序列见文献报道(Wilson K J,Sessitsch A,Corbo J C,et al.β-Glucuronidase(GUS)transposons for ecological and genetic studies of rhizobia and other Gram-negative bacteria[J].Microbiology,1995,141(7):1691-1705.)。
使用无缝克隆和组装试剂盒(pEASY-Uni Seamless Cloning and Assembly Kit,北京全式金生物技术有限公司),将上游同源片段、下游同源片段和线性化的质粒pKR6K连接,得到可部分缺失budA基因的自杀质粒pKR6K-ΔbudA。
(2)budA基因部分缺失的产酸克雷伯氏菌的构建
将pKR6K-ΔbudA转化至大肠杆菌S17-1(λpir)(北京全式金生物技术有限公司)中,得到供体菌大肠杆菌S17-1(λpir)(pKR6K-ΔbudA)。将供体菌大肠杆菌S17-1(λpir)(pKR6K-ΔbudA)与受体菌产酸克雷伯氏菌PDL-0进行双亲本杂交,使pKR6K-ΔbudA上的budA基因上游同源片段和下游同源片段与产酸克雷伯氏菌PDL-0的基因组发生同源重组,从而使产酸克雷伯氏菌PDL-0的budA基因缺失200bp,达到使budA基因缺陷的目的,具体方法为:
a.分别接种活化后的供体菌和受体菌至5mL LB液体培养基中,37℃摇床中,200rpm培养2-3小时,当供体菌和受体菌同时生长到OD620nm为0.5-0.8;将5mL供体菌菌液离心,无菌生理盐水洗两遍;将1mL受体菌菌液离心,无菌生理盐水洗两遍;将上述供体菌和受体菌的菌体一共用100μL无菌生理盐水重悬,将重悬液全部滴在LB固体培养基平板中间,平板正面放置,37℃培养12-18小时。
b.将步骤a中LB固体培养基平板上的菌落用无菌生理盐水和刮刀刮下,无菌生理盐水洗两遍,进行适当稀释,涂布于加入了50μg/mL卡那霉素的M9固体培养基平板,37℃培养24-36小时。
c.挑取步骤b中M9固体培养基平板上生长的单菌落至加入50μg/mL卡那霉素的5mL LB液体培养基中,37℃,200rpm培养12小时。转接菌液至新鲜的5mL LB液体培养基中(不添加卡那霉素),37℃,200rpm培养12小时。
d.将上述菌液适当稀释,涂布于LAS固体培养基平板,25℃培养24小时。
e.挑取步骤d中LAS固体培养基平板上生长的单菌落至5mL LB液体培养基中,37℃,200rpm培养12小时,提取基因组DNA,使用引物budA-1:5’-ACATGATTACGAATTCATGAACCATTCTGCTGAATG-3’(如SEQ ID NO:10所示)和引物budA-4:5’-TACCGAGCTCGAATTCTTAGTTTTCGACTGAGCGAA-3’(如SEQ ID NO:13所示)进行PCR验证。获得的budA基因缺陷的菌株。
其中,所述LB液体培养基配方为:蛋白胨10g/L、酵母粉5g/L和NaCl10g/L,121℃灭菌20分钟。
其中,所述LB固体培养基配方为:蛋白胨10g/L、酵母粉5g/L、NaCl 10g/L和琼脂粉15g/L,121℃灭菌20分钟。
其中,所述M9固体培养基配方为:Na2HPO4·12H2O 1.7g/L、KH2PO40.3g/L、NaCl 0.05g/L、NH4Cl 0.1g/L、柠檬酸三钠0.5g/L和琼脂粉15g/L,121℃灭菌20分钟。
其中,所述LAS固体培养基配方为:蛋白胨10g/L、酵母粉5g/L、蔗糖150g/L和琼脂粉15g/L,115℃灭菌20分钟。
(3)用于在budA基因缺陷的菌株中部分缺失budB基因的载体的构建
根据budB基因序列(如SEQ ID NO:2所示)设计引物,PCR扩增budB基因的上游和下游同源片段。以产酸克雷伯氏菌PDL-0的基因组DNA为模板,使用引物budB-1:5’-ACGCGAATTCGTGGATAATCAACATCAACCGCGCC-3’(如SEQ ID NO:14所示)和引物budB-2:5’-ACGCGGATCCGGGGCGTCCCTGCTCGGC-3’(如SEQ ID NO:15所示)进行PCR扩增,得到上游同源片段;使用引物budB-3:5’-ACGCGGATCCATCGCCCGCTATCTCTACAGCTTCC-3’(如SEQ ID NO:16所示)和引物budB-4:5’-ACGCCTGCAGATTTGACTGAGATGAAGCTGGCCCA-3’(如SEQ ID NO:17所示)进行PCR扩增,得到下游同源片段。PCR扩增条件为:95℃5分钟;95℃30秒,60℃30秒,72℃1分钟,共30个循环;72℃5分钟。PCR反应结束后,将PCR扩增产物进行1.0%琼脂糖凝胶电泳,回收并纯化,得到上游和下游同源片段。
使用限制性内切酶EcoRI和BamHI酶切上游同源片段,使用限制性内切酶BamHI和PstI酶切下游同源片段,使用限制性内切酶EcoRI和PstI酶切自杀质粒pKR6K(Wang et al.,J.Biol.Chem.2014,289:6080-6090),将酶切产物进行1.0%琼脂糖凝胶电泳,回收并纯化,得到线性化的质粒pKR6K和带有粘性末端的上游和下游同源臂片段。
使用T4连接酶(NEB公司)将带有粘性末端的上游同源片段、下游同源片段和线性化的质粒pKR6K连接,得到可部分缺失budB基因的自杀质粒pKR6K-ΔbudB。
(4)budB基因部分缺失的产酸克雷伯氏菌的构建
将pKR6K-ΔbudB转化至大肠杆菌S17-1(λpir)中,得到供体菌大肠杆菌S17-1(λpir)(pKR6K-ΔbudB)。将供体菌大肠杆菌S17-1(λpir)(pKR6K-ΔbudB)与受体菌budA基因缺陷的菌株进行双亲本杂交,使pKR6K-ΔbudB上的budB基因上游同源片段和下游同源片段与budA基因缺陷的菌株的基因组发生同源重组,从而budA基因缺陷的菌株的budB基因缺失722bp,达到使budB基因缺陷的目的。具体方法与步骤(2)相同,只是PCR验证时,使用引物budB-1:5’-ACGCGAATTCGTGGATAATCAACATCAACCGCGCC-3’(如SEQ ID NO:14所示)和引物budB-4:5’-ACGCCTGCAGATTTGACTGAGATGAAGCTGGCCCA-3’(如SEQ ID NO:17所示)进行PCR验证。获得的budA基因和budB基因缺陷的菌株,命名为产酸克雷伯氏菌PDL-1。
实施例3:产酸克雷伯氏菌PDL-1中醛脱氢酶基因(adhE)的缺陷
(1)用于在产酸克雷伯氏菌PDL-1中部分缺失adhE基因的载体的构建
根据adhE基因序列(如SEQ ID NO:3所示)设计引物,PCR扩增adhE基因的上游和下游同源片段。以产酸克雷伯氏菌PDL-1的基因组DNA为模板,使用引物adhE-1:5’-ACATGATTACGAATTCATGGCTGTTACTAATGTCGC-3’(如SEQ ID NO:18所示)和引物adhE-2:5’-TGCTGTCTGTTGGCGTTACGGGTCTTCAGG-3’(如SEQ ID NO:19所示)进行PCR扩增,得到上游同源片段;使用引物adhE-3:5’-CGTAACGCCAACAGACAGCATTCAGCCAGT-3’(如SEQ ID NO:20所示)和引物adhE-4:5’-TACCGAGCTCGAATTCTTAAGCGGATTTTTTCGCTT-3’(如SEQ ID NO:21所示)进行PCR扩增,得到下游同源片段。PCR扩增条件为:95℃5分钟;95℃30秒,60℃30秒,72℃1分钟,共30个循环;72℃5分钟。PCR反应结束后,将PCR扩增产物进行1.0%琼脂糖凝胶电泳,回收并纯化,得到上游和下游同源片段。
使用限制性内切酶EcoRI酶切自杀质粒pKR6K(Wang et al.,J.Biol.Chem.2014,289:6080-6090),将酶切产物进行1.0%琼脂糖凝胶电泳,回收并纯化,得到线性化的质粒pKR6K。
使用无缝克隆和组装试剂盒(pEASY-Uni Seamless Cloning and Assembly Kit,北京全式金生物技术有限公司),将上游同源片段、下游同源片段和线性化的质粒pKR6K连接,得到可部分缺失adhE基因的自杀质粒pKR6K-ΔadhE。
(2)adhE基因部分缺失的产酸克雷伯氏菌的构建
将pKR6K-ΔadhE转化至大肠杆菌S17-1(λpir)中,得到供体菌大肠杆菌S17-1(λpir)(pKR6K-ΔadhE)。将供体菌大肠杆菌S17-1(λpir)(pKR6K-ΔadhE)与受体菌产酸克雷伯氏菌PDL-1进行双亲本杂交,使pKR6K-ΔadhE上的adhE基因上游同源片段和下游同源片段与产酸克雷伯氏菌PDL-1的基因组发生同源重组,从而使产酸克雷伯氏菌PDL-1的adhE基因缺失1876bp,达到使adhE基因缺陷的目的。具体方法与实施例2相同,只是PCR验证时,使用引物adhE-1:5’-ACATGATTACGAATTCATGGCTGTTACTAATGTCGC-3’(如SEQ ID NO:18所示)和引物adhE-4:5’-TACCGAGCTCGAATTCTTAAGCGGATTTTTTCGCTT-3’(如SEQ ID NO:21所示)进行PCR验证。获得的adhE基因缺陷的菌株,命名为产酸克雷伯氏菌PDL-2。
实施例4:产酸克雷伯氏菌PDL-2中乙酸激酶和乙酰磷酸转移酶基因(ackA-pta)的缺陷
(1)用于在产酸克雷伯氏菌PDL-2中部分缺失ackA-pta基因的载体的构建
根据ackA-pta基因序列(如SEQ ID NO:4所示)设计引物,PCR扩增ackA-pta基因的上游和下游同源片段。以产酸克雷伯氏菌PDL-2的基因组DNA为模板,使用引物ackA-pta-1:5’-ACATGATTACGAATTCATGTCGAGTAAGTTAGTACT-3’(如SEQ ID NO:22所示)和引物ackA-pta-2:5’-CACGCGCGGTCCTCAGCGATACCGATCAGG-3’(如SEQ ID NO:23所示)进行PCR扩增,得到上游同源片段;使用引物ackA-pta-3:5’-ATCGCTGAGGACCGCGCGTGGCCATGCTCT-3’(如SEQ ID NO:24所示)和引物ackA-pta-4:5’-TACCGAGCTCGAATTCTTATGCTTGCTGCTGGGACG-3’(如SEQ ID NO:25所示)进行PCR扩增,得到下游同源片段。PCR扩增条件为:95℃5分钟;95℃30秒,60℃30秒,72℃1分钟,共30个循环;72℃5分钟。PCR反应结束后,将PCR扩增产物进行1.0%琼脂糖凝胶电泳,回收并纯化,得到上游和下游同源片段。
使用限制性内切酶EcoRI酶切自杀质粒pKR6K(Wang et al.,J.Biol.Chem.2014,289:6080-6090),将酶切产物进行1.0%琼脂糖凝胶电泳,回收并纯化,得到线性化的质粒pKR6K。
使用无缝克隆和组装试剂盒(pEASY-Uni Seamless Cloning and Assembly Kit,北京全式金生物技术有限公司),将上游同源片段、下游同源片段和线性化的质粒pKR6K连接,得到可部分缺失ackA-pta基因的自杀质粒pKR6K-ΔackA-pta。
(2)ackA-pta基因部分缺失的产酸克雷伯氏菌的构建
将pKR6K-ΔackA-pta转化至大肠杆菌S17-1(λpir)中,得到供体菌大肠杆菌S17-1(λpir)(pKR6K-ΔackA-pta)。将供体菌大肠杆菌S17-1(λpir)(pKR6K-ΔackA-pta)与受体菌产酸克雷伯氏菌PDL-2进行双亲本杂交,使pKR6K-ΔackA-pta上的ackA-pta基因上游同源片段和下游同源片段与产酸克雷伯氏菌PDL-2的基因组发生同源重组,从而使产酸克雷伯氏菌PDL-2的ackA-pta基因缺失2749bp,达到使ackA-pta基因缺陷的目的。具体方法与实施例2相同,只是PCR验证时,使用引物ackA-pta-1:5’-ACATGATTACGAATTCATGTCGAGTAAGTTAGTACT-3’(如SEQ ID NO:22所示)和引物ackA-pta-4:5’-TACCGAGCTCGAATTCTTATGCTTGCTGCTGGGACG-3’(如SEQ ID NO:25所示)进行PCR验证。获得的ackA-pta基因缺陷的菌株,命名为产酸克雷伯氏菌PDL-3。
实施例5:产酸克雷伯氏菌PDL-3中丙酮酸氧化酶基因(poxB)的缺陷
(1)用于在产酸克雷伯氏菌PDL-3中部分缺失poxB基因的载体的构建
根据poxB基因序列(如SEQ ID NO:5所示)设计引物,PCR扩增poxB基因的上游和下游同源片段。以产酸克雷伯氏菌PDL-3的基因组DNA为模板,使用引物poxB-1:5’-ACATGATTACGAATTCATGAAACAGACCGTGGCGGC-3’(如SEQ ID NO:26所示)和引物poxB-2:5’-AAAATCCCCCGGGTTGAGACCAGTTCACAG-3’(如SEQ ID NO:27所示)进行PCR扩增,得到上游同源片段;使用引物poxB-3:5’-GTCTCAACCCGGGGGATTTTCTCTCGCTGG-3’(如SEQ ID NO:28所示)和引物poxB-4:5’-TACCGAGCTCGAATTCTTACCTTAGCCAGTTAGTTT-3’(如SEQ ID NO:29所示)进行PCR扩增,得到下游同源片段。PCR扩增条件为:95℃5分钟;95℃30秒,60℃30秒,72℃1分钟,共30个循环;72℃5分钟。PCR反应结束后,将PCR扩增产物进行1.0%琼脂糖凝胶电泳,回收并纯化,得到上游和下游同源片段。
使用限制性内切酶EcoRI酶切自杀质粒pKR6K(Wang et al.,J.Biol.Chem.2014,289:6080-6090),将酶切产物进行1.0%琼脂糖凝胶电泳,回收并纯化,得到线性化的质粒pKR6K。
使用无缝克隆和组装试剂盒(pEASY-Uni Seamless Cloning and Assembly Kit,北京全式金生物技术有限公司),将上游同源片段、下游同源片段和线性化的质粒pKR6K连接,得到可部分缺失poxB基因的自杀质粒pKR6K-ΔpoxB。
(2)poxB基因部分缺失的产酸克雷伯氏菌的构建
将pKR6K-ΔpoxB转化至大肠杆菌S17-1(λpir)中,得到供体菌大肠杆菌S17-1(λpir)(pKR6K-ΔpoxB)。将供体菌大肠杆菌S17-1(λpir)(pKR6K-ΔpoxB)与受体菌产酸克雷伯氏菌PDL-3进行双亲本杂交,使pKR6K-ΔpoxB上的poxB基因上游同源片段和下游同源片段与产酸克雷伯氏菌PDL-3的基因组发生同源重组,从而使产酸克雷伯氏菌PDL-3的poxB基因缺失919bp,达到使poxB基因缺陷的目的。具体方法与实施例2相同,只是PCR验证时,使用引物poxB-1:5’-ACATGATTACGAATTCATGAAACAGACCGTGGCGGC-3’(如SEQ ID NO:26所示)和引物poxB-4:5’-TACCGAGCTCGAATTCTTACCTTAGCCAGTTAGTTT-3’(如SEQ ID NO:29所示)进行PCR验证。获得的poxB基因缺陷的菌株,命名为产酸克雷伯氏菌PDL-4。
实施例6:产酸克雷伯氏菌PDL-4中富马酸还原酶基因(frdA)的缺陷
(1)用于在产酸克雷伯氏菌PDL-4中部分缺失frdA基因的载体的构建
根据frdA基因序列(如SEQ ID NO:6所示)设计引物,PCR扩增frdA基因的上游和下游同源片段。以产酸克雷伯氏菌PDL-4的基因组DNA为模板,使用引物frdA-1:5’-ACATGATTACGAATTCGTGCAAACTTTTCAAGCCGA-3’(如SEQ ID NO:30所示)和引物frdA-2:5’-GTAGATGCCGAGCCGGTTTTATCGGCAGCG-3’(如SEQ ID NO:31所示)进行PCR扩增,得到上游同源片段;使用引物frdA-3:5’-AAAACCGGCTCGGCATCTACCGTACGCCGG-3’(如SEQ ID NO:32所示)和引物frdA-4:5’-TACCGAGCTCGAATTCTCAGCCATTCGTCGTCTCCT-3’(如SEQ ID NO:33所示)进行PCR扩增,得到下游同源片段。PCR扩增条件为:95℃5分钟;95℃30秒,60℃30秒,72℃1分钟,共30个循环;72℃5分钟。PCR反应结束后,将PCR扩增产物进行1.0%琼脂糖凝胶电泳,回收并纯化,得到上游和下游同源片段。
使用限制性内切酶EcoRI酶切自杀质粒pKR6K(Wang et al.,J.Biol.Chem.2014,289:6080-6090),将酶切产物进行1.0%琼脂糖凝胶电泳,回收并纯化,得到线性化的质粒pKR6K。
使用无缝克隆和组装试剂盒(pEASY-Uni Seamless Cloning and Assembly Kit,北京全式金生物技术有限公司),将上游同源片段、下游同源片段和线性化的质粒pKR6K连接,得到可部分缺失frdA基因的自杀质粒pKR6K-ΔfrdA。
(2)frdA基因部分缺失的产酸克雷伯氏菌的构建
将pKR6K-ΔfrdA转化至大肠杆菌S17-1(λpir)中,得到供体菌大肠杆菌S17-1(λpir)(pKR6K-ΔfrdA)。将供体菌大肠杆菌S17-1(λpir)(pKR6K-ΔfrdA)与受体菌产酸克雷伯氏菌PDL-4进行双亲本杂交,使pKR6K-ΔfrdA上的frdA基因上游同源片段和下游同源片段与产酸克雷伯氏菌PDL-4的基因组发生同源重组,从而使产酸克雷伯氏菌PDL-4的frdA基因缺失991bp,达到使frdA基因缺陷的目的。具体方法与实施例2相同,只是PCR验证时,使用引物frdA-1:5’-ACATGATTACGAATTCGTGCAAACTTTTCAAGCCGA-3’(如SEQ ID NO:30所示)和引物frdA-4:5’-TACCGAGCTCGAATTCTCAGCCATTCGTCGTCTCCT-3’(如SEQ ID NO:33所示)进行PCR验证。获得的frdA基因缺陷的菌株,命名为产酸克雷伯氏菌PDL-5。
实施例7:产酸克雷伯氏菌PDL-5中D-乳酸脱氢酶基因(dldh)的缺陷
(1)用于在产酸克雷伯氏菌PDL-5中缺失dldh基因的载体的构建
根据dldh基因序列(如SEQ ID NO:7所示)设计引物,PCR扩增dldh基因的上游和下游同源片段。以产酸克雷伯氏菌PDL-5的基因组DNA为模板,使用引物dldh-1:5’-ACATGATTACGAATTCATGGAGCATCTGCACATGAA-3’(如SEQ ID NO:34所示)和引物dldh-2:5’-AAAGGGAAAGGAATAAAGACTTTTCTCCAGTGATAATACC-3’(如SEQ ID NO:35所示)进行PCR扩增,得到上游同源片段;使用引物dldh-3:5’-CTGGAGAAAAGTCTTTATTCCTTTCCCTTTTGTGCTC-3’(如SEQ ID NO:36所示)和引物dldh-4:5’-TACCGAGCTCGAATTCTTATCAGAACTGATCTTCTT-3’(如SEQ ID NO:37所示)进行PCR扩增,得到下游同源片段。PCR扩增条件为:95℃5分钟;95℃30秒,60℃30秒,72℃1分钟,共30个循环;72℃5分钟。PCR反应结束后,将PCR扩增产物进行1.0%琼脂糖凝胶电泳,回收并纯化,得到上游和下游同源片段。
使用限制性内切酶EcoRI酶切自杀质粒pKR6K(Wang et al.,J.Biol.Chem.2014,289:6080-6090),将酶切产物进行1.0%琼脂糖凝胶电泳,回收并纯化,得到线性化的质粒pKR6K。
使用无缝克隆和组装试剂盒(pEASY-Uni Seamless Cloning and Assembly Kit,北京全式金生物技术有限公司),将上游同源片段、下游同源片段和线性化的质粒pKR6K连接,得到可缺失dldh基因的自杀质粒pKR6K-Δdldh。
(2)dldh基因缺失的产酸克雷伯氏菌的构建
将pKR6K-Δdldh转化至大肠杆菌S17-1(λpir)中,得到供体菌大肠杆菌S17-1(λpir)(pKR6K-Δdldh)。将供体菌大肠杆菌S17-1(λpir)(pKR6K-Δdldh)与受体菌产酸克雷伯氏菌PDL-5进行双亲本杂交,使pKR6K-Δdldh上的dldh基因上游同源片段和下游同源片段与产酸克雷伯氏菌PDL-5的基因组发生同源重组,从而使产酸克雷伯氏菌PDL-5的dldh基因缺失990bp,达到dldh基因缺陷的目的。具体方法与实施例2相同,只是PCR验证时,使用引物dldh-1:5’-ACATGATTACGAATTCATGGAGCATCTGCACATGAA-3’(如SEQ ID NO:34所示)和引物dldh-4:5’-TACCGAGCTCGAATTCTTATCAGAACTGATCTTCTT-3’(如SEQ ID NO:37所示)进行PCR验证。获得的dldh基因缺陷的菌株,命名为产酸克雷伯氏菌PDL-6。
实施例2-7给出了构建budA、budB、adhE、ackA-pta、poxB、frdA和dldh酶失活菌株的方法,即通过基因同源重组的方式来实现基因缺陷从而导致其所编码的酶失活。然而导致酶失活的方式并不限于基因同意重组,还可以是:小RNA干扰,点突变,加入相关酶的抑制剂等等。
实施例8:将凝结芽孢杆菌2-6的L-乳酸脱氢酶基因(lldh)插入产酸克雷伯氏菌PDL-6的α-乙酰乳酸脱羧酶基因(budA)在基因组中上游的启动子之后
(1)用于在产酸克雷伯氏菌PDL-6的budA基因在基因组中上游的启动子之后插入凝结芽孢杆菌2-6的lldh基因的载体的构建
根据产酸克雷伯氏菌基因组序列设计引物,PCR扩增budA基因在基因组上游和下游的同源片段。以产酸克雷伯氏菌PDL-6的基因组DNA为模板,使用引物budA-lldh-1:5’-ACATGATTACGAATTCATTGCCCTCGACCTGATGTAAC-3’(如SEQ ID NO:38所示)和引物budA-lldh-2:5’-ATTGACTTTTTTCATTACCCGCTTCCTCGTTCAAC-3’(如SEQ ID NO:39所示)进行PCR扩增,得到上游同源片段;使用引物budA-lldh-3:5’-GCACCGATATTGTAAGTCACTACAGAAGGAATCCAGCAAT-3’(如SEQ ID NO:40所示)和引物budA-lldh-4:5’-TACCGAGCTCGAATTCGCACGGCAAACTTACTTGAGCTA-3’(如SEQ ID NO:41所示)进行PCR扩增,得到下游同源片段。根据凝结芽孢杆菌2-6的lldh基因序列(如SEQ ID NO:8所示)设计引物,PCR扩增凝结芽孢杆菌2-6的lldh基因。以凝结芽孢杆菌2-6的基因组DNA为模板,使用引物lldh-1:5’-ACGAGGAAGCGGGTAATGAAAAAAGTCAATCGTATTGCA-3’(如SEQ ID NO:42所示)和引物lldh-2:5’-TCCTTCTGTAGTGACTTACAATATCGGTGCCATTGTTT-3’(如SEQ ID NO:43所示)进行PCR扩增,得到凝结芽孢杆菌2-6的lldh基因片段。PCR扩增条件为:95℃5分钟;95℃30秒,60℃30秒,72℃1分钟,共30个循环;72℃5分钟。PCR反应结束后,将PCR扩增产物进行1.0%琼脂糖凝胶电泳,回收并纯化,得到budA基因在基因组上游和下游的同源片段,以及凝结芽孢杆菌2-6的lldh基因片段。
使用限制性内切酶EcoRI酶切自杀质粒pKR6K(Wang et al.,J.Biol.Chem.2014,289:6080-6090),将酶切产物进行1.0%琼脂糖凝胶电泳,回收并纯化,得到线性化的质粒pKR6K。
使用无缝克隆和组装试剂盒(pEASY-Uni Seamless Cloning and Assembly Kit,北京全式金生物技术有限公司),将budA基因在基因组上游和下游的同源片段,以及凝结芽孢杆菌2-6的lldh基因片段与线性化的质粒pKR6K连接,得到用于在产酸克雷伯氏菌PDL-6中替换budB基因为凝结芽孢杆菌2-6的lldh基因的载体pKR6K-ΔbudA::lldh。
(2)携带凝结芽孢杆菌2-6的lldh基因的产酸克雷伯氏菌的构建
将pKR6K-ΔbudA::lldh转化至大肠杆菌S17-1(λpir)中,得到供体菌大肠杆菌S17-1(λpir)(pKR6K-ΔbudA::lldh)。将供体菌大肠杆菌S17-1(λpir)(pKR6K-ΔbudA::lldh)与受体菌产酸克雷伯氏菌PDL-6进行双亲本杂交,使pKR6K-ΔbudA::lldh上的budA基因上游同源片段和下游同源片段与产酸克雷伯氏菌PDL-6的基因组发生同源重组,从而将凝结芽孢杆菌2-6的lldh基因插入到产酸克雷伯氏菌PDL-6的budA基因在基因组中上游的启动子之后。具体方法与实施例2相同,只是PCR验证时,使用引物budA-lldh-1:5’-ACATGATTACGAATTCATTGCCCTCGACCTGATGTAAC-3’(如SEQ ID NO:38所示)和budA-lldh-4:5’-TACCGAGCTCGAATTCGCACGGCAAACTTACTTGAGCTA-3’(如SEQ ID NO:41所示)进行PCR验证。获得的携带凝结芽孢杆菌2-6的lldh基因的菌株,命名为产酸克雷伯氏菌PLL CCTCC M 2016186。
实施例9:使用产酸克雷伯氏菌PLL CCTCC M 2016186分批发酵生产1,3-PD和L-LAC
(1)菌株选择:选用产酸克雷伯氏菌PLL CCTCC M 2016186;
(2)种子培养:选用步骤(1)的菌株,在无菌条件下接种至甘油培养基中,培养温度为30℃,摇床震荡转速为200rpm,培养时间为15小时,制得种子培养液;
(3)发酵:将步骤(2)中制得的种子培养液接种至装有甘油培养基的发酵罐中,接种量为5%(v/v),发酵温度为30℃,通气量为0.5vvm,搅拌转速为200rpm,发酵过程中,使用25%(w/v)氢氧化钠水溶液作为中和剂调节发酵液pH为7.0,发酵方式为分批发酵,当甘油培养基中的甘油耗尽时,停止发酵。
其中,所述甘油培养基的配方为:酵母粉2g/L、K2HPO4·3H2O 5g/L、KH2PO41g/L、NH4Cl 2g/L、MgSO4·7H2O 0.1g/L、FeCl3·6H2O 10mg/L、CoCl2·6H2O 10mg/L和甘油60g/L;121℃灭菌20分钟。
发酵14小时后,甘油培养基中的甘油耗尽,停止发酵,检测发酵液中的产物组成和浓度。发酵的主要产物为1,3-PD和L-LAC,1,3-PD浓度为22.7g/L,L-LAC浓度为32.4g/L。副产物仅能检测到少量的乙酸、甲酸和琥珀酸,乙酸浓度为0.7g/L,琥珀酸浓度为0.3g/L,丙酮酸浓度为0.4g/L。发酵液中检测不到2,3-丁二醇、乙醇和甲酸。1,3-PD的摩尔转化率为45.8%,L-lac的摩尔转化率为50.0%。两种产物的总转化率为95.8%。
实施例10:使用产酸克雷伯氏菌PLL CCTCC M 2016186分批补料发酵生产1,3-PD和L-LAC
(1)菌株选择:选用产酸克雷伯氏菌PLL CCTCC M 2016186;
(2)种子培养:选用步骤(1)的菌株,在无菌条件下接种至甘油培养基中,培养温度为37℃,摇床震荡转速为150rpm,培养时间为12小时,制得种子培养液;
(3)发酵:将步骤(2)中制得的种子培养液接种至装有甘油培养基的发酵罐中,接种量为2.5%(v/v),发酵温度为37℃,通气量为1vvm,搅拌转速为250rpm,发酵过程中,使用25%(w/v)的氢氧化钙和水的混合乳液作为中和剂调节发酵液pH为6.5,发酵方式为分批补料发酵,当甘油培养基中的甘油耗尽时,通过向发酵罐中补加700g/L的甘油溶液,控制发酵液中甘油浓度为5-30g/L,当发酵液中1,3-PD或L-LAC浓度不再升高时,停止发酵。
其中,所述甘油培养基的配方为:酵母粉5g/L、K2HPO4·3H2O 10g/L、KH2PO42g/L、NH4Cl 1g/L、MgSO4·7H2O 0.1g/L、FeCl3·6H2O 20mg/L、CoCl2·6H2O 15mg/L和甘油20g/L;121℃灭菌20分钟。
发酵30小时后,发酵液中1,3-PD和L-LAC浓度不再升高时,停止发酵,检测发酵液中的产物组成和浓度。发酵的主要产物为1,3-PD和L-LAC,1,3-PD浓度为70.0g/L,L-LAC浓度为104.0g/L。副产物仅能检测到少量的乙酸和琥珀酸,乙酸浓度为1.5g/L,琥珀酸浓度为0.7g/L。发酵液中检测不到2,3-丁二醇、乙醇、甲酸等其他副产物。1,3-PD的摩尔转化率为42.5%,L-乳酸的摩尔转化率为53.4%。两种产物的总转化率为95.9%。
实施例9和10只是本发明在应用方法上的两个较佳实施方式,其所限定的数值在合理范围内的变化也可以实现同一目的,因此不能以实施例9和10中给出的数值对本发明进行限制。虽然实施例9和10是对产酸克雷伯氏菌PLL的发酵产物进行检测,证明了产酸克雷伯氏菌PLL可以生产高摩尔转化率的1,3-PD和L-LAC,但是并不能仅仅理解为本发明所公开的产酸克雷伯氏菌进行基因改造的技术方案只适用于产酸克雷伯氏菌PLL,而应该理解为只要是通过本发明所公开的对产酸克雷伯氏菌进行基因改造的技术方案所得到的人工菌都具有同时提高1,3-PD和L-LAC的摩尔转化率和减少副产物的能力。此外,实施例1-10是以产酸克雷伯氏菌PDL-0为野生菌进行基因改造的,若通过其他与产酸克雷伯氏菌PDL-0具有相同代谢通路的菌进行同样的基因改造,应当理解为也能同时提高1,3-PD和L-LAC的摩尔转化率和减少副产物。
使用产酸克雷伯氏菌PLL CCTCC M 2016186,以甘油为底物,进行分批补料发酵,得到的发酵液中目标产物1,3-PD和L-LAC浓度高,副产物种类少且浓度低,此外,产酸克雷伯氏菌PLL CCTCC M 2016186易于离心和过滤,这些优势都有利于1,3-PD和L-LAC的高效生物法生产,同时有利于产物的提取,说明本发明的产酸克雷伯氏菌PLL CCTCC M 2016186,具有重要的实际应用价值。
实施例11:外源引入1,3-PD合成途径
(1)菌株选择:选用大肠杆菌K12;
(2)基因工程改造,在K12中引入外源1,3-PD合成途径
选用产酸克雷伯氏菌中的1,3-PD合成途径中的甘油脱水酶编码基因dhaB及1,3-PD氧化还原酶编码基因dhaT,进行PCR克隆后连接到质粒DNA pet-Duet上,并转化到大肠杆菌K12中。该菌确定为K12-dhaBdhaT。
以上详细描述了本发明的较佳具体实施例。应当理解,本领域的普通技术无需创造性劳动就可以根据本发明的构思作出诸多修改和变化。因此,凡本技术领域中技术人员依本发明的构思在现有技术的基础上通过逻辑分析、推理或者有限的实验可以得到的技术方案,皆应在由权利要求书所确定的保护范围内。
- 还没有人留言评论。精彩留言会获得点赞!