重组菌株及其构建方法与在产菜油甾醇中的应用与流程
本发明涉及基因工程技术领域,尤其涉及重组菌株及其构建方法与在产菜油甾醇中的应用。
背景技术:
甾体激素类药物是仅次于抗生素的第二类药物,在治疗呼吸系统疾病、内分泌失调、淋巴性白血病、风湿病等方面有广泛应用。菜油甾醇作为甾体激素类药物的重要前体,对于众多甾体激素类药物如糖皮质激素、性激素等的合成有重大应用。传统菜油甾醇的制备多采用植物提取的方式,该方法操作过程繁琐、收率低、有机物污染严重,极大地限制了菜油甾醇进一步的生产和应用。
利用微生物合成的方式成本较低、产量高、污染小且产品安全性有效保证,被认为是一种有前途的生产方法。菜油甾醇属于一种萜烯类物质,在耶氏解脂酵母体内萜烯类物质的合成均需要一种重要前体——乙酰辅酶a(acetyl-coa)。在耶氏解脂酵母内,当以碳水化合物(如葡萄糖)为碳源时,胞内乙酰coa主要由丙酮酸通过到乙醛再到乙酸,再由乙酰辅酶a合成酶(acetyl-coasynthetase,acs)催化合成,或者由线粒体穿梭到胞质中的柠檬酸在柠檬酸裂解酶(atp-citratelyase,acl)催化下提供;而当以疏水性物质(如油)为碳源时,胞内乙酰coa主要经过油脂的β氧化过程再由acl催化提供,生成的乙酰coa经过mva路径合成下游萜烯类物质。在酿酒酵母体内,提高胞质乙酰coa的合成被证明为一种提高萜烯类物质的有效手段。在耶氏解脂酵母内通过引入酿酒酵母(saccharomycescerevisiae)来源的acs及小鼠(musmusculus)来源的acl可以使乙酰coa含量分别提高131.2%和144.2%,随之α-酮戊二酸产量从36.3g/l分别提高到42.2g/l和46.7g/l。然而,胞内的乙酰coa除了合成萜烯类物质,还作为前体用于脂肪酸的合成,进而形成脂滴,而形成的脂滴对于萜烯类弱极性物质提供储存作用,从而避免与细胞膜结合造成膜毒性,因此,萜烯的与脂肪酸的合成既存在竞争作用又具有联动性。
在此基础上提高耶氏解脂酵母体内的萜烯类目标产物积累可利用两种乙酰coa代谢流,一方面耶氏解脂酵母胞质内的乙酰coa竞争性地分配到萜烯合成或脂肪酸合成:在mva路径限速酶hmgr催化下进入萜烯合成,在脂肪酸合成关键酶苹果酸酶(malicenzyme,mae)催化下进入脂肪酸合成;另一方面脂滴中脂肪酸氧化重新形成胞质乙酰coa用于萜烯或脂肪酸的积累,耶氏解脂酵母酵母的脂肪酸β氧化包含四步:第一步为限速步骤,不同链长脂肪酸由六种酯酰辅酶a氧化酶(acyl-coaoxidases)pox1~pox6催化,第二步和第三步由多功能酶(multifunctionalenzyme,mfe)催化,不饱和acyl-coa转化为3-羟基酯酰coa(3-hydroxyacyl-coa)再转化为3-酮基酯酰coa(3-ketoacyl-coa),最后一步由过氧化物体酮基酯酰coa硫解酶(peroxisomalketoacyl-coathiolase,pot)催化形成少2个碳的acyl-coa及acetyl-coa,acetyl-coa进入tca循环。
因此如何调节两种代谢流间的平衡是提高耶氏解脂酵母内萜烯积累的关键点,而对于耶氏解脂酵母的代谢流的调节与平衡目前还没有报道。
技术实现要素:
有鉴于此,本发明要解决的技术问题在于提供重组菌株及其构建方法与在产菜油甾醇中的应用。本发明通过调节萜烯合成与脂肪酸氧化代谢流来实现菜油甾醇在耶氏解脂酵母体内产量的提高。
本发明提供的重组菌株,表达dhcr7基因且erg5基因失活;所述dhcr7基因的序列如seqidno:1所示。
本发明提供的重组菌株还过表达乙酰coa代谢相关基因;
所述乙酰coa代谢相关基因选自hmgr基因、acl基因、pox2基因、pox4基因、pox5基因或pox6基因。
本发明提供的重组菌株的底盘菌为耶氏解脂酵母。
本发明采用的耶氏解脂酵母为野生型耶氏解脂酵母atcc201249。
本发明提供的重组菌株的基因型为:
erg5::-exp1p-drdhcr7-xpr2tpbr322::hp4d-hmgr-xpr2t;
erg5::-exp1p-drdhcr7-xpr2tpbr322::hp4d-acl-xpr2t;
erg5::-exp1p-drdhcr7-xpr2tpbr322::hp4d-pox2-xpr2t;
erg5::-exp1p-drdhcr7-xpr2tpbr322::hp4d-pox4-xpr2t;
erg5::-exp1p-drdhcr7-xpr2tpbr322::hp4d-pox5-xpr2t;
或erg5::-exp1p-drdhcr7-xpr2tpbr322::hp4d-pox6-xpr2t。
本发明提供的重组菌株中,dhcr7基因的插入位点位于erg5基因内。其以exp1p为启动子,xpr2t为终止子。另外添加gpm1t终止子加强终止效果。
本发明提供的重组菌株中,乙酰coa代谢相关基因的插入位点位于基因组的pbr322platform位置。其中,插入的乙酰coa代谢相关基因hp4d为启动子、xpr2t为终止子。
本发明还提供了保藏编号为cgmccno.14039的重组菌株。
本发明对构建获得的,基因型为erg5::exp1p-dhcr7(daniorerio)-xpr2tpbr322::hp4d-pox2-xpr2t的菌株进行了生物保藏,保藏编号为cgmccno.14039。
本发明提供的重组菌株在生产菜油甾醇的应用。
本发明利用重组耶氏解脂酵母底盘菌株通过调节胞质内乙酰coa合成与脂肪酸合成的代谢流提高菜油甾醇产量,以本发明提供的重组菌株进行发酵生产菜油甾醇,产量可达941.5mg/l。
本发明提供的重组菌株的一级、二级种子培养基包括水和22g/l葡萄糖、20g/l蛋白胨和10g/l酵母浸粉。
本发明提供的重组菌株的三级种子、发酵培养基包括水和3.7%v/v葵花籽油、20g/l蛋白胨和10g/l酵母浸粉。
本发明所述重组菌株的构建方法,包括:
以耶氏解脂酵母为底盘菌,将所述dhcr7基因整合至底盘菌基因组并使其内源erg5基因失活,制得中间菌株;
在所述中间菌株的基因组中,插入乙酰coa代谢相关基因,获得重组菌株;
所述乙酰coa代谢相关基因选自hmgr基因、acl基因、pox2基因、pox4基因、pox5基因或pox6基因。
中间菌株的构建包括:将外源dr_dhcr7基因用goldengate方法构建入puc57k-eno2t-exp1p-xpr2t-gpm1t(可在http://synbioml.org/免费获取)质粒获得dr_dhcr7的puc57k-eno2t-exp1p-dr_dhcr7-xpr2t-gpm1t;经noti酶切后获得外源功能基因表达盒eno2t-exp1p-dr_dhcr7-xpr2t-gpm1t。
质粒peasy-blunt-erg5(700)-ura-eno2t(构建过程参见《engineeringyarrowialipolyticaforcampesteroloverproduction》haoxingdu,wenhaixiao,等发表于plosone,jan2016)与质粒peasy-blunt-gpm1t-erg5(670)(构建过程参见《engineeringyarrowialipolyticaforcampesteroloverproduction》haoxingdu,wenhaixiao,等发表于plosone,jan2016)经noti酶切后获得基因组erg5位置上游整合片段erg5(700)-ura-eno2t和下游整合片段gpm1t-erg5(670)。
采用醋酸锂法将上述获得的所有整合片段(外源功能基因表达盒、上游整合片段、下游整合片段)共转化到野生型耶氏解脂酵母atcc201249底盘菌株中,采用sc-ura固体平板进行筛选、经鉴定获得阳性转化子,为中间菌株。
乙酰coa代谢相关基因的插入采用单拷贝整合型质粒pina1269作为载体,基因构建入载体的方法采用in-fusion组装方法。整合型质粒构建入中间菌株的方式采用醋酸锂法,以sc-ura-leu固体平板进行筛选、经鉴定获得阳性转化子,为重组菌株。
中间菌株和重组菌株的鉴定采用菌落pcr的方式。
本发明还提供了一种生产菜油甾醇的方法,以植物油为碳源,发酵本发明提供的重组菌株,制得含有菜油甾醇的发酵液。
在本发明中,发酵采用流加碳源的方式,植物油的浓度不大于2g/l。
在本发明中,植物油为葵花籽油。
在本发明中,发酵的温度为28℃,ph值为5.5,容氧量>30%、通气量为1vvm。
本发明中,发酵制得含有菜油甾醇的发酵液后,还包括菜油甾醇的提取,方法包括:
将发酵液,6000rpm离心5min收集菌体,水洗两次;
用液氮冷冻细胞、破碎后,加入甲醇配制的1.5mol/lkoh,60℃皂化反应8h~12h;
皂化后加入正己烷(与甲醇的体积比为1:1)涡旋震荡10min后,5000rpm离心10min,收集有机相进行真空冷冻干燥;
然后加入衍生化试剂n-甲基-n-三甲基硅基三氟乙酰胺(mstfa)30℃孵育2小时。
本发明构建了表达dhcr7基因且erg5基因失活的重组菌株,该菌株还过表达乙酰coa代谢相关基因,该重组耶氏解脂酵母底盘菌株通过调节胞质内乙酰coa合成与脂肪酸合成的代谢流提高菜油甾醇产量,实验表明,经工艺优化,本发明提供重组菌株的菜油甾醇产量可达941.5mg/l。
生物保藏说明
sybe_yl02060069,分类命名:解脂耶罗维亚酵母yarrowialipolytica,于2017年04月17日保藏在保藏于中国微生物菌种保藏管理委员会普通微生物中心(cgmcc),地址为:北京市朝阳区北辰西路1号院3号中国科学院微生物研究所。保藏编号为cgmccno.14039。
附图说明
图1示菜油甾醇合成关键基因dr_dhcr7基因组整合示意图;
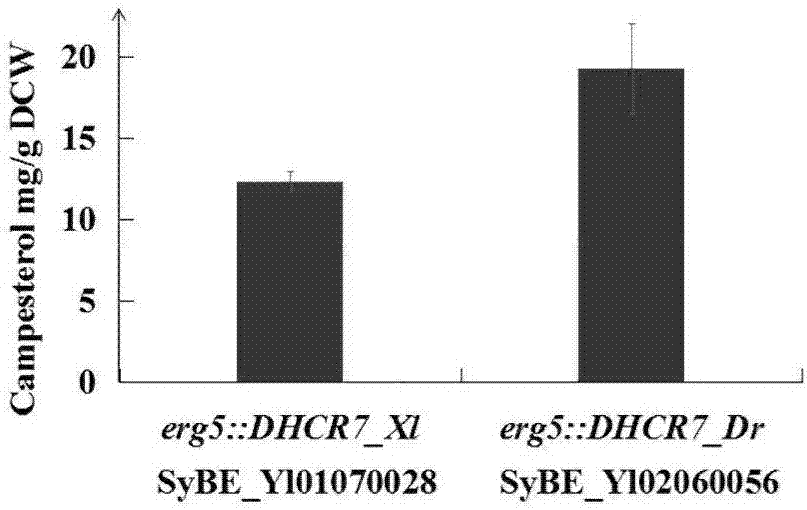
图2示是合成菜油甾醇耶氏解脂酵母菌株sybe_yl02060056与目前最高产耶氏解脂酵母菌株sybe_yl02060028产量比较图;
图3示过表达内源基因的整合型质粒构建过程图;
图4示以葡萄糖为碳源摇瓶产量,其中,图4-a示以葡萄糖为碳源耶氏解脂酵母中心代谢路径形成乙酰coa;图4-b示以葡萄糖为碳源耶氏解脂酵母β氧化形成乙酰coa,图4-c示以葡萄糖为碳源乙酰coa分配;图4-d示以葡萄糖为碳源摇瓶发酵菜油甾醇标准曲线;图4-e示以葡萄糖为碳源乙酰coa代谢相关基因;
图5示葵花籽油为碳源摇瓶产量,其中,图5-a示以葵花籽油为碳源耶氏解脂酵母中心代谢路径形成乙酰coa;图5-b示以葵花籽油为碳源耶氏解脂酵母β氧化形成乙酰coa,图5-c示以葵花籽油为碳源乙酰coa分配;图5-d示以葵花籽油为碳源摇瓶发酵菜油甾醇标准曲线;图5-e示以葵花籽油为碳源乙酰coa代谢相关基因;
图6示发酵罐中重组耶氏解脂酵母发酵曲线图,其中,图6-a示发酵罐发酵产生菜油甾醇标准曲线;图6-b示过表达acl基因的菌株发酵曲线;图6-c示过表达pox2基因的菌株发酵曲线;图6-d示过表达hmgr基因的菌株发酵曲线。
具体实施方式
本发明提供了重组菌株及其构建方法与在产菜油甾醇中的应用,本领域技术人员可以借鉴本文内容,适当改进工艺参数实现。特别需要指出的是,所有类似的替换和改动对本领域技术人员来说是显而易见的,它们都被视为包括在本发明。本发明的方法及应用已经通过较佳实施例进行了描述,相关人员明显能在不脱离本发明内容、精神和范围内对本文的方法和应用进行改动或适当变更与组合,来实现和应用本发明技术。
本发明采用的仪器皆为普通市售品,皆可于市场购得。
下面结合实施例,进一步阐述本发明:
实施例高产菜油甾醇的重组耶氏解脂酵母菌株乙酰coa代谢调节与优化
本发明的高产菜油甾醇的重组耶氏解脂酵母菌株的构建及表征方法为:
1、合成菜油甾醇的重组耶氏解脂酵母的获得
野生型耶氏解脂酵母菌株体内合成的麦角固醇不能向下游甾体激素转化,菜油甾醇在酵母细胞中的合成需将其内源生产麦角固醇的路径截断,敲除固醇c-22还原酶(erg5),同时引入c7位还原酶(dhcr7)基因,即可将代谢流由生成麦角固醇转到菜油甾醇方向。与已报道菜油甾醇最高产量的耶氏解脂酵母构建方法一致(如图1),选取前期实验证明表现最好的斑马鱼(daniorerio)来源dhcr7基因,简写为dr_dhcr7,经耶氏解脂酵母密码子优化并适当规避bsaⅰ限制性内切酶酶切位点,在基因两端额外添加5’端gcggccgcggtctcca;3’taaaggagaccgcggccgc通过人工合成得到,具体基因序列如seqidno:1所示。
将实验室中puc57k-eno2t-exp1p-xpr2t-gpm1t质粒(可在http://synbioml.org/免费获取)及获得的外源dr_dhcr7基因用goldengate方法进行构建,获得dr_dhcr7的puc57k-eno2t-exp1p-dr_dhcr7-xpr2t-gpm1t大肠杆菌游离型多拷贝质粒。将该质粒与实验室质粒peasy-blunt-erg5(700)-ura-eno2t与peasy-blunt-gpm1t-erg5(670)(参见《engineeringyarrowialipolyticaforcampesteroloverproduction》)经noti酶切后获得基因组erg5位置上游整合片段erg5(700)-ura-eno2t,下游整合片段gpm1t-erg5(670),外源功能基因表达盒eno2t-exp1p-dr_dhcr7-xpr2t-gpm1t(参见《engineeringyarrowialipolyticaforcampesteroloverproduction》)。采用醋酸锂法将上述获得的所有整合片段共转化到野生型耶氏解脂酵母atcc201249底盘菌株中,采用sc-ura固体平板(合成酵母氮源ynb6.7g/l,葡萄糖22g/l,缺尿嘧啶的混合氨基酸粉末2g/l,2%的琼脂粉)进行筛选,得到的转化子进行划线分纯培养后进行菌落pcr验证,对验证正确的重组菌株保存甘油菌并命名为sybe_yl02060056,菌株基因组基因型为:erg5::ura3-exp1p-dhcr7(daniorerio)-xpr2t。
2、耶氏解脂酵母菌株sybe_yl02060056与高产菜油甾醇的耶氏解脂酵母菌株sybe_yl01070028摇瓶产量比较
实验材料:耶氏解脂酵母菌株sybe_yl02060056、sybe_yl01070028
试验方法:
一级、二级种子培养基:22g/l葡萄糖、20g/l蛋白胨、10g/l酵母浸粉;
发酵培养基:50g/l葡萄糖、20g/l蛋白胨、10g/l酵母浸粉
将菌株sybe_yl02060056和sybe_yl01070028接种于5ml一级种子培养基中,在28℃、250rpm培养24h,以初始菌体浓度od600=0.2分别接种于二级种子培养基中,在28℃、220rpm培养18h,再以初始菌体浓度od600=0.2分别接种于50ml发酵培养基中,每个实验组做两组平行,于28℃、220rpm条件下培养,发酵148小时结束,测定菌体密度(od600),取10ml菌液提取菜油甾醇。
菜油甾醇提取方法:取10ml发酵液,6000rpm离心5min收集菌体,水洗两次。用液氮冷冻细胞并在研钵中研磨,直到细胞被研磨成白色极细粉末,转移至新10ml离心管中,加入2ml甲醇配制的1.5mkoh,60℃水浴皂化反应过夜。皂化后加入2ml分析纯正己烷涡旋震荡10min对产物进行萃取,5000rpm离心10min收集一定量有机相真空冷冻干燥2h,加入400μl正己烷溶解再次冷冻干燥4h,加入200μl衍生化试剂n-甲基-n-三甲基硅基三氟乙酰胺(mstfa)30℃水浴2h,用2μm有机滤膜过滤后适当稀释利用gc/ms检测菜油甾醇含量。
表1耶氏解脂酵母菌株sybe_yl02060056与sybe_yl01070028摇瓶产量比较实验气相色谱-质谱联用(gc-ms)检测原始数据
试验结果:由表1及图2菌株sybe_yl02060056和sybe_yl01070028的菜油甾醇产量来看,发酵148小时,本专利构建获得的菌株sybe_yl02060056菜油甾醇产量为19.3±2.8mg/gdcw,与目前已报道最高产量菌株,即含有非洲爪蟾(xenopuslaevis)来源dhcr7的耶氏解脂酵母菌株sybe_yl01070028菜油甾醇产量(12.4±0.6mg/gdcw)相比提高56%,为目前的最高产量,因此该菌株可作为后续优化的重要底盘菌株。
2、乙酰coa代谢优化的耶氏解脂酵母菌株的构建
选取13个乙酰coa代谢相关基因(包括碳代谢相关cs、acl和acs基因;乙酰coa分配相关hmgr和mae基因;脂肪酸β氧化相关pox1~pox6、mfe和pot基因),以野生型耶氏解脂酵母atcc201249菌株基因组为模板,利用pcr扩增获得乙酰coa代谢相关基因如hmgr(耶氏解脂酵母菌株中内源基因的基因编号:yali0e04807g),cs(yali0e02684g),acl1(yali0e34793g),acl2(yali0d24431g),acs(yali0f05962g),pox1(yali0e32835g),pox2(yali0f10857g),pox3(yali0d24750g),pox4(yali0e27654g),pox5(yali0c23859g),pox6(yali0e06567g),mfe(yali0e15378g),pot(yali0e18568g),mae(yali0e18634g),通过in-fusion组装方法构建到耶氏解脂酵母单拷贝整合型质粒pina1269上(如图3),以强启动子hp4d为启动子、xpr2t为终止子,分别获得13种质粒pina1269-hmgr,pina1269-cs,pina1269-acl,pina1269-acs,pina1269-pox1,pina1269-pox2,pina1269-pox3,pina1269-pox4,pina1269-pox5,pina1269-pox6,pina1269-mfe,pina1269-pot,pina1269-mae,采用醋酸锂法将13种质粒分别转化到本专利获得的耶氏解脂酵母菌株sybe_yl02060056底盘菌株中,整合于基因组pbr322platform位置从而过表达13种乙酰coa代谢相关基因,采用sc-ura-leu固体平板(合成酵母氮源ynb6.7g/l,葡萄糖22g/l,缺尿嘧啶和亮氨酸的混合氨基酸粉末2g/l,2%的琼脂粉)进行筛选,得到的转化子进行划线分纯培养后提取酵母基因组进行pcr验证,对验证正确的重组菌株保存甘油菌并分别命名为sybe_yl02060063~sybe_yl02060065,sybe_yl02060067~sybe_yl02060076,各菌株基因组基因型如表2:
表2各菌株基因组基因型
3、比较耶氏解脂酵母菌株sybe_yl02060056、sybe_yl02060063~sybe_yl02060065,sybe_yl02060067~sybe_yl02060076分别以葡萄糖或葵花籽油为碳源时菜油甾醇摇瓶产量
实验材料:耶氏解脂酵母菌株sybe_yl02060056、sybe_yl02060063~sybe_yl02060065,sybe_yl02060067~sybe_yl02060076
试验方法:
一级、二级种子培养基:22g/l葡萄糖、20g/l蛋白胨、10g/l酵母浸粉;
以葡萄糖为碳源发酵培养基:50g/l葡萄糖、20g/l蛋白胨、10g/l酵母浸粉
以葵花籽油为碳源发酵培养基:3.7%v/v葵花籽油、20g/l蛋白胨、10g/l酵母浸粉
将上述菌株sybe_yl02060056、sybe_yl02060063~sybe_yl02060065,sybe_yl02060067~sybe_yl02060076接种于5ml一级种子培养基中,在28℃、250rpm培养24h,以初始菌体浓度od600=0.2分别接种于二级种子培养基中,在28℃、220rpm培养18h,再以初始菌体浓度od600=0.2分别接种于50ml以葡萄糖为碳源或以葵花籽油为碳源发酵培养基中,每个实验组做两组平行,于28℃、220rpm条件下培养,发酵148h结束,测定菌体密度(od600),取10ml菌液提取菜油甾醇。
菜油甾醇提取方法:同上。
试验结果:
以葡萄糖为碳源,耶氏解脂酵母菌株sybe_yl02060056、sybe_yl02060063~sybe_yl02060065,sybe_yl02060067~sybe_yl02060076摇瓶发酵产量比较实验气相色谱-质谱联用(gc-ms)检测原始数据如表3:
表3:以葡萄糖为碳源摇瓶发酵产量
由表3及图4~6,菌株sybe_yl02060056、sybe_yl02060063~sybe_yl02060065,sybe_yl02060067~sybe_yl02060076的菜油甾醇产量来看,以葡萄糖为碳源发酵148h,所有乙酰coa代谢相关基因的过表达均对菜油甾醇的产量没有明显影响。
以葵花籽油为碳源,耶氏解脂酵母菌株sybe_yl02060056、sybe_yl02060063~sybe_yl02060065,sybe_yl02060067~sybe_yl02060076摇瓶发酵产量比较实验气相色谱-质谱联用(gc-ms)检测原始数据如表4:
表4:以葵花籽油为碳源摇瓶发酵产量
而由附表4及图5菌株sybe_yl02060056、sybe_yl02060063~sybe_yl02060065,sybe_yl02060067~sybe_yl02060076的菜油甾醇产量来看,以葵花籽油为碳源发酵148h,所有菌株的菜油甾醇产量均高于以葡萄糖为碳源时,因此葵花籽油是耶氏解脂酵母合成菜油甾醇的一种优选碳源。以葵花籽油为碳源过表达acl、pox2和hmgr均使菜油甾醇的含量有明显提高,菌株sybe_yl02060065中过表达acl基因使菜油甾醇产量从22.7±0.3mg/gdcw提高到25.3±0.1mg/gdcw(提高11.5%),菌株sybe_yl02060069中过表达pox2基因使菜油甾醇产量从22.7±0.3mg/gdcw提高到24.7±0.1mg/gdcw(提高8.8%),菌株sybe_yl02060063中过表达hmgr基因使菜油甾醇产量从22.7±0.3mg/gdcw提高到25.8±2.8mg/gdcw(提高13.7%)。该结果表明,以葵花籽油为碳源时,耶氏解脂酵母中过表达内源acl、pox2或hmgr对于菜油甾醇产量提升具有积极作用,可进一步用于发酵工艺优化实验。
4、耶氏解脂酵母菌株sybe_yl02060065、sybe_yl02060069及sybe_yl02060063发酵罐发酵工艺优化的菜油甾醇产量
实验材料:耶氏解脂酵母菌株sybe_yl02060065、sybe_yl02060069和sybe_yl02060063
试验方法:
一级、二级种子培养基:22g/l葡萄糖、20g/l蛋白胨、10g/l酵母浸粉;
三级种子、发酵培养基:3.7%v/v葵花籽油、20g/l蛋白胨、10g/l酵母浸粉
将上述菌株sybe_yl02060065、sybe_yl02060069和sybe_yl02060063接种于5ml一级种子培养基中,在28℃、250rpm培养24h,以初始菌体浓度od600=0.2分别接种于二级种子培养基中,在28℃、220rpm培养24h,再以初始菌体浓度od600=0.2分别接种于50ml三级种子培养基中,每株菌株接种5瓶,于28℃、220rpm条件下培养12h,将全部250ml菌液倒入2.5l发酵培养基中。控制5-l发酵罐中发酵参数,温度、ph值、容氧量及搅拌速率分别控制在28℃、5.5、>30%和1vvm,监测发酵罐中剩余油量,采用碳源限制策略,待葵花籽油耗尽时开始滴加,调节滴加速率使油浓度限制在2g/l以下。发酵142h结束,每隔12h取15ml发酵液,测定菌体密度(od600),发酵液残余油量及菜油甾醇产量。
菜油甾醇提取方法:同上。
耶氏解脂酵母菌株sybe_yl02060065、sybe_yl02060069及sybe_yl02060063发酵罐工艺优化实验气相色谱-质谱联用(gc-ms)检测原始数据如表5
表5:耶氏解脂酵母菌株发酵罐工艺产量
由附表5及图6菌株sybe_yl02060065、sybe_yl02060069和sybe_yl02060063的发酵曲线来看,发酵46h,发酵罐中初始加入的葵花籽油耗尽流加开始。此时三个菌株均进入平台期,菌株sybe_yl02060065、sybe_yl02060069和sybe_yl02060063生物量分别达到20.24mg/gdcw、26.40mg/gdcw和18.34mg/gdcw,说明碳源限制策略很好地控制了菌体生物量的增长,碳代谢流更多地进入菜油甾醇及脂滴的积累。最终,过表达acl基因的菜油甾醇产量为800.8mg/l,过表达pox2基因的菜油甾醇产量为941.5mg/l,过表达hmgr基因的菜油甾醇产量为811.8mg/l,均为国际领先水平。对菜油甾醇产量最高菌株(sybe_yl02060069)进行菌种保藏,保藏编号为cgmccno.14039。
以上仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
sequencelisting
<110>天津大学
<120>重组菌株及其构建方法与在产菜油甾醇中的应用
<130>mp1701565
<160>1
<170>patentinversion3.3
<210>1
<211>1437
<212>dna
<213>人工序列
<400>1
atgatggcctccgatcgagtcagaaagcgacacaagggctccgctaacggcgctcagacc60
gtggagaaagaaccctccaaggaacccgcccaatggggtcgagcctgggaagtcgattgg120
ttctccctctccggcgtcattctgctcctgtgctttgccccctttctggtgtccttcttc180
atcatggcctgtgatcagtaccagtgctccatttcccaccctctgctcgacctgtataac240
ggcgacgccaccctgttcaccatctggaaccgagctccctcctttacctgggctgccgcc300
aagatctacgccatttgggtcaccttccaggtcgtgctctatatgtgcgtccctgacttc360
ctccacaagatcctccccggttacgtgggcggcgtgcaagacggtgccagaactcccgcc420
gggctgattaacaagtacgaggtcaacgggctgcagtgttggctgatcacccacgtgctc480
tgggtcctgaacgcccagcacttccactggttctcccctaccatcatcatcgacaactgg540
atccccctgctgtggtgcaccaatatcctcggctacgctgtctccaccttcgctttcatc600
aaagcctacctcttccccaccaaccccgaggactgcaagttcaccggcaacatgttctat660
aactacatgatgggcatcgagtttaacccccgaatcggtaagtggttcgatttcaagctg720
ttctttaacggccgacccggcattgtcgcctggactctgatcaatctgtcctacgctgcc780
aagcagcaggagctctacggctacgtcaccaactccatgattctcgtgaacgtcctgcag840
gccgtctatgtcgtcgacttcttttggaacgaggcttggtacctcaagactatcgacatc900
tgtcacgaccacttcggctggtacctgggttggggcgattgtgtgtggctgccttttctg960
tacaccctgcaaggcctgtacctggtgtataaccccatccagctctccactccccatgcc1020
gctggtgtcctgatcctcggcctggtcggctactatatcttccgagtcaccaaccatcag1080
aaagacctcttccgacgaaccgagggcaactgctccatctggggcaagaaacccaccttc1140
atcgaatgctcctaccaatccgccgatggcgccatccacaaatccaagctgatgacctct1200
ggcttctggggcgtggcccgacacatgaactacaccggcgacctcatgggctccctcgct1260
tactgtctcgcttgcggcggtaaccatctgctgccctacttctacatcatctacatgacc1320
atcctcctggtgcatcgatgcattcgagatgagcaccgatgctccaacaaatacggcaaa1380
gactgggagcgatacaccgctgctgtctcttaccgactgctgcccaatatcttctaa1437
- 还没有人留言评论。精彩留言会获得点赞!