植物茎杆持汁性调控基因Dry及其编码蛋白的制作方法
本发明涉及一种植物茎杆持汁性调控基因dry及其编码蛋白质序列,属于基因工程领域。
背景技术
植物体内的水分状况涉及许多重要的植物生理活动,但目前对于水分在植物体内的吸收、储藏和转运的研究尚不够深入。甜高粱作为一种新的能源作物,可以用于生物乙醇、糖浆和青贮饲料的生产制作。在其诸多影响因素中,水分含量是极为重要的一个方面。高粱基因组测序和重测序工作的陆续完成,为我们通过正向遗传学手段分析和研究植物体调控其体内水分的机理提供重要的基础。
技术实现要素:
本发明提供一种调控高粱茎秆持汁性的基因dry的dna序列、cdna序列和该基因所编码功能蛋白的氨基酸序列,所述基因的正常表达会使植物体内水分含量显著降低。
本发明的第一个发明目的是:提供一种调控植物持汁性的新基因dry,其特征在于,是选自如下1)或2)或3)或4)或5)的dna分子:
1)seqidno.1所示的dna分子(克隆自中国地方高粱品种ji2731的基因组dna):该dna分子由4155个核苷酸组成,自5′末端第1-169位核苷酸为第一个外显子,第170-1571位核苷酸为第一个内含子,第1572-1837位核苷酸为第二个外显子,第1838-3678位核苷酸为第二个内含子,第3679-4155位核苷酸为第三个外显子;
2)seqidno.2所示的dna分子(克隆自中国地方高粱品种ji2731的cdna);
3)在seqidno.1基础之上经过一至数个碱基替换和\或一至数个碱基的插入和\或缺失以及大片段的核苷酸序列插入\缺失\移位\倒位所形成能够影响植物生育能力的dna分子;
4)在0.1×sspe(或0.1×ssc)、0.1%(w/v)sds的溶液中,65℃条件下杂交并洗膜,能够与seqidno.2的dna分子杂交且编码植物水分调控相关蛋白的dna分子;
5)与seqidno.2的dna分子具有70%以上的同源性且编码植物水分调控相关蛋白的dna分子。
本发明的第二个发明目的是:提供上述基因所编码的调控植物茎杆持汁性的相关蛋白质。
在一个实施方案中,所述基因编码如下1)或2)所述的蛋白质:
1)序列表的seqidno.3所示的氨基酸序列组成的蛋白质;
2)将序列表的seqidno.3经过一个或数个氨基酸残基的取代、和/或缺失、和/或添加且具有影响植物水分含量的相关活性的蛋白质。
本发明的第三个发明目的是:提供含有所述基因的重组表达载体、表达盒、转基因细胞系或重组菌。
本发明第四个发明目的是:提供上述基因在用于转基因改良作物的用途。
在一个实施方案中,所述基因用于调控植物茎秆持汁性,使茎秆水分含量显著降低。
在一个具体实施方案中,所述基因使植物体纤维素含量降低,半纤维素含量升高。
在一个更加具体的实施方案中,所述作物包括但不限于高粱、玉米、水稻、谷子及短柄草。
本发明第五个发明目的是:提供了一种在其它植物中获取dry基因的直系同源基因的方法,以及利用该方法获得的玉米(zeamays)、黍(panicumhallii)、柳枝稷(panicumvirgatum)、狗尾草(setariaviridis)、谷子(setariaitalica)、短柄草(brachypodiumdistachyon)和水稻(oryzasativa)的氨基酸序列。
与现有技术相比,本发明具有如下的有益效果:本发明提供的植物茎杆持汁性调控基因dry直接参与植物体内水分的转运过程,该基因的表达受到抑制后,植物体内水分含量升高,维管束细胞发育不正常。通过植物生物技术途径,本发明在生物乙醇、糖浆和青贮饲料的生产制作中都将发挥重大作用。
术语定义
术语“植物茎杆持汁性调控基因”指一段具有编码蛋白能力的核苷酸序列,该序列特异编码具有调控植物水分含量的蛋白活性多肽,如seqidno.2的第1-912位核苷酸序列及其简并序列。
术语“简并序列”是指,位于seqidno.2的编码框第1-912位核苷酸序列中,有一个或多个密码子被编码相同氨基酸的简并密码子所取代后而产生的序列。由于密码子的简并性,所以与seqidno.2的第1-912位核苷酸序列同源性低至约70%的简并序列也能编码出seqidno.2所编码的氨基酸序列。
所述“植物茎杆持汁性调控基因”,还包括在中度严格条件下,更佳的在高度严格条件下可以与seqidno.2的核苷酸序列杂交的核苷酸序列。其中,中等严格条件可以是0.1×sspe(或0.1×ssc)、0.1%sds(w/v)的溶液中,65℃条件下杂交并洗膜的条件。
所述“植物茎杆持汁性调控基因”,还包括与seqidno.2中从第1-912位核苷酸序列的同源性至少70%,较佳地具有至少80%、82%、85%、86%、88%、89%同源性,更佳地具有至少90%、91%、92%、93%、94%同源性,最佳地具有至少95%、96%、97%、98%、99%同源性的核苷酸序列。
所述“植物茎杆持汁性调控基因”,还包括能编码具有与天然的茎杆持汁性基因dry相同功能蛋白的、seqidno.2开放阅读框序列的变异形式。这些变异形式包括(但并不限于):1个或几个核苷酸的缺失、插入和/或取代,以及在5′或3’端添加几个(通常为60个以内,较佳地为30个以内,更佳地为10个以内,最佳地为5个以内)核苷酸。
所述“植物茎杆持汁性调控基因”,还包括能够翻译一类具备调控水分含量的氨基酸序列,如seqidno.3的氨基酸序列。该类氨基酸序列还包括具有与天然茎杆持汁性蛋白相同功能的seqidno.3的变异形式。这些变异形式包括(但并不限于):1个或几个氨基酸的缺失、插入和/或取代,以及在c末端/或n末端添加一个或几个(通常为20个以内,较佳地为10个以内,更佳地为5个以内)氨基酸。在本领域中,用性能相近或相似的氨基酸进行取代时,通常不会改变蛋白质的功能;在c末端和/或n末端添加一个或数个氨基酸通常也不会改变蛋白质的功能。
另外,所述的“植物茎杆持汁性调控基因”的核苷酸全长序列或其片段通常可以用pcr扩增法、重组法或人工合成的方法获得。对于pcr扩增法,可根据本实施例公开的有关核苷酸序列,尤其是开放阅读框序列来设计引物,并用市售的cdna库或按本领域技术人员已知的常规方法所制备的cdna库作为模板,扩增得到有关序列。一旦获得了有关的序列,可以用重组法来大批量地获得有关序列。通常是将其克隆入载体,然后细胞转化等常规方法从增殖的宿主细胞中分离得到有关序列。此外,还可通过化学合成将突变引入实施例蛋白序列中。除了用重组法产生之外,实施例蛋白的片段还可用固相技术,通过直接合成多肽而加以生产。在体外合成蛋白质可以用人工或自动进行,可以分别化学合成实施例蛋白的各片段,然后用化学方法加以连接以产生全长的蛋白质分子。
附图说明
图1为无汁和持汁高粱的茎秆形态学观察及生理生化分析。(a)茎秆横切面的表型观察(白色标尺代表1厘米),其中茎秆无汁高粱ji2731(左)、茎秆持汁高粱e-tian(中)及杂交后代f1植株(右);(b)茎秆无汁和持汁高粱从拔节期至乳熟期植株茎秆含水量变化,nl:拔节期(第九片叶出现),fls:旗叶期,hs:抽穗期,fs:开花期,af:开花后一周,ms:乳熟期;(c)成熟期高粱茎秆含水量统计,包括茎秆无汁高粱ji2731、茎秆持汁高粱e-tian及重组自交系植株材料(茎秆无汁交换单株和茎秆持汁交换单株),括号内数字代表单株数目。(d至n)高粱茎秆显微观察。高粱茎秆髓部维管束和薄壁细胞半薄切片观察,茎秆无汁材料ji2731(d和e)和茎杆持汁甜高粱e-tian(f和g)。高粱茎秆髓部维管束超薄切片显微观察,ji2731(h)和e-tian(k)。维管束导管周围薄壁细胞显微观察(与白框包括的横切面对应),ji2731(i)和e-tian(l)。细胞壁厚度显微观察(与白框包括的横切面对应),ji2731(j)和e-tian(m)。(n)导管周围薄壁细胞的细胞壁厚度测量。(o和p),茎秆无汁和持汁高粱的茎秆组份分析。(o)纤维素含量测定;(p)木聚糖(半纤维素主要组成成分)含量测定。air,醇不溶物。双边t检验确定统计显著性,*p<0.05,**p<0.01。
图2为dry基因图位克隆。(a)dry基因初定位,ji2731(无汁)与e-tian(有汁)进行正反交,选取198个f2分离群体的交换单株和30对pav和ssr分子标记将dry位点定位到高粱第6号染色体3.25-mb的区间内;(b)dry基因精细定位,利用3343个f2~6的重组自交系将dry基因精细定位到42-kb的物理距离区间(m10-m11标记之间),白色长矩形代表e-tian基因型,黑色长矩形代表ji2731基因型,白色加黑点长矩形代表杂合基因型;(c)dry基因所在物理图谱区间内功能基因示意图,区间内包含四个基因,粉色框代表候选基因sobic.006g147400。
图3为dry基因附近基因组序列变异分析及dry基因表达分析。(a)设计16对pcr引物扩增dry基因上游7-kb至下游2-kb区域的基因组序列,发现以ji2731和显性交换单株的基因组dna为模版,除第五对引物p5外,其余的引物均扩增出目的条带,其中p10和p15两个pcr产物的长度大于以参考基因组btx623为模版的产物。而e-tian及隐性交换单株,从p5至p15均未获得pcr产物,包含dry基因前两个外显子及内含子约3.6-kb的序列,推测此处片段发生序列变异或者丢失,引起dry基因功能丧失。(b)dry基因位点附近基因组序列变异示意图。与无汁高粱品种ji2731的dry基因组序列相比,btx623参考基因组包含两个大片段缺失,其中1.8-kb缺失位于该基因的第二个内含子区段,3.0-kb的缺失发生在dry基因上游2.5-kb区域;相比ji2731,e-tian基因组上dry基因区段发生较大变化,推测发生约9.1-kb的缺失(包含dry基因组序列约3.6-kb)。(c)以ji2731、e-tian、显性交换单株(ril37-18)和隐性交换单株(ril41-11)为模版进行dry基因表达分析,sbactin为内参基因。
图4为dry基因全长cdna的序列比对。四条序列分别为:以ji2731和btx623为模版,race方法克隆dry基因的全长cdna;phytozome网站下载的btx623参考转录本序列;高粱转录组morokoshi数据库下载的dry基因转录本序列。蓝色下划线代表utr区域序列,橙色下划线代表dry基因编码区dna序列。彩色矩形代表起始密码子和终止密码子。
图5为dry蛋白亚细胞定位及原位杂交分析。(a)dry与gfp融合蛋白在烟草表皮细胞进行亚细胞定位分析,左:绿色荧光蛋白,右:明场。阳性对照p35s::gfp,标尺40μmp35s::dry-gfp图的标尺10μm。(b)dry基因高粱茎秆原位杂交分析,左:反义探针;右:正义探针(对照)。标尺10μm。
图6为dry蛋白的转录活性确定及自激活活性区间分析。trp:色氨酸;his:组氨酸;ade:腺嘌呤;x(x-α-gal):酵母半乳糖苷酶(mel1)的显色底物;nam(noapicalmeristem):dry蛋白第11到156位氨基酸,包含五个亚结构域;1-304:包含dry蛋白第1到304位氨基酸;1-268:包含dry蛋白第1到268位氨基酸;1-239:包含dry蛋白第1到239位氨基酸;1-235:包含dry蛋白第1到235位氨基酸;1-226:包含dry蛋白第1到226位氨基酸;1-209:包含dry蛋白第1到209位氨基酸;1-156:包含dry蛋白第1到156位氨基酸;157-304:包含dry蛋白第157到304位氨基酸;252-304:包含dry蛋白第252到304位氨基酸;260-304:包含dry蛋白第260到304位氨基酸;283-304:包含dry蛋白第283到304位氨基酸。
图7为高梁成熟期不同组织部位dry基因表达情况。(a)dry基因在高梁ji2731不同组织部位的表达情况。(b)dry基因在高梁ji2731和e-tian成熟期不同组织部位的半定量表达谱分析。lf,叶片;lv,叶脉;sp,茎髓部;ess,茎皮;esi,节间茎皮;imp,节间茎髓;pa,幼穗;rt,根。相对表达量:设叶片的dry基因表达量为1,其余不同组织中dry基因表达均与其进行比较。三个生物学重复,统计学显著性用字母表示。
图8为高梁ji2731抽穗期茎秆不同节间的dry基因定量pcr相对表达分析。相对表达量:没茎秆第一节(底部)dry基因相对表达量为1,其余不同节的dry基因表达均与第一节进行比较。三个生物学重复,统计学显著性用字母表示。
图9为dry基因转基因互补实验。(a)dry基因表达载体构建。pc系列为dry基因自身启动子及其编码区;pg系列为玉米ubi启动子及dry基因的基因组序列。空载体为pcambia3300。高粱转基因受体材料为甜高粱keller,该材料与e-tian具有相同的dry基因缺失。筛选标记基因为barstar。(b)高粱转基因t1代植株表型观察、分子检测及茎秆含水量测定。
图10为茎秆无汁高粱ji2731和持汁高粱e-tian和keller中dry基因组序列比对分析。蓝色矩形代表nam结构域,红色下划线代表外显子区域序列。
图11为高粱t2代转基因植株与对照keller农艺性状调查分析,杆重(a),株高(b),茎粗(c),穗重(d),穗长(e)和穗柄长(f)。
图12为高粱转基因材料与对照材料高粱茎秆髓部细胞显微观察。高粱茎秆髓部薄壁组织观察,keller(a)和pc4(b)。维管束显微观察,keller(c)和pc4(f)。维管束导管周围薄壁细胞显微观察(与白框包括的横切面对应),keller(d)和pc4(g)。细胞壁厚度显微观察(与白框包括的横切面对应),keller(e)和pc4(h)。(i)导管周围薄壁细胞的细胞壁厚度测量。(j和k)转基因与对照高粱的茎秆组份分析。(j)纤维素含量测定;(k)木聚糖(半纤维素主要组成成分)含量测定。air,醇不溶物。双边t检验确定统计显著性,*p<0.05,**p<0.01。
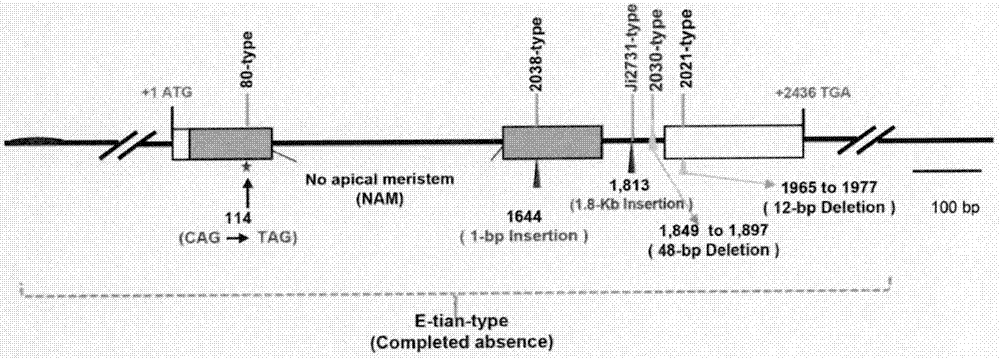
图13为dry基因单倍型分析。茎秆无汁高粱两种单倍型ji2731-type和2030-type;有汁高粱单倍型导致dry基因功能失能,分别为e-tian-like单倍型的dry基因缺失,2038-type单倍型1644位有一个碱基g插入,导致移码,2021-type单倍型在基因保守结构域缺失12个碱基,80-type单倍型在第一个外显子的114位有一个碱基突变,导致编码提前终止。
图14为dry基因翻译的氨基酸序列与其在高粱、水稻、玉米、谷子、黍、二穗短柄草、柳枝稷和狗尾草中直系同源基因翻译产物的进化树分析。clustalw进行多重序列比对,mega6.0软件中的邻位相连法(neighbor-joining)对dry基因的同源基因的氨基酸序列构建进化树。
具体实施方式
以下的实施例便于更好地理解本发明,但并不限定本发明的应用范围。下述实施例中的所有技术和科学术语,如无特殊说明,均为本发明所属领域普通技术人员通常所理解的相同含义。除非有相反指明,本发明所使用或提及的技术均为本领域普通技术人员公认的标准技术。所述试验材料,如无特别注明,均为本发明领域通用的试验材料。下述实施例中所用的试验试剂,如无特殊说明,均为自常规生化试剂商店购买得到的。材料、方法和实施例谨作阐述用,而非加以限制。
本发明也包括利用序列表所述序列调控植株茎杆的持汁性,即利用本发明提供的基因序列在基因组、和/或转录组、和/或蛋白质组水平影响其它植物中相同或同源基因的功能来调控茎杆持汁性的目的。例如,下述方法但不限于下述方法:通过天然序列的变异导致基因表达抑制或蛋白质功能的丧失、通过向植物中转入所述基因的反义序列或引入发卡结构、或将所述基因与其它序列(dna或rna)相结合产生新的具有功能活性的dna或rna链,来影响或改变植物基因的功能。或其它本领域技术人员已知的可用于影响茎杆持汁性的技术方法中的任何一种技术方法。
本发明包括高粱dry基因,其显性等位基因对控制高粱茎秆持汁性具有关键作用,功能缺失性的隐性等位基因会导致茎秆内水分含量升高。该基因位于高粱6号染色体,其基因具体位置如图2所示。
该基因序列及其同源序列可从以下植物中获得,包括但不限于高粱(sorghumbicolor)、玉米(zeamays)、黍(panicumhallii)、柳枝稷(panicumvirgatum)、狗尾草(setariaviridis)、谷子(setariaitalica)、短柄草(brachypodiumdistachyon)和水稻(oryzasativa)。获得方法包括但不限于:通过dry基因序列利用blastx、blastn或通过氨基酸序列利用blastp从其它植物的基因组序列数据库、和/或cdna序列数据库、和/或蛋白质序列数据库中调取;以高粱dry基因的dna或cdna或rna序列为参考序列设计引物,直接从其它植物的基因组dna或cdna或rna中利用pcr的方法直接获得;以高粱dry的基因序列设计探针,利用核酸杂交的方法从基因组文库中分离含有同源基因序列的dna或cdna或rna片段。
“dry基因同源序列”指在与seqidno.3的氨基酸进行tblastn比较分析后,identities大于或等于35%、positives大于或等于50%,且识别区域位于seqidno.3的11至156位氨基酸即保守的nac结构域内的植物基因的dna序列。进行blastx时,所有参数均遵照http://blast.ncbi.nlm.nih.gov/所示的默认设置进行。下文通过说明和阐述提供了更为详细的描述,但这并非意欲对本发明的范围加以限制。
实施例1.茎杆持汁性的鉴定
从用途上来分,高粱可以分为粒用高粱、甜高粱、饲草高粱和帚高粱。与粒用高粱相比,甜高粱茎秆内含有大量的水分(图1.a)。我们通过以下两种方式对高粱茎杆的持汁性进行了鉴定的:一是用钳子挤压高粱茎杆节间,如果有汁液渗出,则判定高粱茎杆持汁,否则判定为无汁;二是在成熟期分别称量烘干前后高粱茎杆的重量,计算其含汁量。研究发现,在开花以后,高粱品种ji2731茎秆中的水分含量快速下降,而高粱品种e-tian茎秆中的水分含量保持在较高水平。(图1.b)。取高粱品种ji2731和e-tian的中部节间,经戊二醛和锇酸固定后进行细胞学观察。发现ji2731的薄壁组织细胞较为饱满充实,而e-tian的薄壁组织细胞极为不规则(图1.d至g)。同时ji2731的维管组织中导管细胞规则,导管周围细胞的细胞壁较厚,而e-tian的维管组织中导管不规则,导管周围细胞的细胞壁变薄(图1.h至m)。细胞壁厚度测量结果显示,e-tian的细胞壁厚度显著小于ji2731(图1.n)。对ji2731和e-tian的细胞壁成分进行化学分析,发现与ji2731相比,e-tian中纤维素的含量显著高于ji2731,而木聚糖的含量显著低于ji2731(图1.o,p)。
实施例2.dry基因精细定位
茎杆持汁性遗传分析,共构建了4个杂交组合,分别为ji2731×e-tian,e-tian×ji2731,keller×ji2731和btx623×ji2731。其中f1代茎秆的表型与ji2731相同,表现为茎秆无汁的表型(图1.a)。f2群体的大小分别为214、213、114和221个单株,相应的无汁单株与持汁单株的比例分别为150∶64,150∶63,84∶30,156∶65,卡方测验结果显示无汁单株和持汁单株符合3∶1的分离比(表1)。
表1.茎秆茎秆持汁性的分离比
在进行基因定位时,用到的杂交群体是e-tian×ji2731,其杂交后代在在含水量上有较大差异(图1.c)。在高粱的每条染色体上选取2-3对分子标记进行pcr扩增和凝胶电泳。结果显示:位于6号染色体上的两个分子标记m1(sm06069)和m2(6-40-2),表现为ji2731基因型的个体数均超过90%,表明它们与dry基因紧密连锁(图2.a)。利用单株传代的方式对群体进行扩繁,最终利用3343个单株,将dry基因的候选区间定位到分子标记m10(6-51797)和m11(6-51840)之间,区间大小为42kb(图2.b)。从网站phytozome上调取取测序品种btx623的基因组序列,发现在该42kb区间内共有4个功能基因(图2.c)。
实施例3.dry基因的克隆和序列分析
基于高粱btx623基因组序列信息,设计16对引物对两个杂交亲本ji2731和e-tian及显性交换单株37-18和隐性交换单株41-11中该基因进行扩增。其中在ji2731和显性交换单株37-18中,除去第五对引物p5无结果外,扩增共得到包括dry基因的编码区序列以及上游7-kb至下游2-kb区域的基因组序列,其中引物p10和p15对应pcr产物的长度大于以参考基因组btx623为模版的产物长度,而在e-tian和隐性交换单株41-11中,从p5至p15均未获得pcr产物,仅在引物p16处得到符合目的的条带(图3.a)。序列分析发现在ji2731中该基因全长4155bp(seqidno.1),与之相比,高粱参考基因组btx623包含两个大片段缺失,其中1.8-kb缺失位于该基因的内含子区段,3.0-kb的缺失发生在dry基因上游2.5-kb区域(图3.b)。序列进行blastn分析,表明其在高粱基因组内为单拷贝基因。
取两个杂交亲本ji2731和e-tian及显性交换单株37-18和隐性交换单株41-11黄化幼苗,用液氮速冻保存,用于提取总rna进行半定量分析。sbactin基因作为内参基因,引物cdse3_f(5′-agccatgggagcttccagac-3′)和cdse3_r:(5′-actgcgatgcgccggcaaa-3′),用于检测dry基因表达,引物sbactin:正向引物actin_f(5′-acattgccctggactacgac-3′)和反向引物actin_r(5′-aatgaaggatggctggaaga-3′),用于检测sbactin基因的表达。结果显示dry在ji2731和显性交换单株37-18有表达,而在e-tian和隐性交换单株41-11中无表达(图3.c)。
以ji2731和btx623的rna为模板,通过3′race和5′race方法,在两个亲本中扩增dry基因全长cdna序列(seqidno.2)。其中3′race经过两轮pcr,第一轮pcr的引物为3p1(5′-tgatgaacggagcgatgt-3′)和b26(5′-gactctagacgacatcga(t)18-3′),第二轮pcr过程中扩增ji2731的引物为3p2(5′-tctacgacgacagtggct-3′)和b25(5′-gactctagacgacatcga-3′),扩增btx623的引物为3p2-b(5′-ccgacgatgatggggcta-3′)和b25(5′-gactctagacgacatcga-3′);5′race是根据试剂盒5′fullracekit(takara,japan)的实验方案进行的,两条特异引物分别为gsp1(5′-ccacatcgctccgttcat-3′)和gsp2(5′-tgtcttgctcgctgtctttc-3′)。序列比对发现,dry基因在ji2731和btx623中存在核苷酸位点的差异,在btx623中dry基因第114位核苷酸存在一个碱基突变(c-t),导致编码提前终止(图4)。并且参考基因组btx623中关于dry基因cds序列的注释存在错误,将第一个外显子中的部分序列注释为内含子序列。。
通过对cdna序列进行模拟翻译,该基因编码含有304个氨基酸的蛋白(seqidno.3),将该蛋白序列与genbankswissprot蛋白质数据库进行blastp分析,显示该蛋白含有一个nac保守结构域。
通过dry基因在高粱品种ji2731中的基因组序列(seqidno.1)和cdna序列(seqidno.2)的比对分析,发现该基因的结构特征如下:含有3个外显子(exon)和2个内含子(intron),其中自5′末端第1-169位核苷酸为第一个外显子,第170-1571位核苷酸为第一个内含子,第1572-1837位核苷酸为第二个外显子,第1838-3678位核苷酸为第二个内含子,第3679-4155位核苷酸为第三个外显子。
实施例4.dry编码的蛋白是一个的转录因子
将dry的cds序列连接到载体pcambia1205,构建表达载体35s::dry-gfp。注射烟草后发现,dry蛋白仅在细胞核中存在表达(图5.a)。将dry的cds序列连接到载体pgbkt7,转化酵母菌株ah109,发现能够激活下游报告基因的表达,并且通过一些的序列分析发现,dry的激活区域位于236-259aa区域。以上试验说明dry编码的蛋白是一个的转录因子(图6)。
实施例5.dry的转录水平分析
取成熟期ji2731和e-tian的叶片、叶脉、茎皮、髓、节间皮、节间髓、幼穗和根,经液氮速冻保存,用于提取总rna;取抽穗期ji2731的节间,剥掉茎皮后,将髓用液氮速冻保存,用于提取总rna。上述样本均采取实时定量pcr(real-timepcr,rt-pcr)进行dry基因的表达量检测。qrt-pcr以sbactin基因作为内参报告基因,引物qpcr_f(5′-gcttcagggaccgcaagt-3′)和qpcr_r(5′-gctgtctttccgcttctgg-3′)用于在qrt-pcr反应中检测dry基因表达,引物actin_f(5′-acattgccctggactacgac-3′)和actin_r(5′-aatgaaggatggctggaaga-3′)用于检测sbactin基因的表达。qrt-pcr中每个样品的dry基因和sbactin基因的pcr反应均设置3个独立重复试验。
qrt-pcr显示在ji2731的叶片、叶脉、茎皮、髓、节间皮、节间髓、幼穗和根中均检测到dry基因的表达,并且在茎皮和髓中有较高的表达量(图7);在e-tian的上述组织中未检测到dry基因的表达。在ji2731的不同节间中均检测到dry基因的表达,并且在含水量较低的节间该基因的表达量较高。这也暗示dry基因表达调控茎秆含水量变化(图8)。
取播种后一月的ji2731和e-tian的幼嫩节间,经固定液faa固定后进行原位杂交试验。合成探针时用到的特异引物为insitu-f(5′-gacagcgagcaagacaacg-3′)和insitu-r(5′-caggccaccaataccgag-3′)。显色反应显示dry主要在ji2731的节间的薄壁组织和导管中有表达,而在e-tian的上述位置无表达(图5.b)。
实施例6.dry的功能互补试验
以载体pcambia3300为骨架,分别构建以dry基因自身启动子驱动的其编码区序列以及以玉米ubi启动子驱动的dry基因的基因组序列的载体(图9.a)。高粱转基因受体材料为持汁高粱品种keller,该材料与e-tian基因型相同,均表现为dry基因的缺失(图10)。对得到的转基因材料进行鉴定,发现转基因材料的含水量降低,分子检测显示在转基因材料中存在dry基因的表达(图9.b)。大田试验显示,除含水量外,转基因材料在杆重,株高,茎粗,穗重,穗长和穗柄长与keller相同(图11.a至f)。细胞学观察显示,与keller相比,转基因材料的薄壁细胞饱满充实(图12.a,b)。维管组织中导管规则,导管周围细胞的细胞壁较厚(图12.c至h)。细胞壁厚度测量结果显示,转基因材料的细胞壁厚度显著大于keller(图12.i)。细胞壁成分测定显示,转基因材料细胞壁中的纤维素含量低于keller,而木聚糖的含量高于keller(图12.j,k)。
实施例7.dry基因的自然变异分析
引物p14、p15和p16可用于在不同的高粱品系中特异克隆dry基因全长。三个引物组合能分别扩增该基因的-17~+1305、+1214~+3958、+3715~+4832片段,通过测序对扩增的基因序列进行验证。所有pcr扩增均使用kod-plus-neo(toyobo,japan),并按照产品说明的反应体系和条件,在abiveriti96wellthermalcycler进行pcr扩增。pcr产物送往北京生工生物工程公司进行测序。经序列比对发现,dry基因主要包括6种单倍型,其中茎秆无汁高粱有两种单倍型ji2731-type和2030-type;有汁高粱单倍型导致dry基因功能失能,其中在e-tian-type单倍型的dry基因缺失,2038-type单倍型在1644位有一个碱基g插入,导致移码;2021-type单倍型在基因保守结构域缺失12个碱基;80-type单倍型在第一个外显子114位一个碱基突变,导致编码提前终止(图13)。
实施例8.dry基因在玉米、黍、柳枝稷、狗尾草、谷子、短柄草和水稻中的同源性分析
利用dry基因的蛋白质序列,通过blastp获得了该基因在玉米、柳枝稷、黍、谷子、狗尾草、短柄草和水稻中的直系同源基因。玉米中两个同源基因为grmzm2g043813和grmzm2g081930,柳枝稷中两个同源基因为pavir.ga01251和pavir.gb00655,黍中同源基因为pahal.g01069,谷子中同源基因为seita.7g166500,狗尾草中同源基因为sevir.7g175600,短柄草中同源基因为bradi5g15587,水稻中同源基因为loc_os04g43560(图14)。
尽管本发明的具体实施方式已经得到详细的描述,本领域技术人员将会理解。根据已经公开的所有教导,可以对那些细节进行各种修改和替换,这些改变均在本发明的保护范围之内。本发明的全部范围由所附权利要求及其任何等同物给出。
- 还没有人留言评论。精彩留言会获得点赞!