一种可降解生物胺的多铜氧化酶重组酶的制作方法
本发明涉及一种可降解生物胺的多铜氧化酶重组酶,属于发酵技术领域。
背景技术:
酱油是我国及东南亚各国特有的调味品,以大豆、小麦或麸皮等为原料,利用米曲霉、酵母菌、乳酸菌等微生物,通过发酵作用酿成具有特殊色、香、味的液体调味品。酱油中含有丰富的蛋白质,大量的游离氨基酸,游离的氨基酸在氨基酸脱羧酶的作用下会生成生物胺。
生物胺是一类具有生物活性的小分子量含氮有机物的总称,是生物体内正常的生理活性物质,微量生物胺在机体细胞活动中发挥着重要作用,当人体摄入过量的生物胺尤其是同时摄入多种生物胺时,会引起诸如头痛、恶心、心悸、血压变化、呼吸紊乱等过敏反应,严重的还会危及生命。
目前主要从三个方面控制食品中的生物胺,1、从源头控制:控制游离氨基酸的含量,局限性在于会影响某些高蛋白食品的风味;采用无氨基酸脱羧酶活性的菌株代替原生产菌种,局限性在于仅适用于封闭的单菌发酵体系。2、过程控制:通过对生产过程中的原料、温度和盐度进行合理选择,抑制生物胺产生菌的生长,以达到抑制生物胺产生的目的,局限性在于加工温度和盐度等因素多由食物特性决定,而低温贮藏会提高设备和能耗费用,且有些微生物低温下也能产生生物胺;添加降解生物胺的菌株,局限性在于人们担心添加菌株的安全性以及可能会影响食物风味。3、末端消除:通过向发酵食品中添加生物胺降解酶,不损害食品营养,不产生新的毒性物质,局限性在于现有生物胺降解酶降解效果差,降解类型少,最适 pH偏酸性,不适用酱油等酸性发酵食品。
目前,利用生物胺降解酶降低酱油中的生物胺主要有三个方面的难点:第一,酱油中的环境偏酸性,添加进酱油的生物胺降解酶需要具有耐酸特性;第二、酱油中的环境是高盐环境,添加进酱油的生物胺降解酶需要具有耐盐特性;第三,酱油中生物胺种类多,添加进酱油的生物胺降解酶若专一降某种生物胺,则无法满足酱油中应用的需要。鉴于以上背景,急需找到一种降解效果好,最适pH偏酸性的生物胺降解酶来降低酱油中生物胺的含量。
多铜氧化酶是一类可通过其光谱学性质,同源性的序列及与底物的反应机理定义的一类含铜氧化酶家族,它将生物胺催化氧化成对应的醛、氨和水,达到降低生物胺含量的目的。由于野生酶无法满足应用的需求,因此,采用基因工程的手段提高酱油中生物胺的降解效率显得十分必要。
技术实现要素:
本发明的目的是提供一种可降解生物胺的多铜氧化酶重组酶,用多铜氧化酶降解生物胺的方法来降低生物胺,从而提供一种降低发酵食品中生物胺的方法。
本发明的第一个目的是提供一种基因工程菌,以大肠杆菌为宿主,表达SEQ ID NO.2所示的多铜氧化酶。
在本发明的一种实施方式中,以大肠杆菌BL21为宿主。
在本发明的一种实施方式中,以pET系列质粒为表达载体。
本发明的第二个目的是提供所述基因工程菌的构建方法,是将SEQ ID NO.1所示的基因序列与载体连接,转化至大肠杆菌细胞中。
在本发明的一种实施方式中,所述载体为pET28a。
本发明的第三个目的是提供一种多铜氧化酶的生产方法,所述方法是将所述基因工程菌接种至培养基中,于35~37℃培养30~48h,收集菌体细胞,破碎细胞获得酶液。
在本发明的一种实施方式中,所述方法将所述基因工程菌接种至LB培养基中,培养至 OD600为0.6~0.8时,加入IPTG诱导。
本发明的第四个目的是提供所述基因工程菌在食品领域降低生物胺方面的应用。
在本发明的一种实施方式中,所述应用包括降低发酵食品中的生物胺含量。
在本发明的一种实施方式中,所述应用是将所述基因工程菌生产的多铜氧化酶添加至酱油中,降解酱油中的生物胺。
在本发明的一种实施方式中,所述生物胺包括色胺、苯乙胺、腐胺、尸胺、组胺、酪胺、精胺、亚精胺中的至少一种。
有益效果:本发明提供的多铜氧化酶重组酶具有较强的降解生物胺的性能,降解谱广,能够在24h内降解色胺、苯乙胺、腐胺、尸胺、组胺、酪胺,精胺、亚精胺,降解率达2%~75%,较野生菌株中的多铜氧化酶酶活提高10~15倍,并且该重组酶耐酸,耐高盐,对色胺、苯乙胺、腐胺、尸胺、组胺、酪胺、精胺、亚精胺这几种食品中常见的生物胺具有较明显的降解作用,有助于使发酵食品的安全性得到进一步提高。
附图说明
图1为(副干酪乳杆菌)(Lactobacillus paracasei)中多铜氧化酶基因的验证图;其中,M Marker;1阴性对照;2-3Lactobacillus paracasei中多铜氧化酶基因;
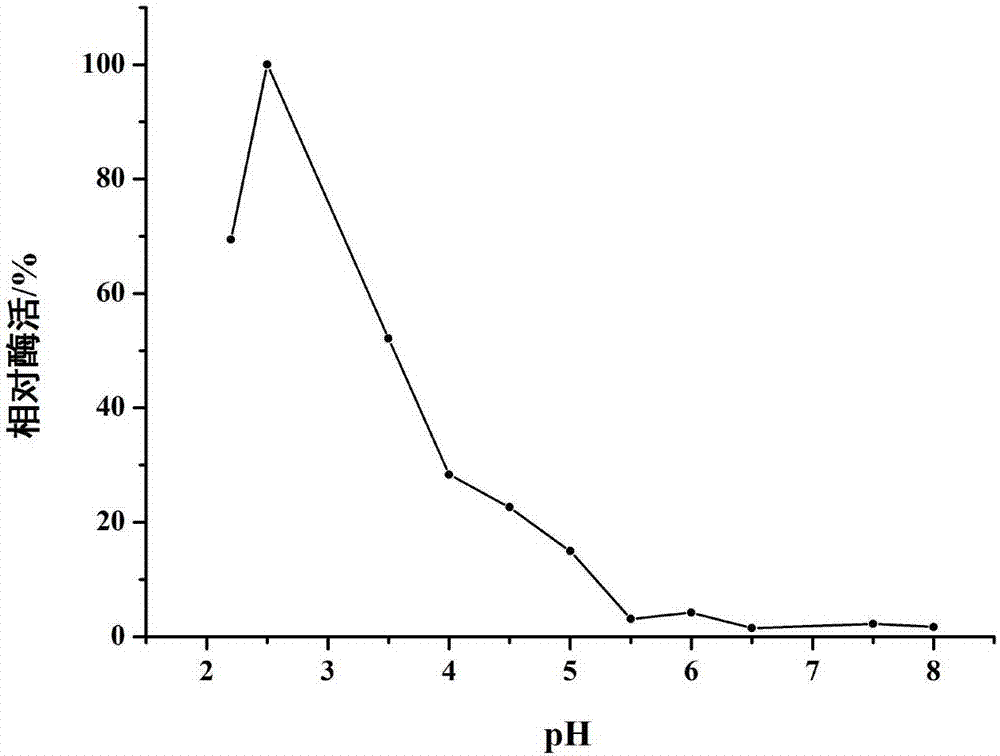
图2为不同pH下的多铜氧化酶酶活;
图3为不同盐浓度下的多铜氧化酶酶活。
具体实施方式
生物胺采用HPLC法进行检测,检测方法参考李志军《食品中生物胺及其产生菌株检测方法研究》。
生物胺降解率=(初始浓度一终浓度)/初始浓度。
实施例1
根据NCBI中的Lactobacillus paracasei strain L9(GenBank登录号为:CP012148.1)中的多铜氧化酶的基因序列,设计引物,以本研究室保藏的副干酪乳杆菌(Lactobacillus paracasei) 基因组为模板,扩增多铜氧化酶基因。扩增所需的引物如下:
上游:ATGAAAACCTATACGGACTATTTC
下游:TTA CATTTTCATCCCCATTT
PCR反应程序:95℃下先预变性3min,然后进行以下循环:95℃下变性30s,55℃下退火30s,72℃下延伸1min 30s;34个循环后,72℃下延伸10min,4℃下保温。
以Lactobacillus paracasei的基因组为模板,PCR扩增,扩增结束后由1%琼脂糖凝胶电泳显示扩增结果,如图1所示有1500bp左右的条带,与目的条带大小一致,表明Lactobacillus paracasei(副干酪乳杆菌)中含有多铜氧化酶基因,将PCR产物纯化测序,测序结果如SEQ ID NO.1所示。
实施例2
1、获得目的片段:以Lactobacillus paracasei的基因组为模板,PCR扩增结束后由1%琼脂糖凝胶电泳显示扩增结果,有1500bp左右的目的条带。用DNA胶回收试剂盒将PCR产物纯化,得到目的片段。
2、获得载体:将含有质粒pET-28a的大肠杆菌划线到含卡那霉素的LB平板上,培养12 h后,挑单菌落到20mL/250mL摇瓶LB培养基中(含卡那霉素),37℃下200r·min-1过夜培养,使用质粒提取试剂盒提质粒。
3、酶切连接:根据基因和载体的序列选择合适的限制性内切酶EcoRI、HindIII,37℃金属浴中反应45min。
表1 50μL限制性内切酶酶切反应体系
酶切后的基因片段和质粒经过回收纯化后,按摩尔比4~10:1的比例混合,通过Solution I连接酶进行连接反应,于16℃金属浴中连接过夜。
4、转化:将感受态细胞大肠杆菌JM109(克隆宿主)加入5μL连接产物,均匀混合后于冰中静置30min。42℃水浴90s后立即取出于冰中放置2-5min。加入700μL LB培养基摇床(37℃,200r·min-1)振荡培养1h。4000r·min-1离心2min,弃去大部分上清液,留100μL 左右上清液重悬菌体。菌液均匀涂布于含有卡那霉素的平板,置于37℃培养箱中过夜培养。次日挑取单菌落,通过菌落PCR筛选阳性转化子。
5、筛选出含重组质粒的转化菌落,提质粒,酶切验证。
6、选2~3个酶切验证结果正确的重组质粒送至上海生工测序,挑选测序正确的重组质粒转化至大肠杆菌BL21感受态细胞中,获得重组菌。
7、将重组菌接种到LB培养基中,于37℃、220r/min条件下过夜培养。将种子液按 3%的接种量转接到TB培养基中,于37℃、220r/min条件下培养至OD600为0.6~0.8,加入终浓度为0.1mM的Cu2+并且加入终浓度为0.1mM的IPTG,20℃振荡培养20h。将培养后的培养液于7000r/min离心5min收集菌体,按原体积加入pH为7.4的PBS缓冲液,洗涤菌体,重复2次,超声波破碎菌体细胞,将得到的破碎液,5000r/min离心15min,收集上清液,上清液即为多铜氧化酶重组酶粗酶液,经测定,酶活为27.4U/mL。
Lactobacillus paracasei strain L9对生物胺的降解率为1~17%,多铜氧化酶酶活为 3.17U/mL,酶活低,降解效果差。
实施例3
取2ml粗酶液与2mL某品牌市售酱油进行混合,将混合物置于37℃培养箱中静置48h 后,于4℃、10000r/min离心5min取上清液,用HPLC法检测生物胺的含量。对照组以PBS 缓冲液代替粗酶液与酱油混合共置。
表1酱油中生物胺的含量
实施例4多铜氧化酶重组酶的酶学性质
(1)多铜氧化酶的最适pH
酶促反应分别在不同pH值的缓冲液(柠檬酸盐缓冲液pH2.2~6.0、磷酸盐缓冲液pH 6.0~8.5)中反应,反应结束测定不同pH条件下的酶活,确定最适反应pH。以最高的酶活为100%。结果如图2所示,pH为2.5时酶活最高,pH为3时,酶活还能保持在最适pH下酶活的50%以上。
(2)盐浓度对多铜氧化酶酶活的影响
将粗酶液在不同质量分数的NaCl缓冲液中保温1h(对照组不添加NaCl),测定酶活,以未添加NaCl测得酶活为100%,计算各NaCl质量分数条件下相对酶活,确定NaCl对酶活稳定性的影响。如图3所示,随着NaCl浓度的增加酶活降低,当NaCl质量分数达到15% (Na+浓度2.5mmol/L),对酶有激活作用,酶活提高了4.67倍。
虽然本发明已以较佳实施例公开如上,但其并非用以限定本发明,任何熟悉此技术的人,在不脱离本发明的精神和范围内,都可做各种的改动与修饰,因此本发明的保护范围应该以权利要求书所界定的为准。
SEQUENCE LISTING
<110> 江南大学
<120> 一种可降解生物胺的多铜氧化酶重组酶
<160> 4
<170> PatentIn version 3.3
<210> 1
<211> 1530
<212> DNA
<213> 人工序列
<400> 1
atgaaaacct atacggacta tttctttgat gagccagcgt ttgatctcca cgatggcggg 60
tacgtgccgc ttgaggtcag tgatgcgcct gagaagccct taaatgtgcc gccgttgttg 120
aaaccggata aagagacagc gaccgacgtt tattacacgg tgacagcaga ggctggggaa 180
acacaactgt tgccaggggc gaagaccaag acgtggggct ataataccag tctgttaggt 240
cagacgattg tgtatcgccg tggtcagcat acgcatgtga cactgaaaaa caccttgcct 300
gagttgacca cttttcattg gcatggggct aatgtcagtg gtccttatgt tgatggcgga 360
tgccatgcgc cggtttatcc gggtgaaagt aagcatatcg acttcacgtt ggaccaaccg 420
gcgacgactt tgtggctgca cgcgcatccg tgcccatcca cagccgagca ggtttggcat 480
ggtttggctg ccatggtgat tgtcaaagac gaccatgaag ccagcctgcc attgccaagg 540
aactatggcg ttgacgatat tccggtcatt ttgcaagacc ggcgttttca cgagaacaac 600
cagtgggact accgggctga ttatgatcct gacggtgttg ctgggccaac cgcaatgatt 660
aacggtacaa tcaatcccta ttttgatgtc accacgcaaa aggtccggtt gcgttttctg 720
gatggtgcta atcgccgtga atggcggttg catttttccg atgacctacc atttacgcaa 780
attggcggag atggctcatt gttaccggag ccggtcaaat tcacccatct gatgctgact 840
tgtgctgagc gtgccgaagt gattgttgat tttggccaat accatgaagg cgacgaggtc 900
accttatata cggatgatgt gccattgcta aagttccgca ttcatgcgtt caaaccggat 960
cagacaactt tgcctgataa gttgttcgac gtgaaggcgc ctgtggttga tccggctttg 1020
ccagttcgcc atgttgtgat gcaggggatg gacgaaggtg ttgcgattga cggtaaaaag 1080
ttcgccatgc agcggattga tgccacgcag ccaattggca aagcccagta ctgggatgtt 1140
accaatagca atgatgcgcc tggaatggtt catccattcc atgtgcatgg aacccaattc 1200
ttagtcttgt cgcggaatgg gcatgcgccg tatccaaatg aacatggatt taaagatacc 1260
gttggcgtaa atcccgggga aacggttcga ctgctggttc gctttgattt gccaggggtt 1320
tatatgtatc actgccatat cattgaacac gaagatggcg gcatgatggc gcagattgaa 1380
acttttgatc cagccaagcc gaagcaagaa tataagttga tggatatgga tacgttaatg 1440
atggccttgg ctaaagaacg tggcgtcaaa ccatctgaga tttggatggg cggtatgcag 1500
tcttatgaaa aaatggggat gaaaatgtaa 1530
<210> 2
<211> 509
<212> PRT
<213> 人工序列
<400> 2
Met Lys Thr Tyr Thr Asp Tyr Phe Phe Asp Glu Pro Ala Phe Asp Leu
1 5 10 15
His Asp Gly Gly Tyr Val Pro Leu Glu Val Ser Asp Ala Pro Glu Lys
20 25 30
Pro Leu Asn Val Pro Pro Leu Leu Lys Pro Asp Lys Glu Thr Ala Thr
35 40 45
Asp Val Tyr Tyr Thr Val Thr Ala Glu Ala Gly Glu Thr Gln Leu Leu
50 55 60
Pro Gly Ala Lys Thr Lys Thr Trp Gly Tyr Asn Thr Ser Leu Leu Gly
65 70 75 80
Gln Thr Ile Val Tyr Arg Arg Gly Gln His Thr His Val Thr Leu Lys
85 90 95
Asn Thr Leu Pro Glu Leu Thr Thr Phe His Trp His Gly Ala Asn Val
100 105 110
Ser Gly Pro Tyr Val Asp Gly Gly Cys His Ala Pro Val Tyr Pro Gly
115 120 125
Glu Ser Lys His Ile Asp Phe Thr Leu Asp Gln Pro Ala Thr Thr Leu
130 135 140
Trp Leu His Ala His Pro Cys Pro Ser Thr Ala Glu Gln Val Trp His
145 150 155 160
Gly Leu Ala Ala Met Val Ile Val Lys Asp Asp His Glu Ala Ser Leu
165 170 175
Pro Leu Pro Arg Asn Tyr Gly Val Asp Asp Ile Pro Val Ile Leu Gln
180 185 190
Asp Arg Arg Phe His Glu Asn Asn Gln Trp Asp Tyr Arg Ala Asp Tyr
195 200 205
Asp Pro Asp Gly Val Ala Gly Pro Thr Ala Met Ile Asn Gly Thr Ile
210 215 220
Asn Pro Tyr Phe Asp Val Thr Thr Gln Lys Val Arg Leu Arg Phe Leu
225 230 235 240
Asp Gly Ala Asn Arg Arg Glu Trp Arg Leu His Phe Ser Asp Asp Leu
245 250 255
Pro Phe Thr Gln Ile Gly Gly Asp Gly Ser Leu Leu Pro Glu Pro Val
260 265 270
Lys Phe Thr His Leu Met Leu Thr Cys Ala Glu Arg Ala Glu Val Ile
275 280 285
Val Asp Phe Gly Gln Tyr His Glu Gly Asp Glu Val Thr Leu Tyr Thr
290 295 300
Asp Asp Val Pro Leu Leu Lys Phe Arg Ile His Ala Phe Lys Pro Asp
305 310 315 320
Gln Thr Thr Leu Pro Asp Lys Leu Phe Asp Val Lys Ala Pro Val Val
325 330 335
Asp Pro Ala Leu Pro Val Arg His Val Val Met Gln Gly Met Asp Glu
340 345 350
Gly Val Ala Ile Asp Gly Lys Lys Phe Ala Met Gln Arg Ile Asp Ala
355 360 365
Thr Gln Pro Ile Gly Lys Ala Gln Tyr Trp Asp Val Thr Asn Ser Asn
370 375 380
Asp Ala Pro Gly Met Val His Pro Phe His Val His Gly Thr Gln Phe
385 390 395 400
Leu Val Leu Ser Arg Asn Gly His Ala Pro Tyr Pro Asn Glu His Gly
405 410 415
Phe Lys Asp Thr Val Gly Val Asn Pro Gly Glu Thr Val Arg Leu Leu
420 425 430
Val Arg Phe Asp Leu Pro Gly Val Tyr Met Tyr His Cys His Ile Ile
435 440 445
Glu His Glu Asp Gly Gly Met Met Ala Gln Ile Glu Thr Phe Asp Pro
450 455 460
Ala Lys Pro Lys Gln Glu Tyr Lys Leu Met Asp Met Asp Thr Leu Met
465 470 475 480
Met Ala Leu Ala Lys Glu Arg Gly Val Lys Pro Ser Glu Ile Trp Met
485 490 495
Gly Gly Met Gln Ser Tyr Glu Lys Met Gly Met Lys Met
500 505
<210> 3
<211> 24
<212> DNA
<213> 人工序列
<400> 3
atgaaaacct atacggacta tttc 24
<210> 4
<211> 20
<212> DNA
<213> 人工序列
<400> 4
ttacattttc atccccattt 20
- 还没有人留言评论。精彩留言会获得点赞!
- Corynascus thermophilus 生产耐热超氧化物歧化酶(SOD)的方法
- Thermoanaerobacter thermocopriae 生产耐热超氧化物歧化酶(SOD)的方法
- Stilbella thermophila生产耐热超氧化物歧化酶(SOD)的方法
- 普通嗜热放线菌生产耐热超氧化物歧化酶(sod)的方法
- 中间型嗜热放线菌生产耐热超氧化物歧化酶(sod)的方法
- Spirochaeta thermophila生产耐热超氧化物歧化酶(SOD)的方法
- Platyamoeba thermophila生产耐热超氧化物歧化酶(SOD)的方法
- Thermoleophilum minutum生产耐热超氧化物歧化酶(SOD)的方法
- Cryptococcus thermophilus生产耐热超氧化物歧化酶(SOD)的方法
- Thermobispora bispora生产耐热超氧化物歧化酶(SOD)的方法