色氨酸衍生物和L-蛋氨酸用于蛋白制剂的用途的制作方法
相关专利申请的交叉引用本申请要求2018年8月8日提交的美国临时申请第62/716,239号的权益,该美国临时申请各自通过引用整体并入本文。本公开涉及包含多肽、n-乙酰基-dl-色氨酸和l-蛋氨酸的液体制剂,以及其生产和使用方法。
背景技术:
:治疗性蛋白(包括单克隆抗体(mab))的生物活性取决于构象和生化稳定性。氧化是治疗性蛋白开发中许多降解问题之一,因为氧化可能会对药代动力学或生物学活性产生负面影响,特别是如果氧化发生在参与与生理靶标结合的蛋白区域中或效应子功能至关重要的区域中。另外,氧化可以改变治疗性蛋白对聚集的敏感性,从而影响免疫原性特征。管理生物治疗中的氧化风险的常用解决方案为冷冻干燥。然而,这种方法并不总是所期望的,因为其可能增加生产成本,并使药物的生产和临床使用更加复杂。经由对易于氧化的氨基酸残基的突变从而进行蛋白再造也是减轻氧化风险的可能方法。然而,靶向突变并不总是可行的解决方案,因为尽管其可以降低氧化的可能性,但其也可以降低蛋白对其靶标的结合亲和力,从而降低蛋白效价。因此,需要在制造、储存和使用过程中控制治疗性蛋白氧化的替代或补充策略。多肽制剂的实例在wo2010/030670、wo2014/160495、wo2014/160497和wo2017/117304中公开。本文所引用的所有参考文献,包括专利申请、专利公开、非专利文献和uniprotkb/swiss-prot/genbank登录号通过引用整体并入本文,如同个别参考文献各自特定地和个别地指示为通过引用并入一样。技术实现要素:为了满足上述和其他需求,本文公开了一种液体制剂,其包含多肽(例如,治疗性多肽,诸如抗体)、n-乙酰基-dl-色氨酸(nat)和l-蛋氨酸,其中该nat和该l-蛋氨酸以足以减少或防止多肽中一个或多个氨基酸残基的(例如,色氨酸残基、蛋氨酸残基等)氧化的量提供。本公开至少部分地基于以下发现,尽管添加nat可有效地在氧化应激期间保护两种示例性抗体的可变区色氨酸残基,但包括对nat使得fc蛋氨酸残基对氧化敏感。然而,发现对于两种示例性抗体,将l-蛋氨酸添加至包括nat的制剂中可以有效地保护色氨酸和蛋氨酸残基免于氧化(参见实例1)。本公开还至少部分基于以下发现,两种赋形剂在体内具有良好的耐受性(参见实例1),表明nat和l-蛋氨酸作为生物治疗制剂中的抗氧化剂赋形剂可以是安全有效的。因此,在一个方面中,本文提供了一种液体制剂,其包含多肽、n-乙酰基-dl-色氨酸(nat)和l-蛋氨酸,其中所述nat以足以防止所述多肽中一个或多个色氨酸残基的氧化的量提供,并且其中所述l-蛋氨酸以足以防止所述多肽中一个或多个蛋氨酸残基的氧化的量提供。在一些实施例中,制剂中nat的浓度为约0.01mm至约25mm。在一些实施例中,制剂中nat的浓度为约0.05mm至约1.0mm。在一些实施例中,制剂中nat的浓度为约0.05mm至约0.3mm。在一些实施例中,制剂中nat的浓度为选自由约0.05mm、约0.1mm、约0.3mm和约1.0mm组成的组的浓度。在一些实施例中,制剂中l-蛋氨酸的浓度为约1mm至约125mm。在一些实施例中,制剂中l-蛋氨酸的浓度为约5mm至约25mm。在一些实施例中,制剂中l-蛋氨酸的浓度为约5mm。在一些实施例中,制剂中nat的浓度为约0.3mm,并且制剂中l-蛋氨酸的浓度为约5.0mm。在一些实施例中,制剂中nat的浓度为约1.0mm,并且制剂中l-蛋氨酸的浓度为约5.0mm。在本发明的一些实施例中,多肽为抗体。在一些实施例中,多肽的一个或多个色氨酸残基位于抗体的可变区内。在一些实施例中,一个或多个色氨酸残基包括w103,其中残基编号根据kabat编号。在一些实施例中,一个或多个色氨酸残基位于抗体的hvr内。在一些实施例中,一个或多个色氨酸残基位于抗体的hvr-h1和/或hvr-h3内。在一些实施例中,一个或多个色氨酸残基包括w33、w36、w52a、w99、w100a和/或w100b,其中残基编号根据kabat编号。在一些实施例中,一个或多个蛋氨酸残基位于抗体的可变区内。在一些实施例中,一个或多个蛋氨酸残基包括m34和/或m82,其中残基编号根据kabat编号。在一些实施例中,一个或多个蛋氨酸残基位于抗体的恒定区内。在一些实施例中,一个或多个蛋氨酸残基包括m252和/或m428,其中残基编号根据eu编号。在一些实施例中,抗体为igg1抗体、igg2抗体、igg3抗体或igg4抗体。在一些实施例中,抗体为多克隆抗体、单克隆抗体、人源化抗体、人类抗体、嵌合抗体、多特异性抗体或抗体片段。在本发明的一些实施例中,相对于缺少nat的液体制剂中的所述多肽中一个或多个相应色氨酸残基的氧化,所述多肽中一个或多个色氨酸残基的氧化减少。在一些实施例中,相对于缺少l-蛋氨酸的液体制剂中的所述多肽中一个或多个相应蛋氨酸残基的氧化,所述多肽中一个或多个蛋氨酸残基的氧化减少。在一些实施例中,相对于缺少nat和l-蛋氨酸的液体制剂中的所述多肽中一个或多个相应色氨酸残基和一个或多个相应蛋氨酸残基的氧化,所述多肽中一个或多个色氨酸残基和一个或多个蛋氨酸残基的氧化减少。在一些实施例中,氧化减少约40%、约50%、约75%、约80%、约85%、约90%、约95%或约99%。在本发明的一些实施例中,制剂中的多肽浓度为约1mg/ml至约250mg/ml。在一些实施例中,制剂的ph值为约4.5至约7.0。在一些实施例中,制剂进一步包含一种或多种赋形剂。在一些实施例中,一种或多种赋形剂选自由稳定剂、缓冲剂、表面活性剂和张度剂组成的组。在本发明的一些实施例中,制剂为适于施用于受试者的药物制剂。在一些实施例中,药物制剂适于皮下、静脉内或玻璃体内施用。在一些实施例中,受试者为人类。在一些方面,本发明提供了一种制品或试剂盒,其包含本文所述的液体制剂。在一些方面,本发明提供了一种减少水性制剂中多肽的氧化的方法,其包括将nat和l-蛋氨酸添加至所述制剂,其中nat以足以防止所述多肽中一个或多个色氨酸残基的氧化的量提供,并且其中l-蛋氨酸以足以防止所述多肽中一个或多个蛋氨酸残基的氧化的量提供。在一些实施例中,将nat添加至该制剂,达到约0.01mm至约25mm的浓度。在一些实施例中,将nat添加至该制剂,达到约0.05mm至约1mm的浓度。在一些实施例中,将nat添加至该制剂,达到约0.05mm至约0.3mm的浓度。在一些实施例中,将nat添加至该制剂,达到选自由约0.05mm、约0.1mm、约0.3mm和约1.0mm组成的组的浓度。在一些实施例中,将l-蛋氨酸添加至该制剂,达到约1mm至约125mm的浓度。在一些实施例中,将l-蛋氨酸添加至该制剂,达到约5mm至约25mm的浓度。在一些实施例中,将l-蛋氨酸添加至该制剂,达到约5mm的浓度。在一些实施例中,将nat添加至该制剂,达到约0.3mm的浓度,并且其中将l-蛋氨酸添加至该制剂,达到约5.0mm的浓度。在一些实施例中,将nat添加至该制剂,达到约1.0mm的浓度,并且其中将l-蛋氨酸添加至该制剂,达到约5.0mm的浓度。在本发明的一些实施例中,多肽为抗体。在一些实施例中,多肽的一个或多个色氨酸残基位于抗体的可变区内。在一些实施例中,一个或多个色氨酸残基包括w103,其中残基编号根据kabat编号。在一些实施例中,一个或多个色氨酸残基位于抗体的hvr内。在一些实施例中,一个或多个色氨酸残基位于抗体的hvr-h1和/或hvr-h3内。在一些实施例中,一个或多个色氨酸残基包括w33、w36、w52a、w99、w100a和/或w100b,其中残基编号根据kabat编号。在一些实施例中,一个或多个蛋氨酸残基位于抗体的可变区内。在一些实施例中,一个或多个蛋氨酸残基包括m34和/或m82,其中残基编号根据kabat编号。在一些实施例中,一个或多个蛋氨酸残基位于抗体的恒定区内。在一些实施例中,一个或多个蛋氨酸残基包括m252和/或m428,其中残基编号根据eu编号。在一些实施例中,抗体为igg1抗体、igg2抗体、igg3抗体或igg4抗体。在一些实施例中,抗体为多克隆抗体、单克隆抗体、人源化抗体、人类抗体、嵌合抗体、多特异性抗体或抗体片段。在本发明的一些实施例中,相对于缺少nat的液体制剂中的所述多肽中一个或多个相应色氨酸残基的氧化,所述多肽中一个或多个色氨酸残基的氧化减少。在一些实施例中,相对于缺少l-蛋氨酸的液体制剂中的所述多肽中一个或多个相应蛋氨酸残基的氧化,所述多肽中一个或多个蛋氨酸残基的氧化减少。在一些实施例中,相对于缺少nat和l-蛋氨酸的液体制剂中的所述多肽中一个或多个相应色氨酸残基和一个或多个相应蛋氨酸残基的氧化,所述多肽中一个或多个色氨酸残基和一个或多个蛋氨酸残基的氧化减少。在一些实施例中,氧化减少约40%、约50%、约75%、约80%、约85%、约90%、约95%或约99%。在本发明的一些实施例中,制剂中的多肽浓度为约1mg/ml至约250mg/ml。在一些实施例中,制剂的ph值为约4.5至约7.0。在一些实施例中,制剂进一步包含一种或多种赋形剂。在一些实施例中,一种或多种赋形剂选自由稳定剂、缓冲剂、表面活性剂和张度剂组成的组。在一些实施例中,制剂为适于施用于受试者的药物制剂。在一些实施例中,药物制剂适于皮下、静脉内或玻璃体内施用。在一些实施例中,受试者为人类。应当理解,上述和本文所述的各种实施例的一个、一些或所有特性可以组合形成本公开的其他实施例。本公开的这些和其他方面对于本领域技术人员将变得显而易见。通过下面的详细描述进一步描述本公开的这些和其他实施例。附图说明图1a-1b示出了2,2′-偶氮-双(2-脒基丙烷)二盐酸盐(aaph)应激下,n-乙酰基-dl-色氨酸(nat)浓度对两种示例性igg1抗体(mab1和mab2)的氧化水平的影响。图1a示出了nat浓度对fv色氨酸氧化水平的影响。图1b示出了nat浓度对fc蛋氨酸氧化水平的影响。图2a-2b示出了在没有蛋氨酸或nat、5mm蛋氨酸、0.3mmnat或5mm蛋氨酸和0.3mmnat的组合的情况下配制的两种示例性igg1抗体(mab1和mab2)在aaph应激后的氧化水平。图2a示出了氧化敏感的fv色氨酸的氧化水平。图2b示出了fc蛋氨酸的氧化水平。图3a-3b示出了在高uv光应激后,nat浓度对两种示例性igg1抗体(mab1和mab2)的氧化水平的影响。图3a示出了nat浓度对hvr色氨酸氧化水平的影响。图3b示出了nat浓度对fc蛋氨酸氧化水平的影响。图4a-4b示出了在没有蛋氨酸或nat、5mm蛋氨酸、0.3mmnat或5mm蛋氨酸和0.3mmnat的组合的情况下配制的两种示例性igg1抗体(mab1和mab2)在高uv光应激后的氧化水平。图4a示出了hvr色氨酸的氧化水平。图4b示出了fc蛋氨酸的氧化水平。图5示出了抗氧化剂减轻化学氧化风险。图6示出了用1mmnat和5mm蛋氨酸保护w52免于氧化。具体实施方式i.定义.在详细描述本公开之前,应当理解,本公开不限于特定的组合物或生物学系统,这些组合物或生物学系统当然可以变化。另外应当了解,本文使用的术语只是为了描述特定实施例的目的,并非旨在进行限制。如本文所使用的,除非另有明确说明,否则单数形式“一个”、“一种”及“该/所述”包括复数个参考物。因此,例如,对“分子”的提及任选地包括两个或更多此类分子的组合等。如本文所用的术语“约”是指为此
技术领域:
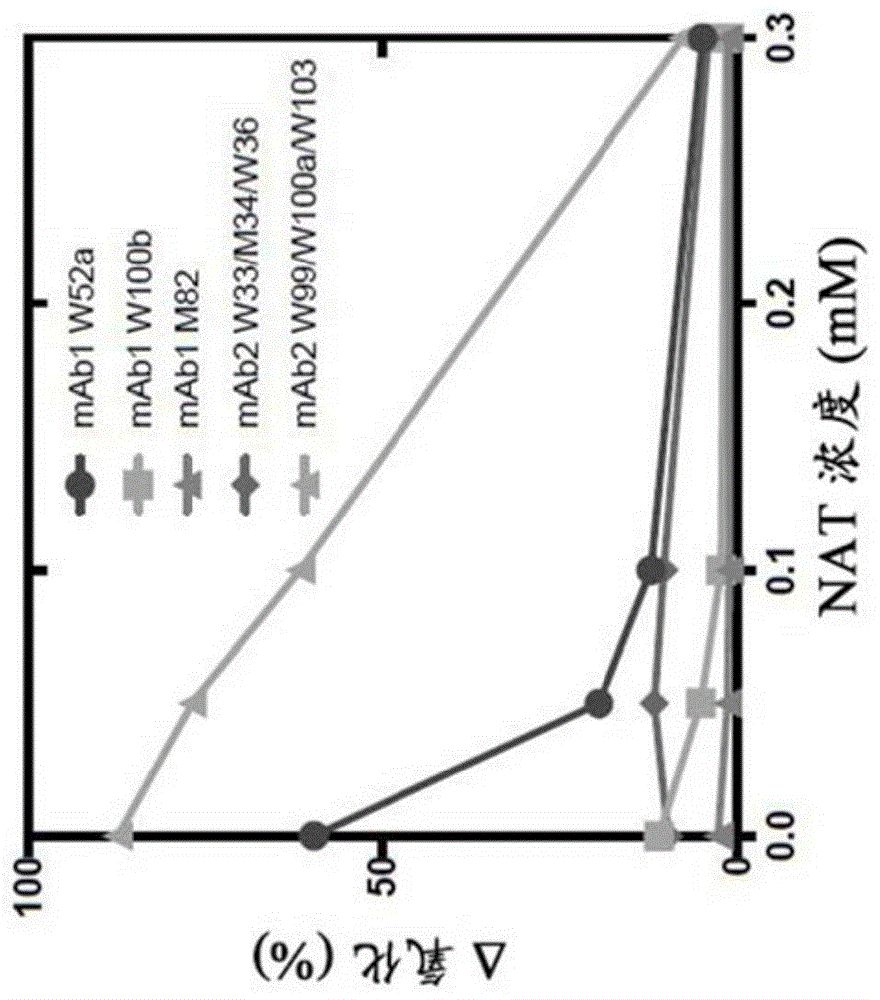
中的技术人员容易知晓的相应值的常见误差范围。在本文中提及“约”值或参数包括(且描述)涉及该值或参数举身的实施例。应当理解,本文所述的本公开的方面和实施例包括“包含”、“由以下组成”及“基本上由以下组成”所指的方面和实施例。如本文所用的术语“和/或”,短语诸如“a和/或b”旨在包括a和b;a或b;a(单独);和b(单独)。同样,如本文所用的术语“和/或”,短语诸如“a、b和/或c”旨在涵盖以下每个的实施例:a、b和c;a、b或c;a或c;a或b;b或c;a和c;a和b;b和c;a(单独);b(单独);和c(单独)。术语“药物制剂”是指处于允许活性成分的生物学活性有效的形式,并且不含对于将被施用制剂的受试者具有不可接受的毒性的另外组分的制备物。此类制剂为无菌制剂。“无菌”制剂是无菌的或者不含或基本上不含所有活微生物及其孢子。“稳定”制剂是其中多肽在储存时基本上保持其物理稳定性和/或化学稳定性和/或生物学活性的制剂。优选地,该制剂在储存时基本上保留其物理和化学稳定性以及其生物学活性。储存期通常基于制剂的预期有效期进行选择。用于测量多肽稳定性的各种分析技术是本领域中现有的,并且综述于例如以下文献中:peptideandproteindrugdelivery,247-301,vincentleeed.,marceldekker,inc.,newyork,n.y.,pubs.(1991)以及例如jones,a.adv.drugdeliveryrev.10:29-90(1993)。可在选定的曝光量和/或温度下测量选定时间段内的稳定性。可以通过各种不同的方式定性和/或定量地评估稳定性,包括聚集体形成评估(例如,使用尺寸排阻色谱法,通过测量浊度和/或通过目测);ros形成评估(例如,通过光应激测定或2,2′-偶氮-双(2-脒基丙烷)二盐酸盐(aaph)应激测定);蛋白的特定氨基酸残基(例如,单克隆抗体的trp残基和/或met残基)的氧化;通过使用阳离子交换色谱、图像毛细管等电聚焦(icief)或毛细管区带电泳评估电荷异质性;氨基末端或羧基末端序列分析;质谱分析;sds-page分析以比较还原抗体和完整抗体;肽图(例如,胰蛋白酶或lys-c)分析;评估蛋白的生物学活性或靶标结合功能(例如,抗体的抗原结合功能);等。不稳定性可能涉及以下一种或多种:聚集、脱酰胺作用(例如,asn脱酰胺作用)、氧化作用(例如,met氧化和/或trp氧化作用)、异构化作用(例如,asp异构化作用)、剪切/水解/片段化作用(例如,铰链区片段化)、琥珀酰亚胺形成、未配对的半胱氨酸、n末端延伸、c末端加工、糖基化差异等。如果通过目视检查颜色和/或透明度或者通过例如uv光散射或者尺寸排阻色谱法测量,多肽没有或几乎没有发生聚集、沉淀、片段化和/或变性的迹象,则认为该多肽在药物制剂中“保持其物理稳定性”。如果给定时间的化学稳定性使得多肽被认为仍保留其生物学活性(如下文所定义),则认为该多肽在药物制剂中“保持其化学稳定性”。化学稳定性可以通过检测和定量化学改变形式的多肽进行评估。化学改变可能涉及多肽的氧化,其可以使用,例如,胰蛋白酶肽图分析、反相高效液相色谱(hplc)和液相色谱-质谱(lc/ms)进行评估。其他类型的化学改变包括多肽的电荷改变,其可以通过,例如,离子交换色谱法或icief进行评估。如果多肽在给定时间的生物学活性为制备药物制剂时表现出的生物学活性的约20%(诸如约10%)以内(在测定误差内),例如,在单克隆抗体的抗原结合测定中测定,则认为该多肽在药物制剂中“保持其生物学活性”。如本文所用,多肽的“生物学活性”是指多肽与其靶标结合的能力,例如单克隆抗体结合抗原的能力。其可以进一步包括可以在体外或在体内测量的生物学应答。这种活性可以为拮抗的或激动的。“易于氧化”的多肽为包括一个或多个已发现易于氧化的残基的多肽,诸如但不限于蛋氨酸(met)、半胱氨酸(cys)、组氨酸(his)、色氨酸(trp)和酪氨酸(tyr)。例如,单克隆抗体的fab部分中的色氨酸氨基酸或单克隆抗体的fc部分中的蛋氨酸氨基酸可能易于氧化。多肽的“氧化不稳定”残基是在氧化测定中具有大于35%氧化的残基(例如,aaph诱发的或热诱发的氧化)。多肽中残基的氧化百分比可以通过本领域已知的任何方法测定,诸如,例如,胰蛋白酶消化,然后通过lc-ms/ms进行位点特异性trp氧化。溶剂中生物分子的“溶剂可及表面积”或“sasa”是溶剂可及的生物分子的表面积。sasa可以以度量单位(例如,平方埃)或溶剂可及的表面积的百分比表示。例如,多肽中氨基酸残基的sasa可以为或30%。sasa可以通过本领域已知的任何方法测定,包括,例如,使用shrake-rupley算法、lcpo法、功率图法或分子动力学模拟。关于目标制剂,术语“等渗”是指具有与人类血液渗透压基本相同的制剂。等渗制剂的渗透压通常为约250mosm至350mosm。等渗性可以,例如,使用蒸气压或冰冻型渗透压计来测定。如本文所用,“缓冲剂”是指通过其酸-碱共轭成分的作用而耐受ph值变化的缓冲溶液。例如,本公开的缓冲剂的ph值在约4.5至约8.0的范围内。例如,将ph值控制在该范围内的缓冲剂的实例为乙酸组氨酸。“防腐剂”是可以任选地包含在制剂中以基本上减少其中的细菌作用,从而促进例如多用途制剂的生产的化合物。潜在的防腐剂的实例包括十八烷基二甲基苄基氯化铵、六甲基氯化铵、苯扎氯铵(烷基苄基二甲基氯化铵的混合物,其中烷基为长链化合物)和苄索氯铵。其他类型的防腐剂包括芳族醇,诸如苯酚、丁醇和苄醇,烷基对羟基苯甲酸酯,诸如对羟基苯甲酸甲酯或对羟基苯甲酸丙酯、邻苯二酚、间苯二酚、环己醇、3-戊醇和间甲酚。在一个实施例中,本文的防腐剂为苯甲醇。如本文所用,“表面活性剂”是指表面活化剂,优选非离子型表面活性剂。本文所述的表面活性剂的实例包括聚山梨醇酯(例如,聚山梨醇酯20和聚山梨醇酯80);泊洛沙姆(例如,泊洛沙姆188);triton;十二烷基硫酸钠(sds);月桂基硫酸钠;辛基糖苷钠;月桂基、肉豆蔻基、亚油基或硬脂酰磺基甜菜碱;月桂基-、肉豆蔻基、亚油基或硬脂酰肌氨酸;亚油基、肉豆蔻基或十六烷基甜菜碱;月桂酰氨基丙基、椰油酰胺丙基、亚油酰胺丙基、肉豆蔻基丙基、棕榈酰氨基丙基或异硬脂酰胺丙基甜菜碱(例如,月桂酰胺丙基);肉豆蔻基胺丙基、棕榈酰丙基或异硬脂酰胺丙基-二甲基胺;甲基椰油酰基钠或甲基油基-牛磺酸二钠;和monaquattm系列(monaindustries,inc.,paterson,n.j.);聚乙二醇、聚丙二醇以及乙烯与丙二醇的共聚物(例如,pluronics、pf68等)等。在一个实施例中,本文所述的表面活性剂为聚山梨醇酯20。在又一实施例中,本文的表面活性剂为泊洛沙姆188。本文所用的“药用”赋形剂或运载体包括本领域熟知的药用运载体、稳定剂、缓冲剂、酸、碱、糖、防腐剂、表面活性剂、张度剂等(remington:thescienceandpracticeofpharmacy,22nded.,pharmaceuticalpress,2012)。药用赋形剂的实例包括:缓冲剂,诸如磷酸盐、柠檬酸盐、乙酸盐及其他有机酸;抗氧化剂,包括抗坏血酸、l-色氨酸和蛋氨酸;低分子量(少于约10个残基)多肽;蛋白质,诸如血清白蛋白、明胶或免疫球蛋白;亲水性聚合物,诸如聚乙烯吡咯烷酮;氨基酸,诸如甘氨酸、谷氨酰胺、天冬酰胺、精氨酸或赖氨酸;单糖、二糖及其他碳水化合物,包括葡萄糖、甘露糖或糊精;金属配合物,诸如锌蛋白配合物;螯合剂,诸如edta;糖醇,诸如甘露醇或山梨糖醇;成盐抗衡离子,诸如钠;和/或非离子型表面活性剂,诸如聚山梨酯、泊洛沙姆、聚乙二醇(peg)和pluronicstm。“药用”赋形剂或运载体是可以合理地施用于受试者以提供有效剂量的所用活性成分并且以所采用的剂量和浓度对暴露于其中的受试者无毒的那些赋形剂或运载体。配制的多肽优选地为基本上纯的并且预期是基本上均质的(例如,无污染的蛋白等)。“基本上纯的”多肽意指基于组合物的总重量,其包括至少约90重量%、优选地至少约95重量%的多肽(例如,单克隆抗体)的组合物。“基本上均质的”多肽意指基于组合物的总重量,其包括至少约99重量%的多肽(例如,单克隆抗体)的组合物。术语“蛋白”、“多肽”和“肽”在本文中可互换使用以指代任何长度的氨基酸聚合物。聚合物可为线性或分支的,其可包含经过修饰的氨基酸,并且其可间插有非氨基酸。这些术语还涵盖已天然修饰或通过介入修饰的氨基酸聚合物;例如,二硫键形成、糖基化、脂质化、乙酰化、磷酸化或任何其他操作或修饰,诸如与标记组分缀合。在定义内还包括,例如,含有氨基酸(包括,例如,非天然氨基酸等)的一种或多种类似物以及本领域中已知的其他修饰的多肽。本文定义中涵盖的多肽的实例包括哺乳动物多肽,诸如,例如,肾素;生长激素,包括人类生长激素和牛生长激素;生长激素释放因子;甲状旁腺激素;促甲状腺激素;脂蛋白;α-1-抗胰蛋白酶;胰岛素a链;胰岛素b链;胰岛素原;卵泡刺激素;降钙素;促黄体素;胰高血糖素;瘦素;凝血因子诸如viiic因子、ix因子、组织因子和维勒布兰德氏(vonwillebrands)因子;抗凝血因子诸如蛋白c;心钠素;肺表面活性剂;纤溶酶原激活剂,诸如尿激酶或人类尿或组织型纤溶酶原激活剂(t-pa);蛙皮素(bombesin);凝血酶;造血生长因子;肿瘤坏死因子-α和-β;肿瘤坏死因子受体诸如死亡受体5和cd120;tnf相关的凋亡诱导配体(trail);b细胞成熟抗原(bcma);b淋巴细胞刺激因子(blys);增殖诱导配体(april);脑啡肽酶;rantes(调节正常表达和分泌的t细胞活化);人类巨噬细胞炎性蛋白(mip-1-α);血清白蛋白诸如人类血清白蛋白;缪勒管(muellerian)抑制物质;松弛素a链;松弛素b链;松弛素原;小鼠促性腺激素相关肽;微生物蛋白,诸如β-内酰胺酶、脱氧核糖核酸酶;ige;细胞毒性t淋巴细胞相关抗原(ctla),诸如ctla-4;抑制素;激活素;血小板衍生内皮细胞生长因子(pd-ecgf);血管内皮生长因子家族蛋白(例如,vegf-a、vegf-b、vegf-c、vegf-d和p1gf);血小板衍生生长因子(pdgf)家族蛋白(例如,pdgf-a、pdgf-b、pdgf-c、pdgf-d及其二聚体);成纤维细胞生长因子(fgf)家族诸如afgf、bfgf、fgf4和fgf9;表皮生长因子(egf);激素或生长因子的受体诸如vegf受体(例如,vegfr1、vegfr2和vegfr3),表皮生长因子(egf)受体(例如,erbb1、erbb2、erbb3和erbb4受体),血小板衍生生长因子(pdgf)受体(例如,pdgfr-α和pdgfr-β),和成纤维细胞生长因子受体;tie配体(血管生成素、angpt1、angpt2);血管生成素受体诸如tie1和tie2;蛋白a或d;类风湿因子;神经营养因子诸如骨源性神经营养因子(bdnf),神经营养蛋白-3、-4、-5或-6(nt-3、nt-4、nt-5或nt-6)或神经生长因子诸如ngf-b;转化生长因子(tgf)诸如tgf-α和tgf-β,包括tgf-β1、tgf-β2、tgf-β3、tgf-β4或tgf-β5;胰岛素样生长因子-i和-ii(igf-i和igf-ii);des(1-3)-igf-i(脑igf-1),胰岛素样生长因子结合蛋白(igfbp);cd蛋白诸如cd3、cd4、cd8、cd19和cd20;促红细胞生成素;骨诱导因子;免疫毒素;骨形态发生蛋白(bmp);趋化因子诸如cxcl12和cxcr4;干扰素诸如干扰素-α、-β和-γ;集落刺激因子(csf),例如,m-csf、gm-csf和g-csf;细胞因子诸如白介素(il),例如,il-1至il-10;中期因子;超氧化物歧化酶;t细胞受体;表面膜蛋白;衰变加速因子;病毒抗原诸如,例如,aids包膜的一部分;转运蛋白;归巢受体;寻址蛋白;调节蛋白;整合素诸如cd11a、cd11b、cd11c、cd18、icam、vla-4和vcam;肝配蛋白;bv8;δ样配体4(dll4);del-1;bmp9;bmp10;卵泡抑素;肝细胞生长因子(hgf)/散射因子(sf);alk1;robo4;esm1;串珠素;egf样结构域7(egfl7);ctgf及其家族成员;血小板反应蛋白诸如血小板反应蛋白1和血小板反应蛋白2;胶原蛋白诸如胶原蛋白iv和胶原蛋白xviii;神经纤毛蛋白诸如nrp1和nrp2;多效生长因子(ptn);颗粒蛋白原;多育曲菌素;notch蛋白诸如notch1和notch4;导蛋白诸如sema3a、sema3c和sema3f;肿瘤相关抗原诸如ca125(卵巢癌抗原);免疫粘附素;以上列出的任何多肽以及抗体(包括抗体片段)的片段和/或变体与一个或多个蛋白(包括,例如,以上列出的任何蛋白)结合。本文的术语“抗体”以最广泛的含义使用,并且具体地覆盖单克隆抗体(包括全长单克隆抗体)、多克隆抗体、多特异性抗体(例如,双特异性、三特异性等)和抗体片段,只要其表现出所需生物学活性。“分离的”多肽(例如,分离的抗体)是已经从其天然环境的组分中鉴别和分离和/或回收的多肽。其自然环境的污染物组分是会干扰对于多肽的研究、诊断或治疗用途的材料,并且可以包括酶、激素和其他蛋白或非蛋白溶质。经分离的多肽包含重组细胞内的原位多肽,因为不存在多肽天然环境的至少一种组分。然而,通常,分离的多肽将通过至少一个纯化步骤进行制备。“天然抗体”通常是约150,000道尔顿的异源四聚体糖蛋白,由两条相同的轻(l)链和两条相同的重(h)链组成。每条轻链通过一个共价二硫键与重链相连,而二硫键的数目在不同免疫球蛋白同种型的重链之间变化。每条重链和轻链还具有规则间隔的链内二硫键。每条重链在一端具有可变结构域(vh),其后是多个恒定结构域。每条轻链的一端具有可变结构域(vl),另一端具有恒定结构域;轻链的恒定结构域与重链的第一恒定结构域对齐,并且轻链的可变结构域与重链的可变结构域对齐。据信特定的氨基酸残基在轻链和重链可变结构域之间形成界面。术语“恒定结构域”是指免疫球蛋白分子的一部分,该部分相对于免疫球蛋白的另一部分(即可变结构域,其包含抗原结合位点)具有更保守的氨基酸序列。恒定结构域包含重链的ch1、ch2和ch3结构域(统称为ch)和轻链的chl(或cl)结构域。抗体的“可变区”或“可变结构域”是指抗体的重链或轻链的氨基末端结构域。重链的可变结构域可以称为“vh”。轻链的可变结构域可以称为“vl。”这些结构域通常是抗体中变化最大的部分,并且包含抗原结合位点。术语“可变的”是指以下事实:可变结构域的某些部分在抗体之间的序列差异很大,并用于每种特定抗体对其特定抗原的结合和特异性。然而,可变性并非在抗体的可变结构域中均匀分布。它集中在轻链和重链可变结构域中的三个称为高变区(hvr)的区段中。可变结构域中保守性更高的部分称为框架区(fr)。天然重链和轻链的可变结构域各自包含四个fr区,其主要采用β折叠结构,由三个hvr连接,这三个hvr形成连接β折叠结构的环并且在一些情况下形成β折叠结构的一部分。每条链中的hvr通过fr区紧密保持在一起,并且与另一条链中的hvr一起,有助于抗体的抗原结合位点的形成(参见kabat等人,sequencesofproteinsofimmunologicalinterest,第五版,美国卫生与公众服务部,国立卫生研究院,马里兰州贝塞斯达(1991))。恒定结构域不直接参与抗体与抗原的结合,但具有各自效应子功能,诸如抗体参与抗体依赖性细胞毒性作用。来自任何哺乳动物物种抗体(免疫球蛋白)的“轻链”基于其恒定结构域的氨基酸序列,可以配属为两种明显不同的类型中的一种,这两种类型分别称为卡帕(“κ”)和兰姆达(“λ”)。如本文所用,术语igg“同种型”或“亚类”是指由免疫球蛋白恒定区的化学和抗原特征定义的免疫球蛋白的任何亚类。根据其重链恒定结构域的氨基酸序列,可以将抗体(免疫球蛋白)分为不同的类别。免疫球蛋白主要分为五类:iga、igd、ige、igg和igm,并且它们中的一些可以进一步分为亚类(同种型),例如,igg1、igg2、igg3、igg4、iga1,以及iga2。对应于不同类别的免疫球蛋白的重链恒定结构域分别称为α、γ、ε、γ和μ。不同类别的免疫球蛋白的亚基结构和三维构型是众所周知的,并在例如总体描述于abbas等人cellularandmol.immunology,4thed.,w.b.saunders,co.,2000。抗体可以是较大融合分子的一部分,该融合分子是通过抗体与一个或多个其他蛋白质或肽的共价或非共价结合形成的。术语“全长抗体”、“完整抗体”和“全抗体”在本文中可互换使用,是指其基本上完整形式的抗体而不是如下文定义的抗体片段。该术语特别是指具有包含fc区的重链的抗体。“抗体片段”包含完整抗体的一部分,优选包含其抗原结合区。抗体片段的实例包括fab、fab′、f(ab′)2和fv片段;双体抗体;线性抗体;单链抗体分子;和由抗体片段形成的多特异性抗体。木瓜蛋白酶消化抗体产生两个相同抗原结合片段,称为“fab”片段,每个片段都有单个抗原结合位点和残留的“fc”片段,其名称反映其容易结晶的能力。胃蛋白酶处理产生的f(ab′)2片段具有两个抗原结合位点并且仍能与抗原交联。fab片段含有重链可变结构域和轻链可变结构域且亦含有轻链的恒定结构域和重链的第一恒定结构域(ch1)。fab′片段与fab片段的不同之处在于fab′片段在重链ch1结构域的羧基末端添加了一些残基,这些残基包括来自抗体铰链区的一个或多个半胱氨酸。fab′-sh是本文中关于其中恒定结构域的半胱氨酸残基带有游离硫醇基的fab′的命名。f(ab′)2抗体片段最初是作为在其间具有铰链半胱氨酸的成对fab′片段而产生的。抗体片段的其他化学偶联也是已知的。“fv”是包含完全的抗原结合位点的最小抗体片段。在一个实施例中,双链fv种类由紧密和非共价结合的一个重链和一个轻链可变结构域的二聚体组成。在单链fv(scfv)物种中,一个重链可变结构域和一个轻链可变结构域可通过柔性肽接头共价连接,使得轻链和重链可缔合成类似于在双链fv物种中的“二聚体”结构。以此构型,每个可变结构域的三个hvr相互作用以在vh-vl二聚体的表面上限定抗原结合位点。六个hvr共同对抗体赋予抗原结合特异性。但是,即使单个可变结构域(或仅包含三个对抗原具有特异性的hvr的fv的一半)也具有识别和结合抗原的能力,尽管其亲和力低于完整结合位点。“单链fv”或“scfv”抗体片段包含抗体的vh和vl结构域,其中这些结构域存在于单个多肽链中。一般地,scfv多肽在vh和vl结构域之间进一步包含多肽接头,使scfv形成所需的抗原结合结构。有关scfv的综述,参见,例如,pluckthun的thepharmacologyofmonoclonalantibodies,第113卷,rosenburg和moore主编,springer-verlag,newyork,pp.269-315,1994。术语“双体抗体”是指具有两个抗原结合位点的抗体片段,其片段包含连接至与同一多肽链(vh-vl)中的轻链可变结构域(vl)的重链可变结构域(vh)。通过使用太短以至于不允许同一条链上两个结构域之间配对的接头,这些结构域被迫与另一条链的互补结构域配对并产生两个抗原结合位点。双体抗体可为二价抗体或双特异性抗体。双体抗体更全面地描述于例如:ep404,097;wo1993/01161;hudson等人,nat.med.9:129-134(2003);以及hollinger等人,proc.natl.acad.sci.usa90:6444-6448(1993)。三体抗体和四体抗体也在hudson等人,nat.med.9:129-134(2003)中进行了描述。本文所使用的术语“单克隆抗体”是指从基本上同质的抗体群中获得的抗体,例如,除了可能存在的少量突变例如天然存在的突变,该抗体群包含的单个抗体是相同的。因此,修饰语“单克隆的”表明抗体的特征不是离散抗体的混合物。在一些实施例中,这样的单克隆抗体通常包括含有结合靶标的多肽序列的抗体,其中靶标结合多肽序列通过包括从多个多肽序列中选择单个靶标结合多肽序列的过程获得。例如,选择过程可以是从多个克隆,例如杂交瘤克隆、噬菌体克隆或重组dna克隆的集合中选择独特的克隆。应当理解,可以进一步改变选择的靶标结合序列,例如,以提高对靶标的亲和力、使靶标结合序列人源化、提高其在细胞培养物中的产生、降低其在体内的免疫原性、产生多特异性抗体等,并且包含改变的靶标结合序列的抗体也是本公开的单克隆抗体。与通常包括针对不同决定簇(表位)的不同抗体的多克隆抗体制剂相反,单克隆抗体制剂中的每种单克隆抗体针对抗原上的单一决定簇。除其特异性外,单克隆抗体制剂的优势还在于其通常不受其他免疫球蛋白的污染。修饰语“单克隆”表示抗体的特征是从基本上同质的抗体群体获得的,并且不应解释为需要通过任何特定方法产生抗体。例如,根据本公开使用的单克隆抗体可以通过多种技术制备,包括,例如,杂交瘤方法(例如,kohler和milstein,nature,256:495-97(1975);hongo等人,hybridoma,14(3):253-260(1995),harlow等人,antibodies:alaboratorymanual,(coldspringharborlaboratorypress,第2版1988);hammerling等人,monoclonalantibodiesandt-cellhybridomas563-681(elsevier,n.y.,1981))、重组dna方法(参见,例如,美国专利号4,816,567)、噬菌体展示技术(参见,例如,clackson等人,nature,352:624-628(1991);marks等人,j.mol.biol.222:581-597(1992);sidhu等人,j.mol.biol.338(2):299-310(2004);lee等人,j.mol.biol.340(5):1073-1093(2004);fellouse,proc.natl.acad.sci.usa101(34):12467-12472(2004);和lee等人,j.immunol.methods284(1-2):119-132(2004))和在动物中产生具有编码人类免疫球蛋白序列的人类免疫球蛋白基因座或基因的部分或全部的人类抗体或类人类抗体的技术(参见,例如,wo1998/24893;wo1996/34096;wo1996/33735;wo1991/10741;jakobovits等人,proc.natl.acad.sci.usa90:2551(1993);jakobovits等人,nature362:255-258(1993);bruggemann等人,yearinimmunol.7:33(1993);美国专利号5,545,807、5,545,806、5,569,825、5,625,126、5,633,425和5,661,016;marks等人,bio/technology10:779-783(1992);lonberg等人,nature368:856-859(1994);morrison,nature368:812-813(1994);fishwild等人,naturebiotechnol.14:845-851(1996);neuberger,naturebiotechnol.14:826(1996)以及lonberg和huszar,intern.rev.immunol.13:65-93(1995))。本文中的单克隆抗体具体地包括“嵌合”抗体,其中重链和/或轻链的一部分与来自特定物种或属于特定抗体类别或亚类的抗体中的相应序列相同或同源,而一条或多条链的其余部分与来自另一物种或属于另一抗体类别或亚类的抗体中的相应序列以及这些抗体的片段相同或同源,只要它们表现出所需的生物学活性即可(参见,例如,美国专利号4,816,567;和morrison等人,proc.natl.acad.sci.usa81:6851-6855(1984))。嵌合抗体包括抗体,其中抗体的抗原结合区来源于通过例如用目标抗原免疫猕猴而产生的抗体。“人源化”形式的非人类(例如,鼠)抗体为包含来源于非人类免疫球蛋白的最小序列的嵌合抗体。在一个实施例中,人源化抗体是人类免疫球蛋白(受体抗体),其中来自受体hvr的残基被来自非人类物种(供体抗体)例如小鼠、大鼠、兔或具有所需特异性、亲和力和/或能力的非人类灵长类动物的hvr的残基取代。在一些情况下,人类免疫球蛋白的fr残基被相应的非人类残基取代。此外,人源化抗体可包含受体抗体或供体抗体中不存在的残基。可以进行这些修饰以进一步改善抗体性能。总体上,人源化抗体将基本上包含所有中的至少一个可变结构域,通常是两个可变结构域,其中所有或基本上所有高变环对应于非人类免疫球蛋白的高变环,并且所有或基本上所有的fr为人类免疫球蛋白序列的fr。人源化抗体还将任选地包含免疫球蛋白恒定区(fc)的至少一部分,该免疫球蛋白通常为人类免疫球蛋白。更多详情,参见,例如,jones等人,nature321:522-525(1986);riechmann等人,nature332:323-329(1988);以及presta,curr.op.struct.biol.2:593-596(1992)。也参见,例如,vaswani和hamilton,ann.allergy,asthma&immunol.1:105-115(1998);harris,biochem.soc.transactions23:1035-1038(1995);hurle和gross,curr.op.biotech.5:428-433(1994);以及美国专利号6,982,321和7,087,409。“人类抗体”是具有对应于由人类产生的抗体的氨基酸序列的抗体和/或使用本文所公开的用于制备人类抗体的任何技术制得的抗体。人类抗体的该定义特别地排除了包含非人类抗原结合残基的人源化抗体。可以使用本领域已知的各种技术产生人类抗体,包括噬菌体展示文库。hoogenboom和winter,j.mol.biol.,227:381(1991);marks等人,j.mol.biol.,222:581(1991)。还可用于制备人类单克隆抗体的方法如cole等人,monoclonalantibodiesandcancertherapy,alanr.liss,p.77(1985);boerner等人,j.immunol.,147(1):86-95(1991)所述。另参见vandijk和vandewinkel,curr.opin.pharmacol.,5:368-74(2001)。可以通过向转基因动物施用抗原来制备人类抗体,该转基因动物已经修饰以对抗原攻击产生应答而产生此类抗体,但其内源基因座已失效,例如,免疫异种小鼠(参见,例如,关于xenomousetm技术的美国专利号6,075,181和6,150,584)。另参见,例如,li等人,proc.natl.acad.sci.usa,103:3557-3562(2006)关于通过人类b细胞杂交瘤技术产生的人类抗体。如本文所用的术语“高变区”、“hvr”或“hv”是指在序列上高变和/或形成结构上限定的环的抗体可变结构域的区域。通常,抗体包含六个hvr;三个在vh中(h1、h2、h3),并且三个在vl中(l1、l2、l3)。在天然抗体中,h3和l3在六个hvr中表现出最多的多样性,尤其是h3被认为在赋予抗体精细特异性方面起着独特的作用。参见,例如,xu等人,immunity13:37-45(2000);johnson和wu,methodsinmolecularbiology248:1-25(lo,ed.,humanpress,totowa,n.j.,2003)。实际上,仅由重链组成的天然存在的骆驼科动物抗体在不存在轻链的情况下是有功能并稳定的。参见,例如,hamers-casterman等人,nature363:446-448(1993);sheriff等人,naturestruct.biol.3:733-736(1996)。在一些实施例中,hvr为互补决定区(cdr)。许多hvr描述得到应用,并且包含于本文中。kabat互补决定区(cdr)基于序列变异性并且是最常用的(kabat等人,sequencesofproteinsofimmunologicalinterest,第5版,美国卫生与公众服务部,国立卫生研究院,马里兰州贝塞斯达(1991))。相反,chothia指的是结构环的位置(chothia和leskj.mol.biol.196:901-917(1987))。abmhvr表示kabathvr和chothia结构环之间的折衷,并且被牛津分子公司(oxfordmolecular)的abm抗体建模软件采用。“接触”hvr基于可用的复杂晶体结构的分析结果。这些hvr中的每个的残基如下文所述。hvr可以包括以下“扩展hvr”:vl中的24-36或24-34(l1)、46-56或50-56(l2)和89-97或89-96(l3),以及vh中的26-35(h1)、50-65或49-65(h2)和93-102、94-102或95-102(h3)。对于这些定义中的每一个,可变结构域残基均根据上述kabat等人的方法进行编号。“框架”或“fr”残基是除本文定义的hvr残基以外的那些可变结构域残基。术语“kabat所述的可变结构域残基编号”、“kabat所述的氨基酸位置编号”、“根据kabat编号的残基编号”及其变型是指在上述kabat等人的文献中(同上)提出的用于抗体的重链可变结构域或轻链可变结构域的编号系统。使用该编号系统,实际线性氨基酸序列可能包含较少或附加的氨基酸,其对应于对可变结构域的fr或hvr的缩短或插入。例如,重链可变结构域可在h2的残基52之后包括单个氨基酸插入(根据kabat编号的残基52a)以及重链fr残基82之后的插入残基(例如,根据kabat编号的残基82a、82b和82c等)。可通过将抗体序列与“标准”kabat编号序列的同源性区域进行比对来测定给定抗体的残基的kabat编号当提及可变结构域中的残基(大约是轻链的残基1-107和重链的残基1-113)时,通常使用kabat编号系统(例如,kabat等人,sequencesofimmunologicalinterest.第5版,美国卫生与公众服务部,国立卫生研究院,bethesda,md.(1991))。当提及免疫球蛋白重链恒定区中的残基时,通常使用术语“eu编号系统”、“eu索引”、“残基编号是根据eu编号”及其变体(例如,kabat等人报告的eu索引,同上)。“kabat所述的eu索引”是指人类igg1eu抗体的残基编号。术语“多特异性抗体”以最广义使用,并且特别涵盖包含具有多表位特异性的抗原结合结构域的抗体(即,能够特异性结合一个生物分子上的两个或多个不同表位,或能够特异性结合两个或多个不同生物分子上的表位)。在一些实施例中,多特异性抗体(诸如双特异性抗体)的抗原结合结构域包含两个vh/vl单元,其中第一vh/vl单元与第一表位特异性结合,第二vh/vl单元与第二表位特异性结合,其中每个vh/vl单元包含重链可变结构域(vh)和轻链可变结构域(vl)。这样的多特异性抗体包括但不限于全长抗体,具有两个或多个vl和vh结构域的抗体,抗体片段诸如fab、fv、dsfv、scfv、双抗体、双特异性双抗体和三抗体,已共价或非共价连接的抗体片段。进一步包含重链恒定区的至少一部分和/或轻链恒定区的至少一部分的vh/vl单元也可以称为“半体(hemimer)”或“半抗体(halfantibodv)”。在一些实施例中,半抗体包含单个重链可变区的至少一部分和单个轻链可变区的至少一部分。在一些这样的实施例中,包含两个半抗体并与两个抗原结合的双特异性抗体包括与第一抗原或第一表位结合但不与第二抗原或第二表位结合的第一半抗体,以及与第二抗原或第二表位结合而不与第一抗原或第一表位结合的第二半抗体。根据一些实施例,多特异性抗体是以5m至0.001pm、3m至0.001pm、1m至0.001pm、0.5m至0.001pm或0.1m至0.001pm的亲和力与各自抗原或表位结合的igg抗体。在一些实施例中,半体包括重链可变区的足够部分,以实现与第二半体形成分子内二硫键。在一些实施例中,半体包括杵突变(knobmutation)或臼突变(holemutation),例如,以实现与包含互补臼突变或杵突变的第二半体或半抗体的异二聚化。杵突变和臼突变将在以下进一步讨论。“双特异性抗体”是包含抗原结合结构域的多特异性抗体,该抗原结合结构域能够特异性结合一个生物分子上的两个不同表位或能够特异性结合两种不同生物分子上的表位。双特异性抗体在本文中也可以被称为具有“双重特异性”或为“双重特异性的”。除非另有说明,双特异性抗体结合的抗原在双特异性抗体名称中列出的顺序是任意的。在一些实施例中,双特异性抗体包含两个半抗体,其中半抗体各自包含单个重链可变区和任选地重链恒定区的至少一部分,以及单个轻链可变区和任选地轻链恒定区的至少一部分。在一些实施例中,双特异性抗体包含两个半抗体,其中半抗体各自包含单个重链可变区和单个轻链可变区,并且不包含多于一个的单个重链可变区并且不包含多于一个的单个轻链可变区。在一些实施例中,双特异性抗体包含两个半抗体,其中半抗体各自包含单个重链可变区和单个轻链可变区,并且其中第一半抗体结合第一抗原而不结合第二抗原,并且第二半抗体结合第二抗原而不结合第一抗原。如本文所用,术语“杵臼(knob-into-hole)”或“knh”技术是指通过在其相互作用的界面将突起(杵)引入一个多肽并将腔(臼)引入其他多肽,从而引导在体内或体外将两个多肽配对在一起的技术。例如,已在抗体的fc:fc结合界面、cl:ch1界面或vh/vl界面中引入knh(参见,例如,us201i/0287009、us2007/0178552、wo96/027011、wo98/050431和zhu等人,1997,proteinscience6:781-788)。在一些实施例中,在多特异性抗体的制造过程中,knh驱动两个不同的重链配对在一起。例如,在其fc区中具有knh的多特异性抗体可以进一步包含与各自fc区连接的单个可变结构域,或进一步包含与相似或不同的轻链可变结构域配对的不同的重链可变结构域。knh技术还可以用于将两个不同的受体胞外域或包含不同靶标识别序列的任何其他多肽序列配对在一起(例如,包括亲和体、肽体和其他fc融合体)。如本文所用,术语“杵突变”是指在多肽与另一多肽相互作用的界面将突起(杵)引入多肽的突变。在一些实施例中,其他多肽具有臼突变(参见,例如,us5,731,168、us5,807,706、us5,821,333、us7,695,936、us8,216,805,其各自通过引用整体并入本文)。如本文所用,术语“臼突变”是指在多肽与另一多肽相互作用的界面将腔(臼)引入多肽的突变。在一些实施例中,其他多肽具有杵突变(参见,例如,us5,731,168、us5,807,706、us5,821,333、us7,695,936、us8,216,805,其各自通过引用整体并入本文)。表述“线性抗体”是指zapata等人在(1995proteineng,8(10):1057-1062)中所述的抗体。简而言之,这些抗体包含一对串联的fd区段(vh-ch1-vh-ch1),其与互补的轻链多肽一起形成一对抗原结合区。线性抗体可以为双特异性或单特异性的。ii.多肽制剂与制备本公开的某些方面涉及一种制剂,其包含多肽、n-乙酰基-dl-色氨酸(nat)和l-蛋氨酸,其中nat和l-蛋氨酸减少或防止多肽的氧化。在一些实施例中,多肽易于氧化。在一些实施例中,多肽中的蛋氨酸、半胱氨酸、组氨酸、色氨酸和/或酪氨酸残基易于氧化。在一些实施例中,多肽中一个或多个色氨酸残基易于氧化。在一些实施例中,多肽中一个或多个蛋氨酸残基易于氧化。在一些实施例中,多肽中一个或多个色氨酸和一个或多个蛋氨酸残基易于氧化。在一些实施例中,多肽为抗体。在一些实施例中,制剂进一步包括根据本文所述的任何多肽的至少一种另外的多肽。在一些实施例中,制剂进一步包含一种或多种赋形剂。在一些实施例中,制剂为液体制剂。在一些实施例中,制剂为水性制剂。在一些实施例中,制剂为药物制剂(例如,适于施用于人类受试者)。在一些实施例中,制剂中nat的浓度为约0.01mm至约25mm(诸如约0.01、0.025、0.05、0.075、0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1.0、2.0、3.0、4.0、5.0、6.0、7.0、8.0、9.0、10.0、11.0、12.0、13.0、14.0、15.0、16.0、17.0、18.0、19.0、20.0、21.0、22.0、23.0、24.0或25.0mm中的任一浓度,包括这些值之间的任何范围),或直至nat可溶于制剂的最高浓度。在一些实施例中,制剂中nat的浓度为约0.05mm至约1mm。在一些实施例中,制剂中nat的浓度为约0.05mm至约0.3mm。在一些实施例中,制剂中nat的浓度为约0.05mm。在一些实施例中,制剂中nat的浓度为约0.1mm。在一些实施例中,制剂中nat的浓度为约0.3mm。在一些实施例中,制剂中nat的浓度为约1.0mm。在一些实施例中,制剂中nat的浓度为约1mm。在一些实施例中,nat减少或防止多肽中一个或多个色氨酸残基的氧化。在一些实施例中,nat减少或防止多肽中一个或多个色氨酸残基被活性氧(ros)氧化。在一些实施例中,活性氧选自单线态氧、超氧化物(o2-)、烷氧自由基、过氧自由基、过氧化氢(h2o2)、三氧化二氢(h2o3)、氢三氧自由基(ho3)、臭氧(o3)、羟基自由基和/或烷基过氧化物。在一些实施例中,多肽为抗体,并且nat减少或防止抗体中一个或多个色氨酸残基的氧化。在一些实施例中,一个或多个色氨酸残基位于抗体的轻链恒定区和/或重链恒定区内。在一些实施例中,一个或多个色氨酸残基位于抗体的轻链可变区(例如,hvr-l1、hvr-l2和/或hvr-l3)和/或重链可变区(例如,hvr-h1、hvr-h2和/或hvr-h3)内。在一些实施例中,一个或多个色氨酸残基位于抗体的重链可变区内。在一些实施例中,一个或多个色氨酸残基位于重链可变区的框架区内。在一些实施例中,一个或多个色氨酸残基包括w103(根据kabat编号)。在一些实施例中,一个或多个色氨酸残基位于抗体(例如,hvr-h1和/或hvr-h3)的hvr-h1、hvr-h2和/或hvr-h3内。在一些实施例中,一个或多个色氨酸残基包括w33、w36、w52、w52a、w99、w100a、w100b和/或w103(根据kabat编号)。在一些实施例中,一个或多个色氨酸残基包括w33和/或w36、w99和/或w100a。在一些实施例中,在本公开的制剂中包括nat减少或防止抗体在残基w33、w36、w52a、ww99、w100a、wl10b和/或wl03处的氧化(例如,与缺少nat的液体制剂中多肽中一个或多个相应色氨酸残基相比)。在一些实施例中,一个或多个色氨酸残基位于抗体的hvr-l1、hvr-l2和/或hvr-l3内。在一些实施例中,一个或多个色氨酸残基包括w94、w31和/或w91。在一些实施例中,制剂中l-蛋氨酸的浓度为约1.0mm至约125.0mm(诸如约1.0、2.0、3.0、4.0、5.0、6.0、7.0、8.0、9.0、10.0、15.0、20.0、25.0、30.0、35.0、40.0、45.0、50.0、55.0、60.0、65.0、70.0、75.0、80.0、85.0、90.0、95.0、100.0、105.0、110.0、115.0、120.0或125.0mm中的任一浓度,包括这些值之间的任何范围),或直至l-蛋氨酸可溶于制剂的最高浓度。在一些实施例中,制剂中l-蛋氨酸的浓度为约5.0mm至约25.0mm。在一些实施例中,制剂中l-蛋氨酸的浓度为约5.0mm。在一些实施例中,l-蛋氨酸减少或防止多肽中一个或多个蛋氨酸残基的氧化。在一些实施例中,l-蛋氨酸减少或防止多肽中一个或多个蛋氨酸残基被活性氧(ros)氧化。在一些实施例中,活性氧选自单线态氧、超氧化物(o2-)、烷氧自由基、过氧自由基、过氧化氢(h2o2)、三氧化二氢(h2o3)、氢三氧自由基(ho3)、臭氧(o3)、羟基自由基和/或烷基过氧化物。在一些实施例中,多肽为抗体,并且l-蛋氨酸减少或防止抗体中一个或多个蛋氨酸残基的氧化。在一些实施例中,一个或多个蛋氨酸残基位于抗体的轻链可变区(例如,hvr-l1、hvr-l2和/或hvr-l3)和/或重链可变区(例如,hvr-h1、hvr-h2和/或和hvr-h3)内。在一些实施例中,一个或多个蛋氨酸残基位于抗体的重链可变区内。在一些实施例中,一个或多个蛋氨酸残基位于重链可变区的框架区内。在一些实施例中,一个或多个蛋氨酸残基包括m82(根据kabat编号)。在一些实施例中,一个或多个色氨酸残基位于抗体的hvr-h1、hvr-h2和/或hvr-h3(例如,hvr-h1)内。在一些实施例中,一个或多个蛋氨酸残基包括m34(根据kabat编号)。在一些实施例中,一个或多个蛋氨酸残基位于抗体的hvr-l1、hvr-l2和/或hvr-l3(例如,hvr-l1)内。在一些实施例中,一个或多个蛋氨酸残基位于轻链内;例如,位于位点m30、m33、m92。在一些实施例中,一个或多个蛋氨酸残基位于重链内;例如,位于位点m82、m99、m57、m58、m62、m64和95-102之间的其他位点。在一些实施例中,一个或多个蛋氨酸残基位于抗体的轻链恒定区和/或重链恒定区内。在一些实施例中,一个或多个蛋氨酸残基位于抗体(例如,igg1抗体)的重链恒定区内。在一些实施例中,一个或多个蛋氨酸残基包括m252、m35和/或m428(根据eu编号)。在一些实施例中,在本公开的制剂中包括l-蛋氨酸减少或防止抗体在残基m34、m82、m252和/或m428处的氧化(例如,与缺少l-蛋氨酸的液体制剂中多肽中一个或多个相应蛋氨酸残基相比)。在一些实施例中,在本公开的制剂中包括nat增加抗体在一个或多个蛋氨酸残基(例如,上述任何蛋氨酸残基,诸如在位置m252和/或m428的fc区蛋氨酸)的氧化。在一些实施例中,在制剂中包括l-蛋氨酸减少或防止抗体中一个或多个蛋氨酸残基(例如,上述任何蛋氨酸残基,诸如在位置m252、m358和/或m428的fc区蛋氨酸)的nat诱导和/或放大的氧化。在一些实施例中,本公开的液体制剂包含本文所述任何浓度的nat和本文所述任何浓度的l-蛋氨酸。在一些实施例中,液体制剂包含浓度为约0.3mm的nat和浓度为约5.0mm的l-蛋氨酸。在一些实施例中,液体制剂包含浓度为约1.0mm的nat和浓度为约5.0mm的l-蛋氨酸。在一些实施例中,本公开提供的液体制剂包含多肽、nat和l-蛋氨酸(其中nat和l-蛋氨酸减少或防止液体制剂中多肽的氧化),其中多肽的氧化(例如,多肽中一个或多个色氨酸残基和/或一个或多个蛋氨酸残基的氧化)减少约40%至约100%(例如,与缺少nat和/或l-蛋氨酸的液体制剂中多肽中一个或多个相应色氨酸残基和/或一个或多个相应蛋氨酸残基相比)。在一些实施例中,多肽的氧化(例如,多肽中一个或多个色氨酸残基和/或一个或多个蛋氨酸残基的氧化)减少约40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%(包括这些值之间的任何范围)中任一值(例如,与缺少nat和/或l-蛋氨酸的液体制剂中多肽中一个或多个相应色氨酸残基和/或一个或多个相应蛋氨酸残基相比)。可以使用本领域已知的任何合适的测量多肽的氧化的方法,包括例如以下实例1中描述的方法(以及其中引用的参考文献)。可以例如使用rp-hplc、lc/ms或胰蛋白酶肽图中的一种或多种来测定多肽中的氧化量。在一些实施例中,使用rp-hplc、lc/ms或胰蛋白酶肽作图中的一种或多种以及以下公式测定多肽中的氧化百分比:在一些实施例中,本公开提供的液体制剂包含多肽、nat和l-蛋氨酸(其中nat和l-蛋氨酸减少或防止液体制剂中多肽的氧化),其中不超过约40%至约0%的多肽被氧化(例如,在多肽中一个或多个色氨酸残基和/或一个或多个蛋氨酸残基处被氧化)。在一些实施例中,不超过约40%、35%、30%、25%、20%、15%、10%、5%、4%、3%、2%、1%或0%中的任何一个(包括这些值之间的任何范围)的多肽被氧化(例如,多肽中一个或多个色氨酸残基和/或一个或多个蛋氨酸残基被氧化)。在一些实施例中,本公开提供的液体制剂包含多肽、nat和l-蛋氨酸(其中nat和l-蛋氨酸减少或防止液体制剂中多肽的氧化),其中多肽中至少一个氧化不稳定色氨酸残基(例如,本文所述抗体的任何一个或多个色氨酸残基)的氧化减少约40%至约100%(例如,与缺少nat的制剂中多肽中一个或多个相应色氨酸残基相比)。在一些实施例中,多肽中氧化不稳定色氨酸残基的氧化减少约40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%中任一值,包括这些值之间的任何范围。在一些实施例中,多肽中每个氧化不稳定色氨酸残基的氧化减少约40%至约100%(诸如约40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%,包括这些值之间的任何范围)。在一些实施例中,本公开提供的液体制剂包含多肽、nat和l-蛋氨酸(其中nat和l-蛋氨酸减少或防止液体制剂中多肽的氧化),其中多肽中不超过约40%至约0%的至少一个氧化不稳定色氨酸残基(例如,本文所述抗体的任何一个或多个色氨酸残基)被氧化。在一些实施例中,多肽中不超过约40%、35%、30%、25%、20%、15%、10%、5%、4%、3%、2%、1%或0%中的任一值(包括这些值之间的任何范围)的氧化不稳定色氨酸残基被氧化。在一些实施例中,多肽中不超过约40%至约0%(诸如不超过约40%、35%、30%、25%、20%、15%、10%、5%、4%、3%、2%、1%或0%(包括这些值之间的任何范围)的每个氧化不稳定色氨酸残基被氧化。在一些实施例中,本公开提供的液体制剂包含多肽、nat和l-蛋氨酸(其中nat和l-蛋氨酸减少或防止液体制剂中多肽的氧化),其中多肽中至少一个氧化不稳定蛋氨酸残基(例如,本文所述抗体的任何一个或多个蛋氨酸残基)的氧化减少约40%至约100%(例如,与缺少l-蛋氨酸的制剂中多肽中一个或多个相应蛋氨酸残基相比)。在一些实施例中,多肽中氧化不稳定蛋氨酸残基的氧化减少约40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%中任一值,包括这些值之间的任何范围。在一些实施例中,多肽中每个氧化不稳定蛋氨酸残基的氧化减少约40%至约100%(诸如约40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%,包括这些值之间的任何范围)。在一些实施例中,本公开提供的液体制剂包含多肽、nat和l-蛋氨酸(其中nat和l-蛋氨酸减少或防止液体制剂中多肽的氧化),其中多肽中不超过约40%至约0%的至少一个氧化不稳定蛋氨酸残基(例如,本文所述抗体的任何一个或多个蛋氨酸残基)被氧化。在一些实施例中,多肽中不超过约40%、35%、30%、25%、20%、15%、10%、5%、4%、3%、2%、1%或0%中的任一值(包括这些值之间的任何范围)的氧化不稳定蛋氨酸残基被氧化。在一些实施例中,多肽中不超过约40%至约0%(诸如不超过约40%、35%、30%、25%、20%、15%、10%、5%、4%、3%、2%、1%或0%(包括这些值之间的任何范围)的每个氧化不稳定蛋氨酸残基被氧化。在一些实施例中,制剂中的多肽(例如,抗体)浓度为约1mg/ml至约250mg/ml。在一些实施例中,多肽(例如,抗体)为治疗性多肽。制剂中的示例性多肽浓度包括约1mg/ml至超过约250mg/ml、约1mg/ml至约250mg/ml、约10mg/ml至约250mg/ml、约15mg/ml至约225mg/ml、约20mg/ml至约200mg/ml、约25mg/ml至约175mg/ml、约25mg/ml至约150mg/ml、约25mg/ml至约100mg/ml、约30mg/ml至约100mg/ml或约45mg/ml至约55mg/ml。在一些实施例中,多肽为抗体。在一些实施例中,抗体为多克隆抗体、单克隆抗体、人源化抗体、人类抗体、嵌合抗体、多特异性抗体(例如,双特异性、三特异性等)或抗体片段。在一些实施例中,抗体源自igg1抗体、igg2抗体、igg3抗体或igg4抗体序列。在一些实施例中,抗体源自igg1抗体序列。在一些实施例中,制剂为水性制剂。在一些实施例中,制剂进一步包含一种或多种赋形剂。本领域已知的任何合适的赋形剂可以用于本文所述的制剂中,包括,例如,稳定剂、缓冲剂、表面活性剂、张度剂及其任何组合。例如,本公开的制剂可以包括单克隆抗体,如本文提供的防止多肽的氧化的nat(例如,在一个或多个色氨酸残基)、如本文提供的防止多肽的氧化的l-蛋氨酸(例如,在一个或多个蛋氨酸残基)和将制剂的ph值维持在理想水平的缓冲剂。在一些实施例中,本文提供的制剂的ph值为约4.5至约9.0。在一些实施例中,本文提供的制剂的ph值为约4.5至约7.0。在一些实施例中,所述ph值在ph值4.0至8.5的范围内、在ph值4.0至8.0的范围内、在ph值4.0至7.5的范围内、在ph值4.0至7.0的范围内、在ph值4.0至6.5的范围内、在ph值4.0至6.0的范围内、在ph值4.0至5.5的范围内、在ph值4.0至5.0的范围内、在ph值4.0至4.5的范围内、在ph值4.5至9.0的范围内、在ph值5.0至9.0的范围内、在ph值5.5至9.0的范围内、在ph值6.0至9.0的范围内、在ph值6.5至9.0的范围内、在ph值7.0至9.0的范围内、在ph值7.5至9.0的范围内、在ph值8.0至9.0的范围内、在ph值8.5至9.0的范围内、在ph值5.7至6.8的范围内、在ph值5.8至6.5的范围内、在ph值5.9至6.5的范围内、在ph值6.0至6.5的范围内、或在ph值6.2至6.5范围内。在一些实施例中,制剂的ph值为6.2或约6.2。在一些实施例中,制剂的ph值为6.0或约6.0。在一些实施例中,制剂进一步包括根据本文所述的任何多肽的至少一种另外的多肽。在一些实施例中,本文提供的制剂为适于施用于受试者的药物制剂。如本文所用,“受试者”、“患者”或“个体”可以指人类或非人类动物。“非人类动物”可以指非分类为人的任何动物,诸如家养、农场或动物园动物,运动、宠物动物(诸如犬、马、猫、牛等),以及用于研究的动物。研究动物可以是指(但不限于)线虫、节肢动物、脊椎动物、哺乳动物、青蛙、啮齿动物(例如,小鼠或大鼠)、鱼(例如,斑马鱼或河豚)、鸟类(例如,鸡)、犬、猫和非人类灵长类动物(例如,恒河猴、食蟹猴、黑猩猩等)。在一些实施例中,受试者、患者或个体为人类。制剂中的多肽和抗体可以使用本领域已知的任何合适方法进行制备。制剂中的抗体(例如,全长抗体、抗体片段和多特异性抗体)可以使用本领域可用的技术进行制备,其非限制性示例性方法在以下部分中更详细地描述。本领域的技术人员可以将本文的方法改编用于制备包括其他多肽诸如基于肽的抑制剂的制剂。关于生产治疗性蛋白的通常熟知的和常用的技术和方法参见molecularcloning:alaboratorymanual(sambrook等人,4thed.,coldspringharborlaboratorypress,coldspringharbor,n.y.,2012);currentprotocolsinmolecularbiology(f.m.ausubel,等人主编,2003);shortprotocolsinmolecularbiology(ausubel等人主编,j.wiley和sons,2002);currentprotocolsinproteinscience,(horswill等人,2006);antibodies,alaboratorymanual(harlow和lane主编,1988);cultureofanimalcells:amanualofbasictechniqueandspecializedapplications(r.i.freshney,6thed.,j.wiley和sons,2010),其所有通过引用整体并入本文。在一些实施例中,根据本文所述的任何制剂(例如,液体制剂),制剂包含两种或多种多肽(例如,制剂是两种或多种多肽的共制剂)。例如,在一些实施例中,制剂是包括两种或多种多肽、nat和l-蛋氨酸的共制剂,其中nat和l-蛋氨酸减少或防止所述两种或多种多肽中至少一种的氧化。在一些实施例中,nat和l-蛋氨酸减少或防止两个或多个多肽中多个的氧化。在一些实施例中,nat和l-蛋氨酸减少或防止两个或多个多肽每个的氧化。在一些实施例中,两个或多个多肽中至少一个为抗体,诸如多克隆抗体、单克隆抗体、人源化抗体、人类抗体、嵌合抗体、多特异性抗体或抗体片段。在一些实施例中,两个或多个多肽中的多个为抗体,诸如独立地选自多克隆抗体、单克隆抗体、人源化抗体、人类抗体、嵌合抗体、多特异性抗体或抗体片段的抗体。在一些实施例中,两个或多个多肽的每个为抗体,诸如独立地选自多克隆抗体、单克隆抗体、人源化抗体、人类抗体、嵌合抗体、多特异性抗体或抗体片段的抗体。在一些实施例中,制剂的一种或多种抗体源自igg1抗体序列。在一些实施例中,制剂为液体制剂。在一些实施例中,制剂为水性制剂。在一些实施例中,制剂为药物制剂(例如,适于施用于人类受试者)。在一些实施例中,药物制剂适于通过任何肠内途径或肠胃外途径施用。术语施用的“肠内途径”是指通过胃肠道任何部分施用。肠内途径的实例包括口服、粘膜、颊和直肠途径或胃内途径。施用的“肠胃外途径”是指肠内途径以外的施用途径。肠胃外途径的施用实例包括静脉内、肌内、皮内、腹腔、肿瘤内、膀胱内、动脉内、鞘内、囊内、眶内、玻璃体内、心内、经气管、关节内、囊下、蛛网膜下腔、脊柱内、硬膜外和胸骨内、皮下或局部施用。在一些实施例中,药物制剂适于皮下、静脉内或玻璃体内施用。在一些实施例中,药物制剂适于皮下或玻璃体内施用。a.抗体制备本文提供的液体制剂中的抗体直接抗目标抗原。优选地,抗原是生物学上重要的多肽,并且向患有病症的哺乳动物施用抗体可以在该哺乳动物中产生治疗益处。然而,还考虑了直接抗非多肽抗原的抗体。当抗原为多肽时,其可以为跨膜分子(例如,受体)或配体诸如生长因子。示例性抗原包括分子诸如血管内皮生长因子(vegf);cd20;ox-ldl;ox-apob100;肾素;生长激素,包括人类生长激素和牛生长激素;生长激素释放因子;甲状旁腺激素;促甲状腺激素;脂蛋白;α-1-抗胰蛋白酶;胰岛素a链;胰岛素b链;胰岛素原;卵泡刺激素;降钙素;促黄体素;胰高血糖素;凝血因子诸如viiic因子、ix因子、组织因子和维勒布兰德氏(vonwillebrands)因子;抗凝血因子诸如蛋白c;心钠素;肺表面活性剂;纤溶酶原激活剂,诸如尿激酶或人类尿或组织型纤溶酶原激活剂(t-pa);蛙皮素;凝血酶;造血生长因子;肿瘤坏死因子受体,诸如死亡受体5和cd120;肿瘤坏死因子-α和-β;脑啡肽酶;rantes(调节正常表达和分泌的t细胞活化);人类巨噬细胞炎性蛋白(mip-1-α);血清白蛋白诸如人类血清白蛋白;缪勒管(muellerian)抑制物质;松弛素a链;松弛素b链;松弛素原;小鼠促性腺激素相关肽;微生物蛋白,诸如β-内酰胺酶、脱氧核糖核酸酶;ige;细胞毒性t淋巴细胞相关抗原(ctla),诸如ctla-4;抑制素;激活素;激素或生长因子的受体;蛋白a或d;类风湿因子;神经营养因子诸如骨源性神经营养因子(bdnf),神经营养蛋白-3、-4、-5或-6(nt-3、nt-4、nt-5或nt-6)或神经生长因子诸如ngf-β;血小板衍生生长因子(pdgf);成纤维细胞生长因子诸如afgf和bfgf;表皮生长因子(egf);转化生长因子(tgf)诸如tgf-α和tgf-β,包括tgf-β1、tgf-β2、tgf-β3、tgf-β4或tgf-β5;胰岛素样生长因子-i和-ii(igf-i和igf-ii);des(1-3)-igf-i(脑igf-1),胰岛素样生长因子结合蛋白;cd蛋白诸如cd3、cd4、cd8、cd19和cd20;促红细胞生成素;骨诱导因子;免疫毒素;骨形态发生蛋白(bmp);干扰素诸如干扰素-α、-β和-γ;集落刺激因子(csf),例如,m-csf、gm-csf和g-csf;白介素(il),例如,il-1至il-10;超氧化物歧化酶;t细胞受体;表面膜蛋白;衰变加速因子;病毒抗原诸如,例如,aids包膜的一部分;转运蛋白;归巢受体;寻址蛋白;调节蛋白;整合素诸如cd11a、cd11b、cd11c、cd18、icam、vla-4和vcam;肿瘤相关抗原诸如her2、her3或her4受体;及以上列出的任何多肽的片段。(i)抗原制备任选地与其他分子缀合的可溶性抗原或其片段可以用作产生抗体的免疫原。对于跨膜分子,诸如受体,可以将其片段(例如受体的细胞外结构域)用作免疫原。可替代地,可以将表达跨膜分子的细胞用作免疫原。这样的细胞可以源自天然来源(例如癌细胞系),或者可以是已经通过重组技术转化以表达跨膜分子的细胞。可用于制备抗体的其他抗原及其形式对于本领域技术人员将是显而易见的。(ii)某些基于抗体的方法优选地通过多次皮下(sc)或腹腔(ip)注射相关抗原和佐剂在动物体内产生多克隆抗体。使用双功能或衍生化试剂,例如,马来酰亚胺基苯甲酰磺基琥珀酰亚胺(maleimidobenzoylsulfosuccinimideester)(通过半胱氨酸残基缀合)、n-羟基琥珀酰亚胺(通过赖氨酸残基缀合)、戊二醛、琥珀酸酐、socl2或r1n=c=nr(其中r和r1是不同的烷基基团),将相关抗原缀合至要免疫的物种中具有免疫原性的蛋白质(例如匙孔血蓝蛋白(keyholelimpethemocyanin)、血清白蛋白、牛甲状腺球蛋白或大豆胰蛋白酶抑制剂),可能会很有用。通过将例如100μg或5μg的蛋白或缀合物(分别用于兔或小鼠)与3体积的弗氏完全佐剂混合,并在多个部位皮内注射溶液,使动物对抗原、免疫原性缀合物或衍生物免疫。一个月后,通过在多个部位进行皮下注射,以弗氏完全佐剂中原始量的肽或缀合物的1/5至1/10的剂量增强动物的免疫力。7至14天后,给动物放血,并测定血清的抗体滴度。增强动物直至滴度稳定。优选地,用相同抗原的缀合物(但是缀合至不同的蛋白质和/或通过不同的交联剂)增强动物。缀合物也可以在重组细胞培养物中作为蛋白融合物制备。而且,聚集剂诸如明矾适合用于加强免疫应答。可以使用杂交瘤方法制备目标单克隆抗体,所述杂交瘤方法首先描述于kohler等人,nature,256:495(1975),并且进一步描述于例如hongo等人,hybridoma,14(3):253-260(1995),harlow等人,antibodies:alaboratorymanual,(coldspringharborlaboratorypress,第2版1988);hammerling等人,monoclonalantibodiesandt-cellhybridomas563-681(elsevier,n.y.,1981)和关于人类-人类杂交瘤的ni,xiandaimianyixue,26(4):265-268(2006)。另外的方法包括例如关于从杂交瘤细胞系生产单克隆人类天然igm抗体的美国专利号7,189,826中描述的那些。人类杂交瘤技术(trioma技术)描述于vollmers和brandlein,histologyandhistopathology,20(3):927-937(2005)以及vollmers和brandlein,methodsandfindingsinexperimentalandclinicalpharmacology,27(3):185-91(2005)。对于各种其他杂交瘤技术,参见,例如,us2006/258841;us2006/183887(完全人类抗体)、us2006/059575;us2005/287149;us2005/100546;us2005/026229和美国专利号7,078,492和7,153,507。使用杂交瘤方法产生单克隆抗体的示例性方案描述如下。在一个实施例中,小鼠或其他合适的宿主动物(诸如仓鼠),被免疫以引发淋巴细胞产生或能够产生特异性结合用于免疫的蛋白质的抗体。通过多次皮下(sc)或腹腔(ip)注射目标多肽或其片段和佐剂(诸如单磷酰脂质a(mpl)/海藻糖二硬脂酸酯(tdm)(ribiimmunochem.research,inc.,hamilton,mont.)来在动物中产生抗体。可以使用本领域熟知的方法,诸如重组方法,制备目标多肽(例如,抗原)或其片段,其中一些在本文中进一步描述。分析来自免疫动物的血清中的抗抗原抗体,并任选地施用加强免疫。从产生抗抗原抗体的动物中分离淋巴细胞。可替代地,可以体外免疫淋巴细胞。然后使用合适的融合剂(诸如聚乙二醇)将淋巴细胞与骨髓瘤细胞融合,以形成杂交瘤细胞。参见,例如,goding,monoclonalantibodies:principlesandpractice,第59-103页(academicpress,1986)。可以使用能有效融合的骨髓瘤细胞,支持所选抗体产生细胞稳定地高水平产生抗体,并对诸如hat培养基之类的培养基敏感。示例性骨髓瘤细胞包括但不限于鼠类骨髓瘤细胞系(诸如源自可从salkinstitutecelldistributioncenter,sandiego,calif.usa获得的mopc-21和mpc-11小鼠肿瘤,和源自可从americantypeculturecollection,rockville,md.usa获得的sp-2或x63-ag8-653细胞)。还已经描述了人类骨髓瘤和小鼠人类异源骨髓瘤细胞系用于人类单克隆抗体的产生(kozbor,j.immunol.,133:3001(1984);brodeur等人,monoclonalantibodyproductiontechniquesandapplications,第51-63页(marceldekker,inc.,newyork,1987))。将由此制备的杂交瘤细胞接种于并培养于合适的培养基(例如,含有一种或多种抑制未融合的亲代骨髓瘤细胞的生长或存活的物质的培养基)中。例如,若亲本骨髓瘤细胞缺乏酶次黄嘌呤鸟嘌呤转磷酸核糖基酶(hgprt或hprt),则用于杂交瘤的培养基通常将包括次黄嘌呤、氨喋呤及胸苷(hat培养基),这些物质阻止缺乏hgprt的细胞的生长。优选地,无血清杂交瘤细胞培养方法用于减少动物来源的血清诸如胎牛血清的使用,描述于例如even等人,trendsinbiotechnology,24(3),105-108(2006)。寡肽作为用于提高杂交瘤细胞培养物生产率的工具描述于franek,trendsinmonoclonalantibodyresearch,111-122(2005)。具体地,标准培养基富含某些氨基酸(丙氨酸、丝氨酸、天冬酰胺、脯氨酸)或蛋白水解物级分,并且由三至六个氨基酸残基组成的合成寡肽可显著抑制细胞凋亡。肽以毫摩尔或更高浓度存在。可以在杂交瘤细胞生长的培养基中测定与本文所述的抗体结合的单克隆抗体的产生。由杂交瘤细胞产生的单克隆抗体的结合特异性可通过免疫沉淀或通过体外结合测定诸如放射免疫测定(ria)或酶联免疫吸附测定(elisa)来测定。单克隆抗体的结合亲和力可以通过例如scatchard分析来测定。参见,例如,munson等人,anal.biochem.,107:220(1980)。一旦鉴定出产生具有所需特异性、亲和力和/或活性的抗体的杂交瘤细胞,便可通过有限稀释法对该克隆进行亚克隆并通过标准方法培养。参见,例如,goding,同上。用于此目的的合适培养基包括例如d-mem或rpmi-1640培养基。另外,杂交瘤细胞可以作为动物的腹水肿瘤在体内生长。通过常规免疫球蛋白纯化方法,诸如,例如,蛋白a-琼脂糖凝胶、羟磷灰石层析、凝胶电泳、透析或亲和层析,适当地将亚克隆物分泌的单克隆抗体与培养基、腹水或血清分离。从杂交瘤细胞分离蛋白质的一种方法描述于us2005/176122和美国专利号6,919,436。该方法包括在结合过程中使用最少的盐(诸如溶致盐),并且优选在洗脱过程中也使用少量的有机溶剂。(iii)某些文库筛选方法可以通过使用组合文库筛选具有所需活性或多种活性的抗体来制备本文所述的制剂和组合物中的抗体。例如,本领域已知多种方法用于产生噬菌体展示文库并筛选此类文库以获得具有所需结合特征的抗体。这样的方法通常描述于hoogenboom等人的methodsinmolecularbiology178:1-37(o’brien等人主编,humanpress,totowa,n.j.,2001)。例如,产生目标抗体的一种方法是通过使用噬菌体抗体文库,如lee等人,j.mol.biol.(2004),340(5):1073-93中所述。原则上,通过筛选含有显示与噬菌体外壳蛋白融合的抗体可变区(fv)的各种片段的噬菌体的噬菌体文库来选择合成抗体克隆。这种噬菌体文库通过亲和层析法对所需抗原进行淘选。表达能够与所需抗原结合的fv片段的克隆被吸附至抗原且因此与文库中的非结合克隆分离。然后将结合克隆从抗原洗脱,并且可通过附加的抗原吸附/洗脱循环进一步富集。抗体中的任一种可以通过以下方式获得:设计合适的抗原筛选程序以选择目标噬菌体克隆,随后使用来自目标噬菌体克隆的fv序列和合适的恒定区(fc)序列构建全长抗体克隆(如kabat等人,sequencesofproteinsofimmunologicalinterest,第五版,nihpublication91-3242,bethesdamd.(1991),第1-3卷所述)。在一些实施例中,抗体的抗原结合结构域由约110个氨基酸的两个可变(v)区形成,各来自轻链(vl)和重链(vh),二者均具有三个高变环(hvr)或互补决定区(cdr)。可变结构域可以以如下形式在功能上展示于噬菌体上:单链fv(scfv)片段,其中vh与vl经由短柔性肽共价连接;或fab片段,其中这些片段各自与恒定结构域融合并且非共价地相互作用,如winter等人,ann.rev.immunol.,12:433-455(1994)所述。如本文所用,编码scfv的噬菌体克隆和编码fab的噬菌体克隆统称为“fv噬菌体克隆”或“fv克隆”。vh与vl基因的谱系可通过聚合酶链反应(pcr)单独克隆并且在噬菌体文库中随机重组,然后可以针对抗原结合克隆对其进行搜索,如winter等人,ann.rev.immunol.,12:433-455(1994)所述。来自经免疫的来源的文库提供针对免疫原的高亲和力抗体,而无需构建杂交瘤。可替代地,初始谱系(naiverepertoire)可经克隆以便在无任何免疫作用下提供针对广泛非自体以及自体抗原的人类抗体的单一来源,如griffiths等人,emboj,12:725-734(1993)所述。最后,还可通过以下方式来制得初始文库:克隆来自干细胞的未重排的v基因区段;以及使用含有随机序列的pcr引物来编码高度可变的cdr3区域并完成体外重排,如由hoogenboom和winter,j.mol.biol.,227:381-388(1992)所述。在一些实施例中,丝状噬菌体用于通过与次要外壳蛋白piii融合来展示抗体片段。抗体片段可以展示为单链fv片段,其中vh和vl结构域通过柔性多肽间隔子连接在同一条多肽链上,例如,如marks等人,j.mol.biol.,222:581-597(1991)所述,或如fab片段,其中一条链与piii融合,另一条链被分泌到细菌宿主细胞周质中,在其中通过置换一些野生型外壳蛋白,fab-外壳蛋白结构的装配体在噬菌体表面上得以展示,例如,如hoogenboom等人,nucl.acidsres.,19:4133-4137(1991)所述。通常,编码抗体基因片段的核酸从人类或动物收获的免疫细胞处获得。如果所需偏向于抗抗原克隆的文库,则用抗原免疫受试者以产生抗体应答,并回收脾细胞和/或循环b细胞以及其他外周血淋巴细胞(pbl)以用于文库构建。在一个实施例中,通过在携带功能性人类免疫球蛋白基因阵列(并且缺少功能性内源抗体生产系统)的转基因小鼠中产生抗抗原抗体应答,使得抗原免疫引起b细胞产生针对抗原的人类抗体来获得偏向于抗抗原克隆的人类抗体基因片段文库。下文描述了产生人类抗体的转基因小鼠的产生。通过使用合适的筛选程序分离表达抗原特异性膜结合抗体的b细胞,例如,通过使用抗原亲和层析法进行细胞分离或将细胞吸附到荧光染料标记的抗原上,然后进行流式激活细胞分选(facs),可以获得抗抗原反应性细胞群的额外富集。可替代地,非免疫供体的脾细胞和/或b细胞或其他pbl的使用提供了更好地可能的抗体谱表示,也可以使用任何动物(人类或非人类)物种构建抗体文库,其中抗原不具有抗原性。对于并入体外抗体基因构建的文库,从受试者收获干细胞以提供编码未重排抗体基因区段的核酸。目标免疫细胞可以从多种动物物种中获得,诸如人类、小鼠、大鼠、兔类、狼类、犬、猫、猪、牛、马和禽类等。从目标细胞中回收并扩增编码抗体可变基因区段(包括vh和vl区段)的核酸。对于重排的vh和vl基因文库,可以通过从淋巴细胞中分离基因组dna或mrna,然后用与重排vh和vl基因的5′和3′末端匹配的引物进行聚合酶链反应(pcr)来获得所需dna,如orlandi等人,proc.natl.acad.sci.(usa,),86:3833-3837(1989)所述,从而制备多种用于表达的v基因谱。可以从cdna和基因组dna扩增v基因,使用编码成熟的v结构域的外显子5′端的反向引物,以及基于在j区段内的正向引物,如orlandi等人(1989)和ward等人,nature,341:544-546(1989)中所述。然而,为了从cdna扩增,反向引物也可以基于jones等人,biotechnol.,9:88-89(1991)中所述的前导外显子,以及如sastry等人,proc.natl.acad.sci.(usa),86:5728-5732(1989)中所述的恒定区内的正向引物。为了最大化互补性,可以在引物中引入简并性如orlandi等人(1989)或sastry等人(1989)所述。在一些实施例中,通过使用靶向v基因家族各自的pcr引物以最大化文库多样性,以扩增存在于免疫细胞核酸样品中的所有可用的vh和vl排列,例如,如marks等人,j.mol.biol.,222:581-597(1991)所述的方法,或如orum等人,nucleicacidsres.,21:4491-4498(1993)所述的方法。为了将扩增的dna克隆到表达载体中,稀有的限制性位点可以如orlandi等人(1989)所述在一端作为标签引入pcr引物内,或如clackson等人,nature,352:624-628(1991)中所述通过使用加标签的引物的进一步pcr扩增。合成的重排v基因谱可以在体外从v基因区段获得。大多数人类的vh基因区段已被克隆和测序(如tomlinson等人,j.mol.biol.,227:776-798(1992)报道),以及图谱分析(如matsuda等人,naturegenet.,3:88-94(1993)报道);这些克隆区段(包括h1环和h2环的所有主要构象)可用于通过编码不同序列和长度的h3环的pcr引物产生多样化的vh基因谱,如hoogenboom和winter,j.mol.biol.,227:381-388(1992)所述。还可以将所有序列多样性集中在单个长度的长h3环中制备vh谱,如barbas等人,proc.natl.acad.sci.usa,89:4457-4461(1992)所述。人类vκ和vλ区段已被克隆和测序(如williams和winter,eur.j.immunol.,23:1456-1461(1993)报道),并且可以用于制备合成的轻链谱。基于一定范围的vh和vl倍数以及l3和h3长度,合成v基因谱将编码具有相当结构多样性的抗体。在扩增编码dna的v基因之后,可以根据hoogenboom和winter,j.mol.biol.,227:381-388(1992)的方法体外重排种系v基因区段。抗体片段谱可以通过几种方式将vh和vl基因谱组合在一起来构建。每个谱可以在不同的载体中创建,并且载体在体外重组,例如,如hogrefe等人,gene,128:119-126(1993)中所述,或通过体内组合感染,例如,loxp系统,如waterhouse等人,nucl.acidsres.,21:2265-2266(1993)所述。体内重组方法利用fab片段的双链性质,以克服大肠杆菌(e.coli)转化效率对文库大小的限制。将初始vh和vl谱分别克隆,一个克隆到噬菌粒中,另一个克隆到噬菌体载体中。然后通过噬菌体感染含噬菌粒的细菌将两个文库合并,以使细胞各自包含不同的组合,并且文库大小仅受存在的细胞数限制(约1012个克隆)。两种载体均包含体内重组信号,因此vh和vl基因重组到单个复制子上,并共同包装在噬菌体病毒体中。这些庞大的文库提供了大量具有良好亲和力的抗体(kd-1约为10-8m)。可替代地,可以将谱依次克隆到相同的载体中,例如,如barbas等人,proc.natl.acad.sci.usa,88:7978-7982(1991)中所述,或通过pcr组装在一起然后克隆,例如,如clackson等人,nature,352:624-628(1991)中所述。pcr组装也可用于将vh和vl的dna与编码柔性肽间隔子的dna连接起来,形成单链fv(scfv)谱。在又一技术中,“细胞内pcr组装”用于通过pcr结合淋巴细胞内的vh和vl基因,然后克隆连接基因谱,如embleton等人,nucl.acidsres.,20:3831-3837(1992)中所述。初始文库(天然或合成)产生的抗体可以具有中等亲和力(kd-1约为106至107m-1),但亲和力成熟也可以通过体外构建和从二级文库中重新选择来模拟,如winter等人(1994)(同上)所述。例如,可以使用易错聚合酶在体外随机引入突变(如leung等人,technique1:11-15(1989)报道),如hawkins等人,j.mol.biol.,226:889-896(1992)中的方法或如gram等人,proc.natl.acad.sciusa,89:3576-3580(1992)中的方法。另外,亲和力成熟可以通过随机突变一个或多个cdr来执行,例如,在选定的单个fv克隆中,使用带有跨目标cdr的随机序列的引物进行pcr,并筛选亲和力更高的克隆。wo9607754(公开于1996年3月14日)描述了在免疫球蛋白轻链的互补决定区诱导诱变以产生轻链基因文库的方法。另一种有效的方法是将通过噬菌体展示选择的vh或vl结构域与从未免疫供体获得的天然存在的v结构域变体谱重组,并筛选几轮链改组以获得更高的亲和力,如marks等人,biotechnol.,10:779-783(1992)中所述。该技术实现生产亲和力为约10-9m或更小的抗体和抗体片段。文库的筛选可通过本领域中已知的各种技术完成。例如,抗原可用于包被吸附板的孔,在附着于吸附板的宿主细胞上表达,或在细胞分选中使用,或缀合至生物素以用于链霉亲和素包被的珠粒的捕获,或在用于淘选噬菌体展示文库的任何其他方法中使用。在适于使至少一部分噬菌体颗粒与吸附剂结合的条件下,将噬菌体文库样品与固定化抗原接触。通常,选择包括ph值、离子强度、温度等的条件以模仿生理条件。洗涤与固相结合的噬菌体,然后用酸洗脱,例如,如barbas等人,proc.natl.acad.sciusa,88:7978-7982(1991)所述,或用碱洗脱,例如,如marks等人,j.mol.biol.,222:581-597(1991)所述,或通过抗原竞争,例如,以类似于clackson等人,nature,352:624-628(1991)的抗原竞争方法的程序进行。单轮选择可使噬菌体富集20-1,000倍。此外,富集的噬菌体可以在细菌培养物中生长,并进行进一步的选择。选择的效率取决于许多因素,包括洗涤过程中解离的动力学,以及单个噬菌体上的多个抗体片段是否可以同时与抗原结合。具有快速解离动力学(和较弱的结合亲和力)的抗体可以通过短时间洗涤、多价噬菌体展示和高固相抗原包被密度来保留。高密度不但通过多价相互作用稳定噬菌体,而且有利于已解离的噬菌体的重新结合。可通过使用如bass等人,proteins,8:309-314(1990)和wo92/09690中所述的长时间洗涤与单价噬菌体展示以及如marks等人,biotechnol.,10:779-783(1992)中所述的低包被密度的抗原来促进具有慢解离动力学(及良好的结合亲和力)的抗体的选择。对于抗原,即使在亲和力略有不同的情况下,也可以在具有不同亲和力的噬菌体抗体之间进行选择。然而,选定抗体的随机突变(例如,如在某些亲和力成熟技术中所进行的)可能产生许多突变体,大多数与抗原结合,而另一些具有更高的亲和力。利用限制性抗原,可以竞争稀有的高亲和力噬菌体。为了保留所有更高亲和力的突变体,可以将噬菌体与过量的生物素化抗原共同孵育,但生物素化抗原的浓度应低于对抗原的目标摩尔亲和常数的摩尔浓度。然后,高亲和力结合噬菌体可被链霉亲和素包被的顺磁珠捕获。这样的“平衡捕获”实现根据抗体的结合亲和力选择抗体,其敏感性实现从具有低亲和力的大量过量的噬菌体中分离出具有低至两倍高亲和力的突变体克隆。还可以基于解离动力学来控制用于洗涤结合至固相的噬菌体的条件以进行区分。可以基于活性选择抗抗原克隆。在一些实施例中,本公开提供了与天然表达抗原或与游离浮动抗原或附着于其他细胞结构的抗原结合的活细胞结合的抗抗原抗体。可以通过以下方式选择与此类抗抗原抗体相对应的fv克隆:(1)如上所述从噬菌体文库中分离抗抗原克隆,并任选地通过在合适的细菌宿主中使该噬菌体克隆的群体逐渐生长而使其扩增;(2)选择需要分别针对其阻断和非阻断活性的抗原和第二蛋白;(3)将抗抗原噬菌体克隆吸附到固定的抗原上;(4)使用过量的第二蛋白洗脱任何非所需克隆,这些克隆识别与第二蛋白的结合决定簇重叠或共享的抗原结合决定簇;(5)洗脱在步骤(4)之后仍吸附的克隆。任选地,可以通过重复一次或多次本文所述的选择程序来进一步富集具有所需阻断/非阻断性质的克隆。编码杂交瘤来源的单克隆抗体或噬菌体展示fv克隆的dna易于使用常规程序进行分离和测序(例如,使用设计为从杂交瘤或噬菌体dna模板中特异性扩增目标重链和轻链编码区的寡核苷酸引物)。一旦分离,可将dna置于表达载体中,然后将该表达载体转染至不另外产生免疫球蛋白的宿主细胞诸如大肠杆菌细胞、猿cos细胞、中国仓鼠卵巢(cho)细胞或骨髓瘤细胞中,以在重组宿主细胞中获得所需单克隆抗体的合成。关于在编码抗体的dna的细菌中的重组表达的综述论文包括skerra等人,curr.opinioninimmunol.,5:256(1993)和pluckthun,immunol.revs,130:151(1992)。可以将编码fv克隆的dna与编码重链和/或轻链恒定区的已知dna序列(例如,合适的dna序列可以从kabat等人(同上)获得)组合以形成编码全长或部分长度的重链和/或轻链的克隆。应当理解,任何同种型的恒定区均可以用于该目的,包括igg、igm、iga、igd和ige恒定区,并且这种恒定区可以从任何人类或动物物种获得。从一个动物(诸如人类)物种的可变结构域dna衍生的fv克隆,然后与另一动物物种的恒定区dna融合,以形成“杂交”的编码序列,全长重链和/或轻链包括在本文所用的“嵌合”和“杂交”抗体的定义中。在一些实施例中,将源自人类可变dna的fv克隆与人类恒定区dna融合,以形成全长或部分长度的人类重链和/或轻链的编码序列。还可以修饰编码来自杂交瘤的抗抗原抗体的dna,例如,通过取代人类重链和轻链恒定结构域的编码序列来代替源自杂交瘤克隆的同源鼠序列(例如,如morrison等人,proc.natl.acad.sci.usa,81:6851-6855(1984)的方法)。可以通过将非免疫球蛋白多肽的全部或部分编码序列共价连接至免疫球蛋白编码序列,进一步修饰编码源自杂交瘤或fv克隆的抗体或片段的dna。以这种方式,制备具有fv克隆或杂交瘤克隆来源的抗体的结合特异性的“嵌合”或“杂交”抗体。(iv)人源化和人类抗体人源化非人类抗体的各种方法是本领域已知的。例如,人源化抗体具有从非人类引入的一个或多个氨基酸残基。这些非人类氨基酸残基通常被称为“导入”残基,其通常取自“导入”可变结构域。人源化可以基本上按照winter和其同事的方法进行(jones等人,nature,321:522-525(1986);riechmann等人,nature,332:323-327(1988);verhoeyen等人,science,239:1534-1536(1988)),通过用啮齿动物的cdr或cdr序列代替人类抗体的相应序列。因此,这样的“人源化”抗体是嵌合抗体(美国专利号4,816,567),其中基本上少于完整的人类可变结构域已被来自非人类物种的相应序列取代。在实践中,人源化抗体通常为人类抗体,其中一些cdr残基和可能的一些fr残基被啮齿动物抗体中类似位点的残基取代。用于制备人源化抗体的人类可变结构域的轻链和重链的选择对于降低抗原性非常重要。根据所谓的“最佳拟合”方法,针对已知的人类可变结构域序列的整个文库筛选啮齿动物抗体的可变结构域的序列。然后将最接近啮齿动物的人类序列作为人源化抗体的人类框架(fr)(sims等人,j.immunol.,151:2296(1993);chothia等人,j.mol.biol.,196:901(1987))。另一方法使用来自轻链或重链特定亚组的所有人类抗体的共有序列的特定框架。相同的框架可用于几种不同的人源化抗体(carter等人,proc.natl.acadsci.usa,89:4285(1992);presta等人,j.immunol.,151:2623(1993))。进一步重要的是将抗体人源化,同时保留对抗原的高亲和力和其他有利的生物特性。为了实现该目的,根据所述方法的一个实施例,通过使用亲本和人源化序列的三维模型对亲本序列和各种概念性人源化产物进行分析的过程来制备人源化抗体。三维免疫球蛋白模型是常规可获得的并且为本领域技术人员所熟悉。提供计算机程序,其说明并显示了所选的候选免疫球蛋白序列的可能的三维构象结构。对这些展示的检查允许分析残基在候选免疫球蛋白序列的功能中的可能的作用,即,分析影响候选免疫球蛋白结合其抗原的能力的残基。以这种方式,可以从受体和导入序列中选择并结合fr残基,从而获得所需的抗体特性,诸如对靶抗原的亲和力增加。通常,高变区残基直接且最大程度地参与影响抗原结合。本文所述的制剂和组合物中的人类抗体可以通过将选自人源噬菌体展示文库的fv克隆可变结构域序列与已知的人类恒定结构域序列组合来构建。可替代地,可以通过杂交瘤方法制备人类单克隆抗体。用于生产人类单克隆抗体的人类骨髓瘤和小鼠-人类异源骨髓瘤细胞系已经,例如,如kozborj.immunol.,133:3001(1984);brodeur等人,monoclonalantibodyproductiontechniquesandapplications,pp.51-63(marceldekker,inc.,newyork,1987);和boerner等人,j.immunol.,147:86(1991)所述。可能产生转基因动物(例如,小鼠),其在免疫后能够在不产生内源性免疫球蛋白的情况下产生人类抗体的完整谱。例如,已经描述了在嵌合和种系突变小鼠中抗体重链连接区(jh)基因的纯合缺失导致内源抗体产生的完全抑制。人类种系免疫球蛋白基因阵列在这种种系突变小鼠中的转移将导致在抗原攻击后产生人类抗体。参见,例如,jakobovits等人,proc.natl.acad.sci.usa,90:2551(1993);jakobovits等人,nature,362:255-258(1993);bruggermann等人,yearinimmuno.,7:33(1993);和duchosal等人nature355:258(1992)。基因改组还可以用于从非人类,例如,啮齿动物抗体获得人类抗体,其中人类抗体与起始非人类抗体具有相似的亲和力和特异性。根据此方法(也称为“表位印迹”),将通过本文所述的噬菌体展示技术获得的非人类抗体片段的重链或轻链可变区替换为人类v结构域基因谱,创建非人类链/人类链scfv或fab嵌合体群。用抗原进行选择可分离出非人类链/人类链嵌合scfv或fab,其中人类链可恢复去除初代噬菌体展示克隆中相应的非人类链时被破坏的抗原结合位点,即表位控制(印迹)人类链配偶体的选择。当重复该过程以取代剩余的非人类链时,获得人类抗体(参见1993年4月1日公开的pctwo93/06213)。与传统的通过cdr移植非人类抗体的人源化不同,该技术提供了完全的人类抗体,其没有非人类来源的fr或cdr残基。(v)抗体片段抗体片段可以通过传统方法(诸如酶消化)或通过重组技术产生。在某些情况下,使用抗体片段而不是全抗体具有优势。片段的较小尺寸允许快速清除,并可以改善对实体瘤的进入。关于某些抗体片段的综述,参见hudson等人(2003)nat.med.9:129-134。已经开发了用于产生抗体片段的各种技术。传统上,这些片段通过完整抗体的蛋白水解消化而获得(参见,例如,morimoto等人,dournalofbiochemicalandbiophysicalmethods24:107-117(1992);和brennan等人,science,229:81(1985))。然而,这些片段现在可以直接由重组宿主细胞产生。fab、fv和scfv抗体片段均可在大肠杆菌中表达并从大肠杆菌中分泌出来,因此可轻松生产大量这些片段。可以从以上讨论的抗体噬菌体文库中分离抗体片段。可替代地,fab′-sh片段可以直接从大肠杆菌中回收,并化学偶联形成f(ab′)2片段(carter等人,bio/technology10:163-167(1992))。根据另一方案,可以直接从重组宿主细胞培养物中分离f(ab′)2片段。具有增加的体内半衰期的包含挽救受体结合表位残基的fab和f(ab′)2片段如美国专利号5,869,046中所述。产生抗体片段的其它技术对熟练技术人员将是显而易见的。在一些实施例中,抗体为单链fv片段(scfv)。参见wo93/16185;美国专利号5,571,894和5,587,458。fv和scfv是具有完整结合位点没有恒定区的仅有的种类;因此,它们可能适合于在体内使用期间减少非特异性结合。可以构建scfv融合蛋白以在scfv的氨基末端或羧基末端产生效应蛋白的融合。参见antibodyengineering,ed.borrebaeck,同上。例如,该抗体片段也可以是“线性抗体”,例如,如美国专利号5,641,870中所述。这样的线性抗体可以是单特异性或双特异性的。(vi)多特异性抗体多特异性抗体对至少两个不同的表位具有结合特异性,其中这些表位通常来自不同的抗原。尽管这样的分子通常仅结合两个不同的表位(即双特异性抗体,bsab),但是当在本文中使用时,该表述包括具有其他特异性的抗体诸如三特异性抗体。可以将双特异性抗体制成全长抗体或抗体片段(例如,f(ab′)2双特异性抗体)。制备双特异性抗体的方法是本领域已知的。全长双特异性抗体的传统生产是基于两个免疫球蛋白重链-轻链对的共表达,其中两条链具有不同的特异性(millstein等人,nature,305:537-539(1983))。由于免疫球蛋白重链和轻链的随机多样性,这些杂交瘤(四源杂交瘤)产生10种不同抗体分子的潜在混合物,其中只有一种具有正确的双特异性结构。通常通过亲和层析步骤完成的正确分子的纯化相当麻烦,并且产物产量低。在wo93/08829和traunecker等人,emboj.,10:3655-3659(1991)中公开了类似的过程。根据不同的方法,将具有所需结合特异性的抗体可变结构域(抗体-抗原结合位点)与免疫球蛋白恒定结构域序列融合。融合优选地使用免疫球蛋白重链恒定结构域,包含铰链的至少一部分、ch2和ch3区。通常具有至少一个融合物中存在的包含轻链结合所需位点的第一重链恒定区(ch1)。将编码免疫球蛋白重链融合物和(如果需要)免疫球蛋白轻链的dna插入单独的表达载体中,并共转染到合适的宿主生物中。当在构建中使用的三个多肽链的不相等比例提供最佳产量时,这在实施例中提供了调节三个多肽片段的相互比例的极大灵活性。然而,当至少两个多肽链以相等的比率表达导致高产量或当比率没有特别意义时,有可能在一个表达载体中插入两条或全部三条多肽链的编码序列。在该方法的一个实施例中,双特异性抗体由在一个臂中具有第一结合特异性的杂交免疫球蛋白重链和在另一臂中具有杂交免疫球蛋白重链-轻链对(提供第二结合特异性)组成。已经发现,这种不对称结构促进了所需的双特异性化合物与不需要的免疫球蛋白链组合的分离,因为仅在双特异性分子的一半中存在免疫球蛋白轻链提供了一种简便的分离方式。该方法在wo94/04690中公开。对于生成双特异性抗体的更多细节,参见,例如,suresh等人,methodsinenzymology,121:210(1986)。根据wo96/27011中描述的另一种方法,可以对一对抗体分子之间的界面进行工程化以最大化从重组细胞培养物中回收的异源二聚体的百分比。一个界面包含抗体恒定结构域的ch3结构域的至少一部分。在该方法中,用较大的侧链(例如,酪氨酸或色氨酸)替换来自第一抗体分子的界面的一个或多个小氨基酸侧链。具有与大侧链相同或相似大小的补偿性“腔”是通过用较小的氨基酸侧链(例如,丙氨酸或苏氨酸)替换大氨基酸侧链在第二抗体分子的界面上创建。这提供了一种机制,可以提高异源二聚体而不是其他不需要的最终产物(诸如同二聚体)的产量。双特异性抗体包括交联或“异源缀合物”抗体。例如,异源缀合物中的一种抗体可以与抗生物素蛋白偶联,另一种与生物素偶联。例如,已经提出了这样的抗体以将免疫系统细胞靶向不需要的细胞(美国专利号4,676,980),以及用于治疗hiv感染(wo9i/00360、wo92/200373和ep03089)。异源缀合物抗体可以使用任何方便的交联方法制备。合适的交联剂是本领域众所周知的,并在美国专利号4,676,980以及许多交联技术中描述。从抗体片段产生双特异性抗体的技术也已在文献中描述。例如,可以使用化学连接来制备双特异性抗体。brennan等人,science,229:81(1985)描述了一种过程,其中完整抗体被蛋白水解裂解以生成f(ab′)2片段。在二硫醇络合剂亚砷酸钠的存在下还原这些片段,以稳定邻位二硫醇并防止分子间二硫化物的形成。然后将生成的fab′片段转化为硫代硝基苯甲酸酯(tnb)衍生物。然后通过巯基乙胺还原将其中一种fab′-tnb衍生物重新转化为fab′-硫醇,并与等摩尔量的另一种fab′-tnb衍生物混合以形成双特异性抗体。产生的双特异性抗体可以用作选择性固定酶的试剂。最近的进展促进了从大肠杆菌中直接回收fab′-sh片段,该片段可以化学偶联以形成双特异性抗体。shalaby等人,j.exp.med.,175:217-225(1992)描述了完全人源化的双特异性抗体f(ab′)2分子的产生。fab′片段各自分别从大肠杆菌中分泌,并在体外进行定向化学偶联以形成双特异性抗体。还已经描述了用于直接从重组细胞培养物中制备和分离双特异性抗体片段的各种技术。例如,已经使用亮氨酸拉链产生了双特异性抗体。kostelny等人,j.immunol.,148(5):1547-1553(1992)。通过基因融合,将来自fos蛋白和jun蛋白的亮氨酸拉链肽连接至两种不同抗体的fab′部分。抗体同源二聚体在铰链区还原形成单体,然后再氧化形成抗体异源二聚体。该方法也可以用于抗体同源二聚体的生产。hollinger等人,proc.natl.acad.sci.usa,90:6444-6448(1993)描述的“双体抗体”技术已经提供了制备双特异性抗体片段的替代机制。片段包含通过接头连接至轻链可变结构域(vh)的重链可变结构域(vl),该接头太短以至于不允许同一条链上的两个结构域之间配对。因此,一个片段的vh和vl,结构域被迫与另一片段的互补的vl,和vh结构域配对,从而形成两个抗原结合位点。还已经报道了通过使用单链fv(sfv)二聚体制备双特异性抗体片段的另一种策略。参见gmber等人,j.immunol.,152:5368(1994)。考虑了具有两个以上价态的抗体。例如,可以制备三特异性抗体。tuft等人j.immunel.147:60(1991)。(vii)单结构域抗体在一些实施例中,本文所述的抗体为单结构域抗体。单结构域抗体为包含抗体的全部或部分重链可变结构域或全部或部分轻链可变结构域的单个多肽链。在一些实施例中,单结构域抗体为人类单结构域抗体(domantis,inc.,waltham,mass.;参见,例如,美国专利号6,248,516b1)。在一个实施例中,单结构域抗体由抗体的全部或部分重链可变结构域组成。(viii)抗体变体在一些实施例中,考虑了本文描述的抗体的氨基酸序列修饰。例如,可能期望改善抗体的结合亲和力和/或其他生物特性。抗体的氨基酸序列变体可以通过向编码抗体的核苷酸序列中引入适当的改变或通过肽合成来制备。此类修饰包括例如抗体氨基酸序列内残基的缺失和/或插入和/或取代。可以进行缺失、插入和取代的任何组合以获得最终构建体,前提条件是所述最终构建体具有所需特性。可以在形成序列时将氨基酸改变引入目标抗体的氨基酸序列中。(ix)抗体衍生物本公开的制剂和组合物中的抗体可以被进一步修饰以包含本领域已知并且易于获得的另外的非蛋白部分。在一些实施例中,适合于抗体衍生的部分是水溶性聚合物。水溶性聚合物的非限制性实例包括但不限于聚乙二醇(peg)、乙二醇/丙二醇的共聚物、羧甲基纤维素、葡聚糖、聚乙烯醇、聚乙烯吡咯烷酮、聚-1,3-二氧戊环、聚-1,3,6-三氧杂环己烷、乙烯/马来酸酐共聚物、聚氨基酸(均聚物或随机共聚物)和葡聚糖或聚(n-乙烯吡咯烷酮)聚乙二醇、丙二醇均聚物、聚环氧丙烷/环氧乙烷共聚物、聚氧乙烯化多元醇(例如,甘油)、聚乙烯醇以及它们的混合物。由于其在水中的稳定性,聚乙二醇丙醛在制造中可具有优势。聚合物可具有任何分子量,并且可以具有支链或不具有支链。附接至抗体的聚合物的数目可变,并且如果附接了多于一个聚合物,那么它们可以为相同或不同的分子。通常,可基于以下考虑因素测定用于衍生化的聚合物的数目和/或类型,包括但不限于抗体待改善的特定特性或功能、抗体衍生物是否将用于限定条件下的疗法等。(x)载体、宿主细胞和重组方法抗体也可以使用重组方法生产。对于抗抗原抗体的重组生产,将编码抗体的核酸分离并插入可复制的载体中以进行进一步克隆(dna的扩增)或表达。编码抗体的dna可以使用常规过程容易地进行分离和测序(例如,通过使用能够与编码抗体的重链和轻链的基因特异性结合的寡核苷酸探针)。许多载体可用。载体组分通常包括但不限于以下一种或多种:信号序列、复制起点、一种或多种标记基因、增强子元件、启动子和转录终止序列。(a)信号序列组分本文所述的制剂和组合物中的抗体不仅可以直接重组产生,而且可以作为与异源多肽的融合多肽重组产生,所述异源多肽优选为信号序列或在成熟蛋白或多肽的n末端具有特定裂解位点的其他多肽。优选地,所选择的异源信号序列是被宿主细胞识别和加工(例如,被信号肽酶裂解)的异源信号序列。对于不识别和加工天然抗体信号序列的原核宿主细胞,该信号序列被原核信号序列取代,该原核信号序列例如选自由碱性磷酸酶、青霉素酶、1pp或热稳定肠毒素ii前导序列组成的组。对于酵母分泌而言,天然信号序列可以被例如酵母转化酶前导序列、因子前导序列(包括酿酒酵母(saccharomyces)和克鲁维酵母(kluyveromyces)α-因子前导序列)或酸性磷酸酶前导序列、白色念珠菌(c.albicans)葡糖淀粉酶前导序列或wo90/13646中描述的信号序列取代。在哺乳动物细胞表达中,可获得哺乳动物信号序列以及病毒分泌前导序列,例如,单纯疱疹gd信号。(b)复制起点表达载体和克隆载体都包含使载体能够在一个或多个所选宿主细胞中复制的核酸序列。通常,在克隆载体中,该序列是使载体能够独立于宿主染色体dna复制的序列,并包括复制起点或自主复制序列。用于多种细菌、酵母和病毒的此类序列是众所周知的。质粒pbr322的复制起点适用于大多数革兰氏阴性细菌,2μ质粒的复制起点适用于酵母,并且各种病毒复制起点(sv40、多瘤病毒、腺病毒、vsv或bpv)可用于在哺乳动物细胞中克隆载体。通常,哺乳动物表达载体不需要复制起点组分(通常仅可使用sv40起点,因为其包含早期启动子)。(c)选择基因组分表达和克隆载体可以包含选择基因,也称为选择性标记物。典型的选择基因编码的蛋白,该蛋白(a)赋予对抗生素或其他毒素(例如,氨苄青霉素、新霉素、氨甲蝶呤或四环素)的耐药性,(b)补足营养缺陷型缺陷,或(c)供应无法从复合培养基中获得的关键营养素,例如,编码杆菌属(bacilli)d-丙氨酸消旋酶的基因。选择方案的一个实例利用一种药物来阻滞宿主细胞的生长。用异源基因成功转化的那些细胞产生赋予药物耐药性的蛋白质,从而在选择方案中生存下来。这种显性选择的实例使用新霉素、霉酚酸和潮霉素。哺乳动物细胞的合适选择标记物的另一实例是那些能够鉴别能够摄取编码抗体的核酸的细胞的标记物,诸如dhfr、谷氨酰胺合成酶(gs)、胸苷激酶、金属硫蛋白-i和金属硫蛋白-ii(优选灵长类金属硫蛋白基因)、腺苷脱氨酶、鸟氨酸脱羧酶等。例如,通过在含有氨甲蝶呤(mtx)(dhfr的竞争性拮抗剂)的培养基中培养转化体来鉴定用dhfr基因转化的细胞。在这些条件下,dhfr基因与任何其他共转化的核酸一起扩增。可以使用缺乏内源dhfr活性的中国仓鼠卵巢(cho)细胞系(例如,atcccrl-9096)。可替代地,通过在含有l-蛋氨酸硫代亚胺(msx)(gs抑制剂)的培养基中培养转化体来鉴定用gs基因转化的细胞。在这些条件下,gs基因与任何其他共转化的核酸一起扩增。gs选择/扩增系统可以与上述dhfr选择/扩增系统联合使用。可替代地,可以通过在含有用于选择性标记物的选择剂(诸如氨基糖苷类抗生素,例如,卡那霉素、新霉素或g418)的培养基中进行细胞生长,来选择用编码目标抗体、野生型dhfr基因和另一种选择性标记物诸如氨基糖苷3′-磷酸转移酶(aph)的dna序列转化或共转化的宿主细胞(特别是包含内源dhfr的野生型宿主)。参见美国专利号4,965,199。用于酵母的合适选择基因是存在于酵母质粒yrp7中的trp1基因(stinchcomb等人,nature,282:39(1979))。该trp1基因为缺乏在色氨酸中生长能力的酵母突变株(例如atcc号44076或pep4-1)提供选择标记物。jones,genetics,85:12(1977)。然后,酵母宿主细胞基因组中trp1病灶的存在为通过在色氨酸不存在下生长来检测转化提供了有效的环境。类似地,leu2缺陷型酵母菌株(atcc20,622或38,626)由带有leu2基因的已知质粒补充。此外,源自1.6μm环状质粒pkd1的载体可用于转化克鲁维(kluyveromyces)酵母。可替代地,报道了采用乳酸克鲁维酵母(k.lactis)来大规模生产重组小牛凝乳酶的表达系统。vandenberg,bio/technology,8:135(1990)。还公开了用于通过克鲁维(kluyveromyces)酵母工业菌株分泌成熟的重组人类血清白蛋白的稳定的多拷贝表达载体。fleer等人,bio/technology,9:968-975(1991)。(d)启动子组分表达和克隆载体通常包含被宿主生物识别并与编码抗体的核酸可操作连接的启动子。适用于原核宿主的启动子包括phoa启动子、β-内酰胺酶和乳糖启动子系统、碱性磷酸酶启动子、色氨酸(trp)启动子系统以及杂合启动子诸如tac启动子。然而,其他已知的细菌启动子也适用。用于细菌系统的启动子还将包含与编码抗体的dna可操作连接的shine-dalgarno(s.d.)序列。真核生物的启动子序列是已知的。实际上,所有真核基因具有富含at的区域,位于转录起始位点上游约25个至30个碱基的位置。在许多基因转录开始的上游发现70个至80个碱基的另一序列是cncaat区,其中n可以是任何核苷酸。大多数真核基因的3′末端是aataaa序列,该序列可能是将polya尾添加到编码序列3′末端的信号。将所有这些序列适当地插入真核表达载体中。与酵母宿主一起使用的合适启动子序列的实例包括3-磷酸甘油酸激酶或其他糖酵解酶的启动子,诸如烯醇酶、3-磷酸甘油醛脱氢酶、己糖激酶、丙酮酸脱羧酶、磷酸果糖激酶、6-磷酸葡萄糖异构酶、3-磷酸甘油酸突变酶、丙酮酸激酶、磷酸三糖异构酶、磷酸葡萄糖异构酶和葡萄糖激酶。其他酵母启动子是具有受生长条件控制的转录的其他优势的诱导型启动子,它们是乙醇脱氢酶2、异细胞色素c、酸性磷酸酶、氮代谢相关的降解酶、金属硫蛋白、甘油三磷酸甘油脱氢酶、以及负责麦芽糖和半乳糖利用的酶。ep73,657中进一步描述了用于酵母表达的合适的载体和启动子。酵母增强子还有利地与酵母启动子一起使用。哺乳动物宿主细胞中载体的抗体转录可以例如通过以下控制:从病毒基因组获得的启动子,该病毒诸如多瘤病毒、鸡痘病毒、腺病毒(诸如腺病毒2)、牛乳头瘤病毒、禽肉瘤病毒、巨细胞病毒、逆转录病毒、乙型肝炎病毒、猿猴病毒40(sv40),或来自异源哺乳动物启动子,例如,肌动蛋白启动子或免疫球蛋白启动子,来自热激启动子,前提是这些启动子与宿主细胞系统相容。sv40病毒的早期和晚期启动子可方便地以sv40限制性片段的形式获得,其中也包含sv40病毒的复制起点。人类巨细胞病毒的立即早期启动子可以以hindiiie限制性片段方便地获得。美国专利号4,419,446公开了使用牛乳头瘤病毒作为载体在哺乳动物宿主中表达dna的系统。该系统的修饰如美国专利号4,601,978所述。另请参见reyes等人,nature297:598-601(1982)关于在来自单纯疱疹病毒的胸苷激酶启动子的控制下,人类β-干扰素cdna在小鼠细胞中的表达。可替代地,劳斯肉瘤病毒(roussarcomavirus)长末端重复序列可用作启动子。(e)增强子元件组分通常通过将增强子序列插入载体中来增加高级真核生物对编码抗体的dna转录。现在从哺乳动物基因中已知许多增强子序列(球蛋白、弹性蛋白酶、白蛋白、α-甲胎蛋白和胰岛素)。然而,通常,将使用来自真核细胞病毒的增强子。实例包括在复制起点(bp100-270)的后侧的sv40增强子、巨细胞病毒早期启动子增强子、在复制起点的后侧的多瘤增强子和腺病毒增强子。另请参见yaniv,nature297:17-18(1982)关于增强真核启动子激活的元件。增强子可以在抗体编码序列的5′或3′位置剪接到载体中,但优选地位于启动子的5′位点。(f)转录终止组分真核宿主细胞(酵母、真菌、昆虫、植物、动物、人类或其他多细胞生物的有核细胞)中使用的表达载体还将包含终止转录和稳定mrna所需的序列。此类序列通常可从真核或病毒dna或cdna的5′(有时是3′)非翻译区获得。这些区域包含在编码抗体的mrna的非翻译部分中转录为聚腺苷酸化片段的核苷酸区段。一种有用的转录终止组分为牛生长激素聚腺苷酸化区。参见wo94/11026及其中公开的表达载体。(g)宿主细胞的选择和转化用于在本文的载体中克隆或表达dna的合适宿主细胞是上述原核生物、酵母或更高等的真核细胞。为此目的,合适的原核生物包括真细菌诸如例如革兰氏阴性或革兰氏阳性生物,例如,肠杆菌科,诸如埃希氏杆菌属(escherichia)例如大肠杆菌(e.coli)、肠杆菌属(enterobacter)、欧文氏菌属(erwinia)、克雷伯氏杆菌属(klebsiella)、变形杆菌属(proteus)、沙门氏菌属(salmonella)例如鼠伤寒沙门氏菌(salmonellatyphimurium)、沙雷氏菌属(serratia)例如粘质沙雷氏菌(serratiamarcescans)和志贺氏杆菌属(shigella),以及芽孢杆菌属(bacilli)诸如枯草芽孢杆菌(b.subtilis)和地衣芽孢杆菌(b.licheniformis)(例如,1989年4月12日公布的dd266,710中公开的地衣芽孢杆菌(b.licheniformis)41p),假单胞菌属(pseudomonas)诸如绿脓假单胞菌(p.aeruginosa),和链霉菌属(streptomyces)。一种优选的大肠杆菌克隆宿主是大肠杆菌294(atcc31,446),但其他菌株例如大肠杆菌b、大肠杆菌x1776(atcc31,537)和大肠杆菌w3110(atcc27,325)也是合适的。这些实例是说明性的而不是限制性的。全长抗体、抗体融合蛋白和抗体片段可在细菌中产生,尤其是在不需要糖基化和fc效应子功能时,诸如当治疗性抗体与自身的细胞毒剂(例如,毒素)偶联时显示出对肿瘤细胞破坏的有效性。全长抗体在循环中具有更长的半衰期。在大肠杆菌中的生产速度更快且更具成本效益。关于抗体片段和多肽在细菌中的表达,参见,例如,美国专利号5,648,237(carter等人)、美国专利号5,789,199(joly等人)、美国专利号5,840,523(simmons等人),这些专利描述了用于优化表达和分泌的翻译起始区(tir)和信号序列。另请参见charlton,methodsinmolecularbiology,第248卷(b.k.c.lo,ed.,humanapress,totowa,n.j.,2003),第245-254页,其描述抗体片段在大肠杆菌中的表达。表达后,可以将抗体以可溶级分的形式从大肠杆菌细胞糊中分离出来,并可以根据同种型通过例如蛋白a或g柱纯化。最终纯化可以类似于纯化例如在cho细胞中表达的抗体的方法进行。除原核生物外,真核微生物诸如丝状真菌或酵母也是编码抗体的载体的合适克隆或表达宿主。酿酒酵母(saccharomycescerevisiae)或普通面包酵母是低等真核宿主微生物中最常用的酵母。然而,许多其他属、种和菌株在本文中是普遍可获得的和有用的,诸如粟酒裂殖酵母(schizosaccharomycespombe);克鲁维酵母属(kluyveromyces)宿主诸如,例如,乳酸克鲁维酵母(k.lactis)、脆壁克鲁维酵母(k.fragilis)(atcc12,424)、保加利亚克鲁维酵母(k.bulgaricus)(atcc16,045)、维氏克鲁维酵母(k.wickeramii)(atcc24,178)、沃氏克鲁维酵母(k.waltii)(atcc56,500)、果蝇克鲁维酵母(k.drosophilarum)(atcc36,906)、耐热克鲁维酵母(k.thermotolerans)和马克斯克鲁维酵母(k.marxianus);耶氏酵母(yarrowia)(ep402,226);巴斯德毕赤酵母(pichiapastoris)(ep183,070);假丝酵母(candida);瑞氏木霉(trichodermareesia)(ep244,234);粗糙脉孢菌(neurosporacrassa);许旺酵母属(schwanniomyces)诸如西方许旺酵母(schwanniomycesoccidentalis);以及丝状真菌诸如,例如,脉孢菌属(neurospora)、青霉属(penicillium)、弯颈霉属(tolypocladium)和曲霉菌(aspergillus)宿主诸如构巢曲霉(a.nidulans)和黑曲霉(a.niger)。有关讨论使用酵母和丝状真菌生产治疗性蛋白的综述,参见,例如,gerngross,nat.biotech.22:1409-1414(2004)。可以选择某些其中糖基化途径已经被“人源化”的真菌和酵母菌株,从而产生具有部分或完全人类糖基化模式的抗体。参见,例如,li等人,nat.biotech.24:210-215(2006)(描述巴斯德毕赤酵母(pichiapastoris)中糖基化途径的人源化);和gemgross等人,同上。用于表达糖基化抗体的合适宿主细胞也来源于多细胞生物(无脊椎动物和脊椎动物)。无脊椎动物细胞的实例包括植物细胞和昆虫细胞。已经鉴定出许多杆状病毒株和变体以及来自宿主的相应的允许昆虫宿主细胞,这些宿主诸如草地贪夜蛾(spodopterafrugiperda)(毛虫)、埃及伊蚊(aedesaegypti)(蚊)、白纹伊蚊(aedesalbopictus)(蚊)、黑腹果蝇(drosophilamelanogaster)(果蝇)和家蚕(bombyxmori)。各种用于转染的病毒株是可公开获得的,例如,苜蓿银纹夜蛾(autographacalifornica)npv的l-1变体和家蚕(bombyxmori)npv的bm-5株,并且此类病毒可以用作本文中根据本发明的病毒,特别是用于转染草地贪夜蛾(spodopterafrugiperda)细胞。棉花、玉米、马铃薯、大豆、矮牵牛、番茄、浮萍(唇形科(leninaceae))、苜蓿(蒺藜苜蓿(m.truncatula))和烟草的植物细胞培养物也可用作宿主。参见,例如,美国专利号5,959,177、6,040,498、6,420,548、7,125,978和6,417,429(描述了用于在转基因植物中产生抗体的plantibodiestm技术)。脊椎动物细胞可以用作宿主,并且脊椎动物细胞在培养(组织培养)中的繁殖已成为常规过程。有用的哺乳动物宿主细胞系的实例是经sv40(cos-7,atcccrl1651)转化的猴肾cv1系;人类胚胎肾细胞系(亚克隆用于在悬浮培养中生长的293或293细胞,graham等人,j.genvirol.36:59(1977));幼仓鼠肾细胞(bhk,atccccl10);小鼠支持细胞(tm4,mather,biol.reprod.23:243-251(1980));猴肾细胞(cv1,atccccl70);非洲绿猴肾细胞(vero-76,atcccrl-1587);人类宫颈癌细胞(hela,atccccl2);犬肾细胞(mdck,atccccl34);水牛大鼠肝细胞(brl3a,atcccrl1442);人类肺细胞(w138,atccccl75);人类肝细胞(hepg2,hb8065);小鼠乳腺肿瘤(mmt060562,atccccl51);tri细胞(mather等人,annalsn.y.acad.sci.383:44-68(1982));mrc5细胞;fs4细胞;以及人类肝癌细胞系(hepg2)。其他有用的哺乳动物宿主细胞系包括中国仓鼠卵巢(cho)细胞,其包括dhfr-cho细胞(urlaub等人,proc.natl.acad.sci.usa77:4216(1980));以及骨髓瘤细胞系诸如ns0和sp2/0。关于适用于抗体生产的某些哺乳动物宿主细胞系的综述,参见,例如,yazaki和wu,methodsinmolecularbiology,第248卷(b.k.c.lo,ed.,humanapress,totowa,n.j.,2003),第255-268页。用上述用于抗体生产的表达或克隆载体转化宿主细胞,并在常规营养培养基中培养,所述营养培养基经适当修饰以诱导启动子、选择转化子或扩增编码所需序列的基因。(h)培养宿主细胞用于产生抗体的宿主细胞可以在多种培养基中培养。市售培养基诸如ham′sf10(sigma)、基本必需培养基((mem),(sigma)、rpmi-1640(sigma)和dulbecco改良的eagle培养基((dmem),sigma)适用于培养宿主细胞。另外,在ham等人,meth.enz.58:44(1979),barnes等人,anal.biochem.102:255(1980),美国专利号4,767,704、4,657,866、4,927,762、4,560,655或5,122,469;wo90/03430;wo87/00195或美国再公告专利30,985中描述的任何培养基可用作宿主细胞的培养基。这些介质中的任何一种都可以根据需要补充激素和/或其他生长因子(诸如胰岛素、转铁蛋白或表皮生长因子)、盐(诸如氯化钠、钙、镁和磷酸盐)、缓冲剂(诸如hepes)、核苷酸(诸如腺苷和胸苷)、抗生素(诸如gentamycintm药物)、微量元素(定义为通常以微摩尔范围的最终浓度存在的无机化合物)和葡萄糖或等效能源。也可以以本领域技术人员已知的适当浓度包括任何其他必要的补充剂。培养条件诸如温度、ph值等,是先前与选择用于表达的宿主细胞一起使用的条件,对于普通技术人员而言是显而易见的。(xi)抗体纯化当使用重组技术时,抗体可以在细胞内在周质间隙中产生,或直接分泌到培养基中。如果抗体在细胞内产生,则作为第一步,例如通过离心或超滤移除宿主细胞或裂解片段的微粒碎片。carter等人,bio/technology10:163-167(1992)中描述了分离分泌到大肠杆菌周质间隙中的抗体的方法。简而言之,将细胞糊状物在乙酸钠(ph3.5)、edta和苯甲基磺酰氟(pmsf)的存在下解冻约30分钟。可以通过离心移除细胞碎片。在抗体被分泌到培养基中的情况下,通常首先使用可商购的蛋白质浓缩过滤器,例如amicon或milliporepellicon超滤单元,浓缩来自此类表达系统的上清液。蛋白酶抑制剂诸如pmsf可以包含在前述任何步骤中以抑制蛋白水解作用并且抗生素可以包含在内以防止外来污染物的生长。可以使用例如羟磷灰石层析、疏水相互作用色谱、凝胶电泳、透析和亲和层析纯化从细胞制备的抗体组合物,其中亲和层析是通常优选的纯化步骤之一。蛋白a作为亲和配体的适用性取决于抗体中存在的任何免疫球蛋白fc结构域的种类和同种型。蛋白a可用于纯化基于人类γ1、γ2或γ4重链的抗体(lindmark等人,j.immunol.meth.62:1-13(1983))。蛋白g被推荐用于所有小鼠同种型和人类γ3(guss等人,emboj.5:15671575(1986))。蛋白l可用于纯化基于κ轻链的抗体(nilson等人,j.immunol.meth.164(1):33-40,1993)。亲和配体所附接的基质大多是琼脂糖,但也可以使用其他基质。机械稳定的基质,诸如可控孔度玻璃或聚(苯乙烯二乙烯基),具有比琼脂糖更快的流速和更短的处理时间。当抗体包含ch3结构域时,bakerbondabxtm树脂(j.t.baker,phillipsburg,n.j.)可用于纯化。也可使用其他蛋白纯化技术诸如在离子交换柱上进行分级、乙醇沉淀、反相hplc、硅胶色谱、肝素色谱、在阴离子或阳离子交换树脂(诸如聚天冬氨酸柱)上进行sepharosetm色谱、色谱聚焦、sds-page和硫酸铵沉淀,取决于待回收的抗体。通常,与上述方法一致和/或被本领域技术人员认为适合于特定目的抗体的制备用于研究、测试和临床的抗体的各种方法是本领域中已经建立的。b.选择生物学活性抗体可以对如上所述产生的抗体进行一种或多种“生物学活性”测定,以从治疗性角度选择具有有益特性的抗体。可以筛选抗体与它所针对的抗原结合的能力。例如,对于抗dr5抗体(例如,曲齐妥单抗),可以在检测与死亡受体5(dr5)结合能力的测定法中评价抗体的抗原结合特性。在另一实施例中,抗体的亲和力可以通过例如饱和结合、elisa和/或竞争测定(例如,ria)所测定。而且,可以对抗体进行其他生物学活性测定,例如,以评价其作为治疗剂的有效性。此类测定是本领域已知的,并且取决于靶抗原和抗体的预期用途。为了筛选与目标抗原上特定表位结合的抗体,需要进行常规的交叉阻断测定诸如antibodies,alaboratorymanual,coldspringharborlaboratory,edharlowanddavidlane(1988)所述。可替代地,表位作图,例如,如champe等人,j.biol.chem.270:1388-1394(1995)中所述,可以用来测定抗体是否结合目标表位。iii.制备制剂的方法本公开的某些方面涉及制备本文所述的任何液体制剂的方法。可以通过将具有所需纯度的多肽与nat和l-蛋氨酸混合来制备液体制剂。在一些实施例中,待配制的多肽未经预先冷冻干燥,并且本文的目标制剂为水性制剂。在一些实施例中,多肽为治疗性蛋白。在一些实施例中,多肽为抗体。在进一步的实施例中,抗体为多克隆抗体、单克隆抗体、人源化抗体、人类抗体、嵌合抗体、多特异性抗体、双特异性抗体或抗体片段。在一些实施例中,抗体为全长抗体。在一些实施例中,制剂中的抗体为抗体片段,诸如f(ab′)2,在这种情况下,可能需要解决使用全长抗体时可能不会发生的问题(诸如将抗体剪接到fab)。制剂中存在的多肽的治疗有效量例如通过考虑所需剂量体积和施用方式来测定。制剂中的示例性多肽浓度包括约1mg/ml至超过约250mg/ml、约1mg/ml至约250mg/ml、约10mg/ml至约250mg/ml、约15mg/ml至约225mg/ml、约20mg/ml至约200mg/ml、约25mg/ml至约175mg/ml、约25mg/ml至约150mg/ml、约25mg/ml至约100mg/ml、约30mg/ml至约100mg/ml或约45mg/ml至约55mg/ml。在一些实施例中,本文所述的多肽易于氧化。在一些实施例中,蛋白中选自蛋氨酸、半胱氨酸、组氨酸、色氨酸和/或酪氨酸的一种或多种氨基酸易于氧化。在一些实施例中,多肽中的一个或多个色氨酸易于氧化。在一些实施例中,多肽中的一个或多个蛋氨酸易于氧化。在一些实施例中,多肽中的一个或多个色氨酸和一个或多个蛋氨酸易于氧化。在一些实施例中,液体制剂进一步包含一种或多种赋形剂,诸如稳定剂、缓冲剂、表面活性剂和/或张度剂。在ph缓冲溶液中制备本公开的液体制剂。本公开的缓冲剂的ph值在约4.0至约9.0的范围内。在一些实施例中,所述ph值在ph值4.0至8.5的范围内、在ph值4.0至8.0的范围内、在ph值4.0至7.5的范围内、在ph值4.0至7.0的范围内、在ph值4.0至6.5的范围内、在ph值4.0至6.0的范围内、在ph值4.0至5.5的范围内、在ph值4.0至5.0的范围内、在ph值4.0至4.5的范围内、在ph值4.5至9.0的范围内、在ph值5.0至9.0的范围内、在ph值5.5至9.0的范围内、在ph值6.0至9.0的范围内、在ph值6.5至9.0的范围内、在ph值7.0至9.0的范围内、在ph值7.5至9.0的范围内、在ph值8.0至9.0的范围内、在ph值8.5至9.0的范围内、在ph值5.7至6.8的范围内、在ph值5.8至6.5的范围内、在ph值5.9至6.5的范围内、在ph值6.0至6.5的范围内、或在ph值6.2至6.5范围内。在本公开的一些实施例中,液体制剂的ph值为6.2或约6.2。在本公开的一些实施例中,液体制剂的ph值为6.0或约6.0。在本公开的一些实施例中,液体制剂的ph值为5.8或约5.8。在本公开的一些实施例中,液体制剂的ph值为5.5或约5.5。将ph值控制在该范围内的缓冲剂的实例包括有机酸和无机酸及其盐。例如,乙酸盐(例如,乙酸组氨酸、乙酸精氨酸、乙酸钠)、琥珀酸酯(例如,组氨酸琥珀酸酯、精氨酸琥珀酸酯、琥珀酸钠)、葡萄糖酸酯、磷酸盐、富马酸酯、草酸酯、乳酸酯、柠檬酸酯及其组合。缓冲剂浓度可以为约1mm至约600mm,这取决于例如缓冲剂和制剂的所需等渗性。在一些实施例中,制剂包含组氨酸缓冲剂(例如,浓度为约5mm至100mm)。组氨酸缓冲剂的实例包括组氨酸氯化物、乙酸组氨酸、磷酸组氨酸、硫酸组氨酸、琥珀酸组氨酸等。在一些实施例中,制剂中的组氨酸为约10mm至约35mm、约10mm至约30mm、约10mm至约25mm、约10mm至约20mm、约10mm至约15mm、约15mm至约35mm、约20mm至约35mm、约20mm至约30mm或约20mm至约25mm。在进一步的实施例中,制剂中的精氨酸为约50mm至约500mm(例如,约100mm、约150mm或约200mm)。本公开的液体制剂可以进一步包含糖,诸如二糖(例如,海藻糖或蔗糖)。本文所用的“糖”包括常规成分(ch2o)n及其衍生物,包括单糖、二糖、三糖、多糖、糖醇、还原糖、非还原糖等。本文糖的实例包括葡萄糖、蔗糖、海藻糖、乳糖、果糖、麦芽糖、右旋糖酐、甘油、右旋糖酐、赤藓糖醇、甘油、阿糖醇、山糖醇、山梨糖醇、甘露糖醇、偏二糖、松三糖、棉子糖、甘露三糖、水苏糖、麦芽糖、乳果糖、麦芽酮糖、葡糖醇、麦芽糖醇、乳糖醇、异麦芽酮糖等。在一些实施例中,制剂包含蔗糖。可以任选地将表面活性剂添加到液体制剂中。示例性表面活性剂包括非离子表面活性剂诸如聚山梨酯(例如,聚山梨酯20、80等)或泊洛沙姆(例如,泊洛沙姆188等)。添加的表面活性剂的量应使得其减少所配制抗体的聚集和/或最小化制剂中颗粒的形成和/或减少吸附。例如,表面活性剂可以约0.001%至超过约1.0%(重量/体积)的量存在于制剂中。在一些实施例中,表面活性剂以约0.001%至约1.0%、约0.001%至约0.5%、约0.005%至约0.2%、约0.01%至约0.1%、约0.02%至约0.06%、或约0.03%至约0.05%(重量/体积)的量存在于制剂中。在一些实施例中,表面活性剂以0.04%或约0.04%的量(重量/体积)存在于制剂中。在一些实施例中,表面活性剂以0.02%或约0.02%的量(重量/体积)存在于制剂中。在一个实施例中,制剂不包含表面活性剂。在一个实施例中,制剂包含上述试剂(例如,抗体、缓冲剂、糖和/或表面活性剂),并且基本上不含一种或多种防腐剂,诸如苯甲醇、苯酚、间甲酚、氯丁醇和苄索氯铵。在另一实施例中,制剂中可以包含防腐剂,特别是其中制剂为多剂量制剂。防腐剂的浓度可以在约0.1%至约2%的范围内,优选约0.5%至约1%。制剂中可以包含一种或多种其他药用运载体、赋形剂或稳定剂,诸如remington’spharmaceuticalsciences16thedition,osol,a.ed.(1980)中描述的那些,只要它们不会不利地影响制剂的所需特征即可。本文的示例性药用赋形剂进一步包含间质药物分散剂诸如可溶性中性活性透明质酸酶糖蛋白(shasegp),例如,人类可溶性ph-20透明质酸酶糖蛋白,诸如rhuph20(baxterinternational,inc.)。某些示例性shasegp和使用方法,包括rhuph20,描述于美国专利公开号2005/0260186和2006/0104968中。在一个方面中,将shasegp与一种或多种另外的糖胺聚糖酶(诸如软骨素酶)组合。制剂可以进一步包含金属离子螯合剂。金属离子螯合剂为本领域技术人员熟知的,包括但不限于氨基聚羧酸盐、edta(乙二胺四乙酸)、egta(乙二醇-双(β-氨基乙基醚)-n,n,n′,n′-四乙酸)、nta(亚硝基三乙酸)、edds(乙二胺二琥珀酸酯)、pdta(1,3-丙二胺四乙酸)、dtpa(二亚乙基三胺五乙酸)、ada(β-丙氨酸二乙酸)、mgca(甲基甘氨酸二乙酸)等。另外,本文的一些实施例包含膦酸酯/膦酸螯合剂。存在张度剂以调节或维持组合物中液体的张度。当与带电荷的大型生物分子(诸如蛋白和抗体)一起使用时,其还可以充当“稳定剂”,因为其可以与氨基酸侧链的带电基团相互作用,从而降低分子间和分子内相互作用的可能性。考虑到其他成分的相对量,张度剂可以以0.1重量%至25重量%,或更优选地以1重量%至5重量%的任何量存在。优选的张度剂包括多元糖醇,优选地三元或更高级的糖醇,诸如甘油、赤藓糖醇、阿拉伯糖醇、木糖醇、山梨糖醇和甘露糖醇。对于所治疗的特定适应症,如有必要本文所述的制剂还可以包含多于一种多肽或小分子药物,优选地具有对其他多肽没有不利影响的互补活性的那些多肽。例如,其中抗体为抗dr5(例如,曲齐妥单抗)时,可以与另一药物(例如,化疗剂和抗肿瘤剂)组合使用。在一些实施例中,制剂施用于体内。在一些实施例中,制剂为无菌的。可以通过无菌滤膜过滤制得无菌制剂。本文中的治疗性制剂通常置于具有无菌进入口的容器中,例如,具有可被皮下注射针刺穿的塞子的静脉内溶液袋或小瓶。施用途径是根据已知和公认的方法,诸如以适当方式通过长时间单次或多次推注或输注,例如,通过皮下、静脉内、腹腔、肌内、动脉内、病灶内、关节内或玻璃体内途径注射或输注,局部施用,吸入或通过持续释放或延迟释放方式。本公开的液体制剂在储存时可以是稳定的。在一些实施例中,液体制剂中的多肽在约0℃至约5℃(诸如约1℃、2℃、3℃或4℃中的任一温度)下储存至少约12个月(诸如至少约15、18、21、24、27、30、33、36个月或更长时间)是稳定的。在一些实施例中,评估或测量液体制剂中多肽的物理稳定性、化学稳定性或生物学活性。可以使用本领域已知的任何方法来评估稳定性和生物学活性。在一些实施例中,通过储存后液体制剂中多肽的氧化来测量其稳定性。可以通过在配制时以及在储存后评估制剂中抗体的物理稳定性、化学稳定性和/或生物学活性来测试其稳定性。可通过各种不同的方式定性和/或定量评估物理和/或稳定性,包括评估聚集体形成(例如,使用尺寸排阻色谱法、通过测量浊度和/或通过目视检查);通过使用阳离子交换色谱或毛细管区带电泳评估电荷异质性;氨基末端或羧基末端序列分析;质谱分析;sds-page分析,用于比较还原态和完整抗体;肽图(例如胰蛋白酶或lys-c)分析;评估抗体的生物学活性或抗原结合功能等。不稳定可能产生聚集、脱酰胺(例如,asn脱酰胺)、氧化(例如,trp氧化)、异构化(例如,asp异构化)、剪切/水解/片段化(例如,铰链区片段化)、琥珀酰亚胺形成、未配对的半胱氨酸、n末端延伸、c末端处理、糖基化差异等。在一些实施例中,使用rp-hplc、lc/ms或胰蛋白酶肽作图中的一种或多种来测定蛋白中的氧化。在一些实施例本文还提供了制备液体制剂或防止液体制剂中的多肽的氧化的方法,该方法包括添加可减少或防止液体制剂中多肽的氧化的一定量的nat和l-蛋氨酸。在一些实施例中,液体制剂包含抗体。减少或防止多肽的氧化的一定量的nat和l-蛋氨酸可以是本文公开的任何量。iv.减少氧化的方法本公开的某些方面涉及减少液体制剂中的多肽(例如,本文描述的任何多肽)的氧化的方法,该方法包括添加可减少或防止液体制剂中多肽的氧化的一定量的nat和一定量的l-蛋氨酸。在一些实施例中,包括nat和l-蛋氨酸的液体制剂是本文所述的任何液体制剂。在一些实施例中,多肽易于氧化。在一些实施例中,多肽中的一种或多种蛋氨酸、半胱氨酸、组氨酸、色氨酸和/或酪氨酸残基易于氧化。在一些实施例中,多肽中一个或多个色氨酸残基易于氧化。在一些实施例中,多肽中一个或多个蛋氨酸残基易于氧化。在一些实施例中,多肽中一个或多个色氨酸和一个或多个蛋氨酸残基易于氧化。在一些实施例中,多肽为治疗性多肽。在一些实施例中,多肽为抗体。在一些实施例中,制剂进一步包括根据本文所述的任何多肽的至少一种另外的多肽。在一些实施例中,制剂进一步包含一种或多种赋形剂。在一些实施例中,制剂为水性制剂。在一些实施例中,制剂为药物制剂(例如,适于施用于人类受试者)。例如,本公开的制剂可以包含如本文提供的单克隆抗体、nat和l-蛋氨酸,其防止单克隆抗体的氧化(例如,在抗体中的一个或多个色氨酸残基和一个或多个蛋氨酸残基处),以及将制剂的ph值维持在期望水平的缓冲剂。在一些实施例中,制剂的ph值为约4.5至约7.0。在一些实施例中,添加至制剂中的nat的量是本文提供的任何浓度的nat。在一些实施例中,添加至制剂中的nat的量为约0.3mm。在一些实施例中,添加至制剂中的nat的量为约1.0mm。在一些实施例中,nat减少或防止多肽中一个或多个色氨酸残基(例如,如本文所述的抗体的一个或多个色氨酸残基中的任何一个)的氧化。在一些实施例中,多肽的氧化(例如,多肽中一个或多个色氨酸残基的氧化)减少约40%至约100%,诸如减少约40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%中任一值,包括这些值之间的任何范围(例如,与缺少nat的液体制剂中多肽中一个或多个相应色氨酸残基相比)。在一些实施例中,不超过约40%至约0%,诸如不超过约40%、35%、30%、25%、20%、15%、10%、5%、4%、3%、2%、1%或0%(包括这些值之间的任何范围)的多肽被氧化(例如,多肽中的一个或多个色氨酸残基被氧化)。在一些实施例中,nat防止多肽被活性氧(ros)氧化。在一些实施例中,添加至制剂中的l-蛋氨酸的量是本文提供的l-蛋氨酸的任何浓度。在一些实施例中,添加至制剂中的l-蛋氨酸的量为约5.0mm。在一些实施例中,l-蛋氨酸减少或防止多肽中一个或多个蛋氨酸残基(例如,本文所述抗体的一个或多个蛋氨酸残基中的任何一个)的氧化。在一些实施例中,多肽的氧化(例如,多肽中一个或多个蛋氨酸残基的氧化)减少约40%至约100%,诸如减少约40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%中任一值,包括这些值之间的任何范围(例如,与缺少l-蛋氨酸的液体制剂中多肽中一个或多个相应蛋氨酸残基相比)。在一些实施例中,不超过约40%至约0%,诸如不超过约40%、35%、30%、25%、20%、15%、10%、5%、4%、3%、2%、1%或0%(包括这些值之间的任何范围)的多肽被氧化(例如,多肽中的一个或多个蛋氨酸残基被氧化)。在一些实施例中,l-蛋氨酸防止多肽被活性氧(ros)氧化。在一些实施例中,制剂中的多肽(例如,抗体)浓度是本文所述的任何多肽浓度(例如,约1mg/ml至约250mg/ml)。在一些实施例中,多肽为治疗性多肽。在一些实施例中,多肽为抗体。在一些实施例中,抗体为多克隆抗体、单克隆抗体、人源化抗体、人类抗体、嵌合抗体、多特异性抗体(例如,双特异性、三特异性等)或抗体片段。在一些实施例中,抗体源自igg1抗体、igg2抗体、igg3抗体或igg4抗体序列。在一些实施例中,抗体源自igg1抗体序列。在一些实施例中,制剂进一步包含一种或多种赋形剂。本领域已知的任何合适的赋形剂可以用于本文所述的制剂中,包括,例如,稳定剂、缓冲剂、表面活性剂、张度剂及其任何组合。在一些实施例中,制剂的ph值约为本文所述的任何ph值(例如,约4.5至约7.0)。v.制剂的施用本公开的某些方面涉及本文所述的任何制剂向受试者的施用。在一些实施例中,本公开的液体制剂可以用于制备适合施用于受试者的药物(例如,用于治疗或预防受试者的癌症)。可以根据已知方法将液体制剂施用于需要接受多肽(例如,抗体)治疗的受试者(例如,人类),诸如推注或通过一段时间内的连续输注的静脉内施用,通过肌内、腹腔、脑脊髓内、皮下、关节内、滑膜内、鞘内、口服、局部、吸入或玻璃体内途径施用。在一些实施例中,通过静脉内、玻璃体内或皮下施用将液体制剂施用于受试者。在一些实施例中,通过玻璃体内施用将液体制剂施用于受试者。在一些实施例中,通过皮下施用将液体制剂施用于受试者。多肽的合适剂量(“治疗有效量”)将取决于,例如,待治疗的病状、病状的严重程度和病程、是否出于预防或治疗目的而施用该多肽、既往治疗、患者的临床病史和对多肽的应答、所用多肽的类型以及主治医师的判断力。该多肽适合一次或经过一系列治疗施用于患者,并且可以从诊断开始的任何时间施用于患者。该多肽可以单独治疗或与可用于治疗所述病症的其他药物或疗法联合施用。如本文所用,术语“治疗”是指治疗性治疗和预防性或预防性(prophylactic或preventative)措施。需要治疗的那些包括已经患有病症的那些以及待预防病症的那些。如本文所用,“病症”是将从治疗获益的任何病状,包括但不限于慢性和急性病症或疾病,包括那些使受试者易于患所述病症的病理性病状。在药理学意义上,在本公开的上下文中,多肽(例如,抗体)的“治疗有效量”是指在预防或治疗抗体对其治疗有效的病症的中有效的量。在一些实施例中,施用的多肽的治疗有效量将在约0.1mg/kg至约50mg/kg(诸如约0.3mg/kg至约20mg/kg或约0.3mg/kg至约15mg/kg)患者体重的范围内,通过一次或多次施用。在一些实施例中,治疗有效量的多肽以日剂量或多次日剂量施用。在一些实施例中,治疗有效量的多肽的施用频率少于每日一次,诸如每周或每月一次。例如,多肽可以以约100mg至约400mg(诸如约100、150、200、250、300、350或400mg中任一值,包括这些值之间的任何范围)的剂量每一周或多周(诸如每1、2、3或4周或更多周,或每1、2、3、4、5或6个月或更多个月)施用,或者以约1.0、1.5、2.0、2.5、3.0、3.5、4.0、4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0、8.5、9.0、9.5、10.0、15.0或20.0mg/kg的剂量每一周或多周(诸如每1、2、3或4周或更多周,或每1、2、3、4、5或6个月或更多个月)施用。该剂量可以以单次剂量或以多次剂量(例如,2、3、4或更多次剂量)的形式施用,诸如输注。疗法的进展可以通过常规技术容易地监测。vi.制品和试剂盒本公开的某些方面涉及包含容纳本公开的任何液体制剂的容器的制品或试剂盒。合适的容器包括,例如,瓶、小瓶和注射器。容器可以由多种材料形成诸如玻璃或塑料。示例性的容器为2-20cc的一次性玻璃小瓶。可替代地,对于多次剂量制剂,容器可以为2-100cc玻璃小瓶。容器容纳制剂,并且容器上或与容器相关的标签可以指示使用说明。制品可以进一步包括从商业和用户角度出发期望的其他材料,包括其他缓冲剂、稀释剂、过滤器、针头、注射器和带有使用说明的包装插页。在一些实施例中,制品或试剂盒进一步包括包装插页,该包装插页包括液体制剂的使用说明。包装插页可以指治疗产品的商业包装中通常包括的说明书,其含有涉及此类治疗产品的使用的有关适应症、用法、剂量、施用、禁忌症和/或警告的信息。还提供了可用于多种目的的试剂盒,例如,用于减少液体制剂中多肽的氧化,或用于筛选液体制剂以减少多肽的氧化。本公开的试剂盒中提供的说明通常是在标签或包装插页(例如,试剂盒中包括的纸张)上的书面说明,但是机器可读的说明(例如,在磁性或光学存储盘上携带的说明)也可以接受。该说明书被认为足以使本领域技术人员能够实施本公开。除了本文中示出和描述的之外,本公开的各种修改对于根据说明书前文的本领域技术人员而言将变得显而易见,并且落入所附权利要求的范围内。实例参考以下实例将更全面地理解本公开。然而,它们不应被解释为限制本公开的范围。应当理解,本文描述的实例和实施例仅用于说明目的,并且鉴于其的各种修改或改变将被建议给本领域技术人员,并且将被包括在本申请的精神和界限内以及本发明所附权利要求的范围内。实例1:nat保护免受氧化的评估。进行以下研究以评估作为生物治疗药物制剂组分的n-乙酰基-dl-色氨酸(nat)和/或l-蛋氨酸的抗氧化有效性和安全性。将2,2′-偶氮-双(2-脒基丙烷)二盐酸盐(aaph),一种偶氮化合物,可产生能够氧化蛋氨酸和色氨酸残基的活性氧(ji等人(2009)j.pharm.sci.98(12):4485-4500),以及光暴露选作以下研究的氧化模型,因为其代表了在制造和/或长期储存过程中抗体可能会暴露的常见氧化途径(grewal等人(2014)mol.pharm.11(4):1259-1272)。材料与方法材料mab1和mab2是具有氧化敏感色氨酸和蛋氨酸残基的igg1单克隆抗体(dion等人,文章待投)。除非另有说明,否则通过一系列层析步骤(包括蛋白a亲和层析和离子交换层析)纯化mab,并在ph值为5.5的低离子强度乙酸钠缓冲剂中配制表面活性剂或其他赋形剂。l-蛋氨酸和n-乙酰基-dl-色氨酸(nat)购自ajinomotonorthamerica(raleigh,nc)。2,2′-偶氮-双(2-脒基丙烷)二盐酸盐(aaph)购自calbiochem(lajolla,ca)。胰蛋白酶(质谱级)购自promega(madison,wi)。高压液相色谱(hplc)级乙腈和水购自fisherscientific(fairlawn,nj)。用于缓冲剂制备的水获自milli-q纯化系统(millipore,bedford,ma)。评估nat抗氧化剂有效性鉴别和监测氧化敏感的残基抗体经受aaph应激,然后进行肽作图分析,以鉴别对氧化敏感的cdr和fc残基(dion等人,文章待投)。kabat编号用于鉴别可变片段(fv)残基,而eu命名法(edelman等人(1969)procnatlacadsciusa63(1):78-85)用于鉴别fc残基。如果残基相对于对照的氧化程度>5%,则在整个实验过程中都将其视为敏感并进行监测。肽图和分析信息如dion等人所报道(文章待投)。简而言之,将样品变性、还原、羧甲基化并进行胰蛋白酶消化。在配有thermoqexactiveplus高分辨率质谱仪的watersacquityh-classuhplc上使用水/乙腈/甲酸梯度,在acquityuplcpeptidecshc18柱上分离肽。使用thermoscientificpepfindertm和xcaliburtm软件处理数据。使用天然和氧化肽的最丰富电荷态,对单同位素m/z的提取离子色谱图进行积分。通过将氧化的肽的峰面积除以天然和氧化的肽的总峰面积来计算氧化百分比。将主要的色氨酸降解产物(对于高氧化位点,除了+4、+20和+48之外,还包括+16和+32)汇总并用于计算色氨酸氧化。仅使用蛋氨酸亚砜(m+16)计算蛋氨酸氧化,因为在这些条件下未观察到蛋氨酸砜(m+32)。如果两个软件包提供不同的答案,则在手动检查数据后报告xcaliburtm数据。aaph化学氧化应激模型在2cc玻璃小瓶中,在20mm乙酸钠,ph值为5.5中将抗体终浓度制备为1mg/ml。在20mm乙酸钠(ph5.5)中从3mmnat的储备溶液,添加nat达到终浓度为0.05mm和0.3mm。对于特定样品,在20mm乙酸钠(ph5.5)中从50mm储备溶液,添加l-蛋氨酸达到终浓度为5mm。从11mm储备溶液,添加aaph达到终浓度为1mm。将等体积的水添加到蛋白等分试样中,以代替aaph作为对照样品。加入aaph或水后,将样品在40℃孵育16h。对照样品也立即在-70℃冷冻。用l-蛋氨酸与aaph的比例为20∶1的l-蛋氨酸淬灭进行产生自由基的反应,然后将样品各自用缓冲剂交换成制剂缓冲剂(20mm乙酸钠,100mm蔗糖,ph5.5),使用pd-10色谱柱(gehealthcare),并使用amicon超离心过滤器(emdmillipore)浓缩达到终浓度为10mg/ml,以准备通过lc-ms肽图分析。光暴露应激模型通过将玻璃瓶中的10mg/ml样品在atlassuntestcps+氙灯箱(chicago,il)中暴露于总剂量为300千勒-小时的可见光和50w·h/m2的近uv(320-400nm)光下进行光稳定性研究。从上述储备溶液添加nat达到终浓度为0.05、0.1、0.3、0.5或1.0mm。将对照样品包裹在铝箔中,并与实验小瓶并排放置。暴露后,将样品储存在-70℃,以备通过lc-ms肽图分析。nat和l-蛋氨酸的安全性评估计算机模拟致突变性和致癌性预测使用计算机模拟建模工具中的dereknexus(程序版本2.0.2.201111291322;lhasalimited,leeds,uk)和(modelapplier版本1.5.0;leadscopeinc.,columbus,oh),评估nat的致突变性和致癌性。体外受体结合和功能评估在结合、细胞和核受体功能以及组织生物测定中评估nat的活性。在内源性表达受体的u373mg人类星形细胞瘤细胞中评估与神经激肽-1(nk-1)受体的结合(eistetter等人(1992)functionalcharacterizationofneurokinin-1receptorsonhumanu373mgastrocytomacells.glia6(2):89-95;heuillet等人(1993)j.neurochem60(3):868-876),并与参比激动剂[sar9,met(o2)11]-sp或参比拮抗剂l733,060进行比较。将nat或参比化合物与u373mg细胞在室温下孵育;一式两份地测定所有浓度。已经显示通过nk-1受体发挥作用的p物质可调节人类和非临床物种的血管张力(coge和regoli,(1994)neuropeptides26(6);385-390;shirahase等人(2000)br.j.pharmacol129(5);937-942)。为了评估nat对nk-1受体的潜在比活力,将具有完整内皮的兔肺动脉环悬挂在充满氧(95%o2/5%co2)的20ml器官浴和预热的(37℃)生理盐溶液(以mm计)中:nacl118.0,kcl4.7,mgso41.2,cacl22.5,kh2po41.2,nahco325和葡萄糖11.0(ph7.4)。在整个实验过程中,存在普萘洛尔(1μm)、美吡拉敏(1μm)、阿托品(1μm)和美西麦角(1μm)分别阻断β-肾上腺素、组胺h1、毒蕈碱和5-ht2受体。将组织连接至用于等距张力记录的力传感器,拉伸至静息张力为2g,然后将其平衡60分钟,在此期间,将它们反复洗涤并重新调节张力。使用具有八个器官浴的半自动化隔离器官系统进行实验,并进行多通道数据采集。测得的参数是化合物各自浓度引起的最大张力变化。为了评估激动剂活性,将组织用去甲肾上腺素(0.1μm)收缩,暴露于低于最大浓度的参比激动剂[sar9,met(o2)11]-sp(0.001μm)中,以验证应答性并获得对照松弛,然后洗涤。此后,组织每隔45分钟用去甲肾上腺素收缩,暴露于浓度增加的nat或参比激动剂中,然后洗涤。将每种化合物浓度均与组织接触,直到获得稳定应答或最多15分钟。如果获得激动剂样应答(松弛),则在30分钟前添加参比拮抗剂spantideii(1μm)的情况下,再次测试化合物的最高浓度,以确认nk1受体参与了该应答。为了评估拮抗剂的活性,将组织用去甲肾上腺素(0.1μm)收缩,暴露于低于最大浓度的参比激动剂[sar9,met(o2)11]-sp(0.001μm)中,以获得对照松弛,然后洗涤。在增加浓度的nat或参比拮抗剂spantideii的存在下,每45分钟重复一次此顺序,每次添加在暴露于[sar9,met(o2)11]-sp之前的30分钟。nat/l-蛋氨酸制剂的体内耐受性在动物中开展的所有程序均符合动物福利法、实验动物的护理和使用指南以及实验动物福利办公室的规定。方案已由适用的机构动物护理和使用委员会批准。兔玻璃体内单次剂量耐受性研究为了评估支持旨在治疗视网膜病症的产品的急性耐受性,对雄性新西兰白(nzw)兔施用等渗赋形制剂(n=2)或含有5mmnat的赋形制剂双侧玻璃体内注射(50ul/眼)和25mml-蛋氨酸(n=3)单次剂量。在研究第1天施用赋形溶液于动物。毒性评估基于临床观察、眼内压(iop)测量和眼科检查。在第8天的尸检中,收集眼睛和视神经并进行苏木素和伊红(h&e)染色处理,并由美国兽医病理学家学院(acvp认证的兽医病理学家)进行显微镜分析。兔玻璃体内重复剂量毒理学研究在雄性和雌性nzw兔中进行了一项优良实验室规范(glp)的毒理学研究,以支持旨在治疗视网膜病症的产品。每隔一周(第1天、第15天、第29天和第43天)通过双侧玻璃体内注射(50ul/眼)施用赋形制剂(等渗溶液,含1mmnat,5mml-蛋氨酸,ph5.5)于动物(n=5/性别)。毒性评估基于临床观察、体重测量、眼科检查、眼压测量、眼部摄影和临床病理。在第45天的尸检时,收集并处理全套组织用于h&e染色,并由acvp认证的兽医病理学家进行显微镜分析。食蟹猴重复剂量毒理学研究-玻璃体内施用在雄性和雌性食蟹猴(macacafascicularis)中开展glp毒理学研究,以支持旨在治疗视网膜病症的产品。每隔一周(第1天、第15天、第29天、第43天、第57天和第71天)为期十周以上,通过双侧玻璃体内注射(50ul/眼)施用赋形制剂(等渗溶液,含1mmnat,5mml-蛋氨酸,ph5.5)于动物(n=5/性别)。毒性评估基于临床观察、身体检查、心电图、眼科检查、频域光学计算机断层扫描(oct)、眼部摄影、荧光素血管造影、视网膜电图和临床病理。在第72天或第99天的尸检时,收集并处理全套组织用于h&e染色,并由acvp认证的兽医病理学家进行显微镜分析。食蟹猴重复剂量研究-皮下施用在雄性和雌性食蟹猴(macacafascicularis)中开展glp毒理学研究,以支持旨在治疗代谢疾病的产品。每周一次(第1天、第8天、第15天、第22天和第29天)为期4周以上,通过皮下注射(0.1ml/kg)施用赋形制剂(等渗溶液,含0.3mmnat,5mml-蛋氨酸,ph5.8)于动物(n=8/性别)。毒性评估基于临床观察、身体检查、神经科和眼科检查、临床病理和尿液分析。在第32天或第99天的尸检时,收集并处理全套组织用于h&e染色,并由acvp认证的兽医病理学家进行显微镜分析。.结果aaph自由基化学氧化应激开展aaph应激测试以测定暴露于溶液中自由基的nat对敏感色氨酸和蛋氨酸残基的抗氧化性能。如先前报道(dion等人,文章待投),mab1的肽图分析显示两个敏感的cdr色氨酸残基w52a和w100b以及fv蛋氨酸hcm82。对于mab2,鉴别出两个肽,肽各自包含多个敏感残基(cdrh1w33/m34/w36和cdrh3w99/w100a和fvw103)。对于具有多个敏感残基的这两种肽,本文显示了肽各自的总氧化值。还发现与fcrn受体相互作用的fc蛋氨酸残基252和428对两个分子的氧化应激均敏感,这与既往文献一致(bertolotti-ciarlet等人,2009)。为了测定nat浓度对抗氧化剂效能的影响,制剂中的nat浓度在0mm至0.3mm之间变化,并且将配制的mab经受aaph应激(图1)。在没有nat的情况下,具有敏感色氨酸残基的fv肽的氧化水平在aaph应激下分别增加11%(mab1的w100b)、60%(mab1的w52a)和87%(mab2的w99/w100a/w103)。这些初始起始值提供了广泛的氧化敏感性,可用于研究nat的影响。稳定色氨酸残基所需的最小nat浓度与残基的初始aaph敏感性相关(图1a)。添加0.05mmnat可以将mab1w100b的氧化降低至5%,而mab1w52a需要添加0.3mmnat才能将氧化降低至5%。相反,分别添加0.05mm、0.1mm和0.3mmnat,mab2的w99/w100a/w103的氧化仅减少至77%、62%和8%。同样,随nat浓度的增加,在mab2上含w33/m34/w36的肽通常会下降,尽管不能明确测定对该肽中单个色氨酸和蛋氨酸残基的相对影响。在不存在nat(3%)的情况下,mab1中敏感较低的m82残基被最小程度地氧化,并且包含nat表现出很小的影响(在0.3mmnat下轻微降低至1%氧化)。还评估了nat浓度对fc蛋氨酸氧化的影响(图1b)。在没有nat的情况下,在暴露于aaph后,两种mab的m252和m428的氧化水平在11%至16%之间。与cdr残基在很大程度上不受nat氧化的保护相反,fc蛋氨酸残基的氧化由于添加nat而加剧。在最高测试水平(0.3mmnat)下,相对于没有nat的相应条件,fc蛋氨酸残基的氧化增加6%-12%。因为nat保护cdr和fv色氨酸残基免受氧化(在0.3mmnat下,氧化<10%氧化)(图1a),但加剧了fc蛋氨酸残基的氧化(图1b),所以包含l-蛋氨酸和nat共配制的实验组被包括在抗氧化剂效能研究中。单独的l-蛋氨酸(5mm)对aaph敏感的色氨酸残基具有混合作用,显示出mab1轻微改善,而且mab2的氧化水平没有影响或轻微加剧(图2a)。在单独添加l-蛋氨酸的情况下,两个分子的fc蛋氨酸的氧化水平降低至2%或更低(图2b)。0.3mmnat和5mml-蛋氨酸的组合有效地降低aaph诱导的cdr色氨酸残基氧化至<5%和fc蛋氨酸残基至<2%,这使得抗氧化剂赋形剂的组合是控制在测试条件下氧化水平最有效的方法(图2a-b)。光暴露应激:高强度uv蛋白(10mg/ml)在不同的nat浓度(0-1mmnat)下暴露于高强度uv组件(300千勒-小时可见光和50w·h/m2的近uv(320-400nm)光超过6小时)的光应激,以测定nat作为抗光氧化作用的抗氧化剂的效能(图3a-b)。与aaph研究相比,基于nat具有光敏性的报道,其nat浓度范围更广(chin等人(2008)j.am.chem.soc.130(22):6912-6913)。在测试条件下,mab1和mab2中的大多数cdr和fv残基的氧化水平≤1%。仅两种肽表现出对所检测的光照条件的敏感性(mab1w52a(3%)和mab2的w99/w100a/w103(6%))(图3a)。添加≥0.1mmnat对在这些位点的氧化的影响最小(mab1w52a变化<1%,mab2w99/w100a/w103增加1-2%)。在测试条件下将nat添加到制剂中时,测定为在无抗氧化剂条件下对光氧化不敏感的残基仍对光不敏感。与fv残基相反,fc蛋氨酸残基对uv光应激和nat的添加敏感(图3b)。例如,在mab1中存在1mmnat的情况下,fc蛋氨酸残基m252的氧化从无nat的8%增加到19%,而对于mab2,其氧化从16%至31%。这些结果表明,与在aaph应激下一样,在uv光应激条件下,nat增加了fc蛋氨酸残基的氧化水平。为了测定是否可以通过添加l-蛋氨酸来降低nat对fc蛋氨酸的敏化作用,评估nat和l-蛋氨酸在uv光条件下的组合作用。单独或与0.3mmnat组合添加5mml-蛋氨酸对在该氧化模型中的cdr和fv残基没有有益影响(图4a)。uv光诱导的fc蛋氨酸的氧化被5mml-蛋氨酸改善(图4b),但效果不如aaph模型中的明显。在这种强uv光氧化模型中,相对于无赋形剂条件或单独使用l-蛋氨酸,l-蛋氨酸(5mm)和nat(0.3mm)组合导致cdr色氨酸或fc蛋氨酸的轻微保护免受光氧化。nat和l-蛋氨酸的安全性评估鉴于nat和蛋氨酸存在于fda肠胃外制剂非活性成分列表中,并已在急性环境中安全使用而未识别危害,因此进行了简短的安全风险评估,以支持将其用于皮下或玻璃体内施用的制剂中。对两种新施用途径均进行了nat和l-蛋氨酸组合的体内耐受性研究。另外,正如文献报道所示,nat可能为nk-1受体的拮抗剂,因此对nat进行了计算机模拟毒性评估和nk-1受体结合的体外评估。nat的计算机模拟评估derek是基于经验/规则的系统,通过将测试化合物(即nat)的结构特征与其数据库中被认为负责毒理作用的部分分子(毒体)进行比较,得出预测。nat的结构已提交至dereknexus数据库,该数据库返回“无报告”结果。是一种定量构效关系(qsar)系统,其中包括用于预测遗传毒性的预训练模型;该系统是与美国fda合作创建的,且具有高敏感性和负预测性(sutter等人(2013)reg.tox.pharm.67(1):39-52)。在总共40个模型中评估了阳性结果的可能性。其中只有2个模型被预测为阳性,其余38个模型被预测为阴性(即无毒性预测)。在遗传毒性类别中,“其他细胞中的姐妹染色单体交换(sce)”模型为阳性,阳性预测概率为0.829。相反,其他两个sce模型(体外sce和cho体外sce)均为阴性。在啮齿动物的致癌性类别中,“carc小鼠雄性”模型被预测为阳性,阳性预测概率为0.622。工具中的0.4-0.6之间的预测概率被视为临界预测。该模型的第二次运行返回了阴性预测,而对小鼠致癌性(雄性和雌性的组合)的总体预测为阴性。体外受体结合和功能评估参比化合物(sar9,met(o2)11)-sp和l733,060)与nk-1受体的激动剂和拮抗剂结合的ic50分别为4.2-10m和4.7-10m。相反,无法计算出nat与nk-1受体的激动剂或拮抗剂结合的ic50值,表明在测定所采用的条件下nat活性不足。体内耐受性评估-兔和食蟹猴毒理学研究通过长达6周的单次和重复剂量施用,对兔通过玻璃体内施用含高达5mmnat和25mml-蛋氨酸的赋形制剂具有良好的耐受性。通过每两周一次长达7周的玻璃体内施用,以及类似地通过每周长达4周的皮下施用,对食蟹猴施用含有0.3mmnat和1mml-蛋氨酸的赋形制剂具有良好的耐受性。在任一物种中均未发现与赋形相关的临床观察或体重、身体检查、神经科或眼科检查、眼内压、oct、眼部摄影、荧光素血管造影、视网膜电图、血液学、凝血和临床化学参数、尿液分析或肉眼或显微镜病理的变化。综上所述,本文提供的研究表明,尽管nat可有效保护cdr色氨酸残基免受aaph降解产生的ros的影响,但其可能具有对化学和光诱导的氧化敏感的fc蛋氨酸残基。在nat中添加l-蛋氨酸可有效保护色氨酸和蛋氨酸残基免受化学诱导的氧化,并导致光氧化水平等于或低于测试条件下不含抗氧化剂制剂中的水平。这些研究表明,nat和l-蛋氨酸的组合能够提供针对生物疗法制造和储存过程中常见的氧化应激类型的保护。重要的是,安全性评估证实两种赋形剂均具有良好的耐受性。因此,本文提供的证据表明,nat和l-蛋氨酸作为生物治疗制剂中的抗氧化剂赋形剂可能是安全有效的,这为色氨酸和/或蛋氨酸在制剂开发中的氧化处理提供了重要的新选择。实例2.抗氧化剂在aaph应激测试中可减少氧化双特异性抗体mab3抗体用于评估nat+蛋氨酸的抗氧化能力。将1mg/ml的mab3与1mm的aaph在40℃下混合16小时,无论有无1mmnat和5mm蛋氨酸。然后如上所述通过质谱法测量mab3的氧化,并通过elisa测量其效价。结果如表1所示。表1.mab3的氧化样品12缓冲剂和ph值his-acph5.5his-acph5.5n-乙酰基-色氨酸-1mml-蛋氨酸-5mm%wwox96.612.3与ag3结合(相对效价%)受影响的89向溶液中添加nat+蛋氨酸可极大减少mab3的氧化。实例3.抗氧化剂的添加减轻化学氧化风险mab4(igg1抗体)在20mm组氨酸hcl、50mm氯化钠、200mm蔗糖、0.04%泊洛沙姆188中配制为100mg/ml。然后将抗体制剂在0mm、5mm、10mm或10mmaaph+1mmnat+5mm蛋氨酸的aaph存在下于40℃孵育24小时。然后如上所述通过ms评估样品。结果如图5所述。在5mmaaph下观察到fcm272约15%氧化。这对应于10%色氨酸氧化。添加1mmnat+5mm蛋氨酸可将色氨酸的氧化减少约50%,将fc蛋氨酸272的氧化减少约80%。在任何水平上均未观察到蛋氨酸cdr的变化。通过elisa测量,添加nat+蛋氨酸改善了mab4与ag4结合的比活力降低。表2.抗体效价aaph比活力%0106563104310+1mmnat/5mm蛋氨酸88实例4.添加nat/蛋氨酸以减轻光氧化风险开展研究以测定nat/蛋氨酸是否可以减少光氧化。将具有与igg1不同同种型的igg抗体mab5在200mm精氨酸琥珀酸酯,ph值为5.5,以150mg/ml的浓度不含nat的条件下配制以及与0.3mmnat+5mm蛋氨酸或0.3mmnat+10mm蛋氨酸配制在一起。样品暴露于300,000lux小时以评估风险。结果在表3中总结。表3.环境光应激nat/蛋氨酸保护mab5免受与环境光有关的氧化。实例5.nat/蛋氨酸的添加提供氧化和效价保护抗体药物产品在保质期结束时可能显示met251约7%-8%氧化,通常在5℃下超过2年。为了模拟这一点,将抗体用5mmaaph处理,产生约15%的met251氧化。用有或没有nat/蛋氨酸的5mmaaph处理样品,然后分析w104和m251的氧化。还测量了抗体的效价。如表4所示,在抗体库中添加0.3mmnat+5mm蛋氨酸可减少w104和m251的氧化,并减少aaph应激后抗体效价的降低。表4.nat/蛋氨酸提供氧化和效价保护另外,如上所述,克隆库经受环境光应激。如表5所示,克隆库经历与上述相同的颜色变化。表5.抗体的光保护样品治疗hmws颜色0mmnat/蛋氨酸箔控制0.71%b5.30mmnat/蛋氨酸环境光0.92%by3.00.3mmnat/5mm蛋氨酸环境光0.74%b5.4实例6.nat/蛋氨酸减轻化学氧化风险aaph应激测试用于评估nat和/或蛋氨酸的氧化保护。mab6,一种双特异性抗体,在有或没有nat和/或蛋氨酸的情况下,在20mm乙酸组氨酸和1mmaaph中以1mg/ml的浓度在40℃下孵育16小时。如上所述分析样品的氧化。如表x所示,0.1mm至0.5mm浓度的nat可提供氧化保护,单独使用蛋氨酸也可提供一些对aaph的保护。表6.nat/蛋氨酸减轻化学氧化风险实例7.nat/蛋氨酸防止化学诱导的氧化。通过在20mm乙酸组氨酸中将1mg/ml的分子与1mmaaph在40℃下孵育16小时,在有或没有nat和蛋氨酸的情况下,评估双特异性抗体mab7的w52位置的化学诱导的氧化。通过肽图分析样品的氧化。如图6所示,nat+蛋氨酸的组合防止w52化学诱导的氧化。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1