4型蓝舌病病毒VP2蛋白中和性B细胞抗原表位多肽及其应用的制作方法
本发明涉及一种抗原表位多肽及其应用,特别涉及4型蓝舌病病毒vp2蛋白的中和性b细胞抗原表位多肽及其在诊断或治疗蓝舌病中的应用,本发明属于医药技术领域。
背景技术:
蓝舌病(bluetongue,bt)是一种在热带和亚热带国家常见,感染反刍动物的虫媒病毒性动物传染病。bt是由蓝舌病病毒(bluetonguevirus,btv)引起的,可感染羊、牛和骆驼等反刍动物的传染病。btv感染动物,植物,昆虫并且通过诸如蚊子、库蠓、蜱虫等节肢动物传播。btv属于呼肠孤病毒科(reoviridae)十二个属之一的环状病毒属(orbivirus)。btv最早发现于1876年,呈世界性分布。目前已从非洲、欧洲、亚洲、北美、南美和大洋洲的多个热带、亚热带和温带地区国家分离到btv,并且其分布范围不断扩大,危害日趋严重。我国已检测出btv-1、btv-2、btv-3、btv-4、btv-5、btv-7、btv-12、btv-15、btv-16、btv-24十种血清型,其中btv-1和btv-16为我国主要的致病血清型。
vp2蛋白是构成btv外壳的主要结构蛋白,大多中和表位都存在于vp2蛋白上,故其能够诱导机体产生对btv的中和反应。vp2蛋白由l2基因编码。有数据表明l2基因在各血清型间差异较大,l2基因的变异与btv血清型之间有密切联系,证明vp2蛋白为型特异性蛋白,可以用来区别btv各血清型间的不同,因此vp2蛋白血清型的主要决定因素。
btv呈现蔓延趋势,近些年不断出现新疫情。btv具有多型性,并且其不断变异,目前已经分离出27个血清型,不同血清型之间无交叉免疫力。同时,4型btv能感染绵羊、骆驼等反刍动物,其发病急,死亡率高,对养殖业造成的损害巨大。因此,建立型特异性鉴别诊断方法从而防控btv尤为重要。
技术实现要素:
本发明所要解决的技术问题是提供一种4型蓝舌病病毒vp2蛋白的中和性b细胞抗原表位多肽及其在检测蓝舌病病毒中的应用。
为了达到上述目的,本发明采用了以下技术手段:
本发明应用噬菌体随机肽库对自行制备成功的4a-1g7单抗进行3轮生物淘选,测序后结果为:12个噬菌体序列一致含有nh,这与4型btvvp2氨基酸序列160-161位氨基酸有一定的相似度。为了进一步对该序列进行鉴定,合成该短肽序列160nh161,采用间接elisa方法与24个血清型的btv阳性血清反应,结果发现该短肽160nh161可与大多数阳性血清呈现明显的阴性反应,与少数几种血清反应呈现阳性。并且158amnsy162,158amnhnsy162与btv-4标准阳性血清分别反应,含有160nh161片段的反应性更强。该结果表明,该抗原表位具有良好的特异性。此外,噬菌体竞争抑制试验与elisa方法证实筛选出的噬菌体多肽的空间构象和天然抗原表位相似,因此提示该短肽可用于4型btv相关表位疫苗的制备。
因此,在上述研究的基础上,本发明提出了一种4型蓝舌病病毒vp2蛋白的中和性b细胞抗原表位多肽,所述的抗原表位多肽含有seqidno.1所示的氨基酸序列。
在本发明中,优选的,所述的抗原表位多肽的氨基酸序列如seqidno.1或seqidno.2所示。
编码所述的中和性b细胞抗原表位多肽的核苷酸序列以及含有所述的核苷酸序列的表达载体也在本发明的保护范围之内。
进一步的,本发明还提出了所述的中和性b细胞抗原表位多肽在制备检测或诊断蓝舌病病毒感染试剂中的应用,以及所述的中和性b细胞抗原表位多肽在制备预防或治疗蓝舌病病毒药物中的用途。
其中,优选的,所述的蓝舌病病毒包括btv-4、btv-8、btv-9、btv-18、btv-20以及btv-9。
本发明的提出,对于4型btv的临床诊断提供了新的技术手段,也为后续建立btv的型特异性鉴别诊断方法以及4型btv相关表位疫苗的制备奠定了物质基础。
附图说明
图1为btv1-24的血清型中vp2的第161-162位序列;
图2为短肽与mab间接elisa鉴定结果;
图3为短肽与不同血清型btv阳性血清间接elisa鉴定结果;
图4为短肽158amnsy162、158amnhnsy162与btv-4阳性血清间接elisa鉴定结果。
具体实施方式
以下实施例进一步说明本发明的内容,但不应理解为对本发明的限制。在不背离本发明精神和实质的情况下,对本发明方法、步骤或条件所作的修改或替换,均属于本发明的范围。若未特别指明,实施例中所用的技术手段为本领域技术人员所熟知的常规手段。
实施例14型蓝舌病病毒vp2蛋白中和性b细胞抗原表位的鉴定
1材料与方法
1.1细胞与主要试剂
抗vp2蛋白mab1g7细胞株由本实验室保存。hrp标记的山羊抗鼠igg购自中杉金桥有限公司;1-24型btv标准阳性血清及阴性血清购自蓝舌病参考实验室pirbrightinstitute;ph.d.-12tm13噬菌体肽库试剂盒购自美国newenglandbiolabs(neb)公司。
1.2基因序列
用于比对的btv1-24型genbank登录号见表1。
表1btv1-24型的genbank登录号
1.3抗原表位筛选
参照ph.d.-12tm13噬菌体肽库说明书,用纯化后的mab腹水作为抗原,加入到微孔板中,对噬菌体肽库进行3次淘选,将三次淘选后的产物测序。
1.4单克隆抗体所识别抗原表位的位置定位及真实性分析
将20个通过序列测定的噬菌体上所展示的随机的12肽序列进行比对分析,确定共有氨基酸序列,并确定该共有序列在编码血清型4型btvvp2蛋白的氨基酸序列上的位置,该位置区域即为单克隆抗体所识别的抗原表位所在区域。将获得的表位序列由上海耀光生物科技股份有限公司进行合成短肽。
同时,为了证明单克隆抗体所识别的抗原表位的真实性,用包被稀释液将短肽倍比稀释,以100μl/孔包被酶标板,分别以4a-1g7上清及纯化后腹水稀释液为抗体,hrp标记的山羊抗鼠igg作为二抗,加入现配制的opd-h2o2底物显色液,显色15min后加入终止液,鉴定短肽反应活性。并且利用上海耀光生物科技股份有限公司合成去除160nh1611的158amnsy162与原btv-4片段158amnhnsy162。分别以158amnsy162、158amnhnsy162为包被抗原,4型btv标准阳性血清及阴性血清为抗体进行间接elisa验证,以证明单克隆抗体所识别的抗原表位的真实性。
1.5单克隆抗体所识别抗原表位的保守性分析及特异性分析
利用ncbi数据库中的blast软件对mab4a-1g7识别的抗原表位序列160nh161进行保守性分析和特异性分析。并以短肽为包被抗原,1-24型btv标准阳性血清及阴性血清为抗体,进行间接elisa验证,以证明单克隆抗体所识别的抗原表位的特异性。
2结果
2.1单克隆抗体所识别抗原表位的dna序列测定及表位分析
利用噬菌体展示技术对所获得的4a-1g7单克隆抗体的表位进行了序列进行比对显示,通过噬菌体基因组测序显示:单克隆抗体4a-1g7所淘选出的20个噬菌体克隆中6个展示出了nh序列,该序列位于本研究所用4btvvp2蛋白的第161-162位的氨基酸位置(表2)。经过序列比对分析该序列在btv1-24的血清型中vp2的第161-162位氨基酸,不保守,特异性较强(图1)。所以推测160nh161(seqidno.1所示)序列可能是一个线性b细胞表位。
表2短肽序列测定结果
2.2间接elisa法鉴定短肽与4a-1g7的反应性
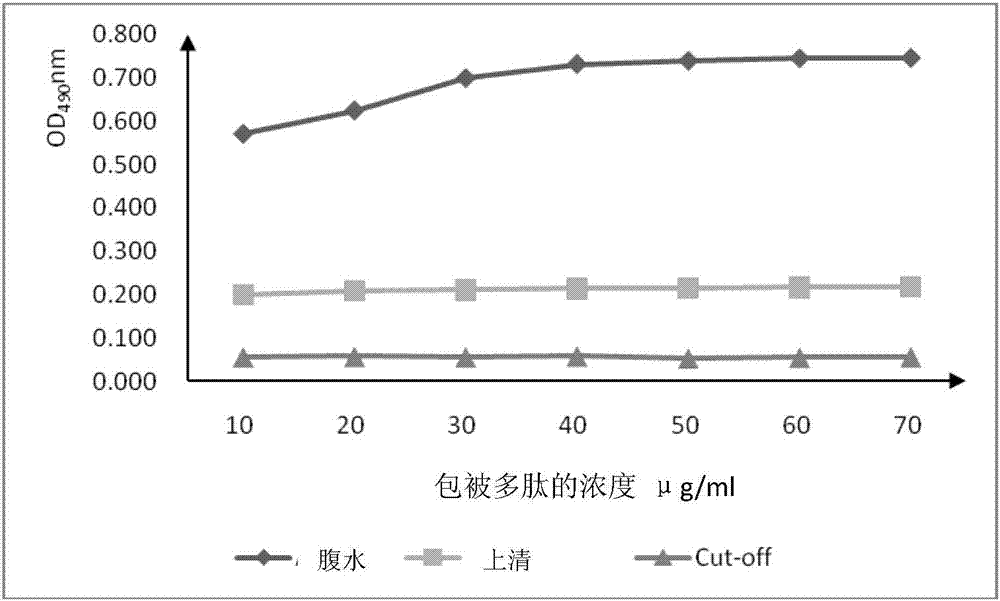
为了证明噬菌体所展示的单克隆抗体所识别构象抗原表位的真实性,针对所识别的氨基酸区域,本着疏水性氨基酸向亲水性氨基酸突变、亲水型不带电荷氨基酸与亲水型带正负电荷的氨基酸相互突变的原则,本发明发明人经人工合成了一条仅含有nh序列的短肽(seqidno.1所示),利用间接elisa法检测短肽序列是否与4a-1g7单抗上清和腹水发生反应。结果如图2,单抗上清及腹水均与短肽发生特异性的反应,短肽可以与单抗上清及腹水中的抗体发生特异性结合。并且与腹水反应的值明显高于上清,这可能是由于腹水中与短肽结合的非特异性抗体较多。
2.3间接elisa法鉴定短肽的特异性
间接elisa法检测合成的短肽与1-24型btv阳性血清的反应,结果如图3,结果表明:可见短肽与btv-4、btv-8、btv-9、btv-18、btv-20阳性血清的反应呈阳性反应,短肽与btv-9阳性血清的反应呈弱阳性。
间接elisa法检测合成敲除160nh161的btv-4片段158amnsy162与原btv-4片段158amnhnsy162(seqidno.2所示),结果如图4。等量158amnsy162,158amnhnsy162与btv-4标准阳性血清反应,短肽amnhnsy的反应性更强。
虽然,上文中已经用一般性说明及具体实施方案对本发明作了详尽的描述,但在本发明基础上,可以对之作一些修改或改进,这对本领域技术人员而言是显而易见的。因此,在不偏离本发明精神的基础上所做的这些修改或改进,均属于本发明要求保护的范围。
序列表
<110>东北农业大学
<120>4型蓝舌病病毒vp2蛋白中和性b细胞抗原表位多肽及其应用
<130>klpi170141
<160>2
<170>patentin3.5
<210>1
<211>2
<212>prt
<213>短肽
<400>1
asnhis
<210>2
<211>7
<212>prt
<213>短肽
<400>2
alametasnhisasnsertyr
- 还没有人留言评论。精彩留言会获得点赞!