Cry1Ab13‑1杀虫基因及应用的制作方法
本发明属于生物技术领域,具体涉及Cry1Ab13-1杀虫基因及其应用。
背景技术:
虫害是影响我国农作物产量的重要因素之一,据国家统计局统计2016年由于虫害的影响我国夏粮总产量较上年减产162.1万吨,带来严重的经济损失。玉米螟(Ostrinia nubilalis),大豆食心虫(Leguminivora glycinivorella Matsumura)等鳞翅目昆虫是主要为害的害虫,农作物不仅仅在生长发育的阶段会受到这些虫害的影响,在后期的运输和储存中也有它们的入侵, 目前为有效的防治这些害虫,使用了大量的化学农药据统计中国农药的单位面积使用量是世界平均水平的2.5倍 [4] ,然而大量广泛的喷洒化学农药严重的影响了人畜和生态环境的安全,也导致害虫产生抗药性,同时也杀死了天敌及有益昆虫,污染进一步加重。因此利用基因工程技术培育具有抗虫性状的品种才是从根本上解决虫害问题的有效途径之一。
苏云金芽孢杆菌(Bacillus thuringiensis,简称Bt)是由日本学者石渡在1901年从染病的家蚕中首次分离得到,1915年被正式定名为Bacillus thuringiensis(Bt ) [5],是种革兰氏阳性土壤芽孢杆菌,在形成芽孢的同时会产生杀死昆虫幼虫的毒性晶体蛋白[6]。1995年Crickmor等根据氨基酸序列的同源性,以45%、75%和95%为界限,对现有的Bt蛋白进行系统的分类和命名。由于Bt杀虫晶体蛋白具有高度的专一性,对害虫天敌及有益昆虫无毒害作用。只对鳞翅目、鞘翅目等靶标害虫具有杀虫效果 [7,8]、又不会对环境造成污染,因而被广泛的应用于害虫的防治。早在1987年Veack[9]就首次报道将Cry1Ab基因转化到烟草中,获得了抗烟草天蛾的转 Bt基因的烟草植株,凸显了Bt基因在基因工程中应用的潜力。郭三堆等[10]通过分子设计技术,人工合成了Cry1A基因并将该基因转入棉花中并大面积推广使我国成为第二个成功培育抗虫棉的国家。但是随着Bt基因及其制剂大量的应用,害虫对其产生了不同程度的抗性[11,12],而且野生型Bt基因mRNA的不稳定也造成了该基因的表达量极低[13]。于是,人们开始运用酶工程技术对野生型的Bt基因进行改造。王阳等[14]利用结构域交换法对杀虫晶体蛋白Cry1Ab进行优化改造,获得新型的杀虫基因CryFLIa,通过室内的生测证明该基因对亚洲玉米螟具有明显的抗性。截至目前cry类Bt基因发掘并注册共有318个[15]。现在有关强毒性和新型杀虫活性的cry类、cyt类、vip类Bt基因的发掘与克隆,仍是国内外对Bt基因研究和利用的热点[16]。
目前Bt蛋白改造主要的方法有结构域转换、密码子优化、DNA Shuffling、易错PCR等,其中易错PCR技术可以简便有效的向已知DNA序列中引入突变[17],其体系最早由Leung[18]建立, Cadwell[19]以该体系进行易错PCR,诱变了编码四膜虫核酶的基因,明显提高了该酶的活性。Shelly Goomber等[20]通过易错PCR法体外进化芽孢杆菌脂肪酶使其在5℃时仍具有活性,增强了该酶的适冷性。
技术实现要素:
本发明的目的是为了解决化学农药污染环境的问题,而提供一种Cry1Ab13-1杀虫基因及应用。
突变基因Cry1Ab13-1,其碱基序列如序列表SEQ ID NO.1所示;
所述的基因为人工合成;
Cry1Ab13-1蛋白,它是序列表SEQ ID NO.1所示的基因表达的蛋白。
Cry1Ab13-1蛋白作为制备杀虫剂的应用;
所述的虫为小菜蛾或玉米螟。
突变基因Cry1Ab13-1在培育抗小菜蛾虫或玉米螟虫转基植物品种的应用。
本发明提供了Cry1Ab13-1杀虫基因,它是通过易错PCR法对Cry1Ab13基因碱基序列进行随机诱变获得的,通过对诱变后的序列分析结果表明:与原始序列同源性达到38.61%,Ser 10.1%、Thr8.0%、和Leu7.9%居多,同原始的序列相比突变后序列GC的含量提高了3%,通过使用在线过敏原数据库在线分析表明,该序列与已知过敏源同源性较低不存在致敏性,通过SDS-PAGE电泳分析结果表明,在79.5KDa左右出现一条明显的特异条带,说明在IPTG的诱导下Cry1Ab13-1 基因得到了正确表达。用诱导表达的蛋白进行抗虫(小菜蛾,玉米螟)试验,结果表明该表达蛋白具有很强的杀虫活性,使小菜蛾幼虫的死亡率达87.81%,使玉米螟幼虫的死亡率达80.00%,而且存活的害虫生长也严重受到抑制,与对照差异明显。初步证明本研究成功诱变一种改良型的Cry1Ab13基因,该基因具有很强的杀虫活性,可以为转基因抗虫育种的培育和工程菌株的构建提供候选基因。
附图说明
图1 不同Mg2+浓度电泳图;M: DL2000 DNA marker;H:无菌水;1-10:反应体系中Mg2+浓度一次为2、2.5、3、3.5、4、4.5、5、5.5、6、6.5mmol/L;
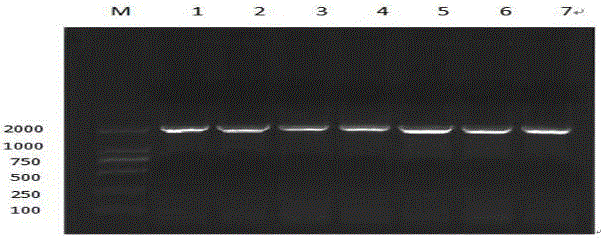
图2 易错PCR的电泳图;M:DL2000 DNA marker;1-7:Cry1Ab13的易错pcr扩增;
图3 Cry1Ab13-1保守结构域;
图4 重组质粒PCR验证;M:DL2000 DNA marker;1-7:重组质粒验证;
图5.CryAb13-1基因质粒双酶切鉴定, M:DL2000 DNA marker;1-2:酶切验证;
图6 pET28a-Cry1Ab13-1蛋白的SDS-PAGE分析,M:蛋白标准分子量;1:Cry1Ab13-1未诱导;2: cry1Ab13诱导;3: cry1Ab13-1诱导;
图7原核表达蛋白抗虫鉴定, A、B、C分别为24、48、72h。
具体实施方式
实施例1易错PCR扩增
使用软件Primer5.0设计引物
s:TGGACAACAATCCTAACATCAAC,
As: TTATCAAAGTTCATCCTTCTCGG (划横线部分为随机置换点) ,
以含有pUC57-Cry1Ab13质粒为模板,进行易错PCR,易错PCR 50ul反应体系包含DDH2O 17.8ul、Buffer 5ul、Mgcl2 20ul、dNTP 1ul、Primer1 2ul、primer2 2ul、模板 1 ul、Taq DNA聚合酶 1.2ul。反应程序:94℃预变性 3min;94℃变性 40s;40℃退火 40s,72℃延伸 3min;70×循环;72℃后延伸 10min; 4℃保存。不进行热启动。1%的琼脂糖凝胶电泳,纯化回收目的片段,以此片段为模板,以同样的条件进行第二轮易错PCR[22,23],经1%的琼脂糖凝胶电泳后,纯化并测序。使用软件DNAman6.0对测序结果进行比对分析,通过SWISS-MODEL(www.expasy.org)对测序结果同源建模并进行物理性质分析,根据外源蛋白质过敏性分析的国家标准,使用在线过敏原数据库(www.allergenonline.org)网站对其致敏性进行在线分析。
以不同镁离子浓度,使用低保真Taq DNA聚合酶,延长每个循环的时间,降低起始模板的终浓度,使用带有置换位点引物,不进行热启动等角度出发进行易错PCR扩增,经过初步的摸索,最终确定加5mmol/L的Mg2+(如图1),70个循环为适合的条件,进行易错PCR可以扩增得到清晰的特异条带(如图2)。
实施例2克隆载体的构建
将经过两轮易错PCR,纯化回收的目的片段与载体PMD-18T进行16℃,过夜连接。将连接产物转化到大肠杆菌E.coli DH5α感受态细胞中,次日挑取单菌落过夜培养,提取质粒进行PCR验证,纯化回收PCR产物送吉林省库美生物技术有限公司测序验证。命名为Cry1Ab13-1基因序列。
Cry1Ab13-1基因序列和保守结构域分析及功能预测
由DNA序列推导,其编码的蛋白质包含706个氨基酸其中亲水氨基酸占41.3%,疏水氨基酸占39.1%,酸性氨基酸占10.6%,碱性氨基酸占9%,理论分子量大小为79.5KDa,等电点5.19。在网站NCBI对该 基因编码蛋白质的氨基酸序列进行CDD(Convsed Domain Database)分析显示,Cry1Ab13-1编码蛋白的Domain I范围是Ser58~Asp265主要参与膜的穿透和孔洞的形成,Domain II对应的范围是Thr273~Glu485主要参与受体的结合,Domain III所对应的范围是Asn487~Pro632,与碳水化合物的结合有关,是δ内毒素活化区的一部分,可以产生杀虫毒素。如图(3)。其与原序列保守结构域相比,在结构域I中第130位的Ser变换成Pro,在结构域II中第383位的Thr变换成Ser。通过在线blast(NCBI)比较同源性显示,该基因与Cry1A最为相似,同源性达78%。在线致敏性分析结果显示,Cry1Ab13-1基因编码的蛋白质80个氨基酸序列与已知过敏原的序列同源性均小于35%,即CryAb13-1基因编码的蛋白不存在致敏性。
实施例3原核表达载体的构建与鉴定
提取pMD18T-Cry1Ab13-1的质粒DNA使用带有BamHI和SalI酶切位点的无缝引物进行正常的PCR扩增,扩增条件:94℃预变性 5min;94℃变性 40s;62℃退火 40s,72℃延伸 2min;72℃后延伸 10min;35×循环。PCR后产物进行1%琼脂糖凝胶电泳,纯化回收目的片段,并使用无缝连接试剂盒,将其与原核表达载体pET-28a进行连接,再转化大肠杆菌E.coli DH5α感受态细胞中,次日挑取全部单菌落过夜培养,并提取其质粒DNA,进行PCR和酶切验证,将初步确定为阳性的PCR产物送吉林省库美生物技术有限公司测序验证。
Cry1Ab13-1基因原核表达载体构建
挑取全部菌落过夜培养,次日提取质粒PCR验证(如图4),筛选出稳定的阳性克隆,对重组质粒DNA进行酶切验证,得到2个条带大小分别5.3kb和2033bp(如图5),测序分析表明序列正确,说明目的基因片段成功的连接到原核表达载体pET28a(+)中。
实施例4 Cry1Ab13-1基因在大肠杆菌中表达
提取pET28a-Cry1Ab13-1的质粒DNA,将其转化到大肠杆菌BL21(DE3)感受态细胞中,挑取单菌落过夜培养,次日将菌液按1:100的比列接种到装有50ml的LB液体培养基中(含Kan100mg/ml),37℃摇床198rpm振荡培养3.5h,使OD600达到0.6时,加入100mM的IPTG的母液至其终浓度1mM,继续振荡培养3h。将三角瓶置于冰上5min,离心收集菌体(4℃、10000r/min、8min),并用预冷的无菌水重悬菌体再次离心,重复两遍。弃上清用PBS Buffer重悬。将悬浮液用超声波破碎(1200W、6s on、5s off 10min),将其离心(4℃、10000r/min、10min)收集菌体和上清液备用。进行SDS-PAGE电泳分析验证。
1.2.4 Cry1Ab13-1基因在大肠杆菌中表达
提取pET28a-Cry1Ab13-1的质粒DNA,将其转化到大肠杆菌BL21(DE3)感受态细胞中,挑取单菌落过夜培养,次日将菌液按1:100的比列接种到装有50ml的LB液体培养基中(含Kan100mg/ml),37℃摇床198rpm振荡培养3.5h,使OD600达到0.6时,加入100mM的IPTG的母液至其终浓度1mM,继续振荡培养3h。将三角瓶置于冰上5min,离心收集菌体(4℃、10000r/min、8min),并用预冷的无菌水重悬菌体再次离心,重复两遍。弃上清用PBS Buffer重悬。将悬浮液用超声波破碎(1200W、6s on、5s off 10min),将其离心(4℃、10000r/min、10min)收集菌体和上清液备用。进行SDS-PAGE电泳分析验证。
Cry1Ab13-1基因在大肠杆菌中表达分析
SDS-PAGE电泳分析,结果显示(图6),经IPTG诱导后含CryAb13-1工程菌(非可溶性组分)在79.5KDa左右,表达出一条明显的特异条带,与预期大小相吻合,表明Cry1Ab13-1基因编码的杀虫蛋白在大肠杆菌BL21中得到正确的表达。
实施例5 Cry1Ab13-1蛋白杀虫活性的生物鉴定
重组菌菌体诱导表达后进行超声波破碎并回收Cry1Ab13-1蛋白,用于以下杀虫活性的鉴定。
小菜蛾杀虫活性的鉴定采用浸叶法:在实验室中选取甘蓝幼嫩一致的叶片,用清水洗净晾干,将其分割成大小相近的块状,将这些大小一致的甘蓝叶片放入待测样品中浸泡10min,晾干,之后平铺在玻璃培养皿中,每个培养皿中接2-3龄的小菜蛾幼虫12只,每个处理三次重复,将培养皿放入人工气候箱中进行培养[24](箱内的培养环境,温度为(25±1.5)℃、相对湿度50%-60%、光照周期14h,黑暗10h)。
玉米螟的杀虫活性鉴定采用人工饲料法:人工饲料的配制参照乔利的方法和条件。以含Cry1Ab13载体的破碎菌液做为阳性对照,以含空载体的破碎菌液为阴性对照,分别于24h、48h、72h调查结果用软件DPS9.5统计分析,三次重复。
Cry1Ab13-1蛋白杀虫活性的生物鉴定
小菜蛾杀虫实验结果表明:在24h之内三组小菜蛾都有大量的取食叶片,随着处理时间的延长实验组和阳性对照组的小菜蛾取食叶片量逐渐减少(叶片的孔洞减少),生长严重受阻,并萎缩出现死虫,而阴性对照组正常取食(如图7),24h、48h、72h实验组小菜蛾的死亡率分别为15.15%、33.31%、87.81%要高于阳性对照组(如表2)。不同的处理时间内,小菜蛾的死亡率差异达到极显著水平。载体之间阳性对照组和实验组的死亡率均与阴性对照组达到极显著的差异水平(如表1)。
玉米螟杀虫实验结果表明:玉米螟幼虫室内饲养24h后,处理组出现变软、发黑和死亡的幼虫,而阴性对照组的幼虫正常生长,随着处理时间的延长,处理组的幼虫逐渐停止活动,死虫的数目增加。24h、48h、72h实验组幼虫死亡率分别为11.11%、23.89%、80%(如表4)。不同处理时间各组的死亡率的差异达到极显著的水平,不同载体之间,实验组和阳性对照组幼虫的死亡率与阴性对照组的差异达到极显著的水平(如表3)。而存活的玉米螟生长也严重受到抑制,72h后实验组单只活虫的重量仅为0.18g,是处理前的7%,阴性对照组活虫重量为0.252g是处理前的98%倍(如表5)。
杀虫活性生物鉴定结果表明:新诱变的基因可以编码出具有杀虫活性的毒蛋白,并且对小菜蛾和玉米螟具有很强的毒杀作用。
本发明通过易错PCR法对Cry1Ab13基因碱基序列进行随机诱变研究,通过对诱变后的序列分析结果表明:与原始序列同源性达到38.61%,Ser 10.1%、Thr8.0%、和Leu7.9%居多,同原始的序列相比突变后序列GC的含量提高了3%,通过使用在线过敏原数据库在线分析表明,该序列与已知过敏源同源性较低不存在致敏性,通过SDS-PAGE电泳分析结果表明,在79.5KDa左右出现一条明显的特异条带,说明在IPTG的诱导下Cry1Ab13-1 基因得到了正确表达。用诱导表达的蛋白进行抗虫(小菜蛾,玉米螟)试验,结果表明该表达蛋白具有很强的杀虫活性,使小菜蛾幼虫的死亡率达87.81%,使玉米螟幼虫的死亡率达80.00%,而且存活的害虫生长也严重受到抑制,与对照差异明显。初步证明本研究成功诱变一种改良型的Cry1Ab13基因,该基因具有很强的杀虫活性,可以为转基因抗虫育种的培育和工程菌株的构建提供候选基因。
<110> 吉林农业大学
<120> Cry1Ab13-1杀虫基因及应用
<160> 1
<210> 1
<211> 2036
<212> DNA
<213> 人工
<400> 1
cttgcatgcc tgcaggtcga cgattatgga caacaatcct aacatcaacg aatgtatccc 60
atacaactgt ttgtcaaatc ctgaggttga ggtgcttggt ggtgagagga tcgagacagg 120
atacactcca atcgacatat cactttctct tacacagttt cttctttctg agttcgtgcc 180
tggtgctgga ttcgtgcttg gacttgtgga catcatctgg ggtatctttg gaccttcaca 240
gtgggacgca ttccttgtgc agatcgagca gttgatcaat cagagaatcg aagagttcgc 300
taggaaccag gctatctcaa ggcttgaagg actttctaac ttgtaccaga tctacgctga 360
atctttcagg gagtgggagg ctgactcaac taatcctgca cttagggaag aaatgagaat 420
acagttcaac gacatgaact ctgctcttac tactgctatc cctcttttcg ctgtgcaaaa 480
ctaccaagtg ccacttcttt cagtgtacgt gcaggcagct aaccttcacc tttctgtgct 540
tagggacgtg tcagtgttcg gtcagaggtg gggttttgac gctgcaacaa tcaattcaag 600
gtacaacgat cttactaggc ttatcggtaa ctatactgac cacgctgtga ggtggtacaa 660
cactggtttg gagagagtgt ggggtccaga ctcaagggac tggatcaggt acaaccagtt 720
cagaagggag ttgactttga ctgttcttga cattgtgtca ttgttcccta actacgattc 780
taggacttat cctatcagga ctgtttctca attgactagg gaaatctaca ctaacccagt 840
tcttgagaac ttcgacggtt ctttcagggg ttcagctcag ggtatcgagg gatctatcag 900
atcaccacac cttatggaca tcttgaactc aatcacaatc tacacagacg cacacagggg 960
tgagtactac tggtcaggtc accagatcat ggcatctcca gtgggtttct ctggtcctga 1020
gttcacattc cctttgtatg gaactatggg aaacgcagct cctcagcaga gaattgtggc 1080
tcagttggga cagggagtgt ataggactct ttcaacaaca ttgtatagga gaccattcaa 1140
cattggaatc aacaatcagc agctttcagt gcttgatggt acagagttcg catacggtac 1200
atcatcaaac ttgccttctg ctgtgtacag gaagtctgga actgtggatt cattggacga 1260
gattccacct caaaacaaca acgtgcctcc aaggcaggga ttctcacaca ggttgtctca 1320
cgtgtctatg ttcaggtctg gattctcaaa ctcatctgtt tctattatca gggcacctat 1380
gttctcatgg atacacaggt cagcagagtt caacaatatc atcccttctt ctcaaatcac 1440
tcagatccct cttactaagt ctacaaactt gggatctgga acatcagtgg tgaagggtcc 1500
aggattcact ggtggtgaca tcttgaggag gacatctcca ggtcagatct ctacacttag 1560
agttaacatc acagctcctc tttctcagag gtacagggtt aggatcaggt atgcatctac 1620
aactaacttg cagttccaca cttcaattga tggaaggcca atcaaccagg gtaacttctc 1680
tgcaactatg tcatcaggtt caaaccttca gtcaggatca ttcaggacag ttggtttcac 1740
aacaccattc aacttctcta atggatcatc agttttcaca ttgtcagcac acgtgttcaa 1800
ctcaggtaat gaagtgtata tcgacaggat tgagttcgtt ccagcagagg ttacattcga 1860
ggcagagtac gacttggaga gggcacagaa ggcagtgaac gagcttttca catcttcaaa 1920
tcaaatcggt ttgaaaacag atgttacaga ctaccatatc gaccaggtgt ctaactccga 1980
gaaggatgaa ctttgataaa atctctagag gatccccggg taccgagctc gaatcg 2036
- 还没有人留言评论。精彩留言会获得点赞!