一种外切型褐藻胶裂解酶、基因及其应用的制作方法
本发明涉及基因工程和酶工程的技术领域,尤其涉及一种外切型褐藻胶裂解酶、基因及其应用。
背景技术:
我国海藻资源丰富。褐藻是一种重要的经济海藻,代表植物包括海带、鹿角菜和马尾菜等。褐藻含有褐藻胶、褐藻淀粉和褐藻糖胶等多糖,占脱脂褐藻干重的40%~80%,其中褐藻胶含量最为丰富。褐藻胶是由β-d-甘露糖醛酸及其c5差向异构体α-l-古罗糖醛酸两种糖醛酸单体通过β-1,4糖苷键连接形成的线性高分子聚合物,广泛应用于食品、医药、化学工业等领域中。褐藻胶在褐藻的细胞壁和细胞内基质中能够大量合成,也可以由一些特定细菌产生。近年来,以褐藻为原料生产乙醇成为藻类生物燃料研究的热点。
褐藻胶裂解酶通过β-消除反应裂解褐藻胶的糖苷键。该酶可作为内切型,切割多糖,也可作为外切型,切割褐藻胶或褐藻胶寡糖末端。利用褐藻胶裂解酶生产褐藻胶寡糖,具有高效、降解条件温和、水解产物不易被破坏、回收工序简单、不污染环境等优点,为褐藻寡糖在功能性食品、多功能化妆品、新产品药物等方面的研究与应用提供了新的手段。除了用于制备褐藻胶寡糖,褐藻胶裂解酶还应用于治疗纤维化囊肿、制备原生质体、从褐藻中提取生物活性物质、分析褐藻胶结构、生产生物能源等中。
褐藻胶裂解酶主要来源于多种微生物、海洋藻类、海洋软体动物和棘皮动物等体内。本发明人研究和设计了一个来自微泡菌alw1(microbulbifersp.alw1)的外切型褐藻胶裂解酶、基因及其应用,本案由此产生。
技术实现要素:
本发明的目的在于提供一种外切型褐藻胶裂解酶、基因及其应用,该酶可用于产生单糖和寡糖,可应用于食品、医药、化学工业、能源等领域中。
为了实现上述目的,本发明解决其技术问题所采取的技术方案是:
一种编码外切型褐藻胶裂解酶algl17的基因,其核苷酸序列如seqidno.1所示。基因大小为2220bp。
一种外切型褐藻胶裂解酶algl17,其氨基酸序列如seqidno.2所示。该蛋白编码739个氨基酸残基。
一种褐藻胶裂解酶表达载体,含有所述的外切型褐藻胶裂解酶algl17基因的表达载体pet-28a-algl17。
一种褐藻胶裂解酶的制备方法,包括采用所述的含有褐藻胶裂解酶基因的表达载体转化寄主细胞,培养转化体,从培养物中获得重组褐藻胶裂解酶。当然,也可将褐藻胶裂解酶algl17基因转染到合适的真核生物宿主中,包括酵母及哺乳动物细胞等。
作为实施例的优选方式,所述的寄主细胞为大肠杆菌。
作为实施例的优选方式,包括采用所述的含有褐藻胶裂解酶基因的表达载体转化至寄主细胞大肠杆菌bl21(de3),经iptg诱导,得到可溶性表达的重组褐藻胶裂解酶。
作为实施例的优选方式,所述的iptg终浓度为0.1mmol/l,诱导温度为18℃。
一种重组褐藻胶裂解酶,包括采用所述的含有褐藻胶裂解酶基因的表达载体转化寄主细胞,培养转化体,从培养物中获得重组酶。
作为实施例的优选方式,以海藻酸钠为底物,该重组褐藻胶裂解酶的最适反应温度为35℃,最适反应ph为8.0。重组algl17在25℃下稳定,在30℃和35℃下不稳定。该酶在ph5.0-8.0具有良好的稳定性。通过研究不同浓度的na+对酶活性的影响,结果表明,na+对重组酶活力有激活作用,0.7mol/lnacl的激活作用最强。抑制剂edta,β-me和dtt可抑制重组酶的活性。该酶对tritonx-100,tween20和tween80具有良好的抗性。相反,sds和ctab大大抑制酶的活性。
重组algl17对海藻酸钠、polym和褐藻胶寡糖具有活性,而对polyg具有较低活性,表明该重组酶是polym特异性寡褐藻胶裂解酶。重组酶可作用于海藻酸钠产生deh,表明该酶是外切型褐藻胶裂解酶。另外,该重组酶能直接降解海带,产生还原糖。
本发明从微泡菌alw1(microbulbifersp.alw1)中克隆获得了一个新的褐藻胶裂解酶algl17基因,发现了该基因编码的酶蛋白在食品、医药、化学工业、能源等领域中具有应用潜力。同时,本次发明也克隆得到了可以大量表达的工程菌,可实现该褐藻胶裂解酶的规模化生产,为后续的工业化应用提供了良好的基础。
附图说明
图1为褐藻胶裂解酶algl17的诱导表达及其纯化检测的sds-page图;其中,1:分子量标准蛋白;2:含pet-28a(+)阴性菌,iptg诱导;3:含pet-28a-algl17阳性菌,未诱导;4:含pet-28a-algl17阳性菌,iptg诱导;5:纯化后的重组蛋白;
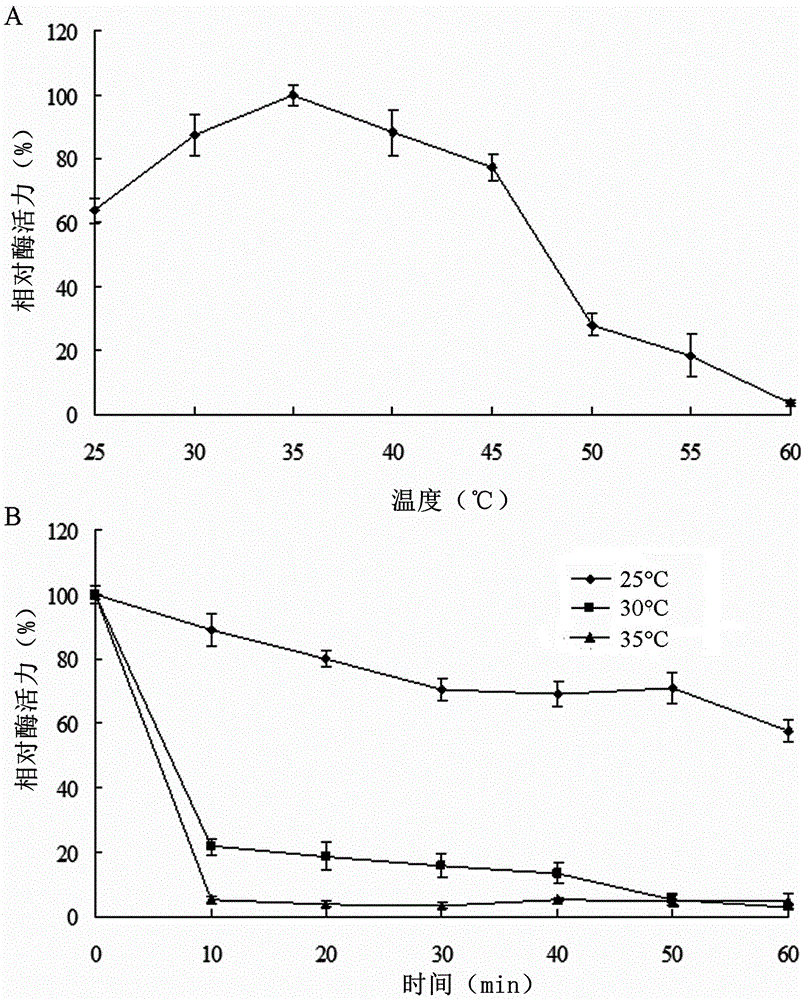
图2为温度对重组褐藻胶裂解酶algl17的活性和稳定性的影响曲线图;a:温度对重组酶活性的影响;b:温度对重组酶稳定性的影响;
图3为ph对重组褐藻胶裂解酶algl17的活性和稳定性的影响曲线图;
图4为金属离子对重组褐藻胶裂解酶algl17活性的影响曲线图;a:不同金属离子对重组酶活性的影响;b:na+对重组酶活性的影响;
图5为重组褐藻胶裂解酶algl17的底物特异性分析图;
图6为重组褐藻胶裂解酶algl17作用于海藻酸钠的酶解产物分析图;a:海藻酸钠酶解处理后的液相色谱分析;b:未经酶解处理的海藻酸钠液相色谱分析;c:5.28min主峰处的产物质谱分析;d:6.94min主峰处的产物质谱分析;
图7为重组褐藻胶裂解酶algl17作用于海带粉的分析图。
具体实施方式
实施例1:褐藻胶裂解酶algl17基因的重组表达质粒与重组菌株的构建
利用细菌基因组提取试剂盒(takara)提取微泡菌alw1(microbulbifersp.alw1)的基因组,利用该酶基因特异性引物,扩增褐藻胶裂解酶algl17基因:
上游引物(seqidno.3):5'-cgcggatccgttgaacatcccaacctggtgatcg-3';
下游引物(seqidno.4):5'-ccgctcgaggttagtaatttggaagagttttgcg-3';
pcr反应条件为:95℃5min;94℃45sec,50℃45sec,72℃,2min,30个循环;72℃10min。pcr产物利用琼脂糖凝胶电泳检测后,割胶回收目的基因,产物与pet-28a(+)载体经bamhi和xhoi酶切后进行连接,将靶基因插入pet-28a(+)载体中,所得构建体命名为pet-28a-algl17。一旦通过测序确认基因序列,将重组质粒导入大肠杆菌bl21(de3)。
实施例2:利用重组表达菌株表达和纯化重组褐藻胶裂解酶
重组表达菌株过夜培养后,按体积比1:100转接到250mllb液体培养基(含50μg/ml卡那霉素)中,37℃、180r/min培养至od600达到0.8,加入终浓度为0.1mmol/liptg,18℃、180r/min培养20h。通过在6,000×g离心10min收集菌体。细胞重悬于结合缓冲液(50mmol/lnah2po4,300mmol/l氯化钠,15mmol/l咪唑,ph8.0)中,在冰上进行超声波破碎处理。4℃下18,000×g离心20min,收集上清,然后进行ni-nta亲和层析,经过洗涤缓冲液(50mmol/lnah2po4,300mmol/l氯化钠,30mmol/l咪唑,ph8.0)洗涤后,利用洗脱缓冲液(50mmol/lnah2po4,300mmol/l氯化钠,250mmol/l咪唑,ph8.0)洗脱,收集洗脱液。将获得的重组褐藻胶裂解酶algl17用50mmol/l磷酸钠缓冲液(ph8.0)透析后,经sds-page检测,重组蛋白的分子量为85kda,结果如图1所示。使用bca蛋白测定试剂盒(pierce,usa)测定蛋白质浓度,重组酶的浓度约为0.6mg/ml。
实施例3:褐藻胶裂解酶的活力检测
通过使用3,5-二硝基水杨酸(dns)方法测定释放的还原糖的量来检测褐藻胶裂解酶的活性。具体操作:将10μl酶液(0.6mg/ml)加入到390μl含有2.5mg/ml海藻酸钠的50mmol/l磷酸钠缓冲液(ph8.0)中,在35℃下温育10min后,加入400μldns试剂,并在100℃下沸水浴加热10min来终止反应。冷却至室温后,使用epoch2t分光光度计(biotek,usa)在540nm处监测还原糖的释放量。一个单位的褐藻胶裂解酶活性定义为在测定条件下每分钟释放1μmol还原糖所需的酶量。
实施例4:温度和ph对重组褐藻胶裂解酶algl17的影响
褐藻胶裂解酶的最适反应温度在25℃–60℃的范围内测定,具体操作:采取前文所用的体系,分别在25、30、35、40、45、50、55和60℃条件下反应10min,测定540nm的吸光值。测定结果(图2a)表明,褐藻胶裂解酶algl17最适反应温度为35℃,并且在25–45℃的温度范围内呈现超过60%的最大活性,在60℃几乎没有活性。酶的热稳定性分析在25、30和35℃下进行测定。具体操作为:将纯化后的酶液在不同温度下处理10–60min。热处理后将样品放在冰上立即冷却。通过所述的标准方法测量酶的剩余活力,以未经处理的酶活性定义为100%。测定结果(图2b)显示,该酶在25℃相对稳定。在30℃处理30min使其84%的相对酶活性损失。该酶在35℃热处理后几乎没有活性。这些结果表明该重组酶具有热不稳定性。
褐藻胶裂解酶的最适反应ph在4.0–11.0的范围内测定。测定使用的缓冲液为:50mmol/l柠檬酸缓冲液(ph4.0–6.0)、50mmol/l磷酸钠缓冲液(ph7.0–8.0)、50mmol/l巴比妥钠-hcl(ph8.5–9.0)和50mmol/l甘氨酸-naoh(ph为9.5–11.0)。测定结果(图3)表明,褐藻胶裂解酶algl17的最适反应ph为8.0,该酶在7.5–9.0的ph范围内显示出超过30%的最大活性。褐藻胶裂解酶的ph稳定性在4.0–11.0范围内进行测定。具体操作为:将酶置于不同ph值的缓冲液中,25℃温育1h后,测定酶的剩余活力,以未经处理的酶活性定义为100%。测定结果(图3)显示,重组酶在ph范围为5.0–8.0时其稳定性保持在最初活性的50%以上,在ph9.5下保持18%的残留活力。
实施例5:添加剂对重组褐藻胶裂解酶algl17活性的影响
在褐藻胶裂解酶中分别添加终浓度为1mmol/l或10mmol/l的不同金属盐离子,包括nacl、kcl、cacl2、mgcl2、fecl2、cucl2、cocl2、mncl2、bacl2、cdcl2和zncl2。置于25℃下放置1h后,测定酶的剩余活力,以未添加金属离子的酶活力为100%。测定结果(图4a)显示,fe2+、cu2+、co2+、mn2+、ba2+、cd2+和zn2+会抑制酶的活性,cd2+和zn2+的影响显著。1mmol/l的k+、ca2+和mg2+对酶的活性没有影响,而在10mmol/l条件下具有抑制作用。1mmol/l和10mmol/l的na+能促进酶的活性。进一步研究不同浓度的na+对酶活性的影响,结果(图4b)表明,0.7mol/lnacl的激活作用最高,具有170%的相对活性,但在1.0mol/lnacl下酶活性被抑制。这些结果表明重组褐藻胶裂解酶是na+活化的酶。
在褐藻胶裂解酶中分别添加终浓度为1mmol/l或10mmol/l的抑制剂(包括edta、β-me和dtt)和0.1%(w/v或v/v)或1%(w/v或v/v)的去垢剂(包括sds、tritonx-100、tween20、tween80和ctab)。置于25℃下放置1h后,测定酶的剩余活力,以未添加抑制剂或去垢剂的酶活力为100%。测定结果如表1所示,edta、β-me和dtt都在所测试的两种浓度下显示出对酶活性的抑制作用。该酶对tritonx-100、tween20和tween80具有良好的抗性。相反,sds和ctab大大抑制酶的活性。在浓度为1%的sds和ctab条件下,酶几乎完全丧失活性。
表1抑制剂和去垢剂对重组褐藻胶裂解酶活性的影响
实施例6:重组褐藻胶裂解酶algl17的底物特异性
将2mg/ml海藻酸钠、polym、polyg和褐藻胶寡糖(3.3kda)分别作为底物,通过分析在235nm处的吸光值大小来测定褐藻胶裂解酶的活性,研究酶的底物特异性。具体操作为:分别向8ml不同底物溶液中加入100μl酶溶液(0.6mg/ml),35℃下反应90min。通过利用uv-5200分光光度计(上海元析仪器有限公司)测定反应混合物在235nm的吸光值,反映产生的不饱和糖醛酸的量。结果(图5)显示,重组褐藻胶裂解酶对polym、海藻酸钠和褐藻胶寡糖具有活性,但对polyg显示出非常低的活性。这些结果表明重组酶是polym特异性寡褐藻胶裂解酶。
实施例7:重组褐藻胶裂解酶algl17作用于海藻酸钠的酶解产物分析
取褐藻胶裂解酶(6u)加入到30ml5.0mg/ml海藻酸钠底物溶液中,在35℃下进行反应。每隔1h取样一次,通过dns法测定释放的还原糖的量。反应8h后,添加3u的褐藻胶裂解酶到反应混合物中,在相同温度下继续反应9h,此时释放的还原糖的量没有变化。通过在沸水中加热混合物10min,然后在冰上冷却5min来终止反应。4℃下18,000×g离心20min后,收集上清液作为水解产物样品。使用配备有acquitybehc18柱(2.1×150mm,1.7μm)的acquityhplc/maldisynaptq-tofms(waters,usa)鉴定酶解产物。两个流动相分别是溶液a:0.1%(v/v)甲酸水溶液,溶液b:乙腈,梯度流速0.3ml/min。使用synaptq-tof质量检测器(waters)获得质谱图。结果(图6)表明,重组algl17降解海藻酸钠产物的液相色谱在保留时间5.28min和6.94min时出现两个主峰(图6a),而未酶解的海藻酸钠没有出现这些峰(图6b)。利用ms测定上述主峰中的产物,在负离子模式下,产物质荷比(m/z)的聚合度(dps)出现的离子峰[dpx–h]—(x=1,2,3,4,5)分别为1(175m/z)、2(351m/z)、3(527m/z)、4(703m/z)和5(879m/z)。质谱结果(图6c-d)显示,重组algl17介导的海藻酸钠降解产物为deh、二糖、三糖和四糖。
实施例8:重组褐藻胶裂解酶algl17作用于海带粉
在蒸馏水中清洗海带片后,将干燥的样品研磨成粉末。将粉末加入至50mmol/l磷酸钠缓冲液(ph8.0)中,制备5mg/ml的海带粉底物。30ml底物中加入6u褐藻胶裂解酶,在35℃下进行酶促反应,每隔1h取样。所取的样品沸水浴加热5min终止反应,然后在冰上冷却5min。4℃下18,000×g离心20min后,利用dns法测定上清液中释放的还原糖的量。反应6h后,向反应混合物中加入3u褐藻胶裂解酶,在相同温度下继续反应14h,此时还原糖的释放量没有变化。将未经重组酶处理的底物用作阴性对照。结果(图7)显示,随着处理时间的延长,还原糖的浓度增加。重组algl17处理底物(150mg)20h后,释放的还原糖稳定,其量达到约16mg。这些结果表明重组褐藻胶裂解酶algl17可以作用于海带产生还原糖。
综上所述,本发明从microbulbifersp.alw1中克隆获得了新型的褐藻胶裂解酶algl17的基因,发现了该基因编码的酶蛋白可降解海藻酸钠产生单糖和寡糖,能降解海带粉产生还原糖,可应用于食品、医药、化学工业、能源等领域中。同时,本次发明也克隆得到了可以大量表达的工程菌,可实现该褐藻胶裂解酶的规模化生产,为后续的工业化应用提供了良好的基础。
本领域的普通技术人员能从本发明公开内容直接导出或联想到的所有变形,均应认为是本发明的保护范围。
sequencelisting
<110>集美大学
<120>一种外切型褐藻胶裂解酶、基因及其应用
<160>4
<170>patentinversion3.3
<210>1
<211>2220
<212>dna
<213>微球茎菌(microbulbifersp.)
<400>1
atgtttggtaaatttttctttaaccagcgtaaagctctgtccgccgccgtcgcggtgctg60
gcaatggctccattgtcgggacttgccgttgaacatcccaacctggtgatcgacgccgca120
gacgtggatgccatgcaaggcgcggtagcaaaacccggtcgctttcgcagcgcatttctc180
gcgtccaaatcagcggtagaccatgcgctgcaagtgccactggcggtaccagtgcccacc240
gacgcgggtggaggctatacccacgagcagcacaaaaagaactatcagctgatgtacaac300
gctggcgtgctttatcagattaccgaagatcccaaatatgcggaacgcgtgcgggatatg360
ctgctggcttatgccgatctgtaccctaccctgccgctccatcccaagcgtcgcccgggg420
gcagaaaatcccggcaagctgttctggcagagcctgaatgaagcggtttggctggtgtac480
accatccaggcctacgatctgattcgcccttcattgtccaatgcagaagcagaaaaaatc540
gaacagggtgcgctgcgcccggtggctaaatttctttccgtagaatccccggccaccttc600
aacaaggtgcataaccacggcacctggttgaccgccggagtgggtatggcgggctatgta660
ttggacgagccagagtgggttgagcaggcgctcctcgacctggacaagtccggcaagggt720
ggcttccttcgccagctgaacaccctgttctcaccggatggctactacaacgaaggtccc780
tactaccagcgctacgcgctgatgccatttgtcaccttcgccaaagcgatcgaaaacaac840
gagccggaacggggcattttcaagtaccgcgatggcattgtgatgaaggccatcgatacc900
accatccagctcagctacaacaatctgtttttcccgatcaacgatgccatcaagagtaaa960
ggcatcgacacctcggagctggtgctgggcgtcaccattgcctatggcgagtccggcaat1020
ccacaactgctggatatcgccgacagacagcatcagattctgttgtccggtgacggcctc1080
aaggttgcgcagggactcgacgccggcgcactacagccttacccgttcaagtctttcgcc1140
ttccgcgatggaaaggacggcgacgaaggcgcactggttgtgctgcgccagcaaaccgac1200
ggcgaccaggcgttagtgttcaagccagccgctcagggcatgggccacggtcactttgac1260
cgcttcctgaatgtggaagccaaaaatggcggccgctatctgcaagaaaacgaaacctgg1380
gccaagcagaccattgcgcacaacacggtggtggtggacgagacctcccacttcgacaac1440
aacctgaaaatcgcaaacaggaaccatcctgagttgctgttcttccacgcggatgaccag1500
gtcaaaatcagtgcggcagaaatcgattccgcctatcccggtgtcagcttgaaacgcact1560
ctggcgctcgttaacaacccggagagcggcaatagcttcgccatcgatgtgttcggtgtg1620
gaatccagccagaaacaccagttggaccttcctttgcactacaacggccagctggtggat1680
accaatttcaggttgcagggctttaccgacagtctcaaggcactgggtaccaacaatggc1740
taccagcacctttggctcaaagcccgcggcaaaccggattccggactggcacaggtcacc1800
tggctgaacgacaacggccgtttctatacccaatccagcctggtagacggcaaaaccgag1860
ctgttgttcaccgagctgggcgccaacgatcccaacttcaacctgcgctccgaaaaaggc1920
tttatcgctcgccgcaatggtgcgcgttcacatacctttgtaagcgtgttggagccccat1980
ggcgagtacaacccgtccaaggaatttaccctggaggcggaaagccaagtgcaggcgcta2040
cagcatcggcaggccggtgacctcgaactgatcgccatcggcatcaagaacggtgccaca2100
cagctgcttgcgtacaaccgcagtagcaatgtgccagaagaactcgaaaacatttttgaa2160
tatgacggccgtaagtaccagttcactggccgcgcaaaactcttccaaattactaactga2220
<210>2
<211>739
<212>prt
<213>微球茎菌(microbulbifersp.)
<400>2
metpheglylysphephepheasnglnarglysalaleuseralaala
151015
valalavalleualametalaproleuserglyleualavalgluhis
202530
proasnleuvalileaspalaalaaspvalaspalametglnglyala
354045
valalalysproglyargpheargseralapheleualaserlysser
505560
alavalasphisalaleuglnvalproleualavalprovalprothr
65707580
aspalaglyglyglytyrthrhisgluglnhislyslysasntyrgln
859095
leumettyrasnalaglyvalleutyrglnilethrgluaspprolys
100105110
tyralagluargvalargaspmetleuleualatyralaaspleutyr
115120125
prothrleuproleuhisprolysargargproglyalagluasnpro
130135140
glylysleuphetrpglnserleuasnglualavaltrpleuvaltyr
145150155160
thrileglnalatyraspleuileargproserleuserasnalaglu
165170175
alaglulysilegluglnglyalaleuargprovalalalyspheleu
180185190
servalgluserproalathrpheasnlysvalhisasnhisglythr
195200205
trpleuthralaglyvalglymetalaglytyrvalleuaspglupro
210215220
glutrpvalgluglnalaleuleuaspleuasplysserglylysgly
225230235240
glypheleuargglnleuasnthrleupheserproaspglytyrtyr
245250255
asngluglyprotyrtyrglnargtyralaleumetprophevalthr
260265270
phealalysalailegluasnasngluprogluargglyilephelys
275280285
tyrargaspglyilevalmetlysalaileaspthrthrileglnleu
290295300
sertyrasnasnleuphepheproileasnaspalailelysserlys
305310315320
glyileaspthrsergluleuvalleuglyvalthrilealatyrgly
325330335
gluserglyasnproglnleuleuaspilealaaspargglnhisgln
340345350
ileleuleuserglyaspglyleulysvalalaglnglyleuaspala
355360365
glyalaleuglnprotyrprophelysserphealapheargaspgly
370375380
lysaspglyaspgluglyalaleuvalvalleuargglnglnthrasp
385390395400
glyaspglnalaleuvalphelysproalaalaglnglymetglyhis
405410415
glyhispheasplysleuthrtrpglnphetyraspargglygluglu
420425430
ilevalthrasptyrglyalaalaargpheleuasnvalglualalys
435440445
asnglyglyargtyrleuglngluasngluthrtrpalalysglnthr
450455460
ilealahisasnthrvalvalvalaspgluthrserhispheaspasn
465470475480
asnleulysilealaasnargasnhisprogluleuleuphephehis
485490495
alaaspaspglnvallysileseralaalagluileaspseralatyr
500505510
proglyvalserleulysargthrleualaleuvalasnasnproglu
515520525
serglyasnserphealaileaspvalpheglyvalglusersergln
530535540
lyshisglnleuaspleuproleuhistyrasnglyglnleuvalasp
545550555560
thrasnpheargleuglnglyphethraspserleulysalaleugly
565570575
thrasnasnglytyrglnhisleutrpleulysalaargglylyspro
580585590
aspserglyleualaglnvalthrtrpleuasnaspasnglyargphe
595600605
tyrthrglnserserleuvalaspglylysthrgluleuleuphethr
610615620
gluleuglyalaasnaspproasnpheasnleuargserglulysgly
625630635640
pheilealaargargasnglyalaargserhisthrphevalserval
645650655
leugluprohisglyglutyrasnproserlysgluphethrleuglu
660665670
alagluserglnvalglnalaleuglnhisargglnalaglyaspleu
675680685
gluleuilealaileglyilelysasnglyalathrglnleuleuala
690695700
tyrasnargserserasnvalproglugluleugluasnilepheglu
705710715720
tyraspglyarglystyrglnphethrglyargalalysleuphegln
725730735
ilethrasn
<210>3
<211>34
<212>dna
<213>人工序列
<400>3
cgcggatccgttgaacatcccaacctggtgatcg34
<210>4
<211>34
<212>dna
<213>人工序列
<400>4
ccgctcgaggttagtaatttggaagagttttgcg34
- 还没有人留言评论。精彩留言会获得点赞!