抗GPC3全人源化抗体、其嵌合抗原受体细胞及应用的制作方法
本发明涉及一种抗gpc3全人源化抗体、其嵌合抗原受体细胞及应用,属于生物技术领域。
背景技术
据发明人所知,肝细胞癌发病广,致死率高,缺乏有效的治疗手段。病人晚期存活率仅有3%[1]。多数病人确诊时已经处于癌变晚期,对化疗药物不敏感,手术切除后复发率高[2,3]。glypican-3(gpc3)是一种硫酸乙酰肝素蛋白聚糖,通过gpi锚定在细胞膜表面。gpc3在胚胎发育早期表达,通过调控发育过程中的关键信号通路(如wnt信号)参与调控细胞增殖。在个体发育成熟后,gpc3表达关闭。大量的研究发现,gpc3在肝细胞癌中特异性高表达,是目前较受关注的肝细胞癌诊断标志物和治疗靶点[4,5]。
由于肝脏的器官功能特殊性,小分子化学药物极易被肝脏代谢清除。相比较而言,生物大分子药物如治疗型抗体在肝癌的治疗中具有高活性、特异性好等优势。由于gpc3是细胞膜表面蛋白,靶向gpc3的抗体除应用于裸抗治疗外,抗体衍生物以及衍生的嵌合抗原受体t细胞均可作为治疗肝细胞癌的可行性手段,具有直接的临床转化潜力。
经检索发现,专利号cn201280029201.3,授权公告号cn103596985b的中国发明专利,公开了一种对磷脂酰肌醇蛋白聚糖3特异的人单克隆抗体及其用途。以该技术方案为代表的现有技术中已经存在了一些抗gpc3抗体。发明人课题组经系统地试验研究后得出了不同于现有技术的抗gpc3全人源化抗体。
技术实现要素:
本发明的主要目的是:针对现有技术存在的问题,提供一种抗gpc3全人源化抗体、以及具有该抗体的嵌合抗原受体细胞,有抗肿瘤应用前景。
本发明解决其技术问题的技术方案如下:
一种抗gpc3全人源化抗体,所述抗体包括重链和轻链;其特征是,所述抗体至少具有以下技术特征之一:
i、所述重链包括重链cdr1,即seqidno:2所示氨基酸序列中的氨基酸残基26-33;
ii、所述重链包括重链cdr2,即seqidno:2所示氨基酸序列中的氨基酸残基50-59;
iii、所述重链包括重链cdr3,即seqidno:2所示氨基酸序列中的氨基酸残基98-102;
iv、所述轻链包括轻链cdr1,即seqidno:2所示氨基酸序列中的氨基酸残基161-165;
v、所述轻链包括轻链cdr2,即seqidno:2所示氨基酸序列中的氨基酸残基182-185;
vi、所述轻链包括轻链cdr3,即seqidno:2所示氨基酸序列中的氨基酸残基223-228。
优选地,所述抗体至少具有以下技术特征之一:
i、所述重链包括重链cdr1,即seqidno:2所示氨基酸序列中的氨基酸残基26-33;所述重链包括重链cdr2,即seqidno:2所示氨基酸序列中的氨基酸残基50-59;所述重链还包括重链cdr3,即seqidno:2所示氨基酸序列中的氨基酸残基98-102;
ii、所述轻链包括轻链cdr1,即seqidno:2所示氨基酸序列中的氨基酸残基161-165;所述轻链包括轻链cdr2,即seqidno:2所示氨基酸序列中的氨基酸残基182-185;所述轻链包括轻链cdr3,即seqidno:2所示氨基酸序列中的氨基酸残基223-228。
优选地,所述抗体的氨基酸序列如seqidno:2所示。
优选地,所述抗体为vh单域结构抗体、fab片段、fab′片段、f(ab)′2片段、单链可变区片段(scfv)、二硫化合物稳定的可变区片段(dsfv)、igg分子、或双特异性抗体。
优选地,所述抗体具有标记,所述标记包括荧光标记、酶标记、以及放射性标记。
本发明还提供:
编码前文所述抗gpc3全人源化抗体的核酸。
优选地,所述核酸的序列如seqidno:1所示。
本发明还提供:
具有前文所述抗gpc3全人源化抗体的细胞或偶联物。
优选地,所述细胞包括嵌合抗原受体t细胞、嵌合抗原受体nk细胞、以及经人工编辑的细胞;所述偶联物包括抗gpc3全人源化抗体-细菌毒素偶联物、抗gpc3全人源化抗体-细菌毒素变体偶联物、抗gpc3全人源化抗体-细胞因子偶联物、以及抗gpc3全人源化抗体-化疗药物偶联物。
本发明还提供:
前文所述抗gpc3全人源化抗体用于制备细胞或偶联物、诊断试剂盒、药物或药物组合物的用途。
前文所述核酸用于制备抗gpc3全人源化抗体、药物或药物组合物的用途。
前文所述细胞或偶联物用于制备药物或药物组合物的用途。
其中,所述药物或药物组合物具有针对gpc3阳性肿瘤的抗肿瘤作用。
本发明通过噬菌体展示技术筛选出抗gpc3的全人源化抗体,该抗体特异性强,亲和力高;具有该抗体的细胞如嵌合抗原受体t细胞对gpc3阳性的肿瘤具有有效地杀伤作用。
附图说明
图1为实施例1中elisa检测富集噬菌体对抗原蛋白结合的示意图。
图2为实施例1中elisa检测噬菌体单克隆对抗原结合的示意图。
图3为实施例2中sds-page检测结果图。
图4为实施例3中抗体32a9对gpc3蛋白的特异性结合elisa检测结果图。
图5为实施例3中抗体32a9对gpc3阳性细胞的特异性结合facs检测结果图。
图6为实施例4中亲和力分析结果图。
图7为实施例5中构建的两个car分子的结构图。
图8为实施例5中慢病毒感染获得两个car-t细胞的流式细胞术检测结果图。
图9为实施例6中32a9car-t细胞的激活情况结果图。
图10为实施例6中32a9car-t细胞特异性杀伤a431-gpc3细胞的结果图。
具体实施方式
下面结合实施例对本发明作进一步详细描述。但是本发明不限于所给出的例子。所用方法如无特别说明均为常规方法,所用试剂和材料如无特别说明均为市售品。
实施例1、筛选抗gpc3全人源化抗体
采用噬菌体展示技术,以人源gpc3(q25-s550)蛋白为gpc3抗原进行筛选。
其中,该gpc3抗原的制备过程如下:构建pfuse-hgpc3(q25-s550)-fc真核细胞表达载体;该表达载体利用il-2的信号肽序列置换了gpc3的信号肽序列,引导gpc3-fc融合蛋白分泌到培养基中;利用lipofectaminetm2000(invitrogen,carlsbad,ca)将上述表达载体转染hek293t细胞,收集上清,并通过proteinaargrose(gehealthcare,piscataway,nj)亲和柱分离纯化gpc3-fc融合蛋白,所得即gpc3抗原。
噬菌体展示的具体过程为:以10μg/ml的上文所得gpc3抗原于4℃包被免疫板过夜;用含有5%脱脂奶粉,0.1%tween-20的pbs溶液室温封闭免疫板1小时;tomlinsoni&j噬菌体文库(genserviceltd.,cambridge,uk,文库大小为1.47x108)以1012pfu与5%脱脂奶粉pbs溶液1:1混合后室温孵育2小时,加入封闭好的免疫板中(100μl/孔),室温孵育1小时;用0.1%tween-20的pbs溶液洗涤免疫板20次;100μlph2.0洗脱缓冲液室温洗脱30分钟;向洗脱的噬菌体中加入30μlph8.0溶液进行中和;将中和后噬菌体感染对数生长期的tg1细胞,扩增和回收后用于下一轮淘选。淘选后elisa分析阳性噬菌体富集情况。
elisa检测:以5μg/ml上文所得gpc3抗原、阴性对照蛋白frizzled-fc、以及bsa分别于4℃包被免疫板过夜;用含有5%脱脂奶粉,0.1%tween-20的pbs溶液室温封闭免疫板1小时;将每一轮富集的噬菌体扩增并回收,以1:1比例与10%脱脂奶粉pbs室温孵育2小时,加入封闭好的免疫板中(100μl/孔),室温孵育1小时;用0.1%tween-20的pbs溶液洗涤免疫板5次;将hrp/anti-m13monoclonalconjugate以1:4000比例与含3%脱脂奶粉,0.05%tween-20的pbs溶液混合,加入洗涤好的免疫板中(50μl/孔),室温孵育1小时;用0.05%tween-20的pbs溶液洗涤免疫板5次;将tmb显色液加入免疫板中(100μl/孔),室温显色3分钟后,加入0.5m硫酸中止显色(100μl/孔);用酶联免疫检测仪在450nm波长下检测吸光值,并分析每轮扩增后噬菌体的亲和力。
elisa检测结果如图1所示,以frizzled-fc和bsa为抗原阴性对照,在三轮富集后,噬菌体群对gpc3抗原的亲和力显著升高。
对第三轮富集的噬菌体单克隆进行抗原结合分析,具体过程为:
用第三轮富集的噬菌体库感染tg1细胞,并从中随机挑取200个单克隆,扩增并回收噬菌体。以5μg/ml上文所得gpc3抗原、阴性对照frizzled-fc蛋白分别于4℃包被免疫板过夜;用含有3%脱脂奶粉,0.05%tween-20的pbs溶液室温封闭免疫板1小时;将200个扩增后的单克隆噬菌体,以1:1比例与6%脱脂奶粉pbs室温孵育1小时,分别加入包被有gpc3抗原和阴性对照frizzled-fc蛋白并封闭好的免疫板中(50μl/孔),室温孵育1小时;用0.05%tween-20的pbs溶液洗涤免疫板5次;将hrp/anti-m13monoclonalconjugate以1:4000比例与含3%脱脂奶粉,0.05%tween-20的pbs溶液混合,加入洗涤好的免疫板中(50μl/孔),室温孵育1小时;用0.05%tween-20的pbs溶液洗涤免疫板5次;将tmb显色液加入免疫板中(100μl/孔),室温显色3分钟后,加入0.5m硫酸中止显色(100μl/孔);用酶联免疫检测仪在450nm波长下检测吸光值,并分析单克隆噬菌体对gpc3抗原的结合能力。检测结果如图2所示,发现9个抗原结合阳性克隆。
分析上述9个抗原结合阳性的噬菌体单克隆序列,发生富集的克隆为抗体32a9,其scfv形式的dna序列为seqidno:1,氨基酸序列为seqidno:2。
seqidno:1:
gaggtgcagctgttggagtctgggggaggcttggtacagcctggggggtccctgagactctcctgtgcagcctctggattcacctttagcagctatgccatgagctgggtccgccaggctccagggaaggggctggagtgggtctcaggtattgcttataatggtgcttctacagcttacgcagactccgtgaagggccggttcaccatctccagagacaattccaagaacacgctgtatctgcaaatgaacagcctgagagccgaggacacggccgtatattactgtgcgaaatctgctggttcttttgactactggggccagggaaccctggtcaccgtctcgagcggtggaggcggttcaggcggaggtggcagcggcggtggcgggtcgacggacatccagatgacccagtctccatcctccctgtctgcatctgtaggagacagagtcaccatcacttgccgggcaagtcagagcattagcagctatttaaattggtatcagcagaaaccagggaaagcccctaagctcctgatctattctgcatcctctttgcaaagtggggtcccatcaaggttcagtggcagtggatctgggacagatttcactctcaccatcagcagtctgcaacctgaagattttgcaacttactattgtcaacaggattatgcttatccttatacgttcggccaagggaccaaggtggaaatcaaa。
seqidno:2:
evqllesggglvqpggslrlscaasgftfssyamswvrqapgkglewvsgiayngastayadsvkgrftisrdnskntlylqmnslraedtavyycaksagsfdywgqgtlvtvssggggsggggsggggstdiqmtqspsslsasvgdrvtitcrasqsissylnwyqqkpgkapklliysasslqsgvpsrfsgsgsgtdftltisslqpedfatyycqqdyaypytfgqgtkveik。
在氨基酸序列中,1-116为抗体重链序列,132-239为抗体轻链序列,117-131为连接肽序列。其中包含的cdr区域如下:氨基酸残基26-33(即gftfssya)为重链cdr1,氨基酸残基50-59(即giayngasta)为重链cdr2,氨基酸残基98-102(即ksags)为重链cdr3;氨基酸残基161-165(即issyl)为轻链cdr1,氨基酸残基182-185(即sass)为轻链cdr2,氨基酸残基223-228(即dyaypy)为轻链cdr3。
此外,该抗体形式可选自vh单域结构抗体、fab片段、fab′片段、f(ab)′2片段、单链可变区片段(scfv)、二硫化合物稳定的可变区片段(dsfv)、igg分子、或双特异性抗体。该抗体可选择具有标记,标记包括荧光标记、酶标记、以及放射性标记。
实施例2、抗体32a9的表达与纯化
将抗体32a9的重链序列和轻链序列分别克隆到表达载体pfuse-chig-hg1和pfuse-clig-hk(invivigen,sandiego,ca)中,制备质粒。用添加10%胎牛血清、100u/ml青霉素、0.1mg/ml链霉素的dmem培养基在细胞培养皿中种5百万个hek293t细胞,置于5%co2,37℃培养箱中培养。当细胞密度到达60-80%时,将5μgpfuse-32a9vh质粒和5μgpfuse-32a9vl质粒加入1mlopti-mem培养基中,静置5分钟;将30μgpei加入混有质粒的opti-mem培养基中,静置20分钟,静置期间为hek293t细胞更换新鲜的添加10%胎牛血清、100u/ml青霉素、0.1mg/ml链霉素的dmem培养基;静置20分钟后将混好的opti-mem培养基加入hek293t细胞培养皿中;每24小时回收上清液,并为hek293t细胞更换新鲜的添加10%胎牛血清、100u/ml青霉素、0.1mg/ml链霉素的dmem培养基继续表达蛋白。将回收的上清液于3500rpm、4℃条件下离心20分钟,并用0.45μm的微孔滤膜抽滤,进一步去除碎片;将上清液通过proteinaargrose(gehealthcare,piscataway,nj)亲和柱分离纯化32a9igg重组蛋白。通过bca法测得32a9igg重组蛋白浓度,并将5μg32a9igg蛋白进行聚丙烯酰胺凝胶电泳,得到32a9igg重组蛋白在变性和非变性的环境下的条带,如图3所示。
实施例3、抗体32a9的特异性分析
(1)抗原蛋白结合特异性
以5μg/ml的gpc3-his蛋白、对照gpc5-his蛋白分别于4℃包被免疫板过夜(gpc3-his蛋白、gpc5-his融合蛋白分别购于r&d);用含有3%脱脂奶粉,0.05%tween-20的pbs溶液室温封闭免疫板1小时;将32a9igg重组蛋白用含有3%脱脂奶粉,0.05%tween-20的pbs溶液稀释至5μg/ml,加入封闭好的免疫板中(50μl/孔),室温孵育1小时;用0.05%tween-20的pbs溶液洗涤免疫板3次(340μl/孔);将goatanti-humanfcγhrp(iacksonimmunoresearch)以1:2000比例与3%脱脂奶粉,0.05%tween-20的pbs溶液混合,加入洗涤好的免疫板中(50μl/孔),室温孵育1小时;用0.05%tween-20的pbs溶液洗涤免疫板3次;将tmb显色液加入免疫板中(100μl/孔),室温显色3分钟后,加入0.5m硫酸中止显色(100μl/孔);使用酶联免疫检测仪在450nm波长下检测吸光值,并分析32a9igg重组蛋白对gpc3蛋白的特异性。
如图4所示,结果表明抗体32a9特异性识别gpc3,但是不识别gpc3的同源蛋白gpc5。
(2)细胞结合特异性
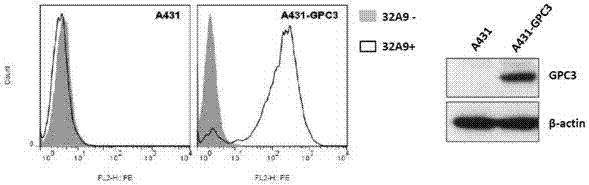
采用的a431细胞(人上皮癌细胞系)购于美国atcc(atcc,manassas,va)。在添加10%胎牛血清、100u/ml青霉素、0.1mg/ml链霉素的dmem培养基中,置于5%co2,37℃培养箱中培养;利用lipofectaminetm2000(invitrogen,carlsbad,ca)将gpc3cdna转染a431细胞,zeocin筛选获得gpc3过表达细胞系a431-gpc3细胞。
收集a431和a431-gpc3细胞,2000rpm,4℃离心5分钟,弃掉上清,并用pbsbuffer重悬,再次离心,弃掉pbs溶液,用含5μg/ml实施例2纯化的32a9igg重组蛋白,含5%bsa的pbs溶液重悬细胞,并放置冰上孵育1小时;2000rpm,4℃离心5分钟,弃掉上清,并用pbsbuffer重悬,并再次离心,弃掉pbs溶液,将gtf(ab)′2anti-humanigg(γ)r-peconjugate(invitrogen)以1:200比例与含5%bsa的pbs溶液混合并重悬细胞,并放置冰上孵育1小时;2000rpm,4℃离心5分钟,弃掉上清,并用pbsbuffer重悬,并再次离心,弃掉pbs溶液,用0.3mlpbs溶液重悬细胞,并用流式细胞仪检测pe荧光标记信号,分析32a9igg重组蛋白对gpc3阳性细胞的特异性。
如图5所示,结果表明抗体32a9特异性识别a431-gpc3细胞,而不识别a431细胞。
实施例4、抗体32a9的亲和力分析
采用elisa实验和流式细胞术实验,分别测定抗体32a9与gpc3-his蛋白以及gpc3阳性细胞的亲和力。
(1)抗原蛋白结合亲和力
使用5μg/ml的gpc3-his蛋白于4℃包被免疫板过夜;用含有3%脱脂奶粉,0.05%tween-20的pbs溶液室温封闭免疫板1小时;将实施例2所得32a9igg蛋白用含有3%脱脂奶粉,0.05%tween-20的pbs溶液以20μg/ml起始,1:2梯度稀释,加入封闭好的免疫板中(50μl/孔),室温孵育1小时;用0.05%tween-20的pbs溶液洗涤免疫板3次(340μl/孔);将goatanti-humanfcγhrp以1:2000比例与3%脱脂奶粉,0.05%tween-20的pbs溶液混合,加入洗涤好的免疫板中(50μl/孔),室温孵育1小时;用0.05%tween-20的pbs溶液洗涤免疫板3次;将tmb显色液加入免疫板中(100μl/孔),室温显色3分钟后,加入0.5m硫酸中止显色(100μl/孔);使用酶联免疫检测仪在450nm波长下检测吸光值,并拟合亲和力曲线分析32a9igg重组蛋白亲和力。
(2)细胞结合亲和力
收集a431和a431-gpc3细胞,2000rpm,4℃离心5分钟,弃掉上清,并用pbsbuffer重悬,再次离心,弃掉pbs溶液,用5%bsa的pbs溶液稀释实施例2所得32a9igg,以60μg/ml为起始,1:2梯度稀释,与细胞混匀后放置冰上孵育1小时;2000rpm,4℃离心5分钟,弃掉上清,并用pbsbuffer重悬,并再次离心,弃掉pbs溶液,将gtf(ab)′2anti-humanigg(γ)r-peconjugate(life)以1:200比例与含5%bsa的pbs溶液混合并重悬细胞,并放置冰上孵育1小时;2000rpm,4℃离心5分钟,弃掉上清,并用pbsbuffer重悬,并再次离心,弃掉pbs溶液,用0.3mlpbs溶液重悬细胞,并用流式细胞仪检测pe荧光标记信号,拟合32a9igg重组蛋白亲和力曲线并计算亲和力。
结果如图6所示,抗体32a9与gpc3-his蛋白的亲和力为1.93nm,抗体32a9与a431-gpc3细胞的亲和力为6.25nm。
实施例5、基于抗体32a9构建嵌合抗原受体t细胞
基于抗体32a9的序列,构建第二代4-1bb型car分子,同时构建fda批准的靶向cd19的car分子fmc63[6]作为对照,两者的结构如图7所示。
分离人pbmc细胞,用dynabeadshumant-activatorcd3/cd28和il-2定向诱导t细胞活化,利用慢病毒系统构建32a9car-t细胞,具体过程为:
合成32a9二代4-1bbcar分子,并克隆入慢病毒表达载体plvx-ef1a-puro中,制备质粒,转染hek293t细胞,收集病毒上清,超速离心2小时(20000rpm,4℃);分装病毒颗粒并测定病毒滴度。用添加10%胎牛血清、100u/ml青霉素、0.1mg/ml链霉素、50u/mlil-2的rpmi培养基复苏pbmc细胞到24孔板中。将dynabeadshumant-activatorcd3/cd28(invitrogen)以2000rpm,4℃离心5分钟,弃掉上清,并用1mlpbsbuffer重悬,并再次离心,弃掉pbs溶液,用50μl添加10%胎牛血清、100u/ml青霉素、0.1mg/ml链霉素的rpmi培养基重悬dynabeadshumant-activatorcd3/cd28,并加入pbmc细胞悬液中混匀;dynabeadshumant-activatorcd3/cd28激活pbmc细胞24小时后,将32a9carlentilvirus以moi=15的浓度混合10μg/mlpolybrene加入细胞悬液中,1000g,20℃离心1小时,并放入37℃细胞培养箱培养过夜。将细胞悬液1000rpm,室温离心5分钟,弃上清并更换为添加10%胎牛血清、100u/ml青霉素、0.1mg/ml链霉素、100u/mlil-2的rpmi培养基。更换培养基后每天检查细胞状态,当细胞密度达到2million/ml或者培养基变黄时为细胞更换新鲜的添加10%胎牛血清、100u/ml青霉素、0.1mg/ml链霉素、100u/mlil-2的rpmi培养基;感染病毒后第九天通过流式细胞术检测gfp荧光信号,检测结果如图8所示。
同时,以基本相同的过程构建fmc63car-t细胞,流式细胞术检测结果如图8所示。
实施例6、评估32a9car-t细胞的抗肿瘤效果
(1)激活检测
将扩增后的32a9car-t细胞(效应细胞)分别与2500个a431细胞和a431-gpc3细胞(靶细胞),按效靶比分别为1.25:1,2.5:1,5:1混合,在添加10%胎牛血清、100u/ml青霉素、0.1mg/ml链霉素的rpmi培养基的96孔细胞培养板中共孵育18小时后,得到上清;通过humanil-1elisakit(联科生物)检测上清中il-2的含量,分析32a9car-t细胞激活效率。
如图9所示,实验结果表明32a9car-t细胞与a431-gpc3细胞孵育后il-2的分泌量显著上升,这证明32a9car-t细胞能够有效地被gpc3阳性靶细胞激活。
(2)杀伤实验
将扩增后的32a9car-t细胞(效应细胞)分别与2500个a431细胞和a431-gpc3细胞(靶细胞),按效靶比分别为1.25:1,2.5:1,5:1混合,在添加10%胎牛血清、100u/ml青霉素、0.1mg/ml链霉素的rpmi培养基的96孔细胞培养板中共孵育18小时后,得到上清;通过cytotox96non-radioactivecytotoxicityassay(promega)试剂盒检测上清液中由裂解细胞释放的乳酸脱氢酶(ldh)的含量,分析32a9car-t细胞杀伤效率。
如图10所示,32a9car-t细胞能够显著地杀伤a431-gpc3细胞,而fmc63car-t细胞对a431-gpc3细胞无显著杀伤,这证明32a9car-t细胞能够有效地杀伤gpc3阳性的肿瘤细胞。
除嵌合抗原受体t细胞外,抗体32a9还可用于制备嵌合抗原受体nk细胞或经人工编辑的细胞,或制备偶联物。偶联物可选自细菌毒素偶联物、细菌毒素变体偶联物、细胞因子偶联物、化疗药物偶联物。
除上述实施例外,本发明还可以有其他实施方式。凡采用等同替换或等效变换形成的技术方案,均落在本发明要求的保护范围。
参考文献
[1].farazipa,depinhora.hepatocellularcarcinomapathogenesis:fromgenestoenvironment.[j].naturereviewscancer,2006,6(9):674-687.
[2].yangjd,robertslr.hepatocellularcarcinoma:aglobalview[j].natrevgastroenterolhepatol,2010,7(8):448-458.
[3].el-seraghb.hepatocellularcarcinoma[j].newenglandjournalofmedicine,2011,365(12):1118-27.
[4].filmusj,capurrom.glypican‐3:amarkerandatherapeutictargetinhepatocellularcarcinoma[j].febsjournal,2013,280(10):2471-2476.
[5].gaow,kimh,fengm,etal.inactivationofwntsignalingbyahumanantibodythatrecognizestheheparansulfatechainsofglypican-3forlivercancertherapy[j].hepatology,2014,60(2):576–587.
[6].cooperljn,jenab,maitis.anti-cd19scfv(fmc63)polypeptide:,us9701758[p].2017.
序列表
<110>南京医科大学
<120>抗gpc3全人源化抗体、其嵌合抗原受体细胞及应用
<160>2
<170>siposequencelisting1.0
<210>1
<211>717
<212>dna
<213>artificialsequence
<400>1
gaggtgcagctgttggagtctgggggaggcttggtacagcctggggggtccctgagactc60
tcctgtgcagcctctggattcacctttagcagctatgccatgagctgggtccgccaggct120
ccagggaaggggctggagtgggtctcaggtattgcttataatggtgcttctacagcttac180
gcagactccgtgaagggccggttcaccatctccagagacaattccaagaacacgctgtat240
ctgcaaatgaacagcctgagagccgaggacacggccgtatattactgtgcgaaatctgct300
ggttcttttgactactggggccagggaaccctggtcaccgtctcgagcggtggaggcggt360
tcaggcggaggtggcagcggcggtggcgggtcgacggacatccagatgacccagtctcca420
tcctccctgtctgcatctgtaggagacagagtcaccatcacttgccgggcaagtcagagc480
attagcagctatttaaattggtatcagcagaaaccagggaaagcccctaagctcctgatc540
tattctgcatcctctttgcaaagtggggtcccatcaaggttcagtggcagtggatctggg600
acagatttcactctcaccatcagcagtctgcaacctgaagattttgcaacttactattgt660
caacaggattatgcttatccttatacgttcggccaagggaccaaggtggaaatcaaa717
<210>2
<211>239
<212>prt
<213>artificialsequence
<400>2
gluvalglnleuleugluserglyglyglyleuvalglnproglygly
151015
serleuargleusercysalaalaserglyphethrphesersertyr
202530
alametsertrpvalargglnalaproglylysglyleuglutrpval
354045
serglyilealatyrasnglyalaserthralatyralaaspserval
505560
lysglyargphethrileserargaspasnserlysasnthrleutyr
65707580
leuglnmetasnserleuargalagluaspthralavaltyrtyrcys
859095
alalysseralaglyserpheasptyrtrpglyglnglythrleuval
100105110
thrvalserserglyglyglyglyserglyglyglyglyserglygly
115120125
glyglyserthraspileglnmetthrglnserproserserleuser
130135140
alaservalglyaspargvalthrilethrcysargalaserglnser
145150155160
ilesersertyrleuasntrptyrglnglnlysproglylysalapro
165170175
lysleuleuiletyrseralaserserleuglnserglyvalproser
180185190
argpheserglyserglyserglythraspphethrleuthrileser
195200205
serleuglnprogluaspphealathrtyrtyrcysglnglnasptyr
210215220
alatyrprotyrthrpheglyglnglythrlysvalgluilelys
225230235
- 还没有人留言评论。精彩留言会获得点赞!