用于治疗疾病的三环DNA反义寡核苷酸、组合物和方法与流程
发明背景
总的来说,本公开涉及合成的反义寡核苷酸(aon),以及使用反义寡核苷酸改变在pre-mrna加工期间发生的剪接事件、或下调含有重复序列例如3’或5’cug、cag和/或ccug的突变mrna的表达的方法。更具体来说,本文公开了三环dna(tc-dna)aon,其能够有效促进pre-mrna加工期间的外显子跳跃、掩蔽pre-mrna中的内含子沉默序列和/或茎-环序列、以及引导rnase介导的mrna破坏。本文描述了这样的tc-dnaaon,通过跳过肌养蛋白基因内突变的外显子、例如突变的外显子23或外显子51以恢复肌养蛋白的功能性,可用于杜氏肌营养不良(duchennemusculardystrophy)的治疗方法中。还描述了这样的tc-dnaaon,其通过掩蔽smn2基因内的内含子沉默序列和/或末端茎-环序列,以产生能够至少部分补偿无功能smn1蛋白、包含由外显子7编码的氨基酸序列的修饰的功能性smn2蛋白,从而可用于脊髓性肌萎缩的治疗方法中。此外本文还描述了这样的tc-dnaaon,其通过引导对包含3’-末端cug重复序列的突变dm1mrna的破坏,可用于steinert肌强直性营养不良的治疗方法中。因此,tc-dnaaon以及一种或多种上述方法可用于恢复与肌病相关的蛋白的功能性。
背景技术:
杜氏肌营养不良(dmd)是最常见的遗传性肌病,3,500名男性中约有1人患有此病,与种族无关。尽管不太常见,在显性携带者中,女孩和妇女也可表现出类似杜氏肌营养不良的症状。dmd的最主要后果是肌纤维变得特别脆,并且自然的肌肉活动就会激起肌肉组织的普遍损伤。在dmd以及许多肌营养不良中观察到的终点,是缓慢退化导致几乎完全纤维变性并伴有脂肪浸润。由于脊柱变形和呼吸困难,在1960年代时的预期寿命约为15岁。在不存在心脏并发症的情况下,现代在管理方法上的改进(即关节固定术和气管切开术通气)已经将预期寿命提高至30岁。
dmd的临床症状在18个月至三岁时变得明显,包括行走和爬行能力滞后、难以从地板上起立和小腿肚异常增大。在约5至6岁时,在足、膝和髋关节中发生肌肉收缩。疾病发展的特征是持续性肌肉消耗,导致在约9至12岁时丧失行走能力。此外,一些杜氏肌营养不良男孩表现出智力迟钝,表明缺失的蛋白也与中枢神经系统有关。
杜氏肌营养不良是x染色体连锁的隐性病。1986年,通过定位克隆方法鉴定到dmd基因座位于x染色体上(xp21.2–omimid:310200),在编码被称为肌养蛋白的蛋白的基因中。肌养蛋白基因中的突变导致在横纹肌中不能产生肌养蛋白。患病男孩的母亲有三分之二的几率携带肌养蛋白突变,而接近三分之一的双亲具有新生突变。一半以上的dmd男孩表现出涵盖一至几个外显子的大基因组缺失;他们中极少数具有大的序列重复。其他人具有难以鉴定的点突变或非常小的缺失或重复。
然而,突变的程度与表型的严重性不直接相关。产生提前终止密码子的框外缺失或无义突变以及随后翻译的失败,导致以严重表型为特征的肌养蛋白缺乏。框内缺失造成较轻的肌病,被称为贝克肌营养不良(beckermusculardystrophy,bmd)。
dmd基因座具有接近250万个碱基对,是曾经检测到的最长的基因,但是只有约14,000个碱基对含有编码序列,其分布在79个外显子中。全长肌养蛋白(dp427)是在所有肌肉中表达的427kda的细胞骨架蛋白,但通过分别位于内含子29、43、55和62中的四个内部启动子的组织特异性的差异使用(在视网膜、中枢神经系统、外周神经系统和非肌肉组织中),产生了各种蛋白质同工型(dp260、dp140、dp116、dp71)。
全长肌养蛋白是肌纤维膜糖蛋白复合物(sgc)的必要组分,所述复合物通过将肌纤维细胞骨架与细胞外基质相连而参与维持肌纤维膜的完整性。序列分析预测肌养蛋白需要几个结构域和重复序列。大体上说,在n-端存在肌动蛋白杂交位点(n-abd);含有可以赋予柔性的4个铰链区段(h)的中央杆结构域(rd;具有24个血影蛋白样重复序列);以及靠近c-端(ct)的富半胱氨酸结构域(crd),其与dpc的其他成员结合。
结构/功能分析已经鉴定了对于蛋白质功能关键的结构域。其以在某些患有轻度疾病的患者中发生的内部缺失为例证,在这些患者中所述缺失涵盖了外显子17至48(编码序列的46%)。england等,nature343(6254):180-2(1990)。这产生了在过去10年中广泛用于基因转移实验中的功能性“微小肌养蛋白”的概念。现在确定,移除n-abd和ct结构域引起功能的中度丧失,而crd是必需的。rd的改变根据截短的程度和性质引起多种多样的表型。例如,rd缺失的肌养蛋白(δr1-r24)是无功能的,然而(δh2-r19)截短的肌养蛋白,其保留了24个血影蛋白样重复序列中的8个完整重复序列,产生具有完全活性的蛋白。
对于杜氏肌营养不良,存在两种充分表征的遗传动物模型。mdx小鼠在肌养蛋白基因的外显子23中带有无义突变,其阻止了全长的野生型肌养蛋白的合成。mdx小鼠显示出对抗退化的代偿性机制,其可以维持再生过程以恢复机械损伤。mdx小鼠不表现出dmd的症状,其生命跨度几乎是正常的。
grmd(金毛猎犬肌营养不良)狗由于内含子6中的剪接位点突变破坏了阅读框而缺乏功能性肌养蛋白。在grmd中,与人类dmd相同,纤维的进行性退化不可避免地导致骨骼肌肉系统消耗,并伴有显著的肌内膜和肌束膜纤维变性。由于其类似dmd的表型,grmd一直是可获得的用于评估dmd潜在疗法的最佳模型。
尽管已经鉴定和表征了肌养蛋白基因中与dmd发作相关的突变并已有适合的动物模型系统来测试有希望的治疗药剂,但在本技术领域中对于治疗这种疾病的组合物和方法,仍存在着需求。在过去10年中进行的几项研究支持了类固醇疗法(强的松和地夫可特)在杜氏肌营养不良男孩中的益处,尽管广泛的统计评估还没有完全完成。利用庆大霉素药物治疗进行提前终止密码子突变的药物诱导的通读,在高达5%的dmd患者中可能也是有效的。尽管在mdx小鼠模型中进行的临床前研究的结果是有争论的,但目前正在美国和意大利进行临床试验。由ptctherapeutics开发的新药(ptc124)似乎非常有希望。使用药物上调utrophin基因的研究也在进行之中,所述基因的产物——肌养蛋白样蛋白utrophin,能够补偿失去的肌养蛋白的功能。
还存在许多其他研究途径;例如,最近显示,通过使用阻断性抗体而拮抗肌生成抑制素,能够在mdx小鼠中提高肌肉强度。这种方法最初是基于将正常成肌细胞多次注入患病肌肉中。partridge等,nature337(6203):176-9(1989)。随后的临床试验(1991-98)失败,尽管改进的细胞制造和递送程序使得在加拿大进行新的i期试验(2002)成为可能。最近的发展还提供了证据,表明来自骨髓或血管来源的干细胞能够通过全身途径靶向骨骼肌,尽管遗传校正的程度仍然不足。
用于dmd的基因治疗依赖于使用基因载体作为介质,将肌养蛋白微小基因原位递送到骨骼纤维中。使用质粒载体中的裸露全长cdna的第一个探索性研究在法国进行(2000-03)。在已试验过的用于肌肉基因治疗的不同载体类型中,腺相关病毒(aav)衍生的载体似乎最有希望。aav载体具有大量优点:(i)它们能够感染种类广泛的细胞类型,包括肌纤维;(ii)因为它们缺少所有病毒基因,并且野生型病毒还没有与人类中的任何疾病相关联,因此它们显得安全;(iii)与整合到宿主细胞的基因组中的野生型aav相反,复制缺陷型aav载体一般保持为游离体,从而限制了插入诱变或激活癌基因的风险;并且(iv)与其他载体系统相反,aav载体不引发显著的免疫应答,因此允许长期表达治疗性转基因(只要它们的基因产物不被排斥)。aav载体也能够以高滴度生产,并且强制性动脉内注射使它们能够通过单次注射实现向重要肌肉区域的基因转移,至少在啮齿动物中如此。尽管aav载体缺少所有病毒基因,但它们的装载量限于4.5kb。因此,选择aav导致开发了约4kb的μ-肌养蛋白变体来代替全长肌养蛋白(14kb)。通过转基因技术或基因转移,这些变体中的几种已经在mdx模型中得到了有利的检验。
已报道,在许多dmd患者以及在mdx小鼠和grmd狗中,肌养蛋白阳性纤维很少。尽管回复体纤维的比例随时间增加,但不幸的是它们的数量太低,不能产生显著的临床益处。引发这些回复体纤维的机制尚不清楚,尽管有研究表明阅读框可能通过外显子跳跃得以恢复。这种自然现象唤起了设计基因修复/调节策略的研究,其基于使用2’-o-甲基反义寡核糖核苷酸以及吗啉干扰剪接,从而诱导外显子跳跃。事实上,这种方法已在体外成功用于mdx、grmd和dmd肌肉细胞中,并在体内成功使用(对2’-o-甲基的1期临床试验在荷兰获得成功,使用吗啉的1期正在英国进行)。然而,这种方法的弱点在于它需要定期给药合成的ao,并且还没有完全实现全身递送。
备选的方法是从载体原位合成作为反义rna分子的目标序列。即使如此,在体内生产“治疗性”反义rna分子仍提出许多问题,例如稳定性和亚细胞定位。已知参与剪接反应的小核rna(snrna)可用作载体以克服这些限制。最近的报道显示,u7snrna携带针对肌养蛋白基因的外显子23或外显子51的剪接接点处的反义序列,在体外以及在体内分别在mdx和δ48-50dmd细胞中诱导了肌养蛋白合成。
已经对具有框外缺失的所有dmd患者进行了计算机搜索,所述患者理论上将从与缺失相邻(任一侧)的单个外显子的跳跃中获益。有趣的是,据预测,跳过外显子51将在22%的病例中恢复微小肌养蛋白(即δ45-50、δ47-50、δ48-50、δ49-50、δ50和δ52)。预期得到的截短蛋白至少有部分功能,因为它们所对应的缺失已在一些bmd患者中发现。此外,鉴定到一些带有δ51-52和δ48-51框内缺失的健康男性。在所选患者中,跳过外显子51将导致产生有功能的较短肌养蛋白,从而改善表型。
智力迟钝是经常与dmd相伴的症状,可能是由于神经元细胞中缺少肌养蛋白。因此,在脑中挽救半功能性肌养蛋白能够矫正或改善认知损害。
脊髓性肌萎缩(sma)总的来说是指各种源自于运动神经元生存(smn)基因中的常见遗传缺陷的病患,所述基因在1990年被作图于染色体5q11.2-13.3。人类5号染色体含有大量重复,因此存在两个smn基因拷贝smn1和smn2。
sma是遗传决定的新生儿死亡的最常见原因。smn相关sma的所有形式的合并发病率约为每6000人中1人。基因频率约为1:80,几乎40人中有1人是携带者。作为携带者没有已知的健康后果。人们可能知道的考虑这种可能性的唯一方式是是否有亲属患病。
sma的特征是脊髓和脑干运动神经元的丧失。一般来说,症状出现越早,预期寿命越短。一旦症状出现后,运动神经元细胞快速退化。所有形式的sma具有由去神经支配所引起的共同的虚弱症、即肌肉萎缩,因为肌肉由于支配性神经的丧失而失去收缩信号。脊髓性肌萎缩只影响运动神经。引起由运动神经元去神经支配造成的虚弱症和由感觉神经元去神经支配造成的感觉损害这两种可遗传的病症,总称为charcot-marie-tooth或遗传性运动感觉神经病来称呼。
sma的过程与虚弱的严重性直接相关。患有重型sma的婴儿常常由于支持呼吸的肌肉的虚弱而死于呼吸疾病。患有较轻型sma的儿童自然活得长得多,尽管他们可能需要大量医疗支持,特别是处于该病谱的更严重一端的儿童。
i型sma、也称为严重婴儿期sma或werdnighoffmann病,是最严重的,并在生命的第一年中就显现。这种类型一般在出生后很快并且出人意料地发作;被诊断患有i型sma的婴儿一般不会活过一岁。由于运动神经元生存的退化,肺炎被认为是最终死因;运动神经元死亡引起主要身体器官系统、特别是呼吸系统(例如肺内合并分泌物的呼出和除去)功能不足。ii型sma或中间型sma,描述那些永远不能站立和行走、但是至少在他们生命中的某些时间能够维持坐姿的儿童。虚弱的发生通常在6至18个月之间的某个时间被意识到。在个体的整个生命期间,虚弱缓慢逐渐地增加。iii型sma患者有时能够行走。
sma典型使用运动神经元生存(smn)基因测试来诊断,这种测试确定是否存在至少一个有功能的smn1基因的拷贝,所述基因通过在完全加工过的mrna中外显子7和8的存在,与高度相似的smn2基因区分开。smn2基因也含有使其在制造蛋白时效率变降低的突变,但是这种效率变降低的水平低下。sma由smn1基因从两条染色体中缺失并且smn2蛋白不能补偿功能性smn1蛋白的缺失所引起。
目前开发sma疗法的策略包括鉴定增加smn2水平、增强残留smn2功能或以其它方式补偿smn1活性丧失的药物。药物例如丁酸盐、丙戊酸、羟基脲和利鲁唑(
肌强直性营养不良(dm)是一种慢性、渐进性、高度可变的遗传性多系统疾病,其可以在从出生到老年的任何年龄时显现。肌强直性营养不良是最常见形式的成年发作性肌营养不良,是仅次于杜氏肌营养不良的第二常见的骨骼肌疾病形式。dm的特征是肌肉消耗(肌营养不良)、后囊下虹彩白内障(眼晶状体混浊)、心脏传导缺陷、内分泌改变和肌强直(肌肉难以松弛)。
目前存在两种已知类型的成年发作性dm,二者都可通过dna分析鉴定:1型肌强直性营养不良(dm1)通常被称为steinert病,其具有可能严重影响婴儿的先天性形式和儿童期发作形式。2型肌强直性营养不良(dm2)被称为promm或近端肌强直性肌病。还怀疑其他的肌强直性营养不良(例如dm3、dm4、dmx),但是它们的存在仍未被证明。虽然dm1和dm2都被认为是缓慢退化性病症,但认为dm2一般比dm1轻。
症状的表现随着形式(dm1/dm2)、严重性乃至不常见的dm2表型而有相当大的变化。dm1患者常表现出肌强直、不能性远端虚弱和严重的认知困难。dm2患者通常表现出肌肉疼痛、僵直、疲劳或发生近端下肢虚弱。day等,neurology60(4):657–64(2003)。虚弱的特征性模式对于dm1和dm2来说是不同的。在dm1中,在面部和下颚肌肉中观察到,眼睑下垂(下垂症),颈部肌肉、手和小腿虚弱。在dm2中,虚弱在那些更接近身体躯干的近端肌肉、颈部、肩部、髋屈肌和大腿中更加明显。
dm1症状包括睡眠过度(白天嗜睡)、肌肉消耗、吞咽困难和呼吸不足。dm1患者可能比dm2患者经历范围更分散的认知困难。取决于他们所患疾病的形式和严重性程度,dm1认知困难的范围可以包括发育迟缓、学习困难、语言、讲话、行为、淡漠或睡眠过度。dm2的认知表现包括执行功能(即组织、凝练、唤词等)问题和睡眠过度。
在dm1中,受影响的基因被称为dmpk(肌强直性营养不良蛋白激酶),并编码在骨骼肌中表达的丝氨酸/苏氨酸蛋白激酶。该基因位于19号染色体的长臂上。在dm1中,dmpk基因的特征为胞嘧啶-胸腺嘧啶-鸟嘌呤(ctg)的三联体重复序列。重复的数量在不同人中有很大变化,但是在健康人中平均数在5至37之间。有时,当dna的重复序列在细胞分裂期间被修复或复制时,细胞机制滑移,三联体重复序列的额外拷贝被添加到序列中。一旦dmpk基因中存在超过37个三联体重复序列,序列变得不稳定并且滑移现象变得更加常见。
患有dm1的人具有超过50个、并可能具有多达2000个ctg重复序列。结果是由于配子发生过程中的滑移,患有dm1的个体的重复序列大小通常在配子发生或早期胚胎发育期间将变得更大,使得患病成年人的孩子典型地表现出比他们的双亲更大的扩增(这种现象被称为早现遗传)。具有更大扩增的个体具有更早的疾病发作和更严重的表型。
dm2同样由染色体3q21上的znf9基因缺陷引起。dm2的重复序列扩增远大于dm1,在75至超过11,000个重复序列的范围内,并且包含4个核苷酸的重复序列。然而,与dm1不同,重复dna扩增的大小没有显示出造成dm2发作年龄或疾病严重性的差异。早现遗传在dm2中表现得不太显著。
目前不存在能够治愈或特异性针对肌强直性营养不良的疗法。与病症相伴的心脏问题、白内障和其他异常情况可以治疗但不能治愈。然而,存在可以缓解一些症状例如肌强直、疼痛和过度嗜睡的医学干预和药物。在例如高通量筛选和反义治疗领域中进行的研究,为将来更有效的定向治疗保留了希望。肌肉特异性氯通道1(clc-1)的剪接改变引起dm1的肌强直表型,并且在小鼠模型中可以使用修改clc-1mrna剪接的吗啉代反义寡核苷酸来逆转。wheeler等,j.clin.invest.117(12):3952-7(2007)。
尽管对于杜氏肌营养不良、脊髓性肌萎缩和steinert肌强直性营养不良的治疗模式正在进行研究,但对治疗这些疾病的有效的化合物和治疗方法,仍存在着迫切需求。
技术实现要素:
本公开通过提供三环dna(tc-dna)反义寡核苷酸(aon)以及使用tc-dnaaon治疗疾病例如杜氏肌营养不良、脊髓性肌萎缩和steinert肌强直性营养不良的方法,满足了这些以及其他相关的需求。
总的来说,本发明还涉及在对象的中枢神经系统细胞中矫正异常基因表达的方法,所述方法包含向对象给药tc-dna反义寡核苷酸,其中所述tc-dna反义寡核苷酸与所述基因编码的rna的一部分互补,并且其中tc-dna反义寡核苷酸以足以矫正所述异常表达的量外周给药于对象。
本发明还涉及治疗由对象中枢神经系统中的异常基因表达引起的遗传疾病的方法,所述方法包含向对象给药tc-dna反义寡核苷酸,其中所述tc-dna反义寡核苷酸与所述基因编码的rna的一部分互补,并且其中所述tc-dna反义寡核苷酸以有效矫正所述异常表达的量外周给药于对象。
本发明还涉及包含tc-dna反义寡核苷酸的药物组合物,其中所述tc-dna反义寡核苷酸与人类基因编码的rna的一部分互补,并且其中所述组合物还包含药用赋形剂。
本发明还涉及tc-dna反义寡核苷酸,其用于治疗由对象中枢神经系统中的异常基因表达引起的遗传疾病,所述tc-dna反义寡核苷酸与所述基因编码的rna的一部分互补,并且所述tc-dna反义寡核苷酸以有效矫正所述异常表达的量外周给药于对象。
当在本文中使用时,术语“外周给药”包括但不限于任何没有暗示直接注射到需要治疗的对象的中枢神经系统中的给药途径。更具体来说,外周给药包括全身注射,例如肌肉内(i.m.)、静脉内(i.v.)、腹膜内(i.p.)、动脉内、皮下或透皮注射。
本发明还涉及用于治疗神经肌肉或肌肉骨骼疾病的tc-dna反义寡核苷酸。所使用的tc-dna反义寡核苷酸如本文下面所进一步详细描述的。更具体来说,tc-dna反义寡核苷酸可以是本文中提出的具体tc-dna之一。
神经肌肉或肌肉骨骼疾病可以源自于基因的改变,其中所述改变是
外显子的框内突变,破坏基因的翻译阅读框的突变,
可以通过在由所述基因编码的mrna中包含非典型外显子加以补偿的有害突变,并且所述tc-dna与所述基因编码的pre-mrna中存在的iss或tsl互补并促进非典型外显子的包含,或者
导致在所述基因编码的mrna中出现有害的3'cug扩增序列的突变。
在具体实施方案中,当改变是外显子的框内突变时,所述tc-dna能够促进所述外显子的跳跃。在另一个实施方案中,当改变是破坏基因的翻译阅读框的突变时,所述tc-dna能够促进外显子的跳跃以便恢复基因的阅读框。在另一个实施方案中,当改变是导致在所述基因编码的mrna中出现有害的3'cug扩增序列的突变时,所述tc-dnaaon能够破坏含有所述扩增序列的mrna。
本文中提出的tc-dnaaon是受限的dnaaon,其与利用例如更常规的2’-o-甲基-硫代磷酸酯或吗啉代化学品的dnaaon相比,显示出与互补的pre-mrna的增加的杂交性质。尽管2’-o-甲基-硫代磷酸酯或吗啉代dnaaon典型地需要20至24个核苷酸来实现与pre-mrna靶的特异性杂交,但本文公开的tc-dnaaon能够与长度在10至18个核苷酸之间、更宽泛在约6至约22个核苷酸之间、尤其是8至20个核苷酸之间的pre-mrna靶特异性杂交。
正如下文更详细描述的,外显子跳跃在pre-mrna的成熟加工过程中在核中实现。它包括使用与pre-mrna内外显子限定序列互补的反义寡核苷酸(aon)来掩蔽参与靶外显子的剪接的关键序列。本文提供的tc-dnaaon,通过掩蔽肌养蛋白pre-mrna内内含子/外显子接点处的剪接位点、或者更普遍来说用于外显子限定的位点,从而促进在pre-mrna向成熟mrna的加工过程中删除有害外显子,因而可以适用于外显子跳跃。这样的tc-dnaaon,通过在包含含有无义、终止、移码突变的外显子或含有有害隐蔽外显子的内含子序列的突变肌养蛋白基因中恢复开放阅读框,将在杜氏肌营养不良的治疗中发现用途。
例如,肌养蛋白基因的外显子23或外显子51中的无义或移码突变,产生了羧基端截短的无功能肌养蛋白。通过分别与内含子23或内含子51中包含肌养蛋白pre-mrna剪接供体位点的核苷酸、以及外显子23或外显子51的相邻5’核苷酸杂交,本文公开的tc-dnaaon能够阻止将突变的外显子23或外显子51包含在成熟的mrna转录本中。该成熟mrna转录本的表达产生有功能的肌养蛋白,其缺失由外显子23或外显子51编码的氨基酸,但是包含那些缺失的氨基酸的n-端和c-端两侧的肌养蛋白氨基酸,因此构成了半功能性的“准肌养蛋白”。
本文公开的用于在肌养蛋白pre-mrna加工期间跳过外显子的tc-dnaaon,含有约6至约22个核苷酸、特别是约8至20个三环核苷酸、特别是约10至18个三环核苷酸,其中tc-dnaaon的8-16个核苷酸与肌养蛋白pre-mrna的内含子剪接供体位点互补,其中tc-dnaaon的2-8个核苷酸与肌养蛋白pre-mrna的外显子区互补,并且其中内含子剪接供体位点与外显子区邻接并位于其5’端。在某些情况中,tc-dnaaon具有12至16个核苷酸或13至15个核苷酸,并包含与内含子剪接供体位点互补的6至14个核苷酸和与外显子区互补的2至5个核苷酸。然而,应该理解,更长的tc-dnaaon可以适用于在肌养蛋白pre-mrna的加工期间实现外显子跳跃。
本文中示例了被设计用于跳过肌养蛋白pre-mrna内突变的外显子23的tc-dnaaon。所述tc-dnaaon包含核苷酸序列5’-aacctcggcttacct-3’(m23d(+02-13),seqidno:1),并与位于肌养蛋白pre-mrna内含子23的3’末端的核苷酸和位于肌养蛋白pre-mrna外显子23的邻接5’末端的核苷酸特异性杂交。还提供了被设计用于跳过肌养蛋白pre-mrna内突变的外显子51的tc-dnaaon。所述tc-dnaaon包含选自5’-agaaatgccatcttc-3’(h51(+68+82),seqidno:2)、5’-aaatgccatcttcct-3’(h51(+70+84),seqidno:3)和5’-tgccatcttccttga-3’(h51(+73+87),seqidno:4)的核苷酸序列,并与位于肌养蛋白pre-mrna内含子51的3’末端的核苷酸和位于肌养蛋白pre-mrna外显子51的5’末端的核苷酸特异性杂交。
在本文中使用了下列命名法:“m”是指小鼠,“h”是指人类,“23”和“51”是指特定外显子,“d”是指供体位点,“a”是指受体位点,跟有数值的“+”表示外显子序列中的核苷酸数量,跟有数值的“-”表示侧翼外显子中的核苷酸数量。因此,例如m23d(+02-13)表示tc-dnaaon涵盖了外显子23的两个3’-端核苷酸和内含子23的13个5’-端核苷酸,所述aon能够掩蔽小鼠肌养蛋白外显子23的供体剪接位点,而h51(+68+82)表示tc-dnaaon跨越人类肌养蛋白外显子51中的从68号核苷酸至82号核苷酸。
本公开的其他方面提供了可以适用于掩蔽运动神经元生存蛋白2(smn2)pre-mrna内的内含子沉默序列(iss)或末端茎-环(tsl)的tc-dnaaon。这样的tc-dnaaon促进在smn2pre-mrna加工成成熟mrna过程中包含非典型外显子。得到的修饰的功能性smn2蛋白含有由被包含的非典型外显子编码的氨基酸序列。这种修饰的功能性smn2蛋白能够补充无功能的smn1蛋白,并且当在体内表达时,能够至少部分逆转由smn1基因中的突变引起的脊髓性肌萎缩。
例如,尽管smn2的外显子7典型通过相应pre-mrna的加工被排除在成熟mrna转录本之外,但加入外显子7产生了修饰的功能性smn2蛋白,其能够在功能上补偿突变的smn1蛋白。通过与smn2pre-mrna内包含smn2iss或tsl的核苷酸杂交,tc-dnaaon能够促进将外显子7包含在成熟mrna转录本中。该成熟mrna转录本的表达产生修饰的功能性smn2蛋白,其包含由外显子7编码的氨基酸以及位于所包含的氨基酸的n-端和c-端两侧的所有其他smn2氨基酸。
因此,本公开提供了在smn2pre-mrna加工期间促进包含外显子7的tc-dnaaon,其中所述tc-dnaaon长度为6-22个三环核苷酸、特别是8-20个三环核苷酸、更特别长度为10-18个三环核苷酸,并且其中所述tc-dnaaon与smn2pre-mrna的内含子沉默序列(iss)或末端茎-环(tsl)互补。这样的tc-dnaaon可以具有13至17个核苷酸、12至16个核苷酸或13至15个核苷酸。本文中示例了包含15个核苷酸的序列5’-cuuucauaaugcugg-3’(smn2i7(10;25),seqidno:5)的tc-dnaaon,所述tc-dnaaon与smn2pre-mrna的iss互补,并且可用于促进将外显子7包含在加工过的smn2mrna中。本文中还示例了包含13个核苷酸的序列5’-uuaauuuaaggaa-3’(smn2e7(39;51),seqidno:6)的tc-dnaaon,所述tc-dnaaon与smn2pre-mrna的tsl2互补,并且也可用于促进将外显子7包含在加工过的smn2mrna中。应该理解,也可以使用本文提出的tc-dnaaon的组合。
本公开的其它方面提供了可以适用于促进突变dm1mrna的破坏的tc-dnaaon。这样的tc-dnaaon包含9-27个三环核苷酸,其中tc-dnaaon与包含有害3’cug扩增序列(n>50)的突变dm1mrna互补,并且其中所述tc-dnaaon能够促进所述dm1mrna的rnase介导的破坏。tc-dnaaon可以包含3至9个、4至8个或5、6或7个连续重复的核苷酸序列5’-cag-3’(seqidno:7)。用于促进突变dm1的破坏的示例性tc-dnaaon包含15个核苷酸的序列5’-cagcagcagcagcag-3’(dm1(cag5),seqidno:8)。用于促进突变dm1的破坏的另一个示例性tc-dnaaon包含21个核苷酸的序列5’-cagcagcagcagcagcagcag-3’(dm1(cag7),seqidno:9)。
在其它方面中,本公开提供了在细胞中用于从肌养蛋白mrna中消除突变外显子的方法、用于将非典型外显子包含在smn2mrna内的方法、以及用于破坏包含病理数量的3’cug扩增序列的dm1mrna的方法。这些方法的每一种都包括将细胞与本文中公开的tc-dnaaon进行接触的步骤。
在某些实施方案中提供了用于从肌养蛋白mrna中消除突变外显子的方法,所述方法包含将表达肌养蛋白pre-mrna的细胞与含有长度为6-22个三环核苷酸、特别是8-20个三环核苷酸、更特别是10至18个三环核苷酸的tc-dnaaon进行接触的步骤,其中所述tc-dnaaon的8-16个核苷酸与肌养蛋白pre-mrna的内含子剪接供体位点互补,其中所述tc-dnaaon的2-8个核苷酸与肌养蛋白pre-mrna的外显子区互补,并且其中外显子区与内含子剪接供体位点邻接并位于其3’端。示例性方法包括将细胞与12至16个核苷酸或13至15个核苷酸的tc-dnaaon进行接触的步骤。适合用于这种方法的tc-dnaaon包含核苷酸序列5’-aacctcggcttacct-3’(m23d(+02-13),seqidno:1)、5’-agaaatgccatcttc-3’(h51(+68+82),seqidno:2)、5’-aaatgccatcttcct-3’(h51(+70+84),seqidno:3)和5’-tgccatcttccttga-3’(h51(+73+87),seqidno:4)。
在其他实施方案中提供了用于将非典型外显子包含在smn2mrna内的方法,所述方法包含将表达smn2pre-mrna的细胞与含有长度为6-22个三环核苷酸、特别是8-20个三环核苷酸、更特别是10至18或11至18个三环核苷酸的tc-dnaaon进行接触的步骤,其中所述tc-dnaaon与smn2pre-mrna的内含子沉默序列(iss)、例如内含子7内的iss-n1互补。示例性方法包括将细胞与12至16个核苷酸或13至15个核苷酸的tc-dnaaon进行接触的步骤。适合用于这种方法的tc-dnaaon包含15个核苷酸的序列5’-cuuucauaaugcugg-3’(smn2i7(10;25),seqidno:5)。在相关的方法中,tc-dnaaon与smn2pre-mrna的末端茎-环(tsl)、例如外显子7内的tsl-2互补。适合用于这种方法的tc-dnaaon包含13个核苷酸的序列5’-uuaauuuaaggaa-3’(smn2e7(39;51),seqidno:6)。
在另外的其他实施方案中提供了用于破坏细胞中包含一个或多个3’cug扩增序列的dm1mrna的方法,所述方法包含将细胞与包含9-27个三环核苷酸的tc-dnaaon进行接触的步骤,其中所述tc-dnaaon与包含一个或多个3’cug扩增序列的突变dm1mrna互补,并且其中所述tc-dnaaon能够促进dm1mrna的rnase介导的破坏。适合用于这种方法的tc-dnaaon包含3至9个、4至8个或5、6、7个连续重复的核苷酸序列5’-cag-3’(seqidno:7),其实例是包含15个核苷酸的序列5’-cagcagcagcagcag-3’(dm1(cag5),seqidno:8)的tc-dnaaon。促进突变dm1的破坏的另一个示例性tc-dnaaon包含21个核苷酸的序列5’-cagcagcagcagcagcagcag-3’(dm1(cag7),seqidno:9)。
在其它方面中,本公开提供了用于治疗杜氏肌营养不良(dmd)的方法、用于治疗脊髓性肌萎缩(sma)的方法和用于治疗steinert肌强直性营养不良(sd)的方法。这些方法的每一种都使用了向患者给药本文中公开的tc-dnaaon的步骤,分别用于从肌养蛋白mrna中消除突变的外显子、用于将非典型外显子包含在smn2mrna内或用于破坏包含一个或多个3’cug扩增序列的dm1mrna。
从下文中提出的详细描述和权利要求书,本公开的这些以及其他实施方案、特点和优点将变得明显。
附图说明
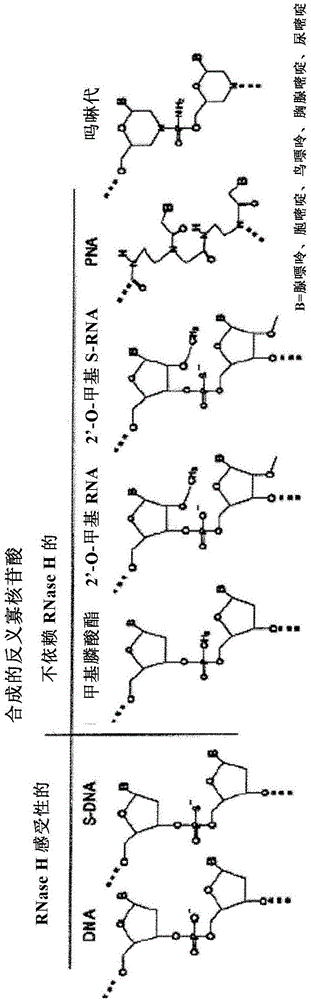
图1是用于生产反义寡核苷酸的各种合成核苷酸的表。
图2是三环dna(tc-dna)的结构图。
图3的图显示了mdx小鼠在肌养蛋白基因的外显子23中携带无义突变,其阻止合成有功能的肌养蛋白。外显子23部分编码重复序列r6和r7,其中c到t的突变产生了终止密码子(taa)。用于在外显子23的下游供体剪接位点处进行外显子跳跃的15个核苷酸的tc-dnaaon被命名为m23d(+02-13),具有核苷酸序列5’-aacctcggcttacct-3’(seqidno:1),并与靶肌养蛋白pre-mrna的外显子23/内含子23接点互补,所述接点由序列5’-外显子23...tcaggtaagccgaggtttggcc…内含子23-3’(seqidno:2)限定,其中大写字母表示外显子的核苷酸,小写字母表示内含子的核苷酸。
图4是巢式rt-pcr反应的琼脂糖凝胶,显示了在利用或不利用oligofectamine下用1、2和10μgtc-dnaaonm23d(+02-13)转染的mdx肌管中肌养蛋白pre-mrna的外显子23的跳过。在48小时后,收获培养物并进行处理以提取mrna。
图5是巢式rt-pcr反应的琼脂糖凝胶,显示了在oligofectamine存在下用0.5、1、2、5和10μgtc-dnaaonm23d(+02-13)转染的mdx肌管中肌养蛋白pre-mrna的外显子23的跳过。在48小时后,收获培养物并进行处理以提取mrna。
图6是巢式rt-pcr反应的琼脂糖凝胶,显示了在oligofectamine存在下用5μgtc-dnaaonm23d(+02-13)转染的mdx肌管中肌养蛋白pre-mrna的外显子23的跳过。在转染后从第0日至第15日收获培养物并进行处理以提取mrna。
图7是巢式rt-pcr反应的琼脂糖凝胶,显示了在用含有100、80、40、20、10和5μgtc-dnaaonm23d(+02-13)的50μlpbs(磷酸盐缓冲盐水)进行胫前肌注射的8周龄mdx小鼠中,肌养蛋白pre-mrna的外显子23的跳过。在3周后处死动物并对肌肉样品进行处理用于mrna分析。
图8是rt-pcr反应的琼脂糖凝胶,显示了在用含有10μgtc-dnaaonm23d(+02-13)的50μlpbs(磷酸盐缓冲盐水)进行胫前肌注射的8周龄mdx小鼠中,肌养蛋白pre-mrna的外显子23的跳过。在4、10和20周后处死动物并对肌肉样品进行处理用于mrna分析。
图9是来自在8周龄时用含有10μgtc-dnaaonm23d(+02-13)的50μlpbs(磷酸盐缓冲盐水)注射的mdx小鼠的胫前肌组织横切片中,肌养蛋白的免疫染色的光学显微照片。在4、10和20周后处死动物并对肌肉样品进行处理用于免疫染色。
图10是用tc-dnaaonm23d(+02-13)注射到海马或脑脊液中的正常和mdx小鼠的cns中肌养蛋白的免疫染色。a-b-c切片是单次鞘内注射后1个月时,正常、mdx和用20μgtc-dnam23d(+02-13)处理的mdx的海马水平处。d-e-f切片是在脑脊液中递送后1个月时,正常、mdx和用200μgtc-dnam23d(+02-13)处理的mdx的小脑水平处。
图11是pre-mrna到成熟mrna的总体加工的示意图。
图12是巢式rt-pcr反应的琼脂糖凝胶,显示了在用含有各种tc-dnaaon的50μlpbs(磷酸盐缓冲盐水)进行胫前肌注射的8至10周龄hdmd小鼠中,肌养蛋白pre-mrna的外显子51的跳过。
图13是smn1和smn2基因的内含子-外显子结构和染色体位置的示意图。
图14是smn2中的点突变(c6t)的示意图,所述smn2明显缺少外显子7,影响了mrna剪接。
图15是通过改进内含子6-外显子7边界处的剪接受体(“sa”)7和外显子7-内含子7边界处的剪接供体(sd)7的使用,来增加外显子7在smn2中的包含的示意图。
图16是smn1和smn2中的外显子7的结构示意图。
图17是tc-dnaaonsmn2e7(39;51)(seqidno:6)的靶序列及其对smn1和smn2外显子7结构的推断作用的示意图。tc-dnaaonsmn2e7(39;51)具有序列5’-uuaauuuaaggaaugug-3’,可能破坏smn2中末端茎环2的结构,从而增加了外显子7在smn1和smn2中的包含。
图18是tc-dnaaonsmn2i7(10;25)(seqidno:5)的靶序列及其对smn1和smn2外显子7包含的推断作用的示意图。tc-dnaaonsmn2i7(10;25)具有序列5’-cacuuucauaaugcugg-3’,可能阻止内含子沉默序列(“iss”)-n1的识别,允许识别外显子7-内含子7边界处的5’剪接位点。exinct是指扩展抑制背景(extendedinhibitorycontext)。基于扩展突变分析,已显示出c6u产生了影响外显子7限定的扩展抑制背景。
图19rt-pcr反应的琼脂糖凝胶(图19b)和归一化图(图19a),显示了在来自sma患者的成纤维细胞(g03813细胞系)中外显子7在smn2中的包含。48小时后,收获培养物并进行处理以提取mrna。实线对应于模拟物处理的对照细胞,不连续线对应于tc-dnaaonsmn2i7(10;25)(被称为tc-i7)处理的细胞,点线对应于tc-dnaaonsmn2e7(39;51)(被称为tc-tsl)处理的细胞。图已经按照每道中smn1+smn2(全长)+smn2(δ7)的总量进行归一化。图19c是western印迹图,显示了用标明的tc-dna寡核苷酸转染的g03813细胞中smn的水平。显示了肌动蛋白作为上样对照。值得注意的是tc-dnaaonsmn2i7(10;25)(被称为tc-iss7)和tc-dnaaonsmn2e7(39;51)(被称为tc-tsl)对smn2生产的累加效应。图19d是光学显微照片,显示了smn在tc-tsl处理的细胞中的核定位(暗点)。核用dapi复染。
图20(图20a)是northern印迹图,显示了在体外,随着转染到dm1成肌细胞中的tc-dnaaondm1(cag7)(被称为tc-dna(cag)7)的量的增加,突变的人类dmpkmrna的水平降低。在3天后收获培养物并进行处理以提取mrna并进行northern印迹分析。图20b是反映了突变体dmpk与正常dmpkmrna的比率的定量图。
图21(图21a)是northern印迹图,显示了随着注射到表达具有700个cug重复序列的人类dmpkmrna的dm1小鼠的ta肌肉中的具有序列5’-cagcagcagcagcagcagcag-3’(seqidno:9)的tc-dnaaondm1(cag7)(被称为tc-dna(cag)7)的量的增加,突变的人类dmpkmrna的水平降低。图21c是northern印迹图,显示了当30或60μgtc-dnaaondm1(cag7)被注射到dm1小鼠(n=4)的ta肌肉中时,突变的人类dmpkmrna的水平降低。图21b和21d分别是反映了图21a和21c的northern印迹中突变的人类dmpk与小鼠dmpkmrna的比率的定量图。
具体实施方式
本公开是基于意外发现三环dna(tc-dna)反义寡核苷酸(aon)可以适用于掩蔽肌养蛋白基因内的pre-mrna剪接位点,用于掩蔽smn2基因内的内含子沉默序列或末端茎-环序列,或用于破坏包含一个或多个3’cug扩增序列的dm1mrna。这些发现将在总的来说遗传疾病的治疗中,更具体来说在杜氏肌营养不良、脊髓性肌萎缩和steinert肌强直性营养不良的治疗中发现广泛应用。
三环dna(tc-dna)属于一类新的受限dna类似物,其表现出与互补rna的改进的杂交能力。ittig等,nucleicacidsres.32:346-353(2004);ittig等,prague,academyofsciencesoftheczechrepublic.7:21-26(coll.symp.series,hocec,m.,2005);ivanova等,oligonucleotides17:54-65(2007);renneberg等,nucleicacidsres.30:2751-2757(2002);renneberg等,chembiochem.5:1114-1118(2004);以及renneberg等,jacs.124:5993-6002(2002)。本文公开的pre-mrna/tc-dnaaon异源双链体对rnaseh具有抗性,并由此阻止所靶向的pre-mrna被破坏。三环dna化学品的优点在于其骨架的结构性质允许减小aon的长度并同时保留与互补核苷酸序列的高亲和性和高特异性杂交。出人意料的是,利用肌肉内施用,tc-dnaaon可以有利地以微克剂量用于体内环境,这比常规反义寡核苷酸技术所需的剂量低至少10倍。此外,tc-dna在减小反义寡核苷酸长度的同时保留了全部活性。因此,例如,在本公开所示例的离体和体内应用中,13至15个核苷酸的tc-dnaaon非常高效。
本文描述的tc-dnaaon与现有的反义寡核苷酸化学品例如2’-o-甲基-硫代磷酸酯或吗啉代化学品相比,还表现出增加的体内稳定性。因此,例如,单次肌肉内注射本公开的tc-dnaaon,能在给药后保留体内效能20周以上。
此外以及非常意外的是,本公开的tc-dnaaon,以被称为m23d(+02-13)的tc-dnaaon为例,可以通过脑实质内或脑室内给药或通过给药到蛛网膜下腔内来递送到中枢神经系统(cns)中,以恢复海马ca1的神经元内或大脑或小脑皮层的神经元内突变的基因,例如突变的肌养蛋白基因。因此,这证实了本文描述的tc-dnaaon能够有效跨过室管膜屏障。
通过参考下面的定义,将最好地理解本公开:
定义
当在本文中使用时,术语“三环dna(tc-dna)”是指一类受限的dna类似物,其中通过引入环丙烷环对每个核苷酸进行修饰,以限制骨架的构象灵活性并优化骨架的扭转角γ的几何学。含有高腺嘌呤和胸腺嘧啶碱基的tc-dna与互补rna形成超常稳定的a-t碱基对。
当在本文中使用时,术语“反义寡核苷酸(aon)”是指能够与pre-mrna或具有互补核苷酸序列的mrna相互作用和/或杂交,由此修改基因表达的寡核苷酸。酶依赖性反义寡核苷酸包括依赖于rnaseh活性来降解靶mrna的形式,并包括单链dna、rna和硫代磷酸酯反义寡核苷酸。空间阻断型(stericblocking)反义寡核苷酸(rnase-h不依赖性反义寡核苷酸)通过与mrna的靶序列结合来干扰基因表达或其他mrna依赖性细胞过程。空间阻断型反义寡核苷酸包括2'-o-烷基反义寡核苷酸、吗啉代反义寡核苷酸和三环dna反义寡核苷酸。正如本文中所述,在某些应用中,可以将tc-dna反义寡核苷酸应用在酶依赖性应用中,例如rnase介导性破坏包含一个或多个3’cug扩增序列的dm1mrna。
当在本文中使用时,“互补”是指核酸分子能够与另一个核酸分子通过互补的核苷或核苷酸之间的常规watson-crick碱基配对或其他非常规类型的配对(例如hoogsteen或反向hoogsteen氢键形成)而形成氢键。对于本公开的tc-dnaaon来说,tc-dnaaon与其互补序列的结合自由能足以允许tc-dnaaon执行相关功能,并具有足够的互补性程度以避免tc-dnaaon与非靶序列在需要特异性结合的条件下、即离体或体内治疗情形中的生理条件下发生非特异性结合。核酸分子结合自由能的确定在本技术领域中是公知的(参见例如turner等,cshsymp.quant.biol.lii:123-133(1987);frier等,proc.nat.acad.sci.usa83:9373-77(1986);以及turner等,j.am.chem.soc.109:3783-3785(1987))。因此,“互补”(或“可特异性杂交”)这种术语是表明存在足够程度的互补性或精确配对,使得在tc-dnaaon与pre-mrna或mrna靶之间能够发生稳定和特异结合。
在本技术领域中应该理解,核酸分子不是必需与靶核酸序列100%互补才能特异性杂交。也就是说,两个或多个核酸分子可以低于完全互补。互补性由核酸分子中能够与第二个核酸分子形成氢键的毗连残基的百分率表示。例如,如果第一个核酸分子具有10个核苷酸并且第二个核酸分子具有10个核苷酸,那么第一与第二个核酸分子之间5、6、7、8、9或10个核苷酸发生碱基配对分别表示50%、60%、70%、80%、90%和100%的互补性。“完全”或“全部”互补的核酸分子是指其中第一个核酸分子的所有毗连残基都将与第二个核酸分子中相同数量的毗连残基形成氢键的核酸分子,其中两个核酸分子具有相同数量的核苷酸(即具有相同长度),或者两个分子具有不同长度。
当在本文中使用时,术语“前体mrna”或“pre-mrna”是指含有一个或多个居间序列(内含子)的信使核糖核酸(mrna)的未成熟的单链。pre-mrna由rna聚合酶从细胞核中的dna模板转录而来,并由内含子和编码区(外显子)的交替序列构成。一旦pre-mrna通过剪掉内含子并连接外显子而被完全加工后,它被称为“信使rna”或“mrna”,其是完全由外显子构成的rna。真核生物的pre-mrna在被完全加工成mrna之前只短暂地存在。当pre-mrna已被适当加工成mrna序列后,它被输出到核外,并最终被细胞质中的核糖体翻译成蛋白质。
当在本文中使用时,术语“剪接”和“加工”是指pre-mrna在转录后的修饰,其中内含子被移除,外显子被连接。(参见图11)。剪接发生在由5个小核核糖核蛋白(snrnps)构成的被称为剪接体的大的rna-蛋白质复合体所催化的一系列反应中。在内含子中,3’剪接位点、5’剪接位点和分支位点为剪接所需。snrnp的rna组分与内含子相互作用,并可能参与催化。
pre-mrna剪接包含两个依次的生物化学反应。两个反应都涉及rna核苷酸之间的剪接体转酯作用。在第一个反应中,内含子内在剪接体装配期间限定的特定分支点核苷酸的2’-oh,对内含子5’剪接位点处的第一个核苷酸执行亲核攻击,形成套索中间体。在第二个反应中,释放出的5’外显子的3’-oh在内含子3’剪接位点处的最后一个核苷酸处执行亲核攻击,由此连接外显子并释放出内含子套索。pre-mrna剪接受到内含子沉默序列(iss)和末端茎环(tsl)序列的调控。
当在本文中使用时,术语“内含子沉默序列(iss)”和“末端茎环(tsl)”分别是指内含子和外显子内的序列元件,其通过结合反式作用蛋白因子控制pre-mrna内的可变剪接,由此导致剪接位点的不同用法。典型地,内含子沉默序列为8至16个核苷酸,并且与外显子-内含子接点处的剪接位点相比保守性较低。末端茎环序列典型为12至24个核苷酸,并由于互补性而在12-24个核苷酸的序列内形成二级环结构并因此结合。
当在本文中使用时,术语“脊髓性肌萎缩(sma)”是指不同临床类型的5号染色体连锁的sma,每种具有共同的遗传病因和由于脊髓与脑干的运动神经元丧失而造成的虚弱表现。脊髓性肌萎缩由运动神经元生存基因smn1内的突变引起。对于正常功能来说,需要smn1基因的至少一个正常等位基因。
5号染色体含有smn(运动神经元生存)基因的区域具有大的复制序列。含有几个基因的大序列在相邻区段中出现两次。因此smn1和smn2存在两个基因拷贝。smn2基因多一个突变,使得它制造蛋白的效率降低,但是这种效率降低的水平低。sma由两条染色体的smn1基因丧失造成。sma的严重性从sma1至sma3,与残留的smn2基因能够弥补smn1丧失的程度部分相关。经常还存在smn2的更多拷贝,并且smn2拷贝数的增加与疾病严重性较低有关。
“对象”是指作为外植细胞的供体或受体的生物体或细胞本身。“对象”也指可以给药本公开的核酸分子的生物体。在一个实施方案中,对象是哺乳动物或哺乳动物细胞。在另一个实施方案中,对象是人类或人类细胞。
当在本文中使用时,术语“治疗有效量”是指在其所给药的对象(例如人类)中足以治疗或预防所陈述的疾病、障碍或病症的tc-dnaaon的量。本公开的tc-dnaaon,可以单独或组合或与其他药物联合用于治疗本文中讨论的疾病或病症。例如,为了治疗特定疾病、障碍或病症,tc-dnaaon可以在适合于治疗的情况下单独或与一种或多种药物组合给药于患者,或可以给药于对于本技术领域的专业人员来说显而易见的其他适合细胞。
当在本文中使用时,词组“可药用”是指分子实体或组合物,当给药于人类时是生理上可耐受的,并且典型不产生过敏或类似的不利反应,例如胃不适、头晕等。优选,当在本文中使用时,术语“可药用”是指得到联邦或州政府的管理机构批准或被列于《美国药典》(u.s.pharmacopeia)或其他广泛认可的药典中,供用于动物、更具体为人类。
当在本文中使用时,术语“分离的”是指所提到的材料从其本源环境例如细胞中移出。因此,分离的生物材料可以不含一些或所有细胞组分,即天然存在本源材料的细胞中的组分(例如细胞质或膜组分)。
当在本文中使用时,术语“纯化的”是指材料已经在减少或消除无关材料的存在的条件下被分离,所述无关材料即污染物,包括从中获得材料的本源材料。例如,纯化的tc-dnaaon优选基本上不含细胞或培养组分,包括组织培养组分、污染物等。当在本文中使用时,术语“基本上不含”在操作上用在材料分析测试的情形中。优选,基本上不含污染物的纯化材料至少为50%纯;更优选至少90%纯,更加优选至少99%纯。纯度可以通过层析、凝胶电泳、免疫测定、组成分析、生物测定和本技术领域已知的其他方法来评估。
在本发明的说明书中,除非另有指明,否则任何浓度范围、百分率范围、比率范围或整数范围应被理解为包括所述范围内的任何整数值,以及在适合情况下的其分数(例如整数的十分之一和百分之一)。此外,除非另有指明,否则本文中所述的与任何物理特性例如聚合物亚基、大小或厚度相关的任何数字范围,应该被理解为包括所述范围内的任何整数。当在本文中使用时,除非另有指明,否则“约”或“基本上由……构成”是指所指范围、值或结构的±20%。
当在本文中使用时,术语“包括”和“包含”同义使用。应该理解,本文中没有数量指示的组分是指“一个或多个”所列举的组分。替换词(例如“或”)的使用应该被理解为是指替换项中的任一个、两者或其任何组合。
术语“约”或“近似于”是指在值的有统计学意义的范围之内。这样的范围可以在一定幅度级内,优选在给定的值或范围的50%以内、优选20%以内、更优选10%以内、更加优选5%以内。术语“约”或“近似于”所涵盖的容许偏差取决于所研究的具体系统,并可以被本技术领域的普通专业人员容易地认识到。
用于治疗杜氏肌营养不良的三环dna反义寡核苷酸
正如上面指出的,在某些实施方案中,本公开提供了可以适用于治疗杜氏肌营养不良(dmd)的tc-dnaaon,所述杜氏肌营养不良是一种严重的隐性x染色体连锁的肌营养不良形式,其特征为肌肉退化的快速发展,最终导致丧失步行能力、瘫痪和死亡。dmd由位于人类x染色体上的肌养蛋白基因内的突变例如无义或移码突变引起。肌养蛋白基因编码肌养蛋白,其是肌肉组织中的重要结构组分,为肌纤维肌膜以及位于细胞膜处的肌养蛋白聚糖复合体(dgc)提供结构稳定性。无义或移码突变引起翻译的过早终止,因此产生了c-端截短的肌养蛋白。
由一个或多个终止突变或框外突变引起的dmd,可以通过切除一个或几个外显子以便恢复翻译阅读框并由此恢复突变下游的mrna序列来缓解。为了实现这一点,作为本公开的一部分,开发了靶向pre-mrna内能够掩蔽一个或多个外显子的剪接体识别的区域的tc-dnaaon。通过用tc-dnaaon靶向这些区域,可以通过可变剪接移除外显子,以获得成熟的、功能性肌养蛋白mrna。
因此,本文描述的tc-dnaaon有效地促进肌养蛋白pre-mrna加工期间肌养蛋白基因中的一个或多个突变外显子的跳过,从而恢复所得到的肌养蛋白mrna的正确阅读框,其在翻译时产生功能性肌养蛋白。因此,本文公开的tc-dnaaon可治疗性用于患有dmd的患者。
当在本文中使用时,术语“外显子跳跃”是指通过用一种或多种互补的反义寡核苷酸(aon)靶向pre-mrna内的剪接供体和/或受体位点来修改pre-mrna剪接。通过阻断剪接体与一个或多个剪接供体或受体位点的接近,aon能够阻止剪接反应,由此引起一个或多个外显子从完全加工的mrna中缺失。外显子跳跃在pre-mrna成熟加工期间在核中实现。它包括使用与pre-mrna内的剪接供体序列互补的反义寡核苷酸(aon)来掩蔽参与被靶外显子剪接的关键序列。本文提供的tc-dnaaon,通过掩蔽肌养蛋白pre-mrna中内含子/外显子接点处的剪接位点,由此促进pre-mrna向成熟mrna的加工过程中突变外显子的缺失,可以适用于外显子跳跃。
例如肌养蛋白基因的外显子23或外显子50内的无义或移码突变产生羧基端截短的无功能肌养蛋白。通过与分别包含内含子23或内含子51中的肌养蛋白pre-mrna剪接供体位点的核苷酸、以及外显子23或外显子51的相邻5’核苷酸杂交,本文公开的tc-dnaaon能够阻止突变的外显子23或外显子51包含在成熟mrna转录本中。该成熟mrna转录本的表达产生功能性肌养蛋白,其缺失了由外显子23或外显子50和51编码的氨基酸,但是包括那些缺失氨基酸的n-端和c-端两侧的肌养蛋白氨基酸。
本文公开的用于在肌养蛋白pre-mrna加工期间跳过外显子的tc-dnaaon,典型地含有6-22个毗连的三环核苷酸、特别是8-20个三环核苷酸、特别是10-18个毗连的三环核苷酸,其中tc-dnaaon的6-16个核苷酸、特别是8-16个核苷酸与肌养蛋白pre-mrna的内含子剪接供体位点互补,其中tc-dnaaon的2-8个核苷酸与肌养蛋白pre-mrna的外显子区互补,并且其中内含子剪接供体位点与外显子区毗连并位于其5’端。取决于所设想的具体应用,tc-dnaaon可以具有12至16个核苷酸或13至15个核苷酸,并可以包含与内含子剪接供体位点互补的6至14个核苷酸和与外显子区互补的2至5个核苷酸。
本文中示例了被设计用于跳过肌养蛋白pre-mrna内突变的外显子23的tc-dnaaon。所述tc-dnaaon包含核苷酸序列5’-aacctcggcttacct-3’(m23d(+02-13),seqidno:1),并与位于肌养蛋白pre-mrna的内含子23的3’末端的核苷酸和位于肌养蛋白pre-mrna的外显子23的毗连5’末端的核苷酸特异性杂交。还提供了被设计用于跳过肌养蛋白pre-mrna内突变的外显子51的tc-dnaaon。所述tc-dnaaon包含选自5’-agaaatgccatcttc-3’(h51(+68+82),seqidno:2)、5’-aaatgccatcttcct-3’(h51(+70+84),seqidno:3)和5’-tgccatcttccttga-3’(h51(+73+87),seqidno:4)的核苷酸序列,并与位于肌养蛋白pre-mrna的内含子51的3’末端的核苷酸和位于肌养蛋白pre-mrna的外显子51的5’末端的核苷酸特异性杂交。
用于治疗脊髓性肌萎缩的三环dna反义寡核苷酸
在其他实施方案中,本公开提供了可以适用于治疗脊髓性肌萎缩(sma)的tc-dnaaon。sma由smn1基因的两个拷贝中的突变引起,所述基因在正常细胞中以在完全加工的mrna中存在外显子7和8为特征。因为正常加工的smn2mrna不含外显子7或8,因此smn2蛋白不能补偿功能性smn1蛋白的丧失。通过掩蔽smn2pre-mrna内的内含子沉默序列(iss)和/或末端茎环(tsl),本文描述的tc-dnaaon能够促进在加工后的smn2pre-mrna中包含非典型外显子7或外显子8,所述pre-mrna被翻译成修饰的功能性smn2蛋白,其能够补偿功能性smn1蛋白的丧失,并且当在体内表达时,修饰的功能性smn2能够至少部分逆转由smn1基因中的突变引起的脊髓性肌萎缩。
因此,本公开提供了用于促进在smn2pre-mrna加工期间包含非典型外显子的tc-dnaaon,其中所述tc-dnaaon长度为6-22个三环核苷酸、特别是8-20个三环核苷酸、更特别长度为10-18个三环核苷酸,并且其中tc-dnaaon与smn2pre-mrna的内含子沉默序列(iss)或末端茎-环(tsl)互补。这样的tc-dnaaon可以具有13至17个核苷酸、12至16个核苷酸或13至15个核苷酸。
本文中示例了包含15个核苷酸的序列5’-cuuucauaaugcugg-3’(smn2i7(10;25),seqidno:5)的tc-dnaaon,所述tc-dnaaon与smn2pre-mrna的iss互补,并且可用于促进将非典型外显子7包含在加工过的smn2mrna中。本文中还示例了包含13个核苷酸的序列5’-uuaauuuaaggaa-3’(smn2e7(39;51),seqidno:6)的tc-dnaaon,所述tc-dnaaon与smn2pre-mrna的tsl2互补,并且也可用于促进将外显子7包含在加工过的smn2mrna中。
用于治疗steinert肌强直性营养不良的三环dna反义寡核苷酸
在其他实施方案中,本公开提供了可以适用于治疗由编码dm1的mrna的3’末端处的cug扩增序列所引起的steinert肌强直性营养不良的tc-dnaaon。据认为,含有过多cug扩增序列的突变dm1mrna被隔绝在核中并积累以形成核灶点(nuclearfoci)。这些灶点是稳定的,并被认为与参与剪接机制的因子结合,从而广泛地影响转录组。作为本公开的一部分,通过使用u7snrna系统,证实了tc-dnaaon可用于靶向cug序列并促进突变的dm1mrna的破坏,从而引起剪接因子的释放和核灶点的移除。不受具体机制理论的束缚,还相信本文公开的tc-dnaaon能够促进含有过量cug扩增序列的mrna的破坏,这是相当出乎意料的。
因此,描述了可以适用于促进包含过量cug扩增序列的突变dm1mrna的破坏的tc-dnaaon。这样的tc-dnaaon包含9-27个三环核苷酸,其中所述tc-dnaaon与包含一个或多个3’cug扩增序列的突变dm1mrna互补,并且其中所述tc-dnaaon能够促进dm1mrna的破坏。取决于所设想的具体应用,tc-dnaaon可以包含3至9个、4至8个或5、6或7个连续重复的核苷酸序列5’-cag-3’(seqidno:7)。用于促进突变dm1的破坏的示例性tc-dnaaon包含15个核苷酸的序列5’-cagcagcagcagcag-3’(dm1(cag5),seqidno:8)。用于促进突变dm1的破坏的另一个示例性tc-dnaaon包含15个核苷酸的序列5’-cagcagcagcagcagcagcag-3’(dm1(cag7),seqidno:9)。
三环dna反义寡核苷酸的合成和分离
tc-dnaaon可以使用本技术领域中已知的方案来合成,例如在下述文献中所描述的:caruthers等,methodsinenzymol.211:3-19(1992);thompson等,pct公布no.wo99/54459;wincott等,nucleicacidsres.23:2677-26841995);wincott等,methodsmol.bio.74:59(1997);brennan等,biotechnolbioeng.61:33-451998);以及brennan,美国专利no.6,001,311。
用于合成tc-dna和tc-dnaaon的方法已被描述,并且在本技术领域中是公知的。参见例如steffens和leumann,j.am.chem.soc.121(14):3249-3255(1999);steffens和leumann,j.am.chem.soc.119:11548-11549(1997);以及wengel,美国专利no.7,034,133。tc-dna可以在商用dna合成仪上由常规固相氰乙基亚磷酰胺化学品产生的亚磷酰胺来合成。tc-dna亚磷酰胺构造单元可以按照steffens和leumann,c.helv.chim.acta80:2426-2439(1997)中的描述来合成。链延伸循环可以与用于天然寡脱氧核糖核酸合成的链延伸循环基本上相同。参见pharmacialkb用户手册(56-1111-56)(geneassemblerspecial/4primers)。
例如,tc-dnaaon的合成可以通过固相亚磷酰胺方法,使用与个人计算机相连的pharmacialkbgeneassemblerspecial仪器或appliedbiosystemspcr-mateepdna合成仪(391型)来实现。试剂溶液可以按照制造商的方案来制备。参见appliedbiosystemspcr_mateepdna合成仪(391型)用户手册(1989),以及pharmacialkb用户手册(56-1111-56)(geneassemblerspecial/4primers)。1h-四唑(在mecn中的0.45m溶液)可以从fluka获得。
三环dnaaon的装配可以按照标准的合成循环来进行,不同之处在于连接时间延长(例如6分钟),11倍过量的亚磷酰胺,以及0.1m的三环腺苷构造单元溶液由于溶解性不良可以使用0.07m来代替。lcaa-cpg(sigma)或聚苯乙烯(pharmacia)结合的天然核苷可用作起始单元。
合成可以在去掉三苯甲基(trityloff)的模式下执行,以5’-去三苯甲基化寡聚物结束。连接效率可以通过在线三苯甲基测定来监测,并且其典型在90至99%之间。在合成后,可以将固相载体悬浮在浓nh3溶液中,并在55℃下放置15小时或在室温下放置2小时。
粗制tc-dnaaon可以通过本技术领域已知的许多方法中的任一种来纯化,例如超滤、凝胶电泳或层析。可以在nucleogendeae60-7(125x4mm)柱上实现离子交换hplc。分离的寡核苷酸可以在sp-pakc-18柱(waters)上脱盐,如sambrook等《分子克隆实验指南》(molecularcloning:alaboratorymanual)11.29(coldspringharborlaboratorypress,plainview,ny,1989)中所述。可以将纯化的tc-dnaaon溶解在150mmnacl,10mmtris-hcl,ph7.0中,并与碱性磷酸酶(1mg/ml)和磷酸二酯酶(2mg/ml)在37℃下温育。5小时后,可以对溶液进行hplc纯化。
具有取代或修饰(碱基、糖或磷酸)的化学合成的核酸分子可以防止它们被血清核糖核酸酶降解,这可以增加它们的效能。参见例如eckstein等,pct公布no.wo92/07065;perrault等,nature344:565(1990);pieken等,science253:314(1991);usman和cedergren,trendsinbiochem.sci.17:334(1992);usman等,pct公布no.wo93/15187;以及rossi等,pct公布no.wo91/03162;sproat,美国专利no.5,334,711;gold等,美国专利no.6,300,074。所有上述参考文献描述了可以对本文描述的tc-dnaaon的碱基、磷酸或糖部分做出的各种化学修饰。
用于体内给药的三环dna的配制
本文描述的tc-dnaaon可以与适合用于制造水性悬液的赋形剂混合。这样的赋形剂是悬浮剂例如羧甲基纤维素钠、甲基纤维素、羟丙基甲基纤维素、藻酸钠、聚乙烯吡咯烷酮、黄蓍树胶和阿拉伯树胶;分散或润湿剂可以是天然存在的磷脂例如卵磷脂,或烯化氧与脂肪酸的缩合产物例如聚氧化乙烯硬脂酸酯,或氧化乙烯与长链脂族醇的缩合产物例如十七氧化乙烯鲸蜡醇,或氧化乙烯与源自脂肪酸和己糖醇的偏酯的缩合产物例如聚氧化乙烯山梨糖醇单油酸酯,或氧化乙烯与源自脂肪酸和己糖醇酐的偏酯的缩合产物例如聚氧化乙烯失水山梨糖醇酐单油酸酯。水性悬液也可以包含一种或多种防腐剂,例如对羟基苯甲酸乙酯或正丙酯。适合通过添加水来制备水性悬液的可分散粉末和颗粒,提供了活性成分与分散或润湿剂、悬浮剂和一种或多种防腐剂的混合。
tc-dnaaon组合物可以是无菌可注射水性或油性悬液的形式。悬液可以按照已知技术,使用上面提到的适合的分散或润湿剂和悬浮剂来配制。无菌可注射制剂也可以是在无毒的可肠胃外使用的稀释剂或溶剂中的无菌可注射溶液或悬液,例如作为在1,3-丁二醇中的溶液。可以使用的可接受的介质和溶剂是水、林格氏(ringer's)溶液和等渗氯化钠溶液。此外,无菌的不挥发油被常规用作溶剂或悬浮介质。对于这种目的来说,可以使用任何品牌的不挥发油,包括合成的甘油单酯或甘油二酯。此外,发现脂肪酸例如油酸可用于制备注射剂。
本公开还包括被制备用于储存或给药的tc-dnaaon组合物,其在可药用载体或稀释剂中包含药物有效量的所需化合物。治疗用的可接受载体或稀释剂在制药技术领域中是公知的,并描述在例如《remington药物学》(remington'spharmaceuticalsciences)(mackpublishingco.,a.r.gennaro主编,1985)中。例如,可以提供防腐剂和稳定剂。它们包括苯甲酸钠、山梨酸、和对羟基苯甲酸的酯。此外,可以使用抗氧化剂和悬浮剂。
本公开提供了tc-dnaaon组合物,以及用于促进pre-mrna中的外显子跳跃或掩蔽内含子沉默或末端茎环或者用于靶向破坏细胞或生物体中的mrna的方法。在相关实施方案中,本公开提供了用于治疗患有上文中所述疾病或具有发生所述疾病的风险的对象,包括人类细胞、组织或个体的方法和tc-dnaaon组合物。在一个实施方案中,方法包括向细胞或生物体例如哺乳动物给药本公开的tc-dnaaon或含有所述tc-dnaaon的药物组合物,以便改变pre-mrna的加工或定向破坏mrna。适合使用本公开的组合物和方法治疗的哺乳动物对象,包括患有适合接受这样的治疗的一种或多种病症例如杜氏肌营养不良、脊髓性肌萎缩或steinert肌强直性营养不良的对象。
本公开的tc-dnaaon组合物可以有效地用作可药用制剂。可药用制剂对于患者中的疾病状态或其他不利状况起到预防、改变其发生或严重性或治疗(以可检测或可测量的程度减轻一种或多种症状)的作用。可药用制剂包括上述化合物的盐,例如酸加成盐例如盐酸、氢溴酸、乙酸和苯磺酸的盐。药物组合物或制剂是指适合于给药、例如全身给药到细胞或患者例如人类中的形式的组合物或制剂。适合的形式部分取决于用法或进入途径,例如透皮或通过注射。这样的形式应该不阻止所述组合物或制剂到达靶细胞(即希望将tc-dnaaon递送给它的细胞)。例如,注射到血流中的药物组合物应该是可溶的。其他因素在本技术领域中是已知的,包括诸如毒性和阻止组合物或制剂行使其效应的形式的考虑。
本公开的药物组合物也可以采取水包油乳液的形式。油性相可以是植物油或矿物油或这些的混合物。适合的乳化剂可以是天然存在的树胶例如阿拉伯树胶或黄蓍树胶、天然存在的磷脂例如大豆、卵磷脂以及源自于脂肪酸和己糖醇、酐的酯或偏酯例如失水山梨糖醇单油酸酯,以及所述偏酯与氧化乙烯的缩合产物例如聚氧化乙烯失水山梨糖醇单油酸酯。
本公开的tc-dnaaon可以使用或不使用稳定剂、缓冲剂等形成适合用于治疗的组合物,以任何标准方式进行给药。当需要使用脂质体递送机制时,可以遵照形成脂质体的标准方案。因此,本公开的tc-dnaaon可以用任何方式给药,例如透皮或通过局部、全身或鞘内注射。
本公开还描述了包含表面修饰脂质体的tc-dnaaon组合物的使用,所述脂质体含有聚(乙二醇)脂类(peg修饰或长循环脂质体或隐形脂质体)。这些制剂提供了用于增加tc-dnaaon在靶组织中的积累的方法。这类药物载体能够抵抗调理作用以及单核细胞吞噬系统(mps或res)的消除,从而能够获得囊化的tc-dnaaon的更长血液循环时间并增加组织对其的暴露(lasic等,chem.rev.95:2601-2627(1995),以及ishiwata等,chem.pharm.bull.43:1005-1011(1995))。长循环脂质体改进了tc-dnaaon的药物动力学和药效学,特别是与已知在mps组织中积累的常规阳离子脂质体相比(liu等,j.biol.chem.42:24864-24870(1995);choi等,pct公布no.wo96/10391;ansell等,pct公布no.wo96/10390;holland等,pct公布no.wo96/10392)。在其避免在代谢上具有攻击性的mps组织例如肝脏和脾脏中积累的基础上,长循环脂质体与阳离子脂质体相比,还可能更大程度地保护tc-dnaaon免于核酸酶降解。
药物有效剂量是预防疾病状态、抑制其发生或对其进行治疗(在一定程度上减轻症状,优选所有症状)所需的剂量。药物有效剂量取决于疾病类型、所使用的组合物、给药途径和所治疗的哺乳动物类型、所考虑的具体哺乳动物的身体特征以及医学领域的专业人员将会意识到的其他因素。例如,取决于本公开的tc-dnaaon的效能,活性成分的给药量介于0.1mg/kg至100mg/kg体重/天之间。
每天每千克体重约0.1mg至约140mg量级的剂量水平可用于治疗上面指出的病症(每位患者每天约0.5mg至约7g)。可以与载体材料合并以生产单剂量形式的活性成分的量,取决于所治疗的宿主和具体的给药方式。剂量单位形式一般含有约1mg至约500mg活性成分。
应该理解,任何特定患者的具体剂量水平取决于各种因素,包括所使用的具体化合物的活性、年龄、体重、总体健康、性别、饮食、给药时间、给药途径、排泄速率、药物组成以及正在治疗的具体疾病的严重性。在根据本公开的配方和方法给药tc-dnaaon组合物后,测试对象与安慰剂治疗的或其他适合的对照对象相比,与被治疗疾病或病症相关的一种或多种症状将表现出约10%直至约99%的减轻。
tc-dnaaon可以通过本技术领域的专业人员已知的各种方法给药到细胞,包括在只包含tc-dnaaon的制剂中或在还包含一种或多种其他组分例如可药用载体、稀释剂、赋形剂、佐剂、乳化剂、缓冲剂、稳定剂、防腐剂等的制剂中给药。在某些实施方案中,tc-dnaaon可以囊封在脂质体中、通过离子电渗给药、或掺入到其他介质例如水凝胶、环糊精、生物可降解纳囊、生物黏附性微球或蛋白质载体中(参见例如pct公布no.wo00/53722)。
本公开的tc-dnaaon的直接注射,不论是皮下、肌肉内还是真皮内,都可以使用标准的针头和注射器方法来进行,或者可以通过无针技术,例如在conry等,clin.cancerres.5:2330-2337(1999)和pct公布no.wo99/31262中所描述的。
用于递送核酸分子、例如本公开的tc-dnaaon的其他方法,描述在例如下列文献中:boado等,j.pharm.sci.87:1308-1315(1998);tyler等,febslett.421:280-284(1999);pardridge等,proc.nat'lacad.sci.usa92:5592-5596(1995);boado,adv.drugdeliveryrev.15:73-107(1995);aldrian-herrada等,nucleicacidsres.26:4910-4916(1998);tyler等,proc.nat'lacad.sci.usa96:7053-7058(1999);akhtar等,trendscellbio.2:139(1992);《用于反义寡核苷酸治疗的递送策略》(deliverystrategiesforantisenseoligonucleotidetherapeutics)(akhtar主编,1995);maurer等,mol.membr.biol.16:129-140(1999);hofland和huang,handb.exp.pharmacol137:165-192(1999);以及lee等,acssymp.ser.752:184-192(2000)。这些方案可用于增补或补充本公开中实际考虑的任何tc-dnaaon的递送。
实施例
上面的公开内容对本公开进行了一般性描述,其将通过下面的实施例进行进一步示例说明。描述这些具体实施例仅仅是出于说明的目的,而不打算限制本公开的范围。尽管在本文中使用了具体的靶、项和值,但这些靶、项和值同样应该被理解为是示例性的,而不对本公开的范围构成限制。。
实施例1
使用三环dna反义寡核苷酸挽救营养不良的肌纤维中的肌养蛋白
杜氏肌营养不良(dmd)是x染色体连锁的隐性病,其源自于编码肌养蛋白的基因中的突变。肌养蛋白基因内的框外缺失编码了截短型肌养蛋白缺陷,其引起严重的dmd表型。开发了使用三环dna(tc-dna)反义寡核苷酸(aon)的外显子跳跃技术以允许有效挽救肌养蛋白基因的框外突变,由此恢复翻译阅读框并因此生产有功能活性的肌养蛋白。描述了tc-dnaaon例如与外显子23/内含子23接点杂交并干扰pre-mrna加工,使得外显子23在得到的加工后mrna中被剪掉。或者,tc-dnaaon与外显子51/内含子51接点杂交,同样地干扰pre-mrna加工,使得外显子51在加工后mrna中被剪掉。因此,得到的肌养蛋白缺失了分别由外显子23或外显子51编码的氨基酸序列,但仍然保有足够的功能性,使得严重dmd表型得到逆转。
实施例2
mdx小鼠模型
mdx小鼠是鼠类dmd模型,其缺少全长肌养蛋白,但是保留了所有较小的肌养蛋白同种型。bulfield等,proc.natl.acad.sci.usa81:1189-1192(1984)。mdx小鼠在肌养蛋白基因的外显子23中带有无义突变,其组织了功能性肌养蛋白合成(参见图3)。外显子23部分编码重复序列r6和r7,其中c到t的突变产生了终止密码子(taa)。
实施例3
体外研究
本实施例证实了用被称为m23d(+02-13)的具有核苷酸序列5’-aacctcggcttacct-3’的15个核苷酸tc-dnaaon转染的mdx肌管,在外显子23的下游供体剪接位点处经历外显子跳跃,使得肌养蛋白pre-mrna被加工成缺失外显子23的mrna。
设计了被称为m23d(+02-13)的tc-dnaaon,使其与外显子23/内含子23剪接接点处的靶序列内含子22–ttttgag[gctc…外显子23...tcag]gtaagccgaggtttggcc–内含子23杂交。
使用或不使用oligofectamine,将mdx肌管用tc-dnaaonm23d(+02-13)(1、2和10μg)转染。留下一个样品不作处理,作为阴性对照。48小时后,收获培养物并使用rneasy小型试剂盒(qiagen)提取mrna。然后如下所述将mrna反转录。将8微升提取的rna(500ng至1μg)与1μldntp和1μl随机六聚物混合,并将混合物在65℃下温育5分钟。然后将混合物在冰上冷却。向混合物加入25mmmgcl2(4μl)、0.1mdtt(2μl)、1μlrnaseout核糖核酸酶抑制剂(40u/μl)、1xtampon(2μl的10x储液)和50usuperscript反转录酶,使反应的终体积为20μl。然后将反应在25℃下温育10分钟,然后在42℃下温育50分钟。接下来,通过在70℃下加热15分钟使反应失活。然后将反应置于冰上并涡旋。然后向反应加入1μlrnaseh,并在37℃下温育20分钟。
接下来,使用下列条件,通过巢式pcr测定外显子23的跳过。将25μlpcr主混合物(taq聚合酶50u/μl,400μmdntp,3mmmgcl2)与3μlcdna、22μlh2o和下列两种引物(储液浓度为100μm)各1μl合并:
ex20fo5’–cagaattctgccaattgctgag–3’
ex26ro5’–ttcttcagcttgtgtcatcc–3’
然后按照下列热循环参数进行反应。94℃5分钟,然后进行30个循环的94℃30秒、55℃1分钟和72℃2分钟。最后,将反应在72℃下进行5分钟温育。
将2微升pcr产物加入到另一个pcr反应中作为模板,使用1μl下列引物(储液浓度为100μm)。反应还包含25μlpcr主混合物和23μlh2o。
ex20fi5’–cccagtctaccaccctatcagagc–3’
ex26ri5’–cctgcctttaaggcttcctt–3’
按照下列热循环参数进行反应–94℃5分钟,然后进行25个循环的94℃30秒、55℃1分钟和72℃2分钟。最后,将反应在72℃下进行5分钟温育。
图4中显示的数据证明:(1)15个核苷酸的tc-dnaaonm23d(+02-13)能够在离体条件下实现mdx的肌养蛋白mrna的突变外显子23的跳过,以及(2)oligofectamine增加了tc-dnaaon的离体摄取。
在oligofectamine存在下将mdx肌管用tc-dnaaonm23d(+02-13)(0.5、1、2、5和10μg)转染。培养物如上所述进行处理。图5中显示的数据证明在存在2μgtc-dnaaonm23d(+02-13)的情况下有明显的跳过。
在oligofectamine存在下将mdx肌管用5μgtc-dnaaonm23d(+02-13)转染。在转染后不同时间点(从第0日到第15日)如上所述对培养物进行处理。图6中显示的数据证明:(1)在第3日(d3)观察到跳过,以及(2)在第15日(d15)仍可检测到跳过,但是从第7日(d7)起减少。
实施例4
体内研究
本实施例证实了用被称为m23d(+02-13)的具有核苷酸序列5’-aacctcggcttacct-3’的15个核苷酸tc-dnaaon注射的mdx小鼠,在外显子23的下游供体剪接位点处经历外显子跳跃,使得肌养蛋白pre-mrna被加工成缺失外显子23的mrna。
对8周龄mdx小鼠在胫前肌中用含有100、80、40、20、10和5μgtc-dnaaonm23d(+02-13)的50μlpbs(磷酸盐缓冲盐水)进行注射。3周后处死动物。对肌肉样品进行处理,使用上面实施例3给出的相同参数进行mrna分析,结果提供在图7中。
图7中显示的数据证明在所有测试条件下发生了外显子跳跃。此外,在横切片中检测到显著水平的肌养蛋白(未显示)。注射2μgtc-dnaaonm23d(+02-13)与5μgtc-dnaaonm23d(+02-13)等效(未显示)。
对8周龄mdx小鼠用含有10μgtc-dnaaonm23d(+02-13)的50μlpbs进行肌肉内注射。在注射后第4、10和20周时处死动物。如上所述测定肌肉样品和横切片的肌养蛋白mrna。结果显示在图8中。
如下所述使用免疫染色测定肌肉横切片的肌养蛋白表达。使用m.o.m.(小鼠对小鼠)试剂盒,向样品加入第一单克隆抗体ncl–dys2(1:100稀释)。然后将样品在pbs中清洗3次。然后将样品与alexafluor488(molecularprobes)标记的第二抗体山羊抗小鼠igg抗体温育2小时,随后进行pbs清洗、0.01%tritonx清洗和最终的pbs清洗。该实验的结果提供在图9中。
图8和9中显示的数据证明(1)在第4和10周时外显子跳跃明显,但是在第20周时没有观察到;(2)从4周至20周可以清楚地检测到肌养蛋白;以及(3)tc-dnaaonm23d(+02-13)在体内似乎比离体更有效。
实施例5
脑免疫染色
对8周龄mdx小鼠在海马或脑脊液(鞘内注射)中用含有20μg(海马)或200μg(鞘内)tc-dnaaonm23d(+02-13)的50μlpbs进行注射。在注射后一个月处死动物。如上面实施例4中所述测定脑切片(图a、b、c)和小脑切片(图d、e、f)的肌养蛋白。结果显示在图10中。图a和d对应于阴性对照,其中小鼠没有进行处理。图b和e对应于来自mdx小鼠的未处理的切片。图c对应于海马处理的mdx小鼠(c),图(f)显示了来自mdx小鼠的鞘内处理的小脑的结果。用dapi对核进行复染。
实施例6
递送(预示性的)
tc-dnaaonm23d(+01-13)可以通过腹膜内和皮下注射来递送(从1g/kg及更低剂量开始)。
实施例7
用于dmd小鼠肌养蛋白援救的三环dna反义寡核苷酸
本实施例证实了针对肌养蛋白的外显子51/内含子51接点设计的、具有序列5’-agaaatgccatcttc-3’(“tc-dnaaonh51(+68+82)”;seqidno:2)、5’-aaatgccatcttcct-3’、(“tc-dnaaonh51(+70+84)”;seqidno:3)和5’-tgccatcttccttga-3’(“tc-dnaaonh51(+73+87)”;seqidno:4)的tc-dnaaon,在来自于表达全长人类肌养蛋白基因的小鼠(“hdmd小鼠”)的肌肉细胞中有效地介导外显子51的跳过。
对于8至10周龄的hdmd小鼠,在胫前肌中注射含有10μg也称为tc-dnaex51esea的tc-dnaaonh51(+68+82)、也称为tc-dnaex51eseb的tc-dnaaonh51(+70+84)或也称为tc-dnaex51esec的tc-dnaaonh51(+73+87)的50μlpbs。4周后处死动物。对肌肉样品进行处理,使用上面实施例3给出的相同参数进行mrna分析,结果提供在图12中。
使用下列条件,通过巢式pcr测定外显子51的跳过。使用accessrt-pcr系统(promega),使用500纳克总rna,在含有下列外部引物的50μl反应中进行rt-pcr:
hex49f2(5’-aaactgaaatagcagttcaagc-3’)
hex53r2(5’-ttgcctccggttctgaagg-3’)
cdna合成在45℃下进行45分钟,然后使用下列参数直接进行20个循环的初级pcr:20个循环的94℃40秒、60℃40秒和72℃40秒。
将2微升pcr产物加入到另一个pcr反应中作为模板,使用下列引物(储液浓度为100μm)。
hex50f5’-aggaagttagaagatctgagc-3’
hex52r25’-ttcttccaactggggacgc-3’
按照下列热循环参数进行反应:94℃5分钟,然后进行30个循环的94℃40秒、60℃40秒和72℃40秒。最后,将反应在72℃下进行5分钟温育。
图12中显示的数据证明,测试的每种tc-dnaaonh51构建物(tc-dnaaonh51(+68+82)、tc-dnaaonh51(+70+84)和tc-dnaaonh51(+73+87))都引起外显子51跳过的增加。
实施例8
定向到smn外显子7/内含子7接点和内含子7iss的三环dna反义寡核苷酸促进外显子7包含在smn2中
本实施例证明了针对smn外显子7/内含子7接点处和内含子7iss设计的tc-dnaaon(分别为“tc-dnaaonsmn2e7(39;51)”和“tc-dnaaonsmn2i7(10;25)”),在从sma患者分离的成纤维细胞(g03813细胞系)中有效介导外显子7包含在smn2中。
tc-dnaaonsmn2i710;25):5’-cuuucauaaugcugg-3’(seqidno:5)
tc-dnaaonsmn2e7(39;51):5’-uuaauuuaaggaa-3’(seqidno:6)
g03813细胞系源自于具有两个smn2拷贝的3岁的i型sma患者。将gm03813细胞培养在含有20%胎牛血清和1%青霉素-链霉素(100u/ml)的dulbecco修改的eagle培养基(dmem)中。tcdnaaon(tc-dnaaonsmn2i7(10;25)和tc-dnaaonsmn2e7(39;51))转染使用oligofectamine(invitrogen),在无血清和抗生素的培养基中进行48小时。在转染后48小时,使用trizol试剂(invitrogen)提取总rna,并使用superscriptii(invitrogen)和随机六聚体进行第一链cdna合成。使用mastermix2xphusiongc(finnzymes)在50μl总体积中进行pcr,其中含有11μlcdna和smn-ex6-fw与smn-ex8-re引物各10μm。然后在3%琼脂糖凝胶上通过电泳分离pcr产物。
smn-ex6-fw5’-gctgatgctttgggaagtatgta-3’
smn-ex8-re5’-attccagatctgtctgatcg-3’
图19中给出的数据显示了tc-dnaaonsmn2e7(39;51)和tc-dnaaonsmn2i7(10;25)两者都促进外显子7包含在smn2mrna中。图19a显示了来自用所指示的tc-dna寡核苷酸处理的gm03813细胞的rna的rt-pcr分析。使用这两种tc-dna寡核苷酸(第3和4道)时,对应于不含外显子7的smn2(“smn2(δ7)”)的条带强度比未处理道(第2道)中的条带低。图19b是凝胶的归一化定量图,显示出在tc-dnaaon转染后,对应于smn1+全长smn2的上部条带增强,而对应于smn2(δ7)的下部条带明显减弱。
图19c显示了来自野生型成纤维细胞和用所指示的tc-dna寡核苷酸转染的gm03813细胞的裂解液的western印迹。在裂解缓冲液(10mmol/lhepesph7.9,00mmol/lkcl,1mmol/ledta,1mmol/l1,4-二硫苏糖醇,1×完全蛋白酶抑制剂混合物(roche),0.5%np-40)中获得蛋白提取物。将同等量的蛋白(通过bradford蛋白测定(bradfordproteinassay,pierce)测定)与2×载样缓冲液(125mmol/ltrisph6.8,2%十二烷基硫酸钠,10%甘油,0.01%溴酚蓝,10%β-巯基乙醇)混合,并使用bradford蛋白测定(pierce)测定蛋白质浓度。将10微克每种蛋白样品通过sds-page4-12%bis-tris凝胶(invitrogen)进行分离,并转移到硝酸纤维素膜上。将膜用含有10%奶的pbs-tween缓冲液阻断,用识别smn1、smn2和截短形式的smn2的兔多克隆smn抗体(1:500稀释;h-195,santacruz)探测,然后与辣根过氧化物酶接合的山羊抗兔第二抗体(1:50,000)温育。使用supersignalwestpico化学发光试剂盒(thermoscientific)检测信号。为了证实蛋白的相等载样量,将膜清洗,重新阻断,并用小鼠单克隆抗肌动蛋白抗体探测,然后与辣根过氧化物酶接合的绵羊抗小鼠第二抗体(1:15,000)温育。如上所述检测信号。第1道:用tc-dnaaonsmn2i7(10;25)转染的gm03813细胞;第2道:用tc-dnaaonsmn2e7(39;51)转染的gm03813细胞;第3道:用tc-dnaaonsmn2i7(10;25)和tc-dnaaonsmn2e7(39;51)两者转染的gm03813细胞;第4道:野生型成纤维细胞;第5道:未转染的gm03813细胞。
图19c中的数据表明,用tc-dna寡核苷酸转染由于将外显子7包含在smn2mrna中,挽救了gm03813细胞中的smn水平。由于外显子7在smn2中的散发性自然包含,对照gm03813细胞表现出一些smn蛋白。
图19d显示了用30μgtc-tsl转染、随后用smn抗体染色的gm03813细胞的光学显微照片。将载片上转染的gm03813细胞用丙酮/甲醇(体积/体积)固定。将固定的细胞在pbs+5%bsa中阻断1小时,然后与兔多克隆smn抗体(1:100,在pbs+1%bsa中;h-195,santacruz)温育1小时。将细胞在pbs中清洗,并与alexa594接合的抗兔第二抗体温育1小时。然后将细胞在pbs中清洗,并与dapi(1:50,000)温育5分钟。使用fluoromount-g(southernbiotech)将载片盖片配合,并在4℃下温育过夜。光学显微照片显示,在用tc-dnatsl转染的gm03813细胞的核(蓝色)中,smn(红色)水平增加。
实施例9
定向于cug重复序列的三环dna反义寡核苷酸降低了dm1成肌细胞中突变dmpkmrna的表达
本实施例证明了针对突变dmpkmrna中的cug重复序列设计的具有序列5’-cagcagcagcagcagcagcag-3’(“tc-dnaaondm1(cag7)”;seqidno:9)的tc-dnaaon,在从“肌库”研究用组织库(tissuebankforresearch“myobank”)获得的肌肉活检样品分离到的人类dm1成肌细胞中,有效降低了具有800个cug重复序列的突变dmpkmrna的表达。
使用lipofectamine(invitrogen),将dm1成肌细胞用量渐增的tc-dnaaondm1(cag7)(0、3.5μg、10μg和20μg)转染。在转染后三天,通过northern印迹检测野生型和突变型dmpkmrna的表达。简单来说,将5-10μg总rna在含有0.66m甲醛的1.3%琼脂糖mops凝胶上分离,并通过用10xssc进行毛细转移,转移到hybond-n+膜(amershampharmaciabiotech)上。将斑点与随机引发的32p标记的(dmpkcdna的bglii-saci片段)探针,在杂交缓冲液(2%sds,10%硫酸葡聚糖,1xsspe,100μg/ml鲑鱼精子dna,2%denhart's)中在68℃下杂交过夜。信号在感光成像仪(molecularimagerfx,bio-rad)上分析,并使用quantityone(bio-rad)定量。
图20a显示出随着转染的tc-dnaaondm1(cag7)的量的增加,突变型dmpk降低。重要的是,野生型dmpk水平不变。使用18srna作为载样对照。图20b是来自图20a的northern印迹的定量图。通过测量突变型dmpk与野生型dmpk的条带强度比率来进行定量。
实施例10
定向于cug重复序列的tc-dna在dm1小鼠中降低突变dmpkmrna的表达
本实施例证实了tc-dnaaondm1(cag7)有效降低在dm1小鼠的ta肌肉中表达的具有700个cug重复序列的突变型人类dmpkmrna的水平。
用量渐增的tc-dnaaondm1(cag7)注射到表达具有700个cug重复序列的人类dmpkmrna的dm1小鼠的ta肌肉中。一周后,使用trizol试剂(invitrogen)提取总rna。通过northern印迹检测人类dmpk和小鼠dmpkmrna。简单来说,将8-10μg总rna在含有0.66m甲醛的1.3%琼脂糖mops凝胶上分离,并通过用10xssc进行毛细转移,转移到hybond-n+膜(amershampharmaciabiotech)上。将斑点与随机引发的32p标记的(dmpkcdna的bglii-saci片段)探针,在杂交缓冲液(2%sds,10%硫酸葡聚糖,1xsspe,100μg/ml鲑鱼精子dna,2%denhart's)中在68℃下杂交过夜。信号在感光成像仪(molecularimagerfx,bio-rad)上分析,并使用quantityone(bio-rad)定量。
图21a和21c显示出随着转染的tc-dnaaondm1(cag7)的量的增加,突变型人类dmpk的水平降低。图21b和21d是分别来自图a和c的northern印迹的定量图。通过测量突变体人类dmpk与野生型小鼠dmpk的条带强度比率来进行定量。
尽管已经在本公开的各种实施方案的每个中对本公开内容进行了描述,但预计本技术领域的专业人员可以对其进行和执行某些修改,而不背离在前面的说明书中提出的并在下面的权利要求书中进一步体现的本公开的真实精神和范围。本公开的范围不受本文描述的具体实施方案的限制。事实上,对于本技术领域的专业人员来说,根据上述说明书和附图,在本文所描述的之外对本公开内容进行各种修改将是显而易见的。这样的修改旨在属于本发明的权利要求书的范围之内。还应该理解,所有值都是近似值,提供它们是出于说明的目的。
在本说明书中提到的所有美国专利、美国专利申请公开、美国专利申请、外国专利、外国专利申请、非专利出版物、图、表格和网址,在此明确以其全文引为参考。
序列表
seqidno:1
人工序列
dna
长度:15
tc-dnaaonm23d(+02-13)
aacctcggcttacct
seqidno:2
人工序列
dna
长度:15
tc-dnaaonh51(+68+82)
agaaatgccatcttc
seqidno:3
人工序列
dna
长度:15
tc-dnaaonh51(+70+84)
aaatgccatcttcct
seqidno:4
人工序列
dna
长度:15
tc-dnaaonh51(+73+87)
tgccatcttccttga
seqidno:5
人工序列
dna
长度:15
tc-dnaaonsmn2i7(10;25)
cuuucauaaugcugg
seqidno:6
人工序列
dna
长度:13
tc-dnaaonsmn2e7(39;51)
uuaauuuaaggaa
seqidno:7
人工序列
dna
长度:12-21
tc-dnaaondm1cagn)
(cag)nn=4-7
seqidno:8
人工序列
dna
长度:15
tc-dnaaondm1(cag5)
cagcagcagcagcag
seqidno:9
人工序列
dna
长度:21
tc-dnaaondm1(cag7)
cagcagcagcagcagcagcag
序列表
<110>肌肉学研究协会
巴黎第六大学
伯尔尼大学
法国国家科学研究中心
<120>用于治疗疾病的三环dna反义寡核苷酸、组合物和方法
<130>spi185523-31
<140>pct/ep2010/054735
<141>2010-04-09
<150>us61/212,384
<151>2009-04-10
<160>22
<170>patentinversion3.3
<210>1
<211>15
<212>dna
<213>artificial
<220>
<223>tc-dnaaonm23d(+02-13)
<400>1
aacctcggcttacct15
<210>2
<211>15
<212>dna
<213>artificial
<220>
<223>tc-dnaaonh51(+68+82)
<400>2
agaaatgccatcttc15
<210>3
<211>15
<212>dna
<213>artificial
<220>
<223>tc-dnaaonh51(+70+84)
<400>3
aaatgccatcttcct15
<210>4
<211>15
<212>dna
<213>artificial
<220>
<223>tc-dnaaonh51(+73+87)
<400>4
tgccatcttccttga15
<210>5
<211>15
<212>dna
<213>artificial
<220>
<223>tc-dnaaonsmn2i7(10;25)
<400>5
cuuucauaaugcugg15
<210>6
<211>13
<212>dna
<213>artificial
<220>
<223>tc-dnaaonsmn2e7(39;51)
<400>6
uuaauuuaaggaa13
<210>7
<211>3
<212>dna
<213>artificial
<220>
<223>tc-dnaaondm1(cagn)
<220>
<221>misc_feature
<222>(1)..(3)
<223>(cag)n:nrepeatof(cag)trinucleotidewithn=4-7
<400>7
cag3
<210>8
<211>15
<212>dna
<213>artificial
<220>
<223>tc-dnaaondm1(cag5)
<400>8
cagcagcagcagcag15
<210>9
<211>21
<212>dna
<213>artificial
<220>
<223>tc-dnaaondm1(cag7)
<400>9
cagcagcagcagcagcagcag21
<210>10
<211>22
<212>dna
<213>musmusculus
<400>10
tcaggtaagccgaggtttggcc22
<210>11
<211>17
<212>dna
<213>artificial
<220>
<223>tc-dnaaonsmn2e7(39;51)
<400>11
uuaauuuaaggaaugug17
<210>12
<211>17
<212>dna
<213>artificial
<220>
<223>tc-dnaaonsmn2i7(10;25)
<400>12
cacuuucauaaugcugg17
<210>13
<211>22
<212>dna
<213>artificial
<220>
<223>ex20fonestedpcrprimer
<400>13
cagaattctgccaattgctgag22
<210>14
<211>20
<212>dna
<213>artificial
<220>
<223>ex26ronestedpcrprimer
<400>14
ttcttcagcttgtgtcatcc20
<210>15
<211>24
<212>dna
<213>artificial
<220>
<223>ex20finestedpcrprimer
<400>15
cccagtctaccaccctatcagagc24
<210>16
<211>20
<212>dna
<213>artificial
<220>
<223>ex26rinestedpcrprimer
<400>16
cctgcctttaaggcttcctt20
<210>17
<211>22
<212>dna
<213>artificial
<220>
<223>hex49f2nestedpcrprimer
<400>17
aaactgaaatagcagttcaagc22
<210>18
<211>19
<212>dna
<213>artificial
<220>
<223>hex53r2nestedpcrprimer
<400>18
ttgcctccggttctgaagg19
<210>19
<211>21
<212>dna
<213>artificial
<220>
<223>hex50fnestedpcrprimer
<400>19
aggaagttagaagatctgagc21
<210>20
<211>19
<212>dna
<213>artificial
<220>
<223>hex52r2nestedpcrprimer
<400>20
ttcttccaactggggacgc19
<210>21
<211>23
<212>dna
<213>artificial
<220>
<223>smn-ex6-fwpcrprimer
<400>21
gctgatgctttgggaagtatgta23
<210>22
<211>20
<212>dna
<213>artificial
<220>
<223>smn-ex8-repcrprimer
<400>22
attccagatctgtctgatcg20
- 还没有人留言评论。精彩留言会获得点赞!