ZmRPH1基因在调控植物株高及抗倒伏能力中的应用的制作方法
本发明涉及生物技术和植物遗传育种领域,具体地说,涉及zmrph1基因在调控植物株高及抗倒伏能力中的应用。
背景技术:
株高作为理想株型的最主要组成因素之一,在作物改良过程中发挥至关重要的作用(donald,1968)。适宜的株高有利于提高收获指数,增强抗倒伏能力进而实现高产稳产等诸多优势,这些优势已经在小麦、水稻的育种过程中得到了充分的利用。我们所说的“绿色革命”就是水稻、小麦的矮化育种带来的巨大的产量提高,具体涉及到水稻中sd1和小麦中rht1基因的利用(pengetal.,1999)。然而到目前为止,在对产量性状不造成不利影响的前提下,玉米中仍然未能找到可利用的能有效降低株高的基因。因此,进一步发掘玉米株高调控相关基因具有重要理论和实践意义。
转基因技术已经在作物改良过程中得到广泛应用,利用转基因技术能使作物获得多种优良性状,例如:抗虫、抗病、抗除草剂、抗旱、高产等。转基因技术的应用,在有效降低玉米株高方面同样能够发挥重要作用。
微管骨架和微丝骨架是细胞骨架的重要成员,在调控植物生长发育等诸多方面都发挥着重要功能。间期细胞的周质微管列阵的排列方向决定了细胞壁中纤维素微纤丝的沉积方向,进而调控细胞的伸长生长:当周质微管以垂直于生长轴方向横向平行排列时,细胞快速伸长;而当周质微管列阵随机或者纵向排布时,细胞生长缓慢甚至停止伸长(lloydandchan,2004)。而微管骨架的动态变化及列阵的取向又受到一系列微管相关蛋白的调控(chenetal.,2016)。在玉米中针对调控微管和微丝的细胞骨架相关蛋白的研究相对较少,而将其应用到降低玉米株高方面的作物改良中还未见报道。
技术实现要素:
本发明的目的是提供zmrph1基因在调控植物株高及抗倒伏能力中的应用。
为了实现本发明目的,第一方面,本发明提供zmrph1基因在调控植物株高及抗倒伏能力中的应用,其中,zmrph1基因编码的蛋白质为如下(a)或(b):
(a)由seqidno:2所示的氨基酸序列组成的蛋白质;
(b)seqidno:2所示序列经取代、缺失或添加一个或几个氨基酸且具有同等功能的由(a)衍生的蛋白质。
具体地,所述调控是指降低植物株高及增强抗倒伏能力。
本发明中,zmrph1基因及其编码蛋白的获得方法如下:首先从玉米数据库maizegdb中查找到zmrph1的蛋白编码核苷酸序列(seqidno:1),并以此全长序列设计引物(zmrph1-f/r),以玉米自交系b73-329的cdna为模板,pcr扩增得到zmrph1全长蛋白编码基因序列。
上游引物:zmrph1-f:5’-atggtggctgccgcggcg-3’
下游引物:zmrph1-r:5’-ctacaattgtgtcgggctctgcttt-3’
前述的应用包括:
1)使植物包含zmrph1基因;或者,
2)使植物过表达zmrph1基因。
本发明中,所述植物为单子叶植物或双子叶植物,优选玉米和拟南芥。
第二方面,本发明提供zmrph1基因在植物育种中的应用。所述育种目的包括降低植物株高及增强抗倒伏能力。
第三方面,本发明提供一种降低玉米株高及增强抗倒伏能力的方法,所述方法为:
1)使玉米包含zmrph1基因;或者,
2)使玉米过表达zmrph1基因。
所述方法包括但不限于转基因、杂交、回交、自交或无性繁殖。
本发明中,过表达zmrph1基因的方法选自以下1)~4),或任选的组合:
1)通过向玉米中导入具有所述基因的质粒;
2)通过增加玉米染色体上所述基因的拷贝数;
3)通过将强启动子与所述基因可操作地连接;
4)通过导入增强子。
在本发明的一个具体实施方式中,过表达zmrph1基因采用玉米组成型启动子ubiquitin1。
第四方面,本发明提供zmrph1基因在制备转基因植物中的应用。
第五方面,本发明分析了通过实时荧光定量pcr分析zmrph1基因的组织表达模式,表明zmrph1在快速生长的节间组织中高表达,说明zmrph1在节间伸长过程中发挥着重要作用。
用于实时荧光定量pcr检测玉米zmrph1基因的特异性引物对为:
上游引物:zmrph1qpcr-f:5’-caaagttgcaacgcaagaac-3’
下游引物:zmrph1qpcr-r:5’-ggctctgcttttggttacgtt-3’
为了分析zmrph1的蛋白性质,本发明首先在玉米原生质体中分析了zmrph1的亚细胞定位。zmrph1的亚细胞定位分析表明,gfp-zmrph1(zmrph1的n端连接gfp)的定位呈弥散状类似于空gfp对照,zmrph1-gfp(zmrph1的c端连接gfp)的定位呈现类似微管的丝状结构。
为了进一步分析zmrph1-gfp的丝状定位,本发明在玉米原生质体中分析了zmrph1-gfp与微管marker蛋白mcherry-α-tubulin1(α-tubulin1的n端连接mcherry)的共定位情况。共定位实验结果表明zmrph1-gfp能与mcherry-α-tubulin1共定位,这一结果证明zmrph1为微管相关蛋白。
为了进一步说明zmrph1与微管的关系,本发明进行了zmrph1与微管的共沉淀实验。实验结果表明,zmrph1能够直接与微管结合,证明zmrph1为微管结合蛋白。
本发明还构建了zmrph1的玉米过表达载体。在优选的实施方案中,玉米表达载体上的表达盒是由来自玉米的ubiquitin1基因的启动子和来自胭脂碱合成酶(nos)基因的3’转录终止区域构成,选择标记基因为草丁膦基因(phosphinothricin)。所述表达载体的出发载体为pbcxun载体(chenetal.,2009),构建得到重组质粒pbcxun-zmrph1,其中zmrph1基因正向插入,并且由玉米ubiquitin1启动子驱动表达。
本发明将含有zmrph1的过表达载体转化玉米自交系b73-329,获得了株高降低抗倒伏能力增强,单株产量不受影响的转基因玉米。
本发明还以玉米另一自交系t13为父本与已收获的纯合b73-329背景的过表达zmrph1基因的转基因玉米进行杂交,获得过表达zmrph1基因的杂交种转基因玉米。
过表达zmrph1基因的转基因杂交种玉米,株高显著降低,抗倒伏能力增强,单株产量不受影响。
进一步地,本发明将zmrph1基因异源过表达到拟南芥中,同样能够使植株株高显著降低。
借由上述技术方案,本发明至少具有下列优点及有益效果:
本发明首次发现zmrph1基因具有调控植物株高及抗倒伏能力的生物学功能。在玉米中调控zmrph1的表达可以调控玉米株高,过表达zmrph1基因可以降低玉米株高,提高玉米抗倒伏能力,并且不影响单株产量。本发明提供一种利用zmrph1基因降低玉米株高的新方法,可用于半矮化的玉米新品种的培育,有利于玉米高产稳产提高玉米收获指数,增强玉米抗倒伏能力。
附图说明
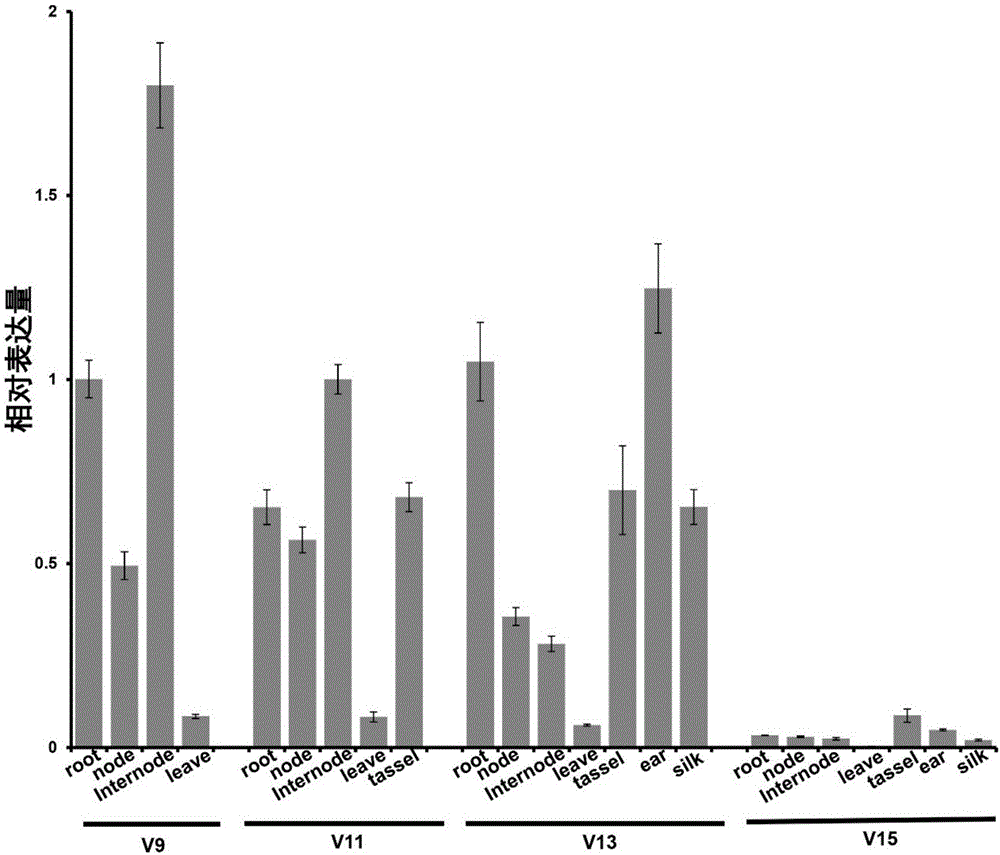
图1为本发明实施例3中分析zmrph1的组织表达模式结果。其中,v9,v11,v13,v15四个不同生长发育时期分别取材,以zmactin1作为内参基因。
图2为本发明实施例4、5中证明zmrph1是一个微管结合蛋白。其中,a、zmrph1的c端连接gfp检测到类似微管结构的丝状定位;b、zmrph1与微管marker共定位;c、zmrph1能直接结合微管,与聚合好的微管共沉淀。
图3为本发明实施例1中构建的zmrph1过表达载体表达盒。其中,启动子为zmubiquitin1的启动子,终止子为nos终止子,zmrph1处于启动子与终止子之间。
图4为本发明实施例6中zmrph1过表达材料zmrph1表达量检测结果。其中,oe1-oe9指9个不同自交系b73-329背景的过表达株系,取材为v3时期幼苗,zmactin1为内参基因。
图5为本发明实施例7中自交系材料田间表型照片。其中,oe1-oe4指自交种4个不同株系,b73-329背景。
图6为本发明实施例7中倒伏率统计结果。其中,a,sz16spr:2016年春季播种于北京上庄;kf16spr:2016年春季播种于河南开封;kf16sum:2016年夏季播种于河南开封。b,2016年播种于河北涿州的材料包括三个不同密度,分别是60000,90000,120000株/公顷。#oe1-#oe4指杂交种4个不同株系,wt背景,wt指b73-329与t13的杂交种。
图7为本发明实施例7中杂交种成熟雌穗照片。其中,2016年播种于河北涿州的材料包括三个不同密度,分别是60000,90000,120000株/公顷。#oe1-#oe4指杂交种4个不同株系,wt背景,wt指b73-329于t13的杂交种。
图8为本发明实施例8中异源过表达zmrph1到拟南芥中表达量鉴定及表型照片。其中,a为rt-pcr半定量鉴定结果,b为6周苗龄的拟南芥植株。col-0为对照,oe1-5和oe2-7为两个过表达株系。
具体实施方式
以下实施例用于说明本发明,但不用来限制本发明的范围。若未特别指明,实施例均按照常规实验条件,如sambrook等分子克隆实验手册(sambrookj&russelldw,molecularcloning:alaboratorymanual,2001),或按照制造厂商说明书建议的条件。
实施例1zmrph1表达载体的构建及遗传转化
选取玉米自交系b73-329v3时期幼苗提取总rna,采用takala公司的cdnasynthesiskit反转录合成cdna,使用特异性引物进行扩增获得zmrph1的cds序列,全长1650bp(seqidno:1)。所述引物对的核苷酸序列为:
上游引物:zmrph1-f:5’-atggtggctgccgcggcg-3’
下游引物:zmrph1-r:5’-ctacaattgtgtcgggctctgcttt-3’
回收pcr产物,将目的片段连接到pbcxun载体上,转化大肠杆菌感受态细胞,得到测序正确的重组载体pbcxun-zmrph1。
表达盒是由zmubiquitin1启动子和来自胭脂碱合成酶(nos)基因的3’转录终止区域构成,选择标记基因为草丁膦基因(phosphinothricin)。其表达框如图3所示。
将测序正确的pbcxun-zmrph1重组质粒采用电击法转化农杆菌。菌落pcr鉴定获得阳性单克隆菌株。
用携带目的基因的pbcxun-zmrph1农杆菌菌株转化玉米自交系b73-329幼胚,筛选获得1-2拷贝的阳性苗。经过筛选鉴定获得单拷贝纯合株系。
实施例2杂交种的制备
以自交系b73-329背景的材料(纯合b73-329背景的过表达zmrph1基因的转基因株系)作为母本,以玉米自交系t13作为父本,进行杂交育种获得过表达zmrph1的杂交种材料。
实施例3zmrph1的组织表达模式分析
分别提取玉米自交系b73-329植株4个不同生长阶段(v9,v11,v13,v15)多个组织器官(根,节,节间,叶片,雌穗,花丝,雄穗)的总rna,反转录为cdna。利用zmrph1基因特异性引物(上游引物:zmrph1qpcr-f:5’-caaagttgcaacgcaagaac-3’
下游引物:zmrph1qpcr-r:5’-ggctctgcttttggttacgtt-3’)进行实时荧光定量pcr扩增,以zmactin1基因作为内参基因,其特异性引物为:上游引物:zmactin1qpcr-f:5’-ttgtcctcagtgggggatct-3’下游引物:zmactin1qpcr-r:5’-ctttcaggtggcgcaatcac-3’。结果如图1所示,显示zmrph1在处于快速伸长阶段的节间组织高表达。
实施例4zmrph1亚细胞定位分析
分别将gfp,gfp-zmrph1,zmrph1-gfp连接到pbcxun载体上,由玉米zmubiquitin1基因启动子启动转录,由nos终止子终止转录。将zmα-tubulin1基因连接到mcherry-p1205载体上作为微管的荧光分子探针,由35s启动子启动转录,nos终止子终止转录。
测序正确的质粒进行大量扩增,提取高浓度质粒,调成终浓度为2μg/μl备用。备用载体包括以下载体:gfp-pbcxun,gfp-zmrph1-pbcxun,zmrph1-gfp-pbcxun,mcherry-p1205和mcherry-zmα-tubulin1-p1205。
玉米原生质体的制备与质粒转化。本实验中离心时参数如无特殊说明均为110g,室温或16℃离心5分钟,具体操作步骤如下:玉米材料培养:玉米种子吸水过夜,28℃长日照条件下(16h/8h,l/d)培养3天,避光培养至第二片叶子长至第一片叶子上10-15cm,避光培养约需7-10天;配制15ml酶解液(1.5%纤维素酶r-10,0.3%,0.6m甘露醇,10mmmes(ph5.7),1mmcacl2,0.1%bsa。配制15ml酶解液时,量取11.25ml0.8m甘露醇,0.75ml0.2mmes,2.985ml超纯水,称取0.225g纤维素酶r-10,0.045g离析酶r-10,混匀后50-55℃水浴加热10分钟加速酶溶解并失活蛋白酶,冷却至室温后加入15μl1mcacl2,0.015gbsa,混匀后用0.45μm的滤膜过滤备用,现用现配)和100ml0.6m甘露醇平衡液等;取第二片叶子(第三片叶子也可)中间6-8cm,将20片左右的叶片叠在一起,用双面刀片切,越小越好,切下后组织浸在0.6m甘露醇平衡液中,平衡10分钟;用200目筛子过滤后,将切碎的组织投入酶解液中,真空渗透30分钟;40rpm,28℃,避光酶解2-4小时(切勿超过6小时),最后80rpm,振荡5分钟,使原生质体尽量分离,酶解完后溶液变黄;200目筛子过滤,将酶解液滤入50ml离心管中;室温或16℃水平离心5分钟,弃上清(用枪头吸取),慢慢沿管壁加入4mlw5溶液(154mmnacl,125mmcacl2,5mmkcl,2mmmes(ph5.7)。轻轻摇晃,悬浮沉淀,离心,弃上清(用枪头吸取);再次加入4mlw5溶液,轻轻摇晃,充分悬浮沉淀,冰浴30分钟;离心,弃上清(用枪头吸取),补mmg液(0.6m甘露醇,15mmmgcl2,4mmmes(ph5.7))至原生质体数量1×106-2×106/ml;在2mlep中,每200μl原生质体,依次加入20μl质粒和220μl30%peg(0.6mmannitol,100mmcacl2,25%peg4000(w/v):配制10ml时,分别量取100μl1mcacl2,称取1.09g甘露醇,加入6ml超纯水,甘露醇溶解后加入3gpeg4000,溶解后定容至10ml,现用现配),轻轻颠倒混匀,室温反应15分钟。加入2倍反应体系的w5溶液终止反应,离心收集细胞,弃上清,最后加1mlw5溶液;将ep管水平放置,28℃避光培养16-20小时;离心收集细胞,用枪将上清吸出,只留一点液体,轻轻摇晃混匀,吸取10μl用于观察。
如图2a,2b所示,zmrph1-gfp呈现丝状定位,并且这种丝状结构能与微管的荧光探针重叠表现出共定位现象。
实施例5zmrph1与微管的共沉淀实验
利用体外转录翻译系统表达zmrph1蛋白。实验步骤按照tntquickcoupledtranscriptions/translationsystems(promega)试剂盒说明书中提供步骤进行操作,所表达蛋白被transcendtrna所标记。
微管蛋白的制备。微管蛋白从猪脑中提取纯化,具体实验方法参考朱蕾博士学位论文(拟南芥微管结合蛋白map18对微管动态调控机理的研究,朱蕾,2007)。
从-80℃取出已纯化的微管蛋白和zmrph1蛋白,冰上融化。4℃,200,000g,离心20min,取上清备用。在200μl反应体系中加入60μl微管蛋白和gtp(终浓度1mm),37℃孵育20min以使蛋白聚合,向聚合体系中加入20μltaxol(紫杉醇,2μm),37℃孵育10min;再加20μltaxol(20μm),37℃孵育10min;20μltaxol(200μm),37℃孵育15min。聚合好的微管在25℃,12,000g,离心10min。用pemt缓冲液(80mmpipes,1mmmgcl2,用naoh调ph到6.9,10μmtaxol)重悬沉淀,混匀后测定微管浓度。共沉淀的反应体系为150μl,其中zmrph1蛋白100μl,一份样品中加入50μl聚合好的微管,另外一份样品中加50μlpemt缓冲液作为对照,37℃反应30分钟。并用不与微管结合的luciferase蛋白作为负对照。25℃100,000g,离心15min。离心后的上清和沉淀分别用10%sds-聚丙烯酰胺凝胶电泳,随后进行westernblot免疫印迹实验。显色试剂及方法步骤按照tntquickcoupledtranscriptions/translationsystems(promega)试剂盒的方法操作。如图2c所示,共沉淀实验结果表明zmrph1能与微管共沉淀,而负对照luciferase则不能与微管共沉淀。
实施例6转基因玉米中zmrph1表达量的鉴定
提取实施例1中获得的阳性转基因玉米幼苗中总rna,反转录为cdna,以cdna为模板,用zmrph1基因特异性引物(上游引物:zmrph1qpcr-f:5’-caaagttgcaacgcaagaac-3’下游引物:zmrph1qpcr-r:5’-ggctctgcttttggttacgtt-3’)进行实时荧光定量pcr扩增。结果如图4所示,各过表达转基因株系oe1-oe9中zmrph1基因的表达量都高于对照自交系b73-329。
实施例7田间表型分析
2015~2017年,分别在北京上庄,河北涿州,吉林公主岭,内蒙古包头,宁夏银川,河南安阳,河南开封,共7个实验站进行了田间实验设计。田间实验方案包括三种类型:自交系实验,杂交种实验,高密度实验。自交系实验是指所种植材料为自交系b73-329背景,种植密度为60000株/公顷;杂交种实验是指所种植材料为b73-329与t13杂交种背景,种植密度为90000株/公顷;高密度实验是指实验包括60000,90000,120000株/公顷三种不同密度,材料为杂交种材料。所有实验均以小区为单位,一个小区包括两行,60000株/公顷小区每行21株,90000株/公顷小区每行31/32株,120000株/公顷小区每行42株。一个小区为一个重复,每组实验均设置三个重复。自交系材料田间表型照片如图5所示。
在乳熟期统计倒伏率,茎倒、根倒、茎折均记为倒伏。如图6所示,过表达材料#oe1-#oe4的倒伏率均显著低于对照组。
籽粒成熟后,测定株高、穗位高、节间长、小区总穗粒重、穗重、穗粒重、百粒重、穗长、秃尖长、穗行数、行粒数等多项重要农艺性状(图7)。本发明中仅选取2016年涿州高密度种植试验数据加以说明,如表1所示。数据分析结果显示,过表达材料株高、穗位高、节间长均短于对照,单株产量无显著性差异,其中#oe2小区产量略有增加,有较好应用前景。图中显示数据为:平均值±标准差,t检验:*p<0.05,**p<0.01,***p<0.001。
实施例8拟南芥中过表达zmrph1基因的表型分析
将zmrph1的cds目的片段连接到psuper1300载体上(出发载体为pcambia1300载体,由super启动子驱动表达),转化大肠杆菌感受态细胞,得到测序正确的重组载体psuper-zmrph1。
zmrph1基因表达盒是由super启动子和来自胭脂碱合成酶(nos)基因的3’转录终止区域构成,选择标记基因为潮霉素磷酸转移酶基因(hpt)。
将测序正确的psuper1300-zmrph1重组质粒采用电击法转化农杆菌。菌落pcr鉴定获得阳性单克隆菌株。
用携带目的基因的psuper1300-zmrph1农杆菌菌株侵染拟南芥生态型col-0花絮,筛选获得阳性苗。经过筛选鉴定获得单拷贝纯合株系。
提取7d幼苗的总rna,反转录为cdna。利用zmrph1基因特异性引物(上游引物:zmrph1qpcr-f:5’-caaagttgcaacgcaagaac-3’,下游引物:zmrph1qpcr-r:5’-ggctctgcttttggttacgtt-3’)进行半定量rt-pcr扩增,以atef1α基因作为内参基因,特异性引物为:上游引物:atef1α-f:5’-gcacgctcttcttgctttcac-3’,下游引物:atef1α-r:5’-gggctccttctcaatctccttac-3’。结果如图8a所示,异源过表达转基因株系oe1-5和oe2-7中zmrph1基因的表达量都高于对照col-0。
观察温室中拟南芥植株表型,结果如图8b所示,异源过表达转基因株系oe1-5和oe2-7的植株株高均显著低于对照col-0。
虽然,上文中已经用一般性说明及具体实施方案对本发明作了详尽的描述,但在本发明基础上,可以对之做一些修改或改进,这对本领域技术人员而言是显而易见的。因此,在不偏离本发明精神的基础上所做的这些修改或改进,均属于本发明要求保护的范围。
参考文献
chen,s.,songkumarn,p.,liu,j.,andwang,g.l.(2009).aversatilezerobackgroundt-vectorsystemforgenecloningandfunctionalgenomics.plantphysiology150,1111-1121.
chen,x.,wu,s.,liu,z.,andfriml,j.(2016).environmentalandendogenouscontrolofcorticalmicrotubuleorientation.trendsincellbiology26,409-419.
donald,c.m.(1968).thebreedingofcropideotypes.euphytica17,385-403.
lloyd,c.,andchan,j.(2004).microtubulesandtheshapeofplantstocome.naturereviewsmolecularcellbiology5,13-22.
peng,j.r.,richards,d.e.,hartley,n.m.,murphy,g.p.,devos,k.m.,flintham,j.e.,james,b.,fish,l.j.,worland,a.j.,pelica,f.,etal.(1999).′greenrevolution′genesencodemutantgibberellinresponsemodulators.nature400.
序列表
<110>中国农业大学
<120>zmrph1基因在调控植物株高及抗倒伏能力中的应用
<130>khp181116655.4
<160>2
<170>siposequencelisting1.0
<210>1
<211>1650
<212>dna
<213>zeamays
<400>1
atggtggctgccgcggcggcggccaccgcgccagatccggaccaatcctcgcgcccgccg60
ctcacgccggcgctcgacaagcccaactccgccgccgcgcggaggaactcgcgctccaac120
aagcccgtctcctcgaggtacctgtccgccgccgcatccccggcctcctccacttcgtcc180
tccacgtcgtcgtcgtcttcctcgtcctcctcccgccgctccctctccgcgcagcgcacc240
cggacgtccactccgccgccgcagcactccacgtcgtccacaaccaccgcgtcggcagcg300
gcagcatccgccaccgcgaccacgatgcggagcctctccgtgtcgttccagggggagtcc360
ttcttctacaagacgtcgcgcgcgccgcgggcttcgtcgccgtcctcaccggcagcgcgc420
cgcgggcccacgccggagcgccggaagagcgtctcctccgtgccggaggccgagaacgcg480
cggccgtcgggacgatggccggcggcgaagcccaaggcgtcggacccgctagcgcggagc540
cttgaatgcagcctcgaccgcaaggactccatcctcgccgccgttcacctcctccgccgt600
tctatggccttcgactcctccacgtcactctcgccgtccgaccgggcagctgcagccgct660
ccagacctctccgcgtcctccgacaccgacagtgcctcgtctggtagcaactccggcgcc720
ggggatcccccgcgccgcggcatcagcgtgccggcgagattctggcaggaaaccaacagc780
cgcctgcgccgacttccagagcctggactgccgctaccgtcgtctggccggaggtcgttt840
tcagacagcccaatgtcacccagactgccaggtcggtccccgtctccatgccgtggaagc900
cggggcgctccgagtccatcaagagggcgcagtggggaggcatcgccaaatggacatacg960
gtgcaggctccagcaaatgcgccatctatcatcagttttgcggcagaggtgagaagggcg1020
aaaaagggggagaagagaatcgaggaggcacacagattgcggctgctggacaatcggcat1080
ttgcagtggcgatgcatcaatgctcggacagatgcgacgcttctggtgcagagcttcact1140
gctgagaaaactcttcacagtgcatggaaagaaatatcaagattgcgtgacaatgtcagt1200
aacaaaaggagcagactccagcttcaaaagcaaaaactcaagctgtttgctattcttagg1260
ggacagatgtcatatctagaggcctggtctcatatcgaaaaacatcattcaagtgcttta1320
tcagcagcaatcaaagctctgaaggctagtactcttcgtcttccagttgttgatggcgca1380
aaggctgatgctcaagccgtgaaggaagcagttaactcagcagtggatgtgatgcacaca1440
atgacatcctcgatttgtaatttgctatctaaggtcgagggaacaagctctgtggtgtct1500
gagcttgccaaagttgcaacgcaagaacaaatgttgttggaccaatcaaaggatctcttg1560
tccacggttgcagcaatacatgtcaaaaattgcagcctgcaaactcatatgctacaacgt1620
aaccaaaagcagagcccgacacaattgtag1650
<210>2
<211>549
<212>prt
<213>zeamays
<400>2
metvalalaalaalaalaalaalathralaproaspproaspglnser
151015
serargproproleuthrproalaleuasplysproasnseralaala
202530
alaargargasnserargserasnlysprovalserserargtyrleu
354045
seralaalaalaserproalaserserthrserserserthrserser
505560
serserserserserserserargargserleuseralaglnargthr
65707580
argthrserthrproproproglnhisserthrserserthrthrthr
859095
alaseralaalaalaalaseralathralathrthrmetargserleu
100105110
servalserpheglnglygluserphephetyrlysthrserargala
115120125
proargalaserserproserserproalaalaargargglyprothr
130135140
progluargarglysservalserservalproglualagluasnala
145150155160
argproserglyargtrpproalaalalysprolysalaserasppro
165170175
leualaargserleuglucysserleuasparglysaspserileleu
180185190
alaalavalhisleuleuargargsermetalapheaspserserthr
195200205
serleuserproseraspargalaalaalaalaalaproaspleuser
210215220
alaserseraspthraspseralaserserglyserasnserglyala
225230235240
glyaspproproargargglyileservalproalaargphetrpgln
245250255
gluthrasnserargleuargargleuprogluproglyleuproleu
260265270
proserserglyargargserpheseraspserprometserproarg
275280285
leuproglyargserproserprocysargglyserargglyalapro
290295300
serproserargglyargserglyglualaserproasnglyhisthr
305310315320
valglnalaproalaasnalaproserileileserphealaalaglu
325330335
valargargalalyslysglyglulysargilegluglualahisarg
340345350
leuargleuleuaspasnarghisleuglntrpargcysileasnala
355360365
argthraspalathrleuleuvalglnserphethralaglulysthr
370375380
leuhisseralatrplysgluileserargleuargaspasnvalser
385390395400
asnlysargserargleuglnleuglnlysglnlysleulysleuphe
405410415
alaileleuargglyglnmetsertyrleuglualatrpserhisile
420425430
glulyshishisserseralaleuseralaalailelysalaleulys
435440445
alaserthrleuargleuprovalvalaspglyalalysalaaspala
450455460
glnalavallysglualavalasnseralavalaspvalmethisthr
465470475480
metthrserserilecysasnleuleuserlysvalgluglythrser
485490495
servalvalsergluleualalysvalalathrglngluglnmetleu
500505510
leuaspglnserlysaspleuleuserthrvalalaalailehisval
515520525
lysasncysserleuglnthrhismetleuglnargasnglnlysgln
530535540
serprothrglnleu
545
- 还没有人留言评论。精彩留言会获得点赞!
- 基于温敏阳离子聚合物的siRNA载体及其制备方法与流程
- 一种提高DC疫苗抗肝癌效果的基因靶标、抑制剂和DC肿瘤疫苗的制造方法与工艺
- 一种基因递送系统的制造方法与工艺
- LncRNA ENST00000424523.1及其基因在诊断和治疗药物中的应用的制造方法与工艺
- 核酸扩增反应方法、核酸扩增反应装置以及核酸扩增反应用试剂与流程
- 一种人胎盘绒毛膜间充质干细胞培养体系的制造方法与工艺
- GO‑PLL‑RGDS/VEGF‑siRNA靶向基因药物的制备、活性和应用的制造方法与工艺
- 纤维变性易感性IL22RA2基因及其用途的制造方法与工艺
- G6PD基因及其表达产物在治疗结直肠癌中的应用的制造方法与工艺
- 基于基因治疗的半导体微针组件及其制造模具的制造方法与工艺